









-
Strawberry (Fragaria × ananassa) is a widely grown horticultural plant worldwide due to the unique flavor and it being rich in health-beneficial nutrients. Correspondingly, the flavor compounds and nutrient components usually determine the quality and commercial value of strawberry[1]. Soluble sugars are the vital metabolites influencing strawberry fruit quality, the content of soluble sugars in strawberry fruit is mainly composed of glucose, fructose and sucrose[2]. Meanwhile, citric acid and malic acid are the major organic acids that determine strawberry acidity and represent important organoleptic quality of fruit flavor[3]. In addition, nutrient compounds such as vitamins and γ-aminobutyric acid (GABA) are crucial antioxidants which are key for maintaining people's health and disease prevention[4−6]. Therefore, improving fruit quality not only meets the increasing demand of more pleasant flavor for consumers, but also supplies various nutrients to satisfy different consumer groups.
Sucrose, fructose and glucose determine fruit sweetness in strawberry[7]. Three key enzymes, sucrose synthase (SUS), invertases (INVs) and sucrose phosphate synthase (SPS) have been extensively studied[8]. Structural genes such as FaSPS3, FaSUS1, FaTPK1, FaVPT1 and FaCWINV1, transcription factors FaGAMYB and FaMYB44.2, as well as protein kinase FaMRLK47 have been shown to be involved in the soluble sugar content in strawberry fruit[9−15]. In addition, some sugar transporters were identified and characterized to be involved in sugar transportation in strawberry, for example sucrose transporters (SUTs) and sugar will eventually be exported transporters (SWEETs)[7, 16]. Moreover, citric acid was reported to be the largest in proportion of total acid in strawberry[2], and several structural genes and transcription factors such as FaGAPC2, FaPKc2.2, FaPEPCK, FaMYB5 were found to be involved in citric acid metabolism[17−18]. In addition, vitamin C (ascorbic acid, AsA) is indispensable for human beings due to the mutation of the last enzyme of AsA biosynthetic pathway in humans[19]. The accumulation of AsA is determined by the biosynthesis, transport and recycling pathway. GDP-L-galactose-phosphorylase (GGP) is the key rate-limiting enzyme for AsA level[20,21]. Additionally, the D-galacturonate reductase (FaGalUR) and aldo-keto reductase FaAKR23 play significant roles in AsA biosynthesis of strawberry fruits[22,23]. Furthermore, AsA degradation is critical for its content, after AsA is oxidized to dehydroascorbic acid (DHA), DHA can be degraded to oxalic acid (OA), therefore, the degradation rate of DHA is essential for AsA pool size[24].
Recently, the conjoint profiling of metabolome with transcriptome has been widely applied to analyze metabolites and characterize candidate genes during fruit ripening[25,26]. Nevertheless, the metabolome dynamics and the relative regulatory networks during the ripening of strawberry fruit are largely unknown, especially between cultivars with markedly different quality.
In this study, two octoploid strawberry cultivars ('Sweet Charlie' and 'OiC berry') with significant difference in fruit flavor and AsA level were selected to explore the metabolic and transcriptomic landscape of strawberry fruits. 'Sweet Charlie' shows characteristics such as resistance to anthracnose and high yield, but it is relatively poor in flavor compared to 'OiC berry'. 'OiC berry' possesses excellent flavor and dramatically high content of AsA and soluble sugars, therefore 'OiC berry' is a higher quality fresh table cultivar. In our current work, through metabolome and transcriptome analysis, the aim is to figure out the underlying structural genes and regulatory factors that contribute to the accumulation of soluble sugars, organic acids, and AsA in strawberry fruits. The data will provide further insights into the formation of flavor-related quality which lays a good foundation for enhancement of quality and nutrition in strawberry fruit.
-
The two strawberry cultivars 'OiC berry' and 'Sweet Charlie' were cultivated in the greenhouse with natural light. According to days post-anthesis (DPA), the fruit stages were divided into small green stage (SG), big green stage (BG), white stage (W), turning stage (T) and red stage (R)[9]. The fruits of two cultivars in each of the five stages were frozen and stored in liquid nitrogen before use.
Widely targeted metabolomic analysis
-
Metabolome profiles were tested by Metware Biotechnology Inc. These collected strawberry fruits were freeze-dried and crushed into a powder. After that, 50 mg of the lyophilized fruits was suspended in 1.2 mL 70% methanol and centrifuged at 12,000 rpm for 3 min, then supernatants were filtrated for further UPLC-MS/MS analysis, the detailed protocol can be found in Chen et al.[27]. The parameters of MS were set as follows: ion spray voltage (IS) 5,500 V; electrospray ionization (ESI) temperature 500 °C; ion source gas I (GSI) 50 psi, gas II (GSII) 60 psi, and curtain gas (CUR) 25 psi, respectively; and using the collision-activated dissociation (CAD) with the highest level.
The software Analyst 1.6.3 was used for processing the metabolome result. After removing the redundant signals, the metabolite identification was performed according to the metware database (
www.metware.cn ). The alignment of the chromatographic peak areas from different samples were calculated[28]. The screening parameters for identifying the specific metabolites is as published by Chen et al.[27].Transcriptome analysis
-
Total RNA of fruits was extracted according to the method of Wei et al.[23]. The library preparation and sequencing were conducted by Biomarker Technologies (Beijing, China) using the platform of Illumina, the clean reads were achieved by removing ploy-N, low-quality reads and adapters. Clean reads were then mapped to the strawberry reference genome (Fragaria × ananassa Camarosa v1.0)[29]. Finally, String Tie (
https://ccb.jhu.edu/software/stringtie/index.shtml ) was used to estimate the transcript levels and calculate the Fragments Per Kilobase of Transcript per Million fragments mapped (FPKM) values. DESeq2 v1.22.1 was used for differential expression analysis[30]. The p-values using the Benjamini and Hochberg's method was conducted to control the false discovery rate (FDR). Adjusted p-value < 0.05 and |log2(fold change)| ≥ 1 were used as thresholds for screening significant differentially expressed genes (DEGs). The DEGs were further analyzed by Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment[31] and Gene Ontology (GO) enrichment analysis[32].Determination of soluble sugars content
-
The contents of soluble sugars were detected by high-performance liquid chromatography (HPLC). In brief, fruits were pulverized in liquid nitrogen, 500 mg of resulting powder was incubated with 4 mL of an 80% ethanol solution for 60 min, at 37 °C. And then, the mixture was centrifuged at 10,00 rpm for 10 min, at 4 °C, and the supernatants were collected and dissolved in 80% ethanol and evaporated at 60 °C, for 4 h. The dried samples were suspended in deionized water (1 mL), and then centrifuged in the conditions as before. The supernatants were then filtered using the membrane with a pore size of 0.22 µm. The HPLC system consisted of a SUGAR SP0810 Column (8.0 mm × 300 mm), a Waters E2695 Separations Module (Waters, USA), and a Waters 2414 Refractive Index Detector (Waters, USA). The column temperature was 30 °C, flow rate 0.5 mL/min, detector temperature was 80 °C, and the injection volume was 10 µL.
Determination of organic acid content
-
HPLC was used for determining the contents of organic acids according to the previous protocol of Zheng et al.[33]. Strawberry fruits were pulverized in liquid nitrogen, 500 mg of the sample was suspended in 6 mL deionized water, centrifuged for 5 min at 10,000 rpm, at 4 °C, and 2 mL supernatant was centrifuged for 10 min at 10,000 rpm, at 4 °C. The supernatant was collected through a 0.22 μm Sep-Pak filter (ANPEL, China) and injected into the Agilent 1100 Series automatic sampler for detection of organic acid content using the Agilent 1260 Infinity HPLC system (Milford, MA, USA). Separation was performed on an Athena C18-WP column (CNW, 4.6 mm × 250 mm, 5 μm) at a column temperature of 40 °C. The mobile phase consisted of a solution of KH2PO4 with a concentration of 0.02 mol/L and pH value set at 2.4. The organic acid concentration was calculated by measuring UV absorbance at a wavelength of 210 nm.
Determination of AsA content
-
The HPLC (Waters 2695) method was employed to detect the total AsA content of strawberry fruits. In Brief, 5 g of fruit powder (frozen) was transferred in solution containing 2 mM EDTA and 2% (w/v) metaphosphoric acid. After chilling on ice (20 min), centrifuging was employed at 13,000 rpm, 20 min, 4 °C. Subsequently, a supernatant volume of 15 µL was injected into the HPLC column. The mobile phase contained 5% methanol (pH 5.8) and 0.1 M sodium acetate/acetic acid. AsA was determined by a C18 column (5 µm 25 cm × 0.46 mm). Finally, total AsA content was analyzed based on a standard curve utilizing peak areas at 254 nm[34].
Combined metabolome and transcriptome analysis
-
For the identification of the genes that potentially regulate fruit quality, WGCNA (v1.66) was conducted to model the co-expression regulatory networks according to the quantitative values of genes and metabolites at different fruit stages[35]. Setting the following parameters: soft threshold power: 7; minModuleSize: 50; cutHeight: 0.25. Finally, visualization the co-expression regulatory networks via Cytoscape (v3.8.1)[36].
-
To generate the comprehensive insight into the metabolome dynamics and the regulatory networks during the strawberry fruit quality formation, we collected strawberry fruits at five different developmental stages (from SG to R) (Fig. 1a) from two strawberry cultivars ('Sweet Charlie' and 'Oic berry'). Then UPLC-MS/MS-based metabolic profiling was carried out on these fruit samples. The 1,153 metabolites were identified in the fruits. In addition, we also carried out transcriptome analysis for the same samples. After removing adapter reads, 188.26 Gb clean reads were obtained (Supplemental Table S1). In total, 63,717 genes were identified expressing in five developmental stages of fruits from the two cultivars. Furthermore, principal component analysis (PCA) was conducted, it was confirmed that those identified metabolites and genes were affiliated to five groups, which clearly corresponded to the specific fruit developmental stages (Fig. 1b, c). The DEMs (Fig. 1d) and DEGs (Fig. 1e) between these two cultivars were analyzed, the results revealed that the most DEMs and DEGs were consistently existed in O-T vs S-T, which suggested that the turning stage should be the critical stage which determines the fruit quality. The heatmap of the cluster analysis showed that the data of metabolome (Fig. 1f) and transcriptome (Fig. 1g) from the biological replicates had good repeatability, in addition, the metabolites and transcripts from the five fruit stages were obviously classified into two distinct groups. Collectively, all these findings indicated that during the different developmental stages, the role of metabolite accumulation in strawberry fruits are well-organized.
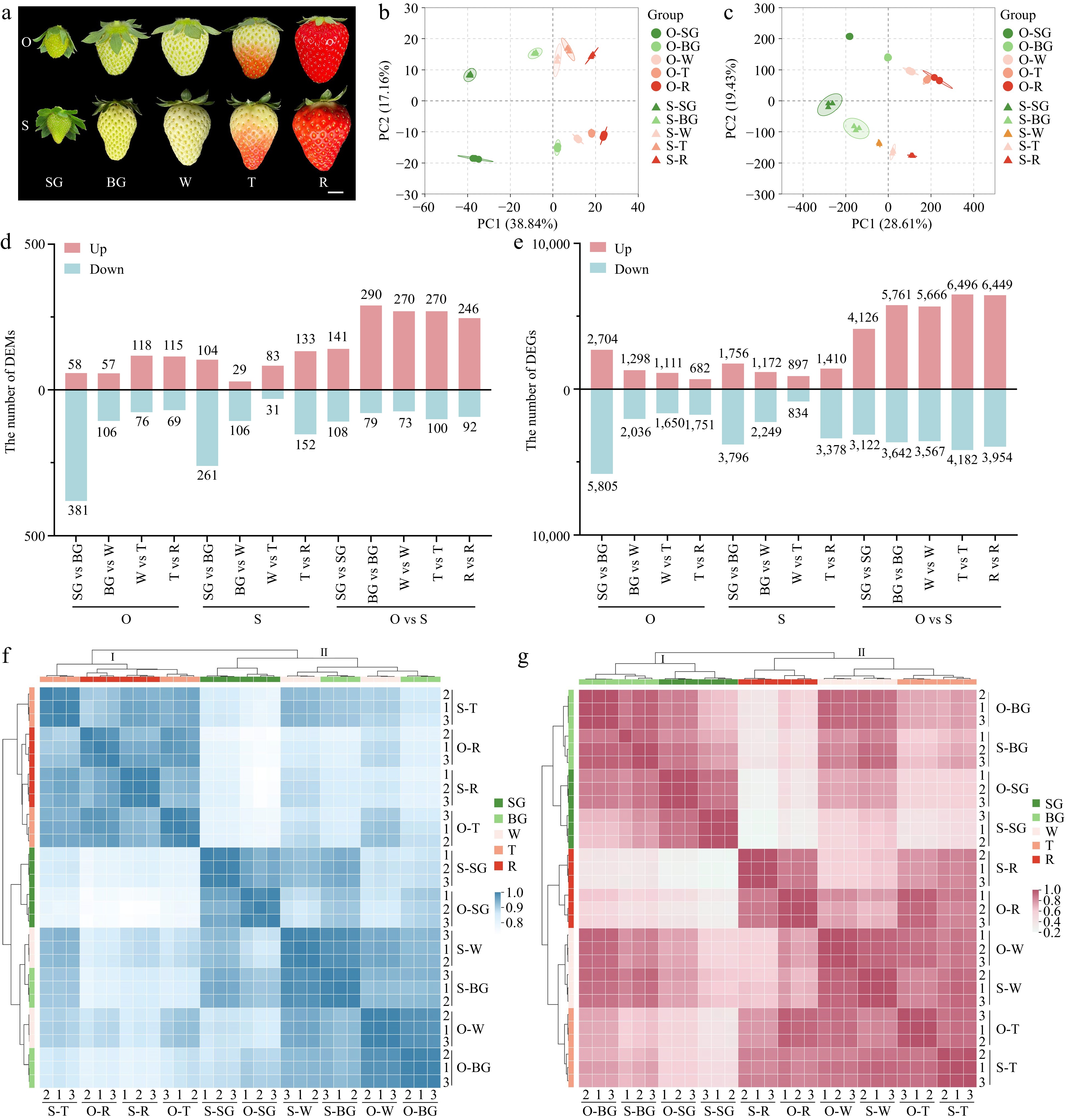
Figure 1.
Overview of metabolome and transcriptome. (a) Five developmental stages of 'OiC berry' and 'Sweet Charlie'. Small green fruit stage (SG), big green fruit stage (BG), white fruit stage (W), turning stage (T), red stage (R), O, 'OiC berry', S, 'Sweet Charlie'. Scale bar: 1 cm. Principal component analysis for (b) metabolome and (c) transcriptome. The number of (d) differentially accumulation metabolites (DAMs) and (e) genes (DEGs). Cluster analysis for (f) metabolome and (g) transcriptome.
Among the 1,153 detected metabolite compounds, including 247 phenolic acids, 213 flavonoids, 117 lipids, 120 amino acids and derivatives, 91 organic acids, 82 alkaloids, 74 saccharides, 59 nucleotides and derivatives, 38 tannins, 37 lignans and coumarins, 37 terpenoids, 21 vitamins and 17 others compounds (Fig. 2a). Soluble sugars, organic acids and vitamins determine the main characteristic flavor of strawberry (Supplemental Table S1). To further clarify the correlation among the corresponding metabolites during fruit ripening, the Pearson correlation coefficients was employed for the 25 representative metabolites. The heatmap in Fig. 2b indicated that there was a significant positive correlation among fructose, glucose, sucrose, maltose, maltotriose, trehalose, melibiose, and raffinose except for galactose and mannose, which had negative correlation with the above eight saccharides. Regarding the nine organic acids, the citric acid content showed positive correlation with that of AsA, vitamin B13, and quininic acid, and the level of malic acid positively correlated with the other organic acids except for succinic acid and ellagic acid. Intriguingly, AsA had the most significantly positive correlation with sucrose, maltose, maltotriose, trehalose, melibiose and raffinose, these implied that there might be an intrinsic association between the accumulation of AsA and saccharides in strawberry fruits. Additionally, AsA exhibited negative correlation with vitamin B2, vitamin B3 and vitamin B5.
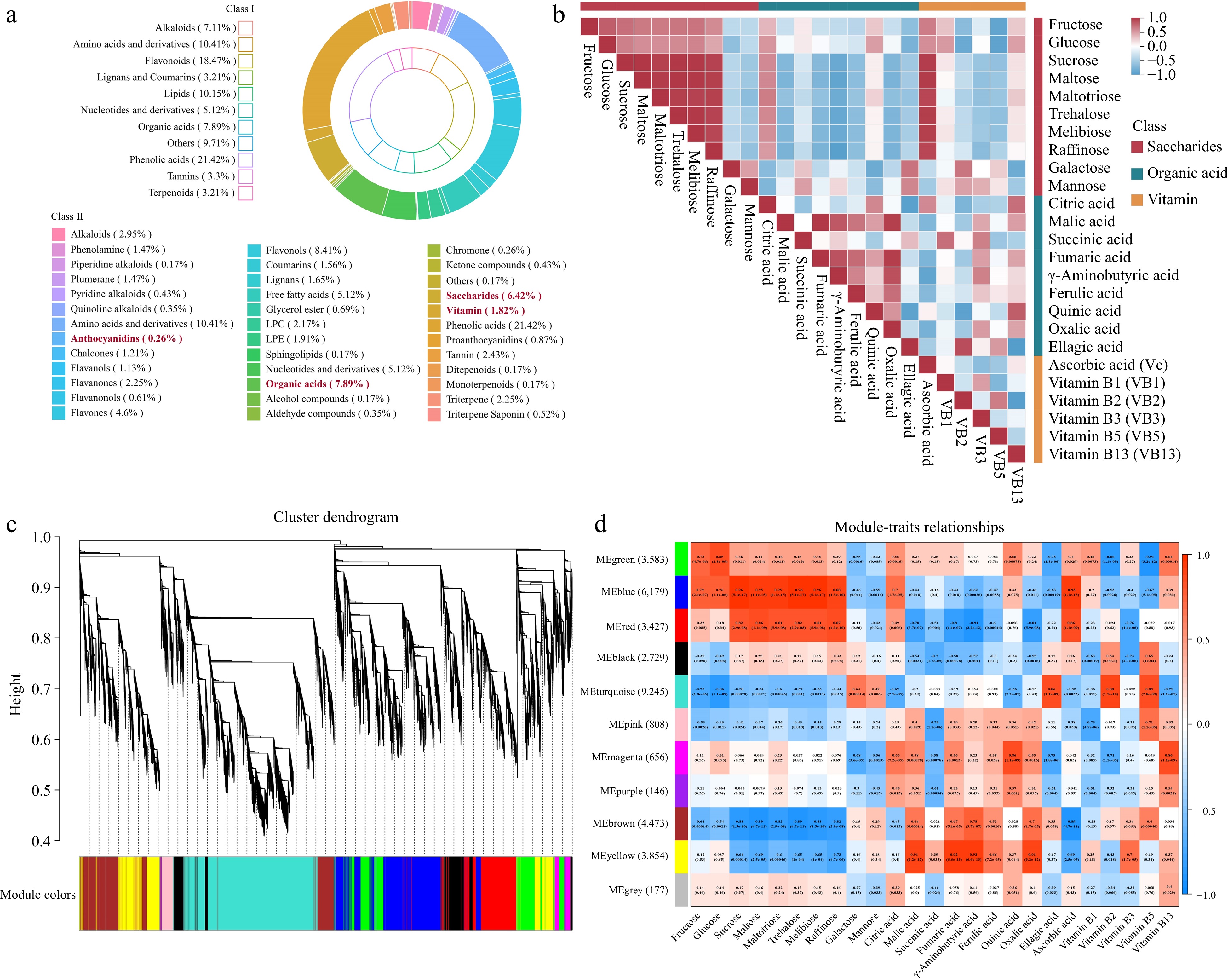
Figure 2.
Correlation analysis of transcriptomic and metabolic during five fruit stages in strawberry. (a) The different categories of metabolites in five developmental stages of 'OiC berry' and 'Sweet Charlie'. (b) Heatmap of correlations between metabolites. (c) Dendrogram indicating co-expression modules analyzed by WGCNA during five fruit stages. (d) Heatmap showing correlations between gene modules and metabolites. Each column in different colors representative a gene module. Every row represents a metabolite. Red color and blue color indicate positive and negative correlation between the gene clusters and the metabolites respectively.
Establishment of 'metabolites vs gene' modules in strawberry fruit
-
To further illustrate the molecular mechanism of the metabolic accumulation involved in the formation of flavor and quality in strawberry fruit, we performed WGCNA to construct the co-expression networks based on DEGs. These DEGs were classified into 11 modules according to the expression patterns (Fig. 2c). Among them, the MEblue module contained the largest number of members (6,179 genes), interestingly, the expression patterns were significantly correlated with content of the flavor-related metabolites included fructose, glucose, sucrose, citric acid and AsA (Fig. 2d). Similarly, the 3,427 genes in MEred module exhibited the significant positive correlation with the mentioned above metabolites. In addition, MEpink (808 genes), MEmagenta (656 genes), MEpurple (146 genes), MEbrown (4,473 genes) and MEyellow (3,854 genes) mudules showed meaningful correlations with most of the organic acids. These modules of genes presented the underlying roles in the regulation of soluble sugars, organic acids and AsA accumulation in strawberry fruits.
Identification of potential regulatory factors in soluble sugar accumulation
-
Sweetness is the most important characteristic contributing to the taste of strawberry and consumer preference. To reveal the difference of soluble sugar accumulation in the two cultivars, the sugars from the five developmental stages were further investigated (Fig. 3c). Among the 12 selected metabolites, melibiose, trehalose, sucrose, maltose, maltotriose, raffinose, fructose and glucose were highly accumulated in 'Oic berry' during fruit maturation, and the accumulation of mannose, galactose, fructose-6P, glucose-6P had no significant differences between the two cultivars. Notably, the content of fructose, glucose and sucrose at the red fruit stage presented highly significant difference between two cultivars (Fig. 3b). Based on the WGCNA data (Fig. 2d), the structural genes regulating soluble sugar accumulation (Fig. 3a) in the MEblue module were examined (Fig. 2d). We identified 43 metabolizing enzymes regulating soluble sugar accumulation including six SWEETs, four SUTs, three SUSs, seven SPSs, six FKs, nine INVs, and eight hexokinases (HK) genes (Fig. 3d), whose transcription levels were positively correlated with the accumulation of three main soluble sugars during fruit ripening. Among these structural genes (Fig. 3d), 23 members of them exhibited significantly higher expression levels in 'Oic berry', based on this, 68 transcription factors were identified by WGCNA analysis, including two WRKYs, 25 MYBs, 12 NACs, six GRASs, seven bZIPs, nine bHLHs, one MADS, one GATA and five AP2s, whose expression were highly positively correlated with the above candidate structural genes and moreover, they formed a compact co-expression network (Fig. 3e). These results indicated that the above transcription factors are potentially involved in soluble sugar accumulation during strawberry fruit ripening.
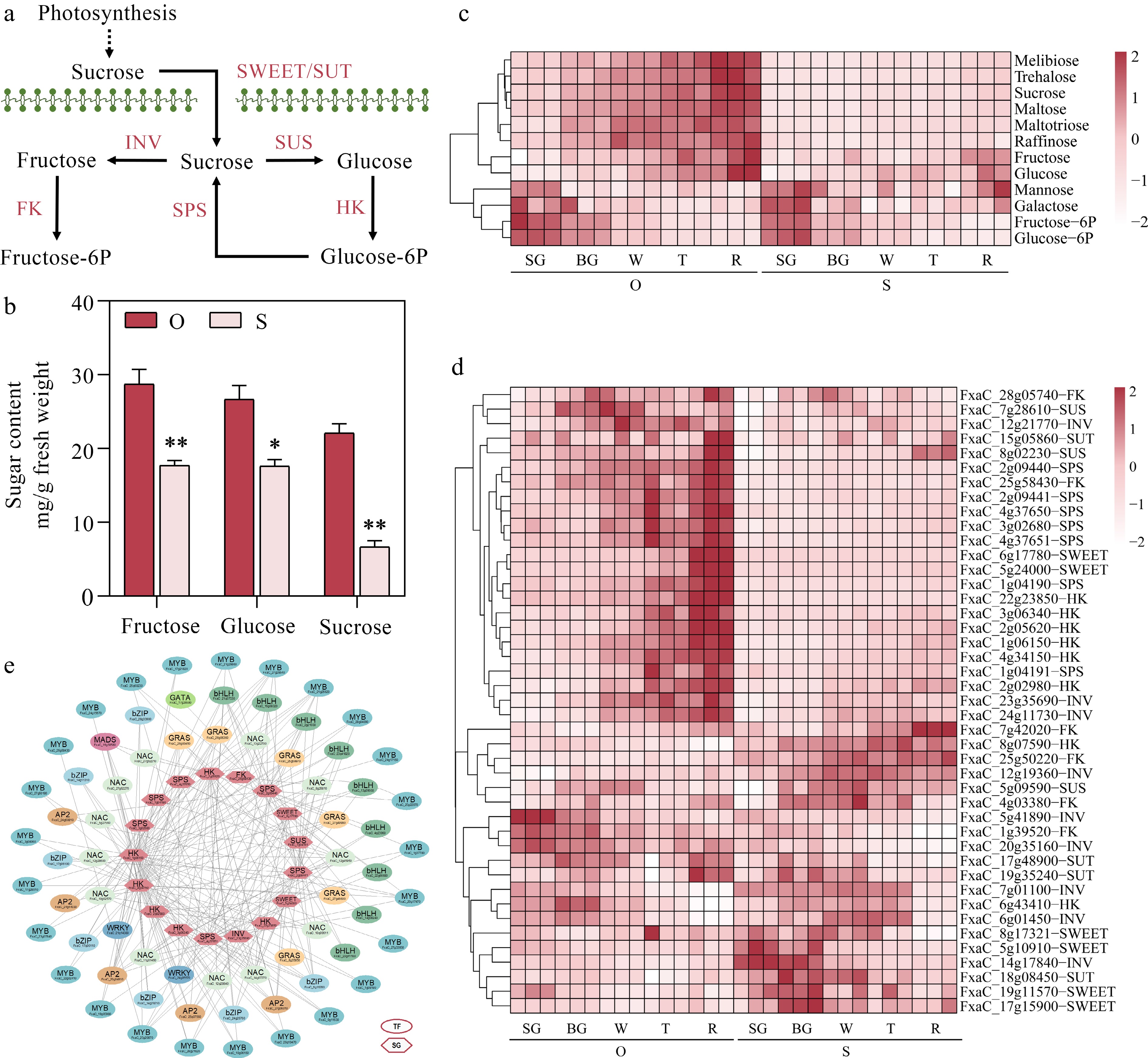
Figure 3.
Identifying potential regulatory factors in strawberry soluble sugar accumulation. (a) Metabolic pathways of soluble sugar. SWEET, Sugars will eventually be exported transporters; SUT, Sucrose transporter; INV, Sucrose invertase; SUS, Sucrose synthase; SPS, Sucrose phosphate synthetases; HK, Hexokinase; FK, Fructokinase. (b) Fructose, glucose and sucrose content during five developmental stages in the two strawberry cultivars. (c) Heatmap showing metabolites in metabolic pathways of soluble sugar. (d) Heatmap showing genes in metabolic pathways of soluble sugar. (e) Soluble sugar metabolic regulatory network. TF, transcription factors; SG, structural genes. Statistical significance was determined by Student's t test (* p < 0.05, ** p < 0.01).
Identification of potential regulatory factors in organic acid accumulation
-
Organic acids such as citric acid and malic acid are critical for the formation of unique strawberry taste. Citric acid is the predominant organic acid in strawberry. To reveal the difference of organic acid accumulation in the two cultivars, the metabolomes from five fruit development stages were investigated (Fig. 4c). Among the nine selected main organic acids, citric acid was highly accumulated in 'Oic berry' during the fruit ripening, and the content of ferulic acid, oxalic acid (OA), γ-aminobutyric acid (GABA), malic acid, succinic acid, fumaric acid, and ellagic acid were significantly accumulated in 'Sweet Charlie', besides, the accumulation of quinic acid showed no significant difference between two cultivars. From Fig. 4b, the data showed that the content of citric acid and malic acid presented significant difference between two cultivars in red stage fruit. To further clarify the potential metabolizing enzymes for citric acid and malic acid biosynthesis, the structural genes involved in citric acid and malic acid metabolism (Fig. 4a) in MEblue and MEyellow modules were analyzed (Fig. 2d). Totally, 41 organic acid metabolizing genes were identified, including 11 pyruvate dehydrogenases (PDH), three citrate synthase (CS), two aconitase (ACO), six isocitrate dehydrogenase (IDH), three 2-oxoglutarate dehydrogenase (OGDC), one succinyl-CoA ligase (SUCL), four succinate dehydrogenase (SDH), two fumarate hydratase (FH) and nine malate dehydrogenase (MDH) genes (Fig. 4d). The transcription levels of these identified genes were positively related to the citric acid and malic acid content. Among these structural genes (Fig. 3d), the 11 members of them presented obviously higher expression level in 'Sweet Charlie', furthermore, 38 transcription factors including four WRKYs, 20 MYBs, one NAC, two GRASs, two bZIPs, three bHLHs, one GATA and five AP2s were identified, whose expression patterns were highly correlated with the 11 structural genes (Fig. 4e). Overall, the results exhibited that these transcription factors might play vital roles in the regulation of the major organic acid accumulation during strawberry fruit ripening.
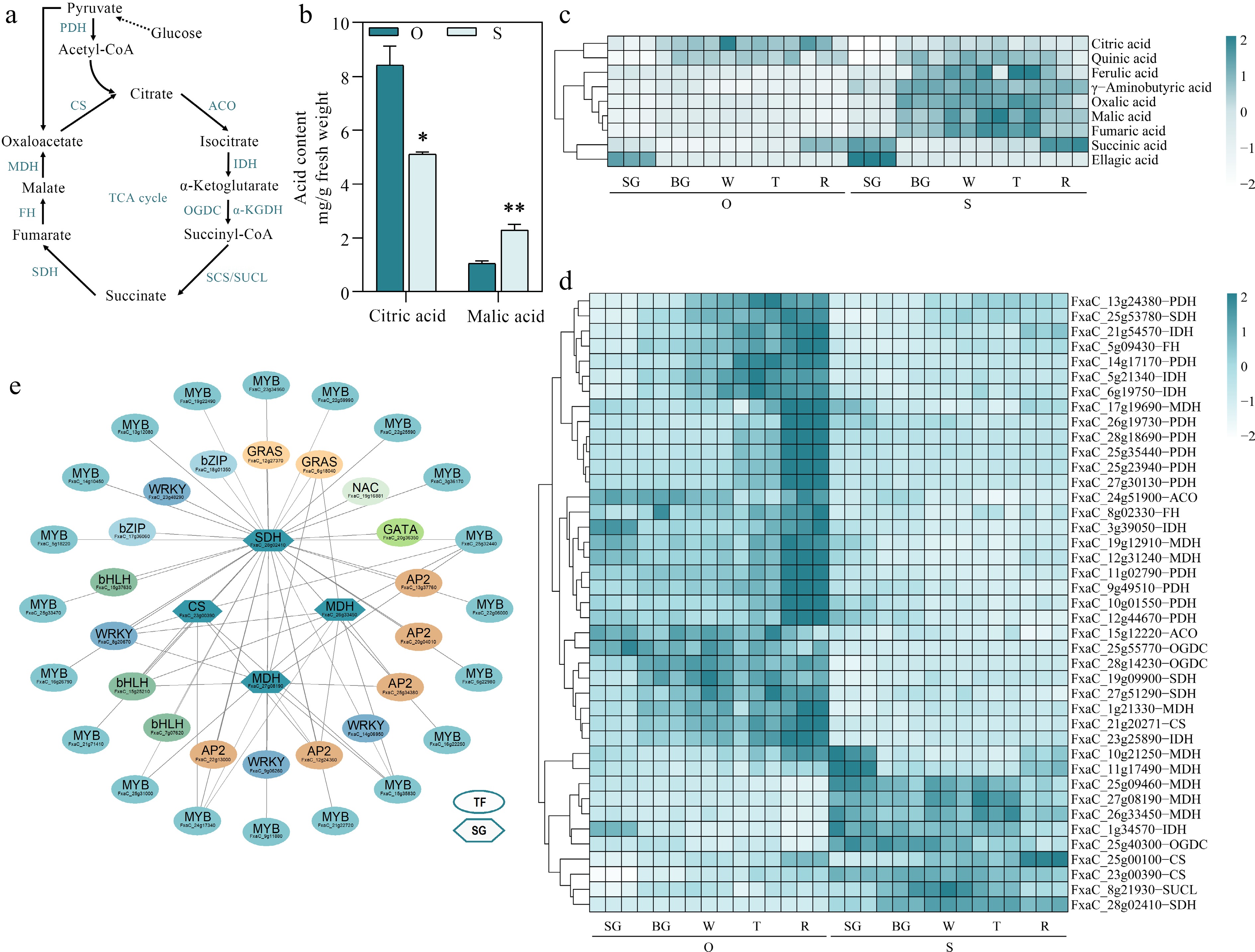
Figure 4.
Identifying potential regulatory factors in strawberry organic acid accumulation. (a) Organic acid metabolism pathways. PDH, Pyruvate dehydrogenase; CS, Citrate synthase; ACO, Aconitase; IDH, Isocitrate dehydrogenase; OGDC, Oxoglutarate dehydrogenase; SUCL, Succinyl-CoA ligase; SDH, Succinate dehydrogenase; FH, fumarate hydratase; MDH, Malate dehydrogenase. (b) Citric acid and malic acid content during five developmental stages in the two strawberry cultivars. (c) Heatmap showing metabolites in organic acid metabolic pathways. (d) Heatmap showing genes in organic acid metabolic pathways. (e) Organic acid metabolic regulatory network. TF, transcription factors; SG, structural genes. Statistical significance was determined by Student's t test (* p < 0.05, ** p < 0.01).
Identification of potential co-regulatory factors for AsA and soluble sugars accumulation
-
Strawberry fruits are rich in vitamins, among them, AsA is the most well-known nutrient component for human being. To reveal the accumulation difference of the vitamin including AsA between the two cultivars, the level of the vitamins from the five fruit developmental stages were investigated (Fig. 5b, c). Among the six delected vitamins, the content of vitamin B2, vitamin B5, vitamin B1 and vitamin B13 presented no significant difference between the two cultivars, and vitamin B3 level was higher in 'Sweet Charlie', notably, the level of AsA from all the five fruit developmental stages was remarkably higher in 'Oic berry' compared to 'Sweet Charlie' (Fig. 5b, c).
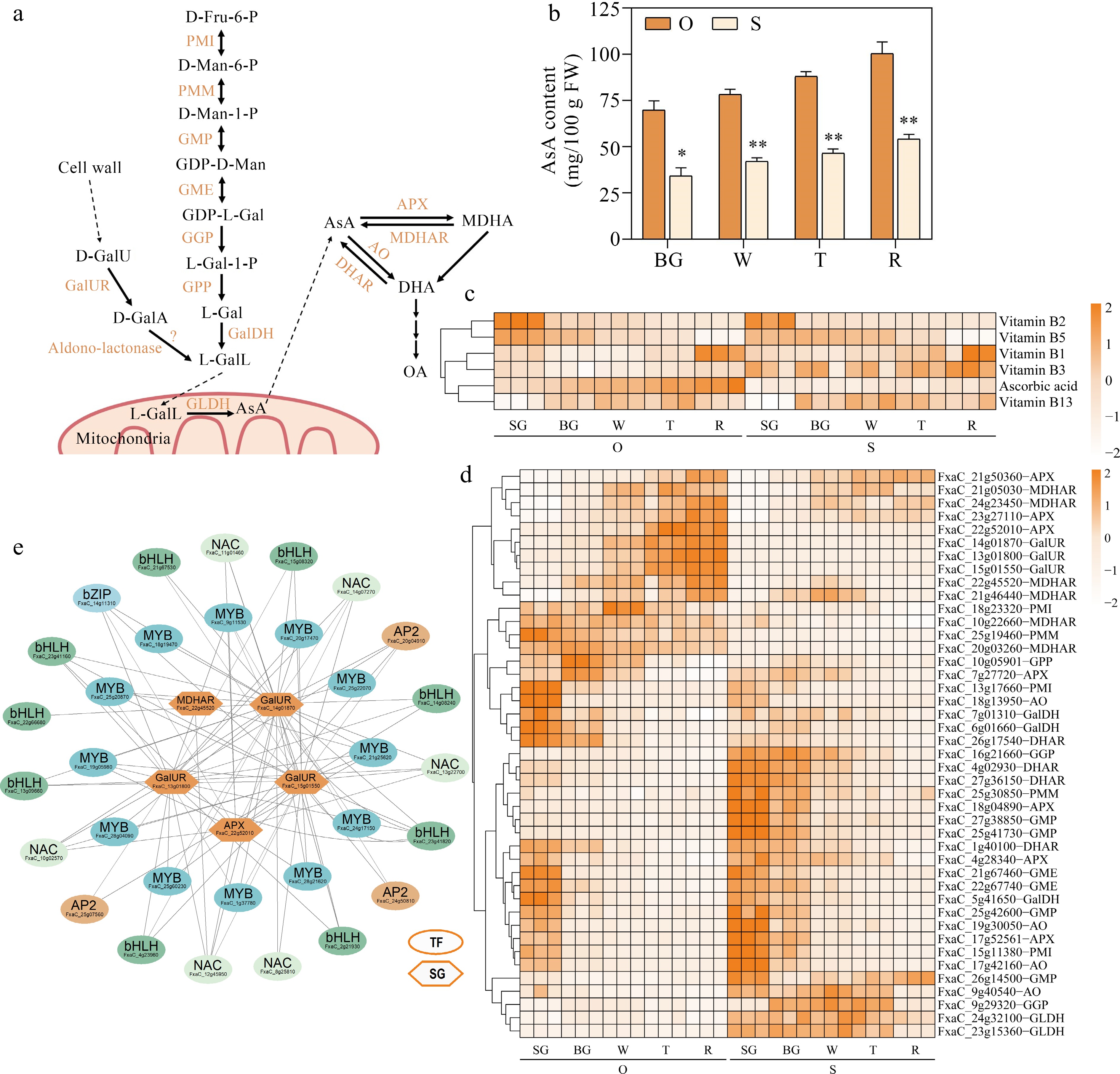
Figure 5.
Identifying potential regulatory factors in strawberry AsA accumulation. (a) AsA metabolism pathways. PMI, Phosphomannose isomerase; PMM, Phosphomannose mutase; GMP, GDP-D-mannose pyrophosphorylase; GME, GDP-D-mannose-3',5'-epimerase; GGP, GDP-L-galactose phosphorylase; GPP, L-galactose-1-phosphate phosphatase; GalDH, L-galactose dehydrogenase; GalUR, D-galacturonate reductase; GLDH, L-galactono-1,4-lactone dehydrogenase; DHAR, dehydroascorbate reductase; AO, L-ascorbate oxidase; MDHAR, monodehydroascorbate reductase; APX, L-ascorbate peroxidase. (b) AsA content during five developmental stages in the two strawberry cultivars. (c) Heatmap showing metabolites in AsA metabolic pathways. (d) Heatmap showing genes in AsA metabolic pathways. (e) AsA metabolic regulatory network. TF, transcription factors; SG, structural genes. Statistical significance was determined by Student's t test (* p < 0.05, ** p < 0.01).
To identify the key metabolizing genes in AsA accumulation, the differently expressed structural genes involved in AsA metabolic pathway were identified (Fig. 5a, d), there were totally 43 AsA metabolizing enzymes including three phosphomannose isomerase (PMI), two phosphomannose mutase (PMM), four GDP-D-mannose pyrophosphorylase (GMP), two GDP-D-mannose-3',5'-epimerase (GME), two GGP, 1 L-galactose-1-phosphate phosphatase (GPP), three L-galactose dehydrogenase (GalDH), three GalUR, two L-galactono-1,4-lactone dehydrogenase (GLDH), four dehydroascorbate reductase (DHAR), four L-ascorbate oxidase (AO), six monodehydroascorbate reductase (MDHAR) and seven L-ascorbate peroxidase (APX) genes (Fig. 5d), whose transcription levels were positively correlated with the AsA content. From Fig. 5d, it was noticeable that the transcription levels of 21 metabolizing genes were higher in 'Oic berry', among them, one third of them were FaMDHARs (Fig. 5d), the above results showed that the recycling efficiency of AsA in 'OiC berry' might be higher than 'Sweet Charlie'. Furthermore, there were 10 members which presented consistent expression patterns with that of AsA accumulation during fruit ripening.
Based on the above metabolizing genes, 31 transcription factors were screened out consisting of 12 MYBs, six NACs, nine bHLHs, three AP2s and one bZIP, whose transcription levels showed positive correlation with the five key AsA metabolic structural genes, including three GalURs, one MDHAR and one APX (Fig. 5e). From the above, the findings implied that these transcription factors putatively participate in AsA accumulation in strawberry fruits.
More interestingly, in our work, the regulation of the metabolism of AsA and soluble sugars highly shared the genes from the MEblue module (Fig. 2d), the transcription factors belonging to MYB, bHLH, NAC, AP2 and bZIP families potentially regulated the accumulation of both soluble sugars and AsA (Figs 3e & 5e, Supplemental Table S3).
-
Conjoint analysis of metabolomic and transcriptomic has become an important technical tool to study the flavor and nutritional quality of fruits[25,26], The global understanding of the accumulation patterns of soluble sugars, organic acids and AsA as well as their underlying transcriptional regulatory mechanisms in strawberry are still limited. In this study, through integrative analysis of the metabolic and transcriptomic data from five fruit developmental stages of two strawberry cultivars ('Sweet Charlie' and 'OiC berry') with significant difference in the quality of flavor and nutrition (Fig. 6), we constructed the strawberry metabolic regulatory networks of soluble sugars, organic acids and AsA, respectively.
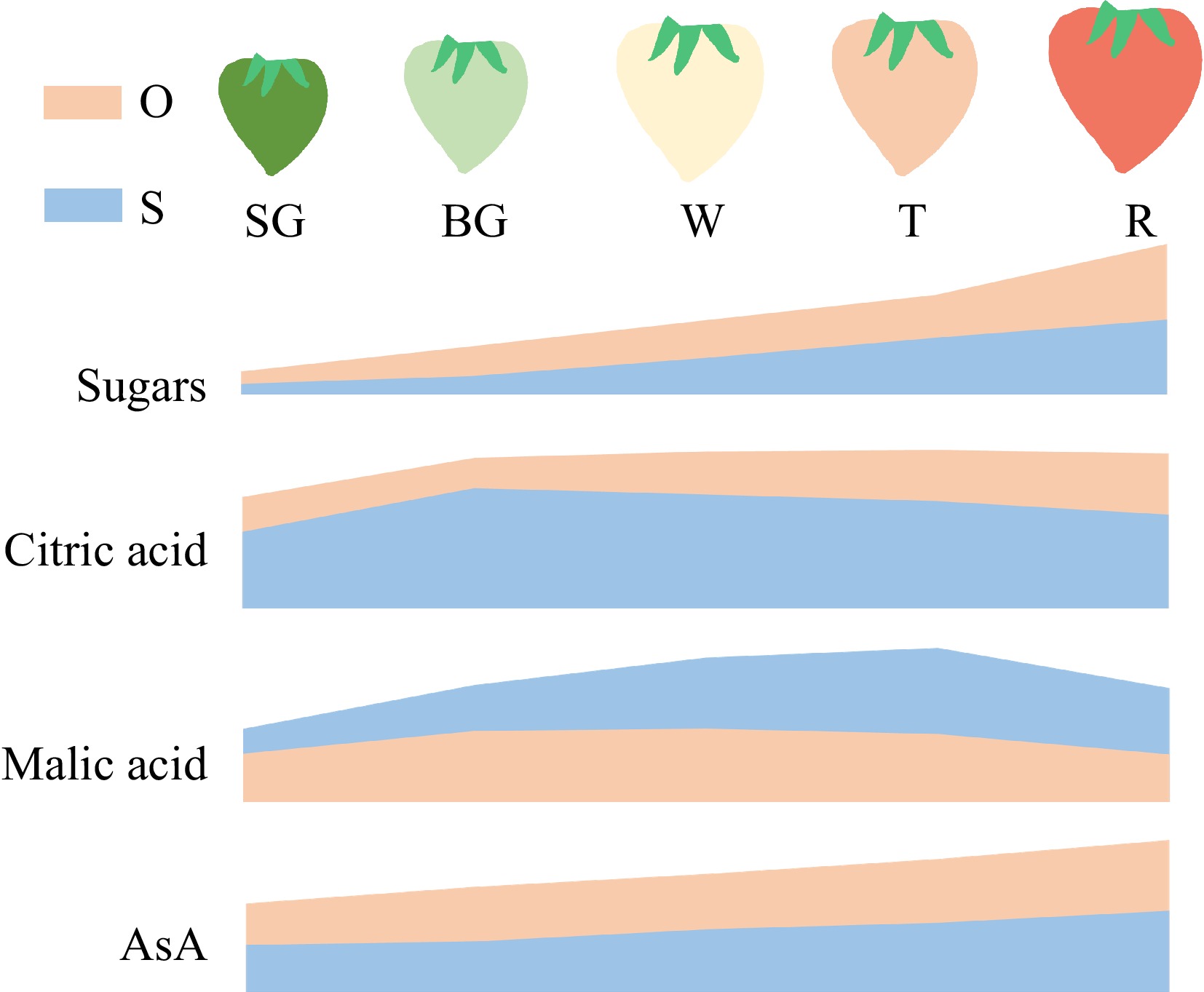
Figure 6.
Schematic diagram indicates the accumulation patterns of major quality metabolites during fruit ripening of two cultivars strawberry. O, 'Oic berry'; S, 'Sweet Charlie'. The content of sugars, citric acid, malic acid and AsA at different fruit stages were standardized to 0−1. The areas of each metabolite from the two cultivars are calculated using the same baseline.
The two strawberry cultivars contain varying levels of flavor compounds due to genetic differences and breeding objectives. Compared to 'Sweet Charlie', the taste of 'OiC berry' is much sweeter, it contains ideal content of soluble sugars, especially more sucrose and fructose (Fig. 3b, c). Although the three soluble sugars present high association with fruit sweetness, sucrose is the only one which contributes the most to consumer preference attributing to its positive association with the total volatiles[37]. To date, multiple structural genes including FaSUT1, FaSPS3, FaSUS1, FaTPK1, FaCWINV, FaVPT1 and FvTST1 have been characterized to modulate sugar accumulation in strawberry fruits[9, 12−14, 38], and some research suggested that transporters SWEETs and SUTs contributed to sugar transportation in fruit[7,16]. Furthermore, in the present study, we identified nine transcription factor families with a total of 68 members which potentially participate in soluble sugar accumulation throughout strawberry fruit ripening (Fig. 3e). Previous studies have demonstrated that transcription factors FaGAMYB[10], FaMYB44.2[15], FaRIF/FvRIF[39−40], FvebZIPs1.1[41] regulate sugar accumulation in strawberry fruit. Furthermore, ClNAC68[42], HpWRKY3[43] and MdbHLH3[44] were reported to be associated with fruit sugar accumulation in watermelon, Pitaya and apple fruits. Overall, our further study is being aimed to investigate whether these identified transcription factors modulate the sugar content during strawberry ripening and explore their transcriptional regulatory mechanisms.
Considering the organic acids, although, from the metabolome result, 'OiC berry' fruits contained a relatively higher level of citric acid than 'Sweet Charlie' (Fig. 4b, c), the flavor of 'OiC berry' is excellent, it indicated that citric acid is not an absolute negative point which brings unpleasant flavor, on the contrary, it should be a contributory metabolite to the pleasant taste. It provides a refreshing tartness and brightness that balance the sweetness of the fruit, enhancing the taste complexity and perceived freshness. From our data, the high level of citric acid is positive to the promotion of comprehensive taste sensation in the cultivar containing high level of sugar. Except citric acid, the content of other organic acids including malic acid, fumaric acid and succinic acid showed higher levels in 'Sweet Charlie' compared with 'OiC berry', which elucidated that the flavor of 'Sweet Charlie' is not ideally satisfactory (Fig. 4b). Herein, 38 transcription factors including the family members of WRKY, MYB, NAC, GRAS, bZIP, bHLH, GATA and AP2 exhibited significant correlation with organic acids levels during strawberry fruit ripening, which were hypothesized to be regulators of organic acids metabolism in strawberry (Fig. 4e). Previously, some studies have demonstrated that the structural genes FaGAPC2, FaPKc2.2, FaPEPCK, and transcription factor FaMYB5 attending citric acid accumulation in strawberry fruits[17,18]. Additionally, recent studies have identified that CitNAC62, CitWRKY1, CitMYB52, CitbHLH2, CitERF13[45−47] participate in citric acid accumulation in citrus fruits. In apple, MdMYB1/10 and MdMYB73 promote malic acid accumulation[48,49], MdMYB44[50] and MdMYB21[51] negatively regulate fruit malic acid content. However, the report of the transcription factors associated with organic acid content in strawberry fruit remains scarce, the molecular mechanism of these candidate genes potentially involved in organic acid regulation is being elucidated in future work.
AsA is an important antioxidant contributing to disease prevention and human health. The recycling pathway also plays a vital role in the accumulation of AsA during fruit development and ripening[26,52]. In our results, 'Sweet Charlie' contain much higher OA in all of the fruit developmental stages than 'OiC berry' (Figs 4c & 5a), which is the downstream metabolite of AsA degradation, furthermore, the expression levels of quite a number of FaMDHARs are obviously lower in 'Sweet Charlie' (Fig. 5d), these results suggested that the recycling efficiency of AsA in 'Sweet Charlie' is much lower than 'OiC berry', leading to the lower AsA content during fruit ripening (Fig. 5b & c), and a recent work reported that the recycling pathway contributes to AsA pool size in plant cells[52]. Except the recycling pathway, the galacturonic acid pathway was firstly reported in strawberry, FaGalUR was identified to be necessary for AsA accumulation in strawberry fruit[53]. From our data, three FaGalUR homologs showed dramatically high expression in 'OiC berry' (Fig. 5d), this illustrates the galacturonic acid pathway is also critical for the AsA accumulation in 'OiC berry' fruits ultimately.
Different from other fruits, such as apple, pear, kiwifruit and jujube, in which, the total AsA content decreases obviously from during fruit ripening[54,55], in strawberry, AsA is accumulated visibly from green fruit to red fruit, and it will come to the highest level when the fruit is fully ripened (Fig. 5b). The accumulation pattern of AsA in strawberry fruit is the same as that of most of the soluble sugars, however, there have been limited studies on this. In our work, the transcription factors belonging to MYB, bHLH, NAC, AP2 and bZIP families showed the potential to regulate the metabolism of both soluble sugars and AsA (Figs 3e & 5e, Supplemental Table S3). Previously, the MYB family transcription factors were reported to directly regulate or indirectly influence the accumulation of soluble sugars and AsA in fruits[15, 25, 56]. In the future, the functional characterization of the possible transcription factors which regulate the accumulation of both of soluble sugars and AsA in strawberry should be conducted.
It is hypothesized that these transcription factors could be valuable genetic resources for the enhancement of flavor-related metabolites contributing to strawberry fruit quality. Further experimental verification is necessary to reveal the biological function of the candidate factors, and the elaborate regulatory network of the transcription factors need to be elucidated, in order to legitimately utilize the gene resources to the molecular breeding for strawberry fruit quality improvement ultimately.
-
In conclusion, we conducted a comprehensive analysis of metabolic and transcriptomic data from five strawberry fruit developmental stages of two cultivars with significant difference in quality of flavor and nutrition, we identified the candidate metabolizing and regulating genes for the accumulation of soluble sugars such as sucrose, fructose, glucose, as well as organic acids including citric acid, malic acid, and AsA. Furthermore, our study provides new insights into the synergistic accumulation of soluble sugars and AsA in strawberry fruits. Our findings not only contribute to deciphering the regulatory networks determining the key flavor and nutrition, but also afford valuable information for molecular breeding which is aimed at thoroughly improving strawberry fruit quality.
-
The authors confirm contributions to the paper as follows: conceptualization, data analysis, data processing and writing original draft: Wei L, Gao Y; data processing: Liu H; metabolites detection and analysis: Ni Y, Xiong R; data curation: Li S, Sun R; strawberry plant cultivation: Wei Y, Zhang H; plant materials: Dong J, Zhong C; formal analysis: Chang L; data investigation: Zhang Y, Sun J; review: Wang G; conceptualization and manuscript modification: Gao Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This work was supported by the Beijing Natural Science Foundation (6244041), China Agriculture Research System of MOF and MARA (CARS-24-A-14), the Beijing Innovation Consortium of Agriculture Research System (BAIC04-2024).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Summary of transcriptome data in strawberry ('OiC berry' and 'Sweet Charlie') at five developmental stages.
- Supplemental Table S2 The major metabolites contributing to fruit quality in strawberry ('OiC berry' and 'Sweet Charlie') at five developmental stages.
- Supplemental Table S3 List of transcription factors regulate both of soluble sugars and AsA.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wei L, Liu H, Ni Y, Xiong R, Li S, et al. 2024. Integrative analysis of metabolome and transcriptome reveals new insights into major quality formation and regulation in two strawberry cultivars with different fruit flavor. Vegetable Research 4: e012 doi: 10.48130/vegres-0024-0011 

Integrative analysis of metabolome and transcriptome reveals new insights into major quality formation and regulation in two strawberry cultivars with different fruit flavor
- Received: 30 January 2024
- Revised: 05 March 2024
- Accepted: 15 March 2024
- Published online: 24 April 2024
Abstract: Soluble sugars, organic acids, and vitamin C (AsA) are the major metabolites contributing to strawberry fruit quality. To date, the metabolomics and genetic information for the above metabolites are still limited. In this study, we integrated the metabolome and transcriptome in two cultivars with remarkably different fruit quality. The significant difference of fructose, sucrose, and malic acid content illustrated the flavor discrepancy in two cultivars, in addition, the recycling pathway was showed to play a vital role in AsA accumulation. Conjoint analysis implied the key metabolizing enzymes related to soluble sugars, organic acids and AsA, respectively, further construction of co-expression regulatory networks identified the transcription factors for the above structural genes. Specifically, it was suggested that the accumulation of soluble sugars and AsA were highly likely to be synchronously regulated by MYB, bHLH, NAC, AP2 and bZIP family members during fruit ripening. This work provides new insights into the accumulation and regulation of core metabolites in strawberry fruits, and the identified gene resources are promising for further improvement of strawberry fruit quality with molecular breeding.
-
Key words:
- Metabolome /
- Transcriptome /
- Strawberry /
- Fruit flavor /
- Co-expression /
- Regulatory factor