









-
Sustainable agricultural productivity depends on the availability of high-quality planting materials. Plant genetic resources provide potential raw materials to breeders for crop improvement to enhance crop productivity and resilience to climate change[1]. Genebanks worldwide conserve the germplasm of a large variety of plant species for current and future use. These collections broaden the available gene pools that will serve different purposes, including breeding and direct use by different users. Also, the genebanks safeguard gene pools of crops whose germplasm is threatened by underutilization or displacement by modern varieties. For instance, traditional African vegetables (TAVs) have been neglected in research and development activities currently[2]. The neglect of TAVs has been threatening their germplasm to disappear due to low utilization despite their significant nutritional and health values. This has led the World Vegetable Center (WorldVeg) to take strategic action in collecting, conserving, distributing, and promoting germplasms of micronutrient-dense TAVs[3,4]. WorldVeg maintains large seed collections in its Eastern and Southern Africa (ESA) genebank based in Arusha, Tanzania with diverse germplasm of TAVs such as African eggplant (AEP), spider plant, amaranths, okra, jute mallow, and African nightshades[3].
Seed storage and in vitro cultures are among the alternative means used to conserve germplasm in genebanks[5]. However, the viability and longevity of stored seeds are affected by storage conditions and storage duration[6]. Cold dry storage is used to store seeds for a long period thereby enabling long-term germplasm conservation at low cost[7]. Nevertheless, seed deterioration tend to accumulate as the storage duration increases, even under cold dry conditions. This is because seed deterioration is actually induced by oxidative reactions of the reactive oxygen species (ROS)[8]. ROS are the results of an incomplete reduction of oxygen in non-enzymatic reactions such as superoxide (O2−•), peroxide (H2O2), hydroxyl radical (•OH), alpha oxygen (α-O) and singlet oxygen (O=O)[9]. Hence, there is a continuous accumulation of reactive oxygen in stored seeds even under cold room conditions[10]. Eventually, this results in accumulation of oxidative damage in stored seeds as the storage duration increases, and or under poor storage conditions, especially with high temperatures and seed moisture content.
As seed age increases, the extensive antioxidant systems such as catalase activities (CAT) and peroxidase activities (POD) are reduced, and seeds are exposed to oxidative reactions[11−13]. Oxidative reactions in seeds, damage stored soluble macromolecules (lipids, proteins, and starch), cytoplasmic organelles (mitochondria and chloroplasts), and lipid cell membranes[8]. Also, they cause genomic damage by inducing single-strand breaks (SSB), double-strand breaks (DSB), cross-links, alkali labile sites, and incomplete excision of repair sites in seeds[14]. These can cause an increase in mutation rates and loss of alleles[7,15]. Also, these deteriorations lead to attacks on DNA which reduces its recovery speed to complete germination during imbibition[16]. Finally, seed deterioration is reflected by the loss of seed viability and vigour. All these deteriorations are threats to the long-term conservation of plant genetic resources.
The advances in molecular biology have generated various novel techniques for assessing the toxicogenomic effect of various external stresses on the genome[17]. The effects of ROS on the genome have been studied using techniques such as random amplified polymorphic DNA (RADP), comet assay, and microsatellite markers[7,14,16]. Comet assay is used to assess the DNA damage down to the individual cell level[18]. The assay gives detailed information on DNA damage but also the repair ability of the cell to recover from oxidative-induced damage[17]. Hence, the comet assay provides detailed information on seed survival mechanisms during storage which is relevant for germplasm conservation in genebanks.
Various studies have evaluated the genetic integrity of stored seeds, loss of seed viability and vigour during seed banking[15,19]. However, the studies on genetic integrity, seed viability, and vigour in ex-situ conserved TAV seeds including African eggplant are lacking. In conserving genetic resources, genebanks usually distribute the genetic materials to different users[20]. Seed characteristics such as 1000-seed weight can be used to precisely determine the number of seeds to be packed for distribution. They can also be used to predict the quality of seeds packed for distribution to various users[21]. Hence, empirical data on the seed viability of ex-situ conserved seeds are critical in seed stock management. The purpose of this study was therefore to investigate the seed viability, vigour, and genetic integrity of seed lots of African eggplant accessions regenerated and stored under cold dry conditions for various time periods (years). It further provided comprehensive information on whether stored seed lots of African eggplants regenerated in different years conserve the same seed quality attributes under cold dry conditions.
-
The seeds used in this study were stored in a cold room (10 °C) at the World Vegetable Center's Eastern and Southern Africa (ESA) genebank in Arusha, Tanzania. Three regeneration years were chosen for each accession (genotype): 2010, 2017, and 2020 corresponding to 12, 5, and 2-year-old seed lots at the time the experiment was set up. The fresh seed materials were exposed to ageing-induced reactions compared to the older seed materials. The study included four African eggplant accessions (Tengeru White, DB3, UG-AE-16, and RW-AE-7) because they were the only accessions with enough seeds required for the experiment. Tengeru White and DB3 genotypes were improved varieties while RW-AE-7 and UG-AE-16 were landraces. The Tengeru White, RW-AE-7 and DB3 are Solanum aethiopicum L. genotypes, while UG-AE-16 is Solanum anguivi genotype. The temperature (°C) and rainfall (in millimetres, mm) prevailing during growing of maternal plants for seed regeneration were collected at WorldVeg's Arusha weather station. They were recorded using a Vantage PRO2 from Davis Instruments California, USA (Table 1).
Table 1. Maternal growing conditions during seed regeneration.
Year Average monthly
temperature (°C)Average monthly
rainfall (mm)Season 2010 24.70 115.30 Rainy 2017 20.40 11.00 Dry 2020 23.22 326.66 Rainy Experimental design and setup
-
A factorial experiment was used with two factors, genotypes and seed age (storage period in years). Four genotypes were used while there were three levels of seed ages, under natural ageing assay (2, 5, and 12 years).
Measuring 1000-seed weight (TSW) and seed moisture content
-
Eight pure seed fraction samples were drawn at random, and 100 seeds were weighed using an analytical balance to obtain 100-seed weight in grams[22]. The counting of 100 pure seed replicates were used to determine 1000-seed weight (in gram)[22]. The seed moisture content was measured using a moisture meter (Model: HygroLab 3, Rotronic AG, Bassersdorf, Switzerland)[23]. First, the seed equilibrium relative humidity (eRH) was measured at 23 °C using the intact seeds to fill 3.2 ml sample holders (three holders) of the moisture meter. The holders were placed in the measuring chamber of an AW-D10 water activity station used in conjunction with a HygroLab 3 display unit. The unit displayed the measurements of water activity and temperature. The water activity generated the value of eRH as per Eqn 1, which, together with the value of temperature, was used to estimate the seed moisture content with the help of the online Kew Seed Information Database of the Royal Botanical Gardens (RBG) (
https://ser-sid.org/viability/moisture-equilibrium ).$\rm Water\; activity = \dfrac{eRH}{100}$ (1) Germination test and measurement of seed viability, vigour, and seedling growth
-
The initial germination data for the selected seed lots were obtained from the genebank inventory as recorded before the storage. The data were recorded from the germination test from 25 seed samples replicated four times for 14 d under screenhouse conditions in organic media. After the storage, the germination test was first done on top of tissue paper in a Petri dish in a germination chamber at 25 °C and alternate periods of 12 h of light and 12 h of dark for 14 d[24]. As the initial germination percentages were poor and thus a seed dormancy is speculated in the tested seeds an alternative growing media, the peat moss was used with a pre-chilling treatment. The seeds were pre-chilled at 4 °C in the refrigerator for 24 h prior to germination test in order to break dormancy as proposed for African eggplant[25]. The treated seeds were then sown in the organic media (peat moss) in four replicates of 100 seeds and germination test was conducted at an average temperature of 24 °C in a screen house for 14 d[24]. The first germination counts were taken at seven days after sowing to estimate germination energy (GE) as shown in Eqn (2)[26]. Final germination counts were recorded at 14 d after sowing to calculate the germination percentage (GP) as indicated in Eqn (3)[24].
$\rm {Germination\; energy}\,(GE)=\dfrac{\begin{array}{c}\rm Initial\; germination\; count\; at \\ \rm 7\; day \;after \;sowing\end{array}}{Total\; number\; of \;seeds\; sown}\times 100$ (2) $\rm Germination\; percentage\,(GP) =\dfrac{\begin{array}{c}\rm Final\; germination\; count \;at\\ \rm 14\; day\; after\; sowing\end{array}}{Total\; number\; of\; seeds\; sown}\times 100 $ (3) Five normal seedlings from the germination test were randomly selected 14 d after sowing to measure the seedling length (SEL) and seedling dry weight (SDW) in order to determine seed vigour index I (SVI) and seed vigour index II (SVII). Seedling length was measured in centimetre (cm) using a ruler. To measure SDW, cotyledons were removed from the 14 d seedlings, and cotyledon-free seedlings were placed in an oven at 80 °C for 24 h. SDW was measured in grams using an analytical balance[27]. SVI and SVII were obtained by multiplying seedling length and seedling dry weight respectively with the germination percentage following the formulas[27], as shown in Eqns (4) and (5).
$\rm {SVI}={Germination\; percentage }\,\times\, {Seedling\; length}\,\left({cm}\right) $ (4) $\rm {SVII}={Germination\; percentage }\,\times\, {Seedling\; dry\; weight }\,\left({g}\right) $ (5) Evaluation of genetic integrity of stored seed lots
-
The genetic integrity of the selected seed lots of African eggplant was studied by using comet assay. The comet assay was conducted in a molecular laboratory at Nelson Mandela African Institution of Science and Technology in Arusha, Tanzania following previous studies[7,16]. The experiment was done on imbibed and non-imbibed seeds for each genotype, and each seed age. Non-imbibed seeds were used to estimate the extent of the genotoxic effect due to seed ageing[7], while seed imbibition was conducted to measure the DNA repair of seeds in the early stage of germination[16]. Seed imbibition was done by placing 20 seeds on top of germination papers in the Petri dishes. The Petri dishes with seeds were then placed in a germination chamber at 25 °C for 24 h[7].
Then, 20 seeds (each from imbibed and non-imbibed seeds) were crushed to remove the embryo and then placed into plastic tubes with 250 μl cold sodium phosphate buffer solution (pH = 7.5). The suspensions were kept in ice for 1 h and then centrifuged for 3 min at 12,000 rpm. The 60 μl aliquots that resulted from centrifugation were transferred into 1% low melting agarose and immediately applied to a microscopic slide pre-coated with high-melting agarose. Two slides were prepared for each treatment. The slides were allowed to solidify at 4 °C before electrophoresis. Solidified slides were placed into the cold alkaline buffer (300 mM of NaOH and 1 mM of EDTA, pH > 13) within an electrophoresis chamber, whereby electrophoresis was run for 20 min at a field density of V = 1V/cm, and the current strength of ~300 mA. After electrophoresis, the slides were washed in sodium phosphate buffer (pH = 7.5) fixed in 70% alcohol and allowed to dry. Finally, the slides were stained for 30 min by ethidium bromide (20 μg/ml or 80 μl) and examined at 200× magnification using the excitation and cut-off filters (490 and 530 nm respectively). A total of eight comet images per treatment, four from each slide, were analysed by using CASP 1.2.3 beta1 software (
https://sourceforge.net/projects/casp/ ). The software directly provides the intensity of the tail and head of the comet image, where the comet tail intensity in comparison to the head informs the number of DNA breaks in the studied sample[28]. Hence, the percentage value of DNA in the tail, as tail intensity (Fig. 1), was used to estimate the DNA damage (genotoxicity) and DNA repair in non-imbibed and imbibed seeds, respectively. For the genotoxicity or DNA damage (in non-imbibed seeds), the higher tail intensity reflected the higher genotoxic effect on the genome. And the higher tail intensity in imbibed seeds represented lower DNA repair (recovery) resulting from DNA damage.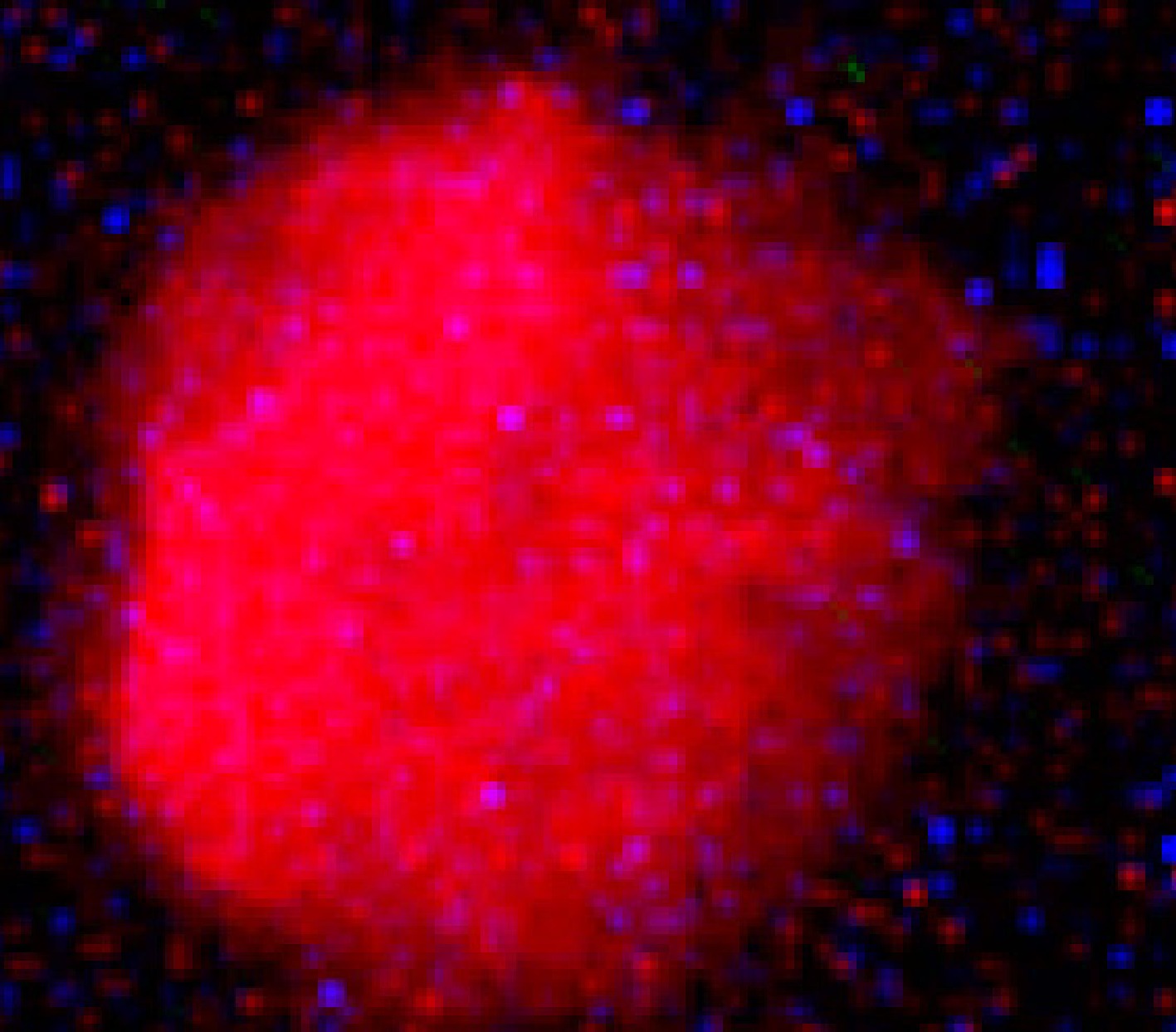
Figure 1.
Comet image of one of nucleoids captured from a microscopic slide (200× magnification).
Data analysis
-
Data were analysed using R-statistical software version 4.0.4[29]. To evaluate the effect of seed ageing on the germination data was analysed using a generalized linear model (glm) with a binomial or quasi-binomial error structure was used to account overdispersion[30]. The variation of germination energy in seed lots of the selected accessions after storage was assessed using with a binomial or quasi-binomial error structure too. To evaluate the effect of growing environmental conditions (temperature and rainfall) on initial seed quality (viability) of selected seed lots, the glm with quasi-binomial error structure was used[30]. The Analysis of Variances (ANOVA) on seed moisture content, 1000-seed weight, SVI, SVII, and the mean tail intensity were done using an Additive Main Effects and Multiplicative Interaction model (AMMI) as per Eqn (6).
${Y}_{jk}= {\text µ} +{G}_{j}+{E}_{k}+{GE}_{jk}+{\varepsilon }_{jk}$ (6) Where,
$ {Y}_{jk} $ $ {G}_{j} $ $ {E}_{k} $ $ {GE}_{jk} $ $ {\varepsilon }_{jk} $ -
Overall, the final viability of each seed lot was lower than the initial viability prior to storage in all the genotypes (Fig. 2). There were significant differences between initial and final mean germination percentages in seeds stored for 12 years (p-value < 0.05). The decrease in seed viability from initial viability was higher in DB3 seeds (−56.8%, the higher negative difference between initial and final seed viability) followed by RW-AE-7 (−28.0%), UG-AE-16 (−23.8%) and Tengeru White (−16.5%) seeds stored for 12 years (Fig. 2). That is the seed viability of DB3 seeds decreased from 68.0% to 11.3% after 12 years of storage (Fig. 2). For the same storage duration, the viability of RW-AE-7, UG-AE-16 and Tengeru White seed decreased from 72.0% to 44.0%, 90.0% to 66.3%, and 75.0% to 58.5%, respectively (Fig. 2). According to the experiments, it appeared that seed viability was significantly reduced in seed 12-year-old seed lots, across all genotypes (p < 0.05). Also, there was significant reduction of seed viability in Tengeru White and UG-AE-16 seed lots after 5 and 2 years of storage respectively (Fig. 2).
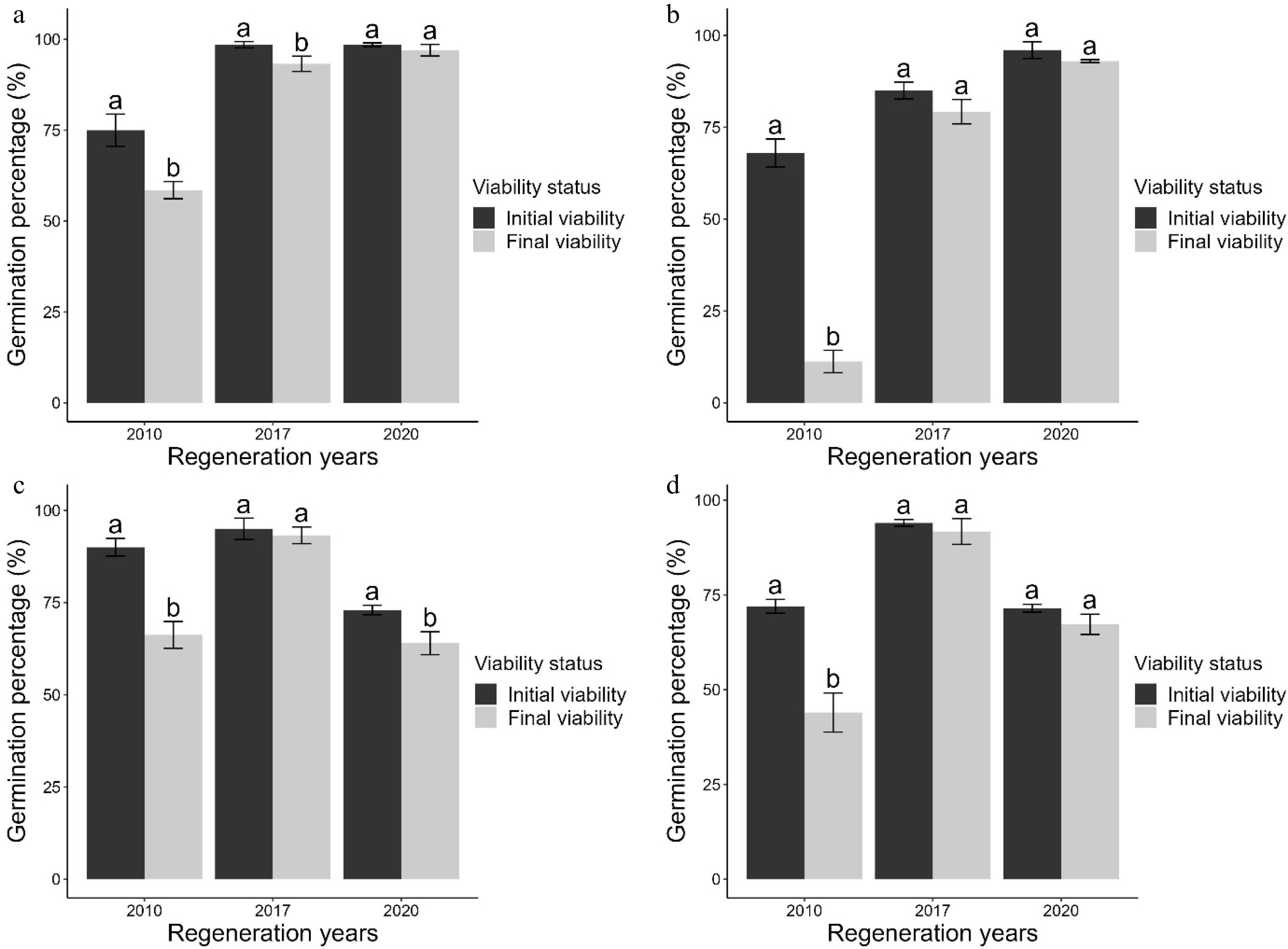
Figure 2.
The initial and final mean germination percentages (mean germination ± standard error) of (a) Tengeru White, (b) DB3, (c) UG-AE-16 and (d) RW-AE-7 seed lots stored for 2, 5 and 12 years at 10 °C.
The temperature prevailing during growing of maternal plants had a significant negative relationship with the initial seed viability (p < 0.001) of the studied seed lots (Fig. 3). However, the rainfall during growing of maternal plants showed no significant relationship with the initial seed viability of the studied seed lots (p = 0.15) (Fig. 3). In addition, there was no significant interaction effect of growing environmental conditions (temperature and rainfall) during the growth of maternal plants on the initial viability of harvested seeds of the selected seed lots (p > 0.05). However, there was a significant interaction effect of the growing temperature and genotypes on the initial viability of harvested seeds of the maternal plants (p = 0.001). Also, the interaction between growing rainfall and genotypes had a significant effect on the initial viability of the selected seed lots (p = 1.19 × 10−10).
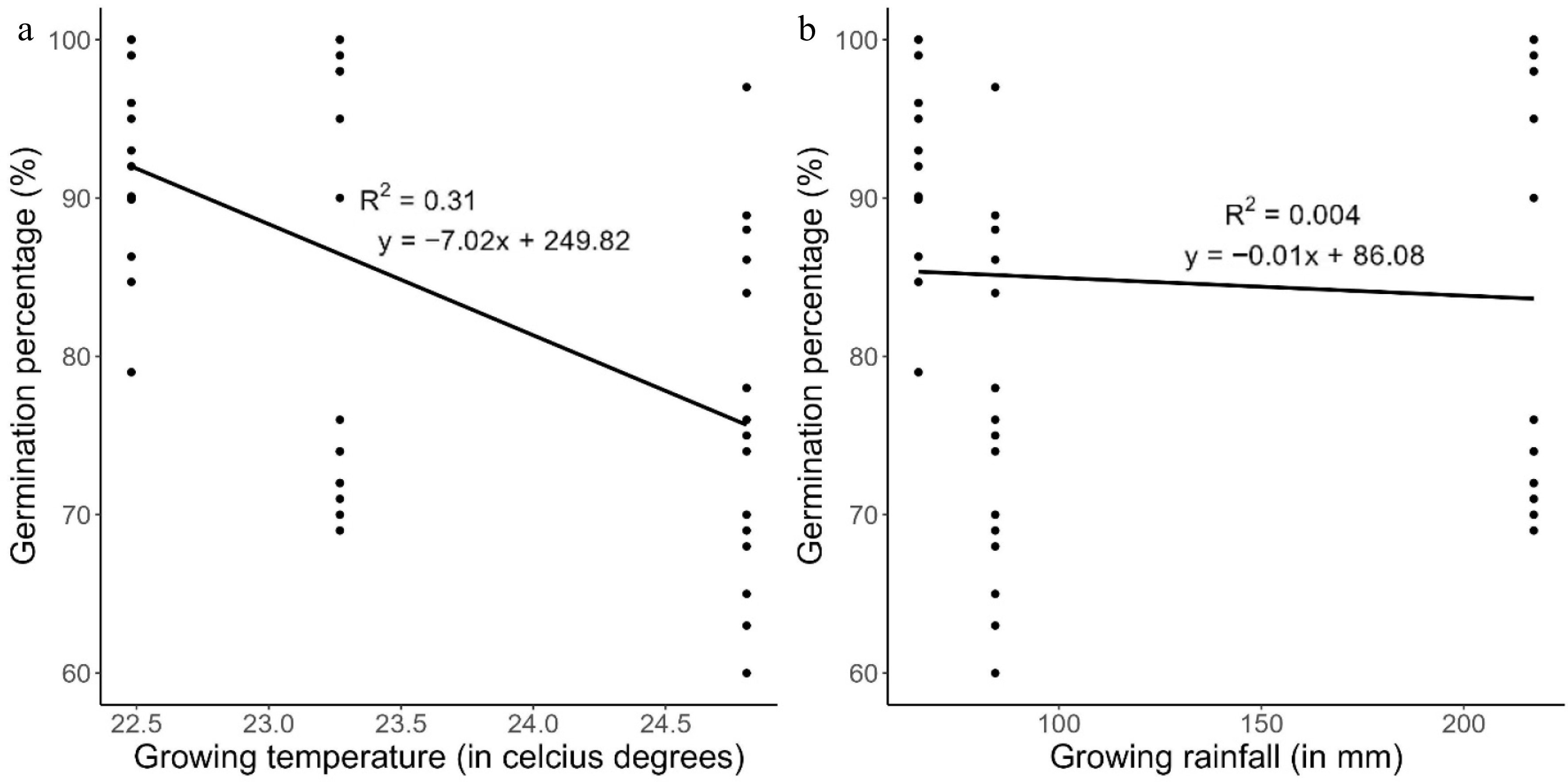
Figure 3.
Germination percentage temperature of African eggplant at 24 °C in screen house for 14 d against maternal plant growing conditions: (a) temperature (°C) and (b) rainfall (mm).
Germination energy, seed vigour, 1000-seed weight (TSW), and seed moisture content of stored seeds
-
Within the studied genotypes there was a significant variation in germination energy of seed lots stored for different years (p < 0.001) (Table 2). However, it is important to note that in UG-AE-16 and RW-AE-7, the initial seed viability of seed lots regenerated in 2020 (which are fresher seeds) was poor (Fig. 2). This poor seed quality observed in 2020 seed lots of UG-AE-16 and RW-AE-7 may have a link with the prevailing environmental conditions during seed production in the field, as can be observed in Fig. 3. Moreover, in DB3 and Tengeru White, the germination energy significantly lowered in the longer stored seeds. The germination energy and germination percentage of stored seeds were significantly and positively correlated (Table 3). There was a significant interaction effect between genotypes and storage period on seed vigour index I (SVI) (p < 0.001) (Table 2). Like on the germination energy, with exception to RW-AE-7 and UG-AE-16, the older seeds (regenerated in 2010) of Tengeru White and DB3 had lower SVI at the time when the experiment was set (Table 2). In fact, the 12-year old DB3 and Tengeru White had extremely lower SVI of 49.5 and 269.1 respectively as compared to other more fresh seed lots of these two accessions (Table 2). The interaction between genotypes and storage periods of seed lots had no significant effect on seed vigour index II (SVII) (p > 0.05) (Table 2). There was a significant positive and high correlation of SVI with both germination percentage and germination energy of stored seed lots (Table 3). There was a significant variation of SVII between the seed lots regenerated in different years in all selected genotypes (p < 0.05), except in Tengeru White (p = 0.148) (Table 2).
Table 2. Seed traits for seed lots regenerated in different years (mean ± standard error).
Parameters Storage period
in yearsAccessions DB3 RW-AE-7 Tengeru White UG-AE-16 Germination energy 2 87.25 ± 1.44aA 25.50 ± 0.6bC 85.25 ± 2.84aB 21.50 ± 5.81bD 5 73.75 ± 3.038bD 75.00 ± 0.91aC 76.50 ± 5.20aB 83.00 ± 3.79aA 12 6.00 ± 1.22cD 13.50 ± 1.94bC 14.25 ± 1.31bB 20.50 ± 3.12bA Seed vigour index I 2 468.72 ± 29.20aA 283.80 ± 20.11abB 419.44 ± 32.12aA 253.44 ± 8.25bB 5 367.72 ± 21.65bB 454.86 ± 28.67aA 406.57 ± 16.31aAB 389.79 ± 12.30aAB 12 49.50 ± 3.73bC 158.40 ± 4.40bB 269.10 ± 12.13bA 274.28 ± 16.17abA Seed vigour index II 2 0.47 ± 0.21abB 0.27 ± 0.04abBC 0.79 ± 0.11aA 0.21 ± 0.05bC 5 0.51 ± 0.15aA 0.39 ± 0.13aA 0.72 ± 0.61aA 0.60 ± 0.20aA 12 0.05 ± 0.01bB 0.19 ± 0.05bAB 0.33 ± 0.18aA 0.29 ± 0.09abA 1000-seed weight (g) 2 2.78 ± 0.04aA 2.69 ± 0.026aB 2.39 ± 0.02aC 2.06 ± 0.22aD 5 2.52 ± 0.02bB 2.54 ± 0.03bA 2.38 ± 0.013bC 2.04 ± 0.03bD 12 2.08 ± 0.029cB 1.94 ± 0.02cC 2.24 ± 0.02cA 1.89 ± 0.03cD Seed moisture content (%) 2 9.03 ± 0.02bB 9.25 ± 0.18bB 9.21 ± 0.17bB 11.60 ± 0.31aA 5 11.20 ± 0.30aB 7.68 ± 0.46aA 10.50 ± 0.05aB 10.70 ± 0.04bB 12 8.80 ± 0.12bB 10.10 ± 0.35bA 8.89 ± 0.67bB 10.30 ± 0.04bA The letters after the mean values represent statistical differences of seed traits between regeneration years down the column (lowercase letters) and between accessions across the row (uppercase letters), at 5% significance level (p < 0.05) (results from One-way ANOVA followed by Tukey test). Table 3. Pearson's correlation (r2) of seed germination, seed vigour, seed metrics and genetic integrity in African eggplant genotypes of stored seed lots.
Parameters GE SVI SVII TSW DNA repair Genotoxic effect GP 0.86*** 0.92*** 0.64*** 0.51*** −0.82*** −0.55*** GE 0.82*** 0.63*** 0.56*** −0.61*** −0.56*** SVI 0.61*** 0.59*** −0.80*** −0.55*** SVII 0.28* −0.49*** −0.33** TSW −0.59*** −0.46*** DNA repair 0.65*** *** Significant at 0.1% level of probability, ** significant at 1% level of probability, and * significant at 5% level of probability. TSW = 1000-seed weight (g), GP = germination percentage (%), GE = germination energy (%), SVI = seed vigour index I, and SVII = seed vigour index II. Furthermore, the 1000-seed weight was significantly lower in seed lots stored for 12 years than seed lots stored for two and five years in all genotypes (p < 0.001) (Table 2). The two-year-old DB3 seeds (regenerated in 2020) recorded a higher average 1000-seed weight (2.78 g) compared with other seed lots in all genotypes. The lowest average 1000-seed weight (1.89 g) was observed in UG-AE-16 seeds that were stored for 12 years (Table 2). The seed moisture content varied significantly among stored seed lots of the selected genotypes (Table 2). However, the recorded variation did not show the pattern loss of seed moisture content in longer stored seed lots as in 1000-seed weight (Table 2). There was a significant weak negative correlation between seed moisture content and 1000-seed weight moisture content (r2 = −0.33, p-value < 0.01). The 1000-seed weight was positively correlated with the germination percentage, germination energy and SVI in the studied genotypes (p < 0.001) (Table 3).
Genetic integrity of stored African eggplant seeds
-
The mean tail intensities (% of tail DNA) obtained from comet images of each nucleoid (Fig. 1) indicating genotoxicity and DNA repair are shown in Table 4. Overall, in non-imbibed seeds significantly higher mean tail intensities were recorded in 12-year-old seed lots compared to fresher 2 and 5-year-old seed lots (p < 0.05), except in UG-AE-16 genotype. The non-imbibed 12-year-old RW-AE-7 seeds had the highest mean tail intensity (41%), while the non-imbibed five-year-old seeds had the lowest mean tail intensity (0.06%) (Table 4).
Table 4. Mean tail DNA intensity (%) (mean ± standard error) of imbibed and non-imbibed seeds of African eggplant genotypes stored under 10˚C and for different durations.
Storage period in years Genotypes DB3 RW-AE-7 Tengeru White UG-AE-16 Non-imbibed seeds 2 0.53 ± 0.512bA 1.54 ± 0.833bA 0.568 ± 0.642bA 3.01 ± 1.77aA 5 0.06 ± 0.045bA 0.33 ± 0.303bA 0.40 ± 0.353bA 1.13 ± 1.08aA 12 8.80 ± 1.59aBC 41.0 ± 5.67aA 18.30 ± 7.76aB 1.69 ± 1.59aC Imbibed seeds 2 0.26 ± 0.161bB 2.67 ± 0.507bB 2.09 ± 0.942aB 16.50 ± 4.05aA 5 8.38 ± 2.03bA 4.69 ± 0.351bA 5.82 ± 5.73aA 6.06 ± 1.36bA 12 28.90 ± 7.15aA 28.60 ± 2.46aA 4.72 ± 2.49aB 4.87 ± 1.40bB The letters after the mean values represent statistical differences of seed traits between regeneration years down the column (lowercase letters) and between genotypes across the row (uppercase letters), at 5% significance level (p < 0.05) (results from One-way ANOVA followed by Tukey test). The same trend is observed in imbibed seeds, where overall older 12-year-old seeds had significantly higher tail intensities (p < 0.05), except in Tengeru White and UG-AE-16. The 12-year-old seeds of DB3, RW-AE-7, and UG-AE-16 had higher mean tail intensity (Table 4). Also, in imbibed seeds, 12-year-old DB3 seeds (regenerated in 2010) had the highest average tail intensity (28.9%), while two-year-old DB3 seeds (regenerated in 2020) had the lowest average tail intensity (0.26%) (Table 4).
The relationship between germination energy and the mean tail intensity of imbibed seeds (accounting for DNA repair) is shown in Fig. 4. Generally, the seed lots with a lower mean tail intensity showed higher germination energy (Fig. 4). However, the 12-year-old seeds of UG-AE-16 and Tengeru White showed a lower mean tail intensity, but their germination energy was poor (Fig. 4). In another case, the two-year-old RW-AE-7 seeds failed to germinate rapidly despite having minimal mean tail intensity (Fig. 4).
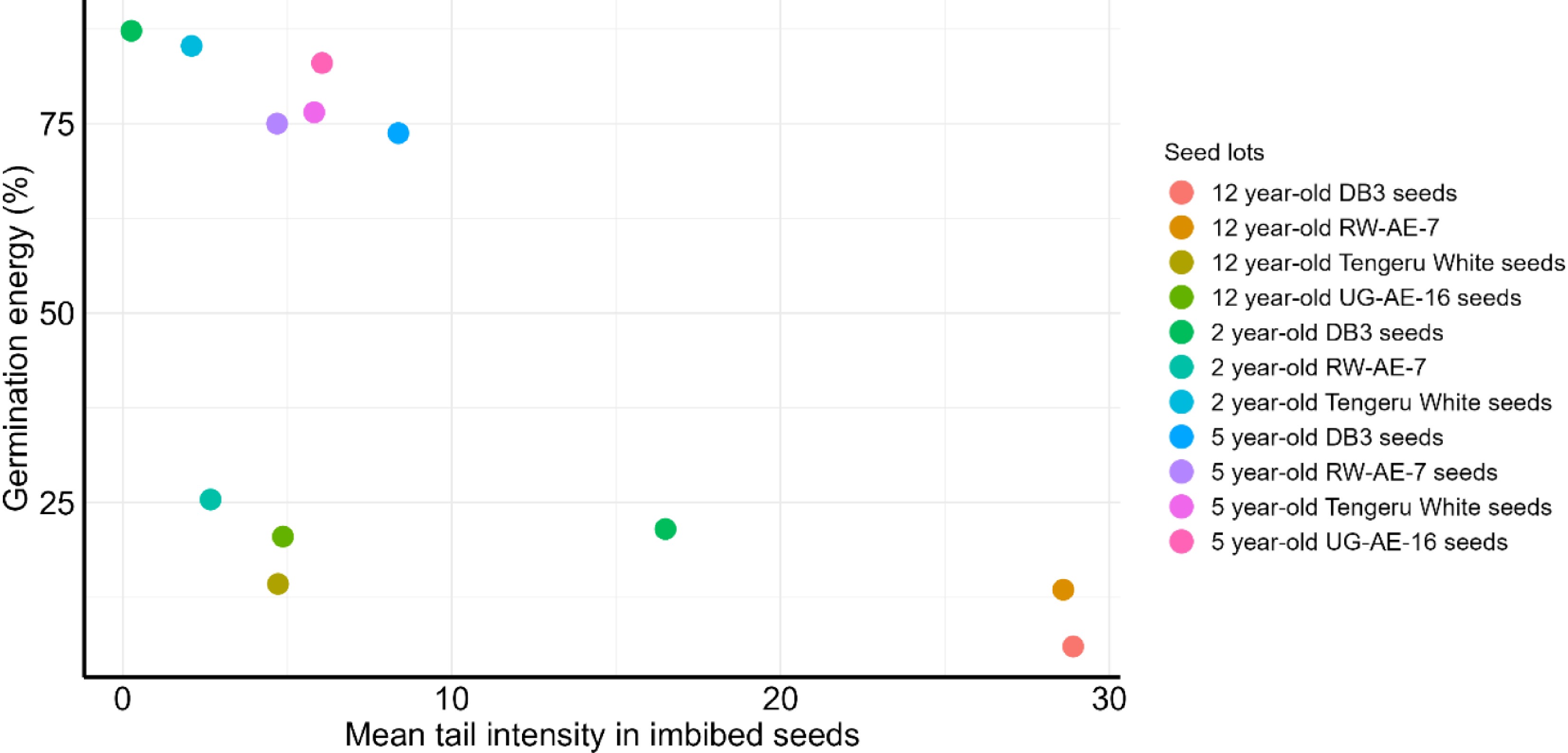
Figure 4.
Relationship of germination energy and mean tail intensity in imbibed seeds of stored genotypes under cold room conditions.
-
Preserving seed quality is the primary goal of genebanks[20]. As the seeds remain viable for a long period of time, the costs incurred by genebanks for regular regeneration of the stored seeds are reduced[7,31]. This study revealed a loss of seed viability in accessions of African eggplant under natural ageing at 10 °C storage in genebank. A decline in seed viability was observed with the storage of all accessions. The older seed lots have been stored for a longer period and might have been exposed to more ageing deteriorative actions than the other fresher seed lots. This corresponded to the fact that the continuous (unceasing) natural accumulation of ageing-induced oxidative damages makes it difficult to prevent seed deterioration even under cold storage conditions[32]. Similar observations have been reported in genotypes of onion and okra[11,33]. However, the initial and final quality of stored seeds differed significantly between genotypes and seed lots. This emphasises the importance of understanding the differences of seed traits between genotypes as this can have implications for the seed longevity prediction, but also the importance of having seeds with high initial viability before storage in genebank.
The rate of seed deterioration is a function of storage conditions (temperature and relative humidity) and internal seed factors (initial seed quality, biochemical composition and genetics)[32]. Our findings showed the significant relationship of initial seed viability on ageing of African eggplant seeds under cold storage conditions. The seeds with higher initial viability seemed to be able to maintain better viability during storage. The seed lots regenerated in 2017 in all accessions were of better quality and were able to maintain viability above 50% after five years of genebank storage. For instance, the Tengeru White, UG-AE-16 and RW-AE-7 seed lots regenerated in 2017 remained with germination percentage of 93.25%, 93.25%, and 91.75% respectively after five years of storage. This demonstrates that having higher initial seed viability could enhance longer storage. Also, our findings indicated that lower initial quality compromised storability and quality of African eggplant seeds. This was more evident in RW-AE-7 and UG-AE-16 seeds regenerated in 2020. These seed lots showed poorer quality only after two years of storage which is partially attributed to their poor initial quality prior to genebank storage. For a long-term storage of germplasm, the genebank curators and seed companies therefore should be aware of factors which may compromise the initial seed quality and take them into account during seed regeneration or seed increase. These factors may include the environmental conditions (temperatures, and rainfall) during the growing of maternal plants, agronomic practices, pests and diseases, harvesting time, and post-harvest handling including proper seed drying before storage[34]. The findings indicated that across all the seed lots and genotypes, there was a significant effect of the prevailing temperature during maternal plants growth on the initial seed viability in studied genotypes. However, the rainfall during seed regeneration had no significant effect on initial seed viability of the studied genotypes. In fact, it has been found elsewhere that the environmental conditions influence the phenological stages of crop, including reproductive stage[35] . The findings of this study corroborate those obtained in Plantago cunninghamii, whereby the temperature during the growth of maternal plants in the field affected seed longevity while field capacity (soil moisture content 2−3 d after rain or irrigation) influenced seed size[36]. Genebanks and seed companies should take this into account to develop high-quality seeds. For instance, appreciating the species (genotypes) specific cardinal environmental conditions to produce high-quality seeds may be useful in planning the critical season for seed regeneration or multiplication[37].
When seed storage duration increases, the oxidative reactions on DNA induce genotoxic effects[38]. The magnitude of genotoxic effect in seed lots of African eggplant regenerated in different years varied considerably between accessions. This basically could be attributed to the inherent storability difference between accessions that can define their ability to tolerate ageing-induced reactions[39]. In this study the genotoxic effect in older seeds of Tengeru White, DB3 and RW-AE-7 after 12 years of storage was accompanied by lower viability and vigour when compared to seed lots stored for 2 and 5 years. Older seeds likely experienced more severe oxidative damage. The deterioration of seed viability during storage is also accompanied by the deterioration of seed vigour and poor seedling growth[31,33,40]. This is what was observed in the older seeds of Tengeru White and DB3 genotypes after 12 years of cold room storage, with a significantly lower germination energy and seed vigour index I as compared with other seed lots. Lowered germination energy and vigour in stored seeds are the consequences of oxidative DNA damage.
Normally during seed germination, the quiescent seeds imbibe water at the initial stage to initiate the chain of events for growth and emergence of the embryonic axis[41]. During germination, seeds initiate DNA replication and metabolic activities that result in cell division and energy production, respectively, for radicle growth and emergence. However, severe oxidative damage on seeds impair the speed of DNA repair, usually causing the seeds to spend much more time in the S-phase (DNA replication) which ultimately delays the completion of germination[42]. The studied seed lots of African eggplant stored under genebank conditions for different years showed significant variation in DNA repair upon imbibition. The significantly lower DNA repair in imbibed DB3 and RW-AE seeds stored for 12 years (reflected by higher tail intensity) was accompanied by reduced germination energy. This indicates that seed quality was impaired during storage. Furthermore, the speedy DNA repair in five-year-old DB3 seeds was reflected by the higher germination energy and germination percentage. The results are in line with findings of previous studies which evaluated the effect of seed ageing on DNA stability in vegetable crops such as pea[9], tomato[14], amaranth[15], and cowpea[43].
Some African eggplant seed lots had the ability to recover from DNA damage. These seed lots showed a reversal response to oxidative damage by displaying a reduced mean tail intensities after imbibition compared to non-imbibed seeds. For instance, after imbibition, the mean tail intensity of two-year-old DB3 seeds regenerated in 2020 was reduced from 0.53% to 0.26%. Also, the mean tail intensity in 12-year-old RW-AE-7 and Tengeru White seeds decreased from 41.00% to 28.60% and 18.30% to 4.72% respectively following imbibition. This form of reversal of seed ageing effect was reported by previous studies[7,44] upon artificial ageing of rice (Oryza sativa L.), pea (Pisum sativum L.) and soybean (Glycine max L.) seed lots. The quick DNA recovery in the seed lots up on imbibition may give insight on possible underlying physiological mechanisms of seed priming in reversing ageing-induced seed weakening. In vegetable crops, different priming treatments have been reported to enhance antioxidant activities in aged seeds[45−48]. In addition, the initial seed quality might also be important in determining the recovery potential of seed lots upon storage. For instance, the poor DNA repair in two-year-old UG-AE-16 seeds would be due to the poor initial quality of seeds.
Implications of the findings for genebanks
-
Seed weight and size indirectly enhance the uniformity of germination and seed vigour under field conditions[49]. In the present study, the germination percentage, germination energy and vigour were significantly and positively correlated with 1000-seed weight of the selected genotypes of AEP. Large seeds were more vigorous, with high germination energy. The 1000-seed weight can therefore be an indicator to monitor seed vigour in AEP genotypes. However, further investigations of seed longevity are required to establish the point at which the stored seed lots should be regenerated. Seed weight can be affected by ageing-induced reactions. For instance, lipid peroxidation, one of the physiological processes involved in seed deterioration[50], modifies membrane lipids into easily volatilized carbonyls with lower molecular weights[51]. These modifications may reduce the number of macromolecules in stored seeds and eventually lower the seed weight. In all the studied genotypes, the 1000-seed weight significantly decreased in older seed lots. These variations in seed weight during storage may also be a warning for genebank staff to monitor seed quality to decide whether regeneration is required to preserve the genetic integrity of the seed. Furthermore, the 1000-seed weight varied significantly between genotypes, as expected given it is controlled by quantitative trait loci (QTLs) on chromosomes[52,53]. This necessitates the use of genotype-specific seed rates but also considers the age of the seed lots for the different genebank activities. This is critical for genebanks because optimal seed rates are required during seed packaging and distribution to seed users. Distributing the required seed amount of each genotype to farmers and other genebank users will reduce seed wastage from stored seed collections.
-
The present study reported on the effect of seed ageing on seed viability, vigour, and genetic integrity in AEP genotypes, under ex-situ genebank conditions. It was shown that natural aging reduced the viability and vigour of stored AEP seeds. However, there was an intra-specific variation of seed traits. This variation should be considered for seed longevity prediction for ex-situ conservation. It was also found that the seed lots with higher initial quality may be able to withstand the ageing induced oxidative damages during storage. Therefore, it is imperative to monitor the growing conditions during seed regeneration to ensure high initial seed quality. Also, in ex-situ conservation the use of molecular tools like comet assay may generate data useful in monitoring seed quality. The comet assay uses few seed samples and gives quick results on DNA breaks which may compromise seed germination and genetic integrity of stored seeds. The comet assay is relatively low cost compared to other molecular tools such as the use of microsatellite and RADP markers.
This research was financially supported by the 'Taiwan-Africa Vegetable Initiative (TAVI)' project and the One CGIAR Initiative on Fruit and Vegetables for Sustainable Healthy Diets (FRESH)’s Work Package 2. Funding for the genebank of traditional African vegetables is provided by strategic long-term donors to the World Vegetable Center: Republic of China (Taiwan), UK aid from the UK government, United States Agency for International Development (USAID), Australian Center for International Agricultural Research (ACIAR), Germany, Thailand, Philippines, Korea, and Japan. The first author is grateful to the World Vegetable Center for the studentship program with Sokoine University of Agriculture and for providing a favourable work environment for this study. We acknowledge the support of laboratory technicians at Nelson Mandela African Institution of Science and Technology for their collaboration in the comet assay experiment. Also, we thank Dr. Fiona Hay for her advice and revision of the initial draft of this paper. The authors appreciate the contributions of anonymous reviewers of this work.
-
The authors confirm contribution to the paper as follows: conceptualization, methodology: Nickas J, N'Danikou S, Shango AJ; formal analysis, investigation, data curation, original draft preparation, visualization: Nickas J; resources, supervision, project administration, funding acquisition: N'Danikou S; article review and editing: N'Danikou S, Kilasi N, Shango AJ. All authors have read and agreed to the published version of the manuscript.
-
The datasets used in the current study are available from the corresponding author, on a reasonable request. The seed samples used in this experiment were acquired by the WorldVeg's genebank following the national and international guidelines and legislations. The code assigned for DB3 is VI059379, Tengeru White is VI059378, UG-AE-16 is VI058824, and RW-AE-7 is VI058808. All these codes are available in WorldVeg’s repository via the following link: https://genebank.worldveg.org/index.html#/?filter=v2z1YE963mbandp=0.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Nickas J, N'Danikou S, Shango AJ, Kilasi N. 2024. Viability and genetic integrity of African eggplant (Solanum spp.) seeds during natural ageing under cold dry storage. Seed Biology 3: e004 doi: 10.48130/seedbio-0024-0003 

Viability and genetic integrity of African eggplant (Solanum spp.) seeds during natural ageing under cold dry storage
- Received: 18 September 2023
- Revised: 07 February 2024
- Accepted: 21 February 2024
- Published online: 25 March 2024
Abstract: Seed deterioration occurs gradually during long-term seed storage. Viability, vigour, and genetic integrity of African eggplant seeds were assessed under natural ageing during genebank storage. The percentage of tail DNA before and after seed imbibition was used to account for genotoxicity and DNA repair during cold dry storage at 10 °C using the comet assay. After 2, 5, and 12 years of storage in the genebank, a reduction of seed viability was observed in all studied African eggplant genotypes. However, seed viability was more significantly reduced in seed lots that had been stored for a longer period in the genebank (p < 0.05). Overall, higher viability loss was recorded in seeds regenerated 12 years before (p < 0.05). The molecular analysis by comet assay showed significant DNA damage during natural aging under genebank conditions, accompanied by important loss of viability and germination energy in older seeds translated into low DNA repair after seed imbibition. The seed lots which had better initial quality were able to maintain higher viability during genebank storage. The findings inform on the storage behaviour of African eggplant genotypes and are relevant for the proper handling of seed lots in genebanks. They are also relevant for future studies on seed longevity prediction in African eggplant.
-
Key words:
- Comet assay /
- DNA repair /
- Genotoxicity /
- Oxidative damage /
- Traditional African Vegetables