









-
Cells are the base unit of the plant multicellular organisms[1]. Cell division is required for growth and development, maturity, organ morphogenesis, and stress response[2−9]. The cell division cycle is a highly regulated and evolutionarily conserved process that is strictly controlled by multiple components, which results in DNA replication and cytoplasmic division to form two daughter cells[10]. Cell division cycle regulation mainly depends on the cell cycle-dependent protein kinase (CDK)/cyclins complex activity. The binding of different CDKs to cyclins regulates the transition from the cell cycle G1/S phase to G2/M phase[11]. However, the enzyme activity of CDKs was suppressed by the interaction between CDKs and Cyclin-dependent kinase inhibitor (CKI)[12]. Based on the similarity and specificity of amino acid sequences, the CKI genes in mammals consist of INK4 and Kip/Cip families. The Kip/Cip family comprises three subfamilies, including p21Cip1, p27Kip1, and p57Kip2[13]. The CKIs mainly consist of p27Kip1 and SIAMESE (SIM) gene family in plants. Due to the similar sequences in the conserved domains between p27Kip1 and Kip/Cip, the p27Kip1 in plants was named as Kip-related-proteins (KRPs)[14]. The evidence indicated that KRPs, an inhibitor of CDKs, could negatively regulate CDKs/cyclins complex activity via binding to CDKs[15].
KRP protein generally carries a conserved cyclin-dependent kinase inhibitor (CDI) domain consisting of about 50 amino acids at the C terminal, required for the binding of KRP/ICKs to CDK-cell cycle phase-specific cyclin complex (CYC)[16]. To date, only a few studies reported that the members of KRP gene family in plant species were identified, such as seven KRPs in Arabidopsis thaliana[17], six KRPs in tomato[18], and nine KRPs in soybean[19]. Accumulating evidence has reported that KRPs played a vital role in the processes of growth and development in plants. In Arabidopsis thaliana, AtKRP2 overexpression promotes trichome branching and endoreduplication[20]. AtKRP6 could regulate the size of rosette leaves[21,22], gametogenesis[23,24], germline development[25], and cytokinesis[26], via inhibiting the cell division of Arabidopsis. ICK3/KRP5 is a positive regulator in Arabidopsis thaliana cell growth and endoreduplication[27]. Although the functions of KRPs involved in plant growth and development were confirmedly demonstrated, few studies showed the functions of KRPs in plant response to abiotic and biotic stresses. The evidence showed that SlKRP2 expression level was significantly up-regulated by cold stress, SlKRP3 expression was markedly up-regulated by the treatment of drought stress in tomatoes, and silencing of SlKRP5 enhanced the susceptibility of tomato to drought stress[18]. In soybean, GmKRP1a, GmKRP2a, and GmKRP4 expression levels were up-regulated by polyethylene glycol stress, and the up-regulation of GmKRP1a, GmKRP2a, GmKRP2b, GmKRP4, and GmKRP5 expression levels treated by salt stress treatment were recorded[19]. However, little is known about the expression of KRPs and their roles in plant response to environmental stressors as of yet.
Eggplant (Solanum melongena) is a popular Solanaceae vegetable cultured all around the world. The growth, development, and yield of eggplant were threatened by multiple environmental stresses, including salt, drought, high temperature, cold, or pathogenic microorganism attack[28,29]. It helps to breed new eggplant varieties with high environmental stress tolerance by exploring the functions of resistance-related genes. Although the knowledge of expression and functions of KRP gene family members in Arabidopsis thaliana was well known[14,17], the information on sequences, expression, and functions of KRPs in eggplant has not been executed. In this study, five KRP gene family members (SmKRPs) in the eggplant HQ-1315 genome were identified, the sequences, structures and expression of SmKRPs in eggplant analyzed under salt and dehydration stress treatments, and the function of SmKRP3 in eggplant response to salt stress explored. The data indicates that SmKRP3 positively functions in eggplant against salt stress via virus-induced gene silencing (VIGS) assay.
-
The method used to identify the members of the KRP gene family in Solanum melongena was as described in previous studies[30,31]. The amino acid sequences of KRP gene family members in Arabidopsis thaliana, which acted as the reference sequence, were obtained from The Arabidopsis Information Resource (
www.arabidopsis.org ) database. The TBtools software (v. 2.027)[32], The National Center for Biotechnology Information (www.ncbi.nlm.nih.gov ) database, and the Eggplant Genome Database (http://eggplant-hq.cn/Eggplant/home/index ; HQ-1315) were used to identify the members of KRP gene family in Solanum melongena. The KRP amino acid sequences from the eggplant inbred lines '67/3' v4.1 (an up-date version of '67/3' v3.0)[33], '67/3' v3.0 (https://solgenomics.net ), as well as GUIQIE-1[34], were obtained to perform the comparative analysis of amino acid sequences of these KRP proteins.Physicochemical properties analysis of KRP proteins in eggplant
-
The physicochemical properties of KRP proteins, including molecular weight, theoretical pI, instability index, and grand average of hydropathicity, were analyzed by searching ProtParam Expasy (
https://web.expasy.org/protparam/ ) website with amino acid sequences of KRP proteins in eggplant[35].Multiple sequence alignment and phylogenetic analysis
-
The amino acid sequences of SmKRPs from Solanum melongena, Solanum lycopersicum, Capsicum annuum, and Arabidopsis thaliana were used to perform multiple sequence alignment and phylogenetic analysis by MEGA 7.0 (v. 7.026) software[18]. The evolutionary tree of KRP genes was generated by MEGA 7.0 software using the neighbor-joining (NJ) method with 1,000 bootstrap replications[36]. The Evolview website (
https://evolgenius.info//evolview-v2/#login ) was used to modify the evolutionary tree.Analysis of chromosomal assignment and collinearity of SmKRPs
-
The distributions of SmKRPs were analyzed using the Gene Location Visualize from the GTF/GFF functional module of TBtools software. Collinearity relationships of SmKRPs were analyzed and visualized by using the Multiple Collinearity Scan toolkit (MCScanX) functional module of TBtools software[37].
Analysis of gene structure, cis-elements, conserved motif, and domains of SmKRPs
-
The gene structures of SmKRPs were visualized using the functional module Visualize Gene Structure (Basic) of TBtools software. The cis-elements within the promoters of SmKRPs were analyzed by searching the PlantCARE website (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ). The conserved motifs of SmKRP proteins were predicted by searching the MEME website (https://meme-suite.org/meme/tools/meme ). The conserved domains of SmKRP proteins were analyzed by searching the SMART website (http://smart.embl-heidelberg.de/ ) with the amino acid sequences of SmKRPs.Plant growth and conditions
-
The seeds of eggplant cultivar ML41, which exhibited middle salt tolerance, were packaged with clean gauze and then placed into the 55 °C water bath at 15 min for pre-germination. The treated seeds were transferred into tap water at room temperature overnight. The seeds were washed and then sowed in the nutrient soil for germination. The eggplant seedlings with two cotyledons were transplanted in a small plastic pot (7 cm × 7 cm) filled with nutrient soil. The seeds of Nicotiana benthamiana were sowed on the wet nutrient soil for germination. The seedlings of Nicotiana benthamiana with two cotyledons and two euphyllas were transplanted in the small plastic pot (7 cm × 7 cm) with nutrient soil. The eggplant and Nicotiana benthamiana seedlings were placed in the illumination incubator with the condition of 25 °C, photoperiod 16 h light/8 h dark, and 60% relative humidity.
Salt and dehydration stresses treatment
-
The plants of eggplant ML41 with 4−6 leaves were gently pulled out from the nutrient soil. The roots were washed with tap water and then cultivated in the Hoagland nutrient solution for 2 days for root restoration. For salt stress treatment, the roots of the eggplant were soaked in 200 mM NaCl solution. The root samples were taken at 0, 2, 6, 12, 24, and 48 h and directly placed in liquid nitrogen. For dehydration stress, the surface of eggplant roots was dried with filter paper and then placed on a table. The root samples were harvested under the condition of liquid nitrogen at the time points of 0, 0.5, 1, 3, 6, and 9 h post-treatment.
Vector construction
-
For subcellular localization assay, the full-length codon sequence (CDS) of SmKRPs were respectively amplified by PCR assay and cloned into the plant overexpression vector pBinGFP2 (containing a GFP protein tag) by using ClonExpress II One Step Cloning Kit (C112-01, Vazyme, China). For the VIGS assay, the specific DNA fragment of SmKRP3 was amplified and cloned into the entry pDONR207 plasmid by BP reaction and then transferred into the pTRV2 vector by LR reaction. The primer pairs sequences used for vector construction in this study are listed in Supplemental Table S1.
Plant total RNA extraction, cDNA synthesis, real-time fluorescent quantitative PCR analysis
-
For total RNA extraction, the roots of eggplant were ground into powder by tissue crusher under liquid nitrogen conditions. The plant cells were broken by lysis buffer TRIpure Reagent (RN0102, Aidlab Biotechnologies Co., Ltd, China). The total RNA was extracted by tri-chloromethane and then sedimented by isopropanol. The concentration of purified total RNA was measured by NanoPhotometer (NP80, Implen, Germany). An agarose gel electrophoresis assay was performed to assess whether total RNA degradation occurs. The cDNA synthesis was carried out following the protocol of HiScript III RT SuperMix for qPCR (+gDNA wiper) (R323-01, Vazyme, China) kit. For the detection of target genes' expression, real-time fluorescent quantitative PCR (RT-qPCR) analysis was performed following the specification of ChamQ Universal SYBR qPCR Master Mix (Q711-02, Vazyme, China). SmActin (Smechr1100649) was regarded as the reference gene to standardize the target genes' expression. Three biological repeats were utilized. The target genes' expression was calculated by using 2−ΔΔCᴛ method[38]. The primer pairs sequences used in this study are listed in Supplemental Table S1.
Agrobacterium tumefaciens cultivation and infiltration as well as subcellular localization analysis of SmKRP proteins
-
Agrobacterium tumefaciens strain GV3101 cells carrying 35S:GFP-SmKRPs or 35S:GFP (acted as a control) constructs grew in the liquid Luria-Bertani (LB) medium with 50 μg/mL kanamycin and 50 μg/mL rifampicin under the condition of 28 °C, 200 rpm overnight. Agrobacterium culture was centrifuged under 28 °C, 8,000 rpm for 10 min, and gathered the bacterium cells. The Agrobacterium cells were resuspended by infiltration buffer (10 mM MES, 10 mM MgCl2, 200 μM acetosyringone, pH = 5.4) to adjust the OD600 to 0.8. Agrobacterium GV3101 cells harboring containing 35S:GFP-SmKRPs or 35S:GFP constructs were incubated in the shaker under the condition of 28 °C, 60 rpm for 1 h. The bacterium was injected into the leaves of Nicotiana benthamiana using a disposable sterile syringe without a needle.
VIGS assay
-
Agrobacterium GV3101 cells containing TRV1, TRV2:SmPDS (encoded a phytoene desaturase, which was used as a marker to prove whether the virus-induced gene silencing assay successful), TRV2:SmKRP3, or TRV2:00 constructs were respectively cultivated in the liquid LB medium. The GV3101 cells were gathered by centrifugation. The OD600 of bacterium solution was adjusted to 0.8 by using an infiltration buffer. The GV3101 cells carrying TRV1 constructs were respectively mixed with the GV3101 cells containing TRV2:SmPDS, TRV2:SmKRP3, or TRV2:00 constructs at a 1:1 ratio. The mixtures were incubated in the shaker at 28 °C, 60 rpm for 1 h, and then injected into the cotyledon of eggplant seedlings with 2−3 leaves. The infiltrated eggplant seedlings were placed in the illumination incubator at 20 °C without light for 48 h. The eggplant seedlings grew in the illumination incubator at 25 °C, photoperiod 16 h light/ 8 h dark for 3 weeks until the leaves of the plants infiltrated with TRV:SmPDS exhibited photobleaching. For survival rate statistics, each biological repeat including SmKRP3-silenced and control contained eight plants, which were treated with 200 mM NaCl solution. After 4 d, the healthy plants and the plants with yellow or wilt leaves of eggplants in each biological repeat were counted to calculate the survival rate.
Measurement of physiological index
-
The physiological indexes, including APX and CAT enzyme activity, were measured as described in previous studies[31,39].
-
According to previous studies, six KRP members were identified in the eggplant '67/3' genome (v3.0)[18,40]. However, only five KRP genes were identified, including SmKRP1-5 in the eggplant HQ-1315 genome. The amino acid sequences of KRP proteins in eggplant '67/3' v4.1, GUIQIE-1, and the HQ-1315 genome were further investigated. The numbers and amino acid sequences of KRP members in eggplant '67/3' v4.1 and v3.0 genome did not change (Supplemental Fig. S1a). In the GUIQIE-1 genome, six KRP members were identified, and five transcripts of EGP26265 and two transcripts of EGP17106 were observed (Supplemental Table S2). It was found that five KRP members in the eggplant '67/3' (v4.1 and v3.0) and GUIQIE-1 genome had corresponding homologs in eggplant HQ-1315 genome. However, the amino acid sequence of KRP protein (SMEL4.1_09g013240.1 and SMEL_002g167200) in eggplant '67/3' (v4.1 and v3.0) and EGP31387.1 in GUIQIE-1 exhibited some difference comparing to its homolog (Smechr0202706) in eggplant HQ-1315 (Supplemental Fig. S1a). In addition, the results of conserved domain prediction showed that SMEL4.1_09g013240.1, SMEL_002g167200, and EGP31387.1 protein respectively contained a conserved CDI domain, while Smechr0202706 protein did not carry a conserved CDI domain (Supplemental Fig. S1b). This may be the reason for the difference in the number of KRP members between in eggplant '67/3', GUIQIE-1, and HQ-1315. In addition, KRP genes were further identified in other Solanaceae plants including six KRP genes in potato, six KRP genes in tobacco, six KRP genes in pepper, and six KRP genes in tomato (Supplemental Table S3). The physicochemical properties of five SmKRPs in eggplant HQ-1315 were further analyzed. As shown in Table 1, SmKRPs encoded 157~222 amino acids with 474~669 base pairs (bp). The molecular weight, theoretical pI, instability index, and grand average of hydropathicity of SmKRP proteins were 18,579.92~24,529.1 Da, 5.11~9.84, 40.16~62.55, and −1.162~ −0.834, respectively (Table 1).
Table 1. Physicochemical properties of the KRP family genes in eggplant.
Gene name Gene ID CDS length
(bp)Protein length
(aa)Molecular weight
(Da)Theoretical pI Instability index Grand average of
hydropathicitySmKRP1 Smechr0902520.1 636 211 23,426.48 9.47 45.52 −0.889 SmKRP2 Smechr0901526.1 483 160 18,779.21 9.84 62.55 −0.834 SmKRP3 Smechr0502598.1 669 222 24,471.22 9.28 50.26 −1.078 SmKRP4 Smechr0100331.1 663 220 24,529.1 5.11 40.16 −0.862 SmKRP5 Smechr0300499.1 474 157 18,579.92 6.98 62.29 −1.162 Phylogenic analysis of KRP genes from eggplant, tomato, pepper, and Arabidopsis thaliana
-
To analyze the phylogenic relationship of SmKRPs with its homologs from the other plant species, we generated the evolutionary tree of SmKRPs with the KRP gene family members from Arabidopsis thaliana, tomato (Solanum lycopersicum), and pepper (Capsicum annuum) using MEGA 7.0 software. We found that the KRP genes from these four plant species were divided into three subgroups, including class I, class II, and class III (Fig. 1). In class I, SmKRP5, and SlKRP5 were grouped into the same branch, implying that the sequence and function of SmKRP5 may be similar to SlKRP5. In class II, there were three SmKRPs, including SmKRP1, SmKRP2, and SmKRP3. In class III, SmKRP4 had a high sequence similar to SlKRP6.
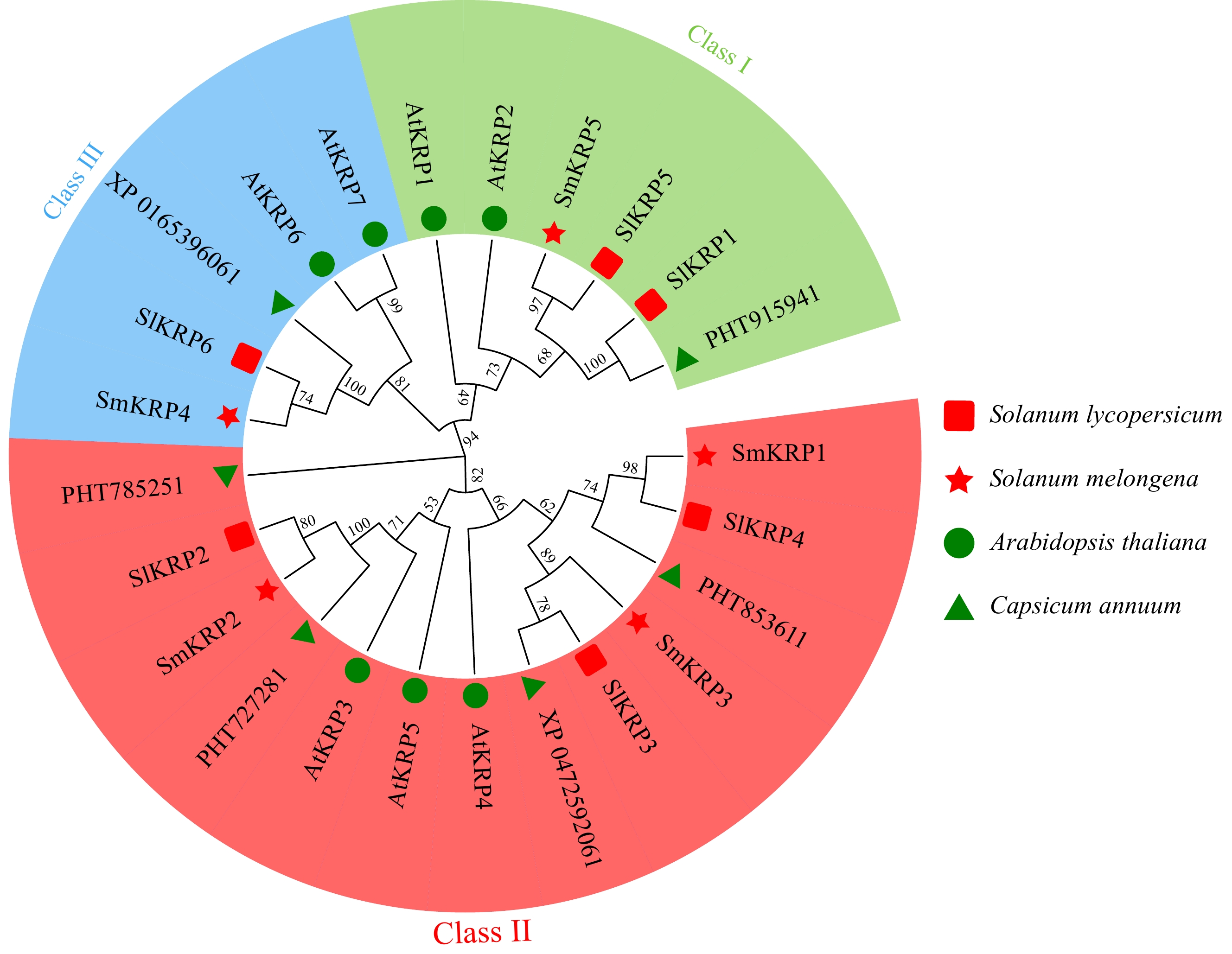
Figure 1.
Phylogenetic relationship analysis of SmKRPs with its homologs from Arabidopsis thaliana, Solanum lycopersicum, and Capsicum annuum by constructing an evolutionary tree. The KRP proteins were distributed into three classes, which is distinguished by different colors.
Analysis of chromosomal location, collinearity relationship, gene structure, conserved domain and motif, and cis-elements of SmKRP members
-
Next, the chromosomal location of SmKRPs were analyzed. The results showed that five SmKRPs were located on four chromosomes including Chr. 1, Chr. 3, Chr. 5, and Chr. 9 (Fig. 2a). Both SmKRP1 and SmKRP2 are located on Chr. 9, while SmKRP3, SmKRP4, and SmKRP5 are located on Chr. 5, Chr. 1, and Chr. 3, respectively. In addition, we also investigated the collinearity relationship among five SmKRPs. We analyzed the homology and sequence similarity of the KRP members between eggplant and tomato. The KRP members SmKRP1~SmKRP5 shared 84%, 64%, 70%, 81%, and 43% sequence similarities with the KRP members SlKRP4, SlKRP2, SlKRP3, SlKRP6, and SlKRP1 in tomato, respectively (Supplemental Table S4). Interestingly, it was found that the locations of SmKRP1 and SmKRP2 on the eggplant Chr. 9 were similar to the locations of their homologs SlKRP4 and SlKRP2 on the tomato Chr. 9 (Fig. 2a), suggesting that these two groups of KRP homologous exhibit similarities in evolution. A collinearity relationship was found between SmKRP1 and SmKRP3 (Fig. 2b), suggesting that a duplicated segment occurred between SmKRP1 and SmKRP3. Moreover, the collinearity relationship of KRP members in eggplant, Arabidopsis thaliana, and tomato were also analyzed. However, the collinearity relationship among these KRP members was not observed (Supplemental Fig. S2).
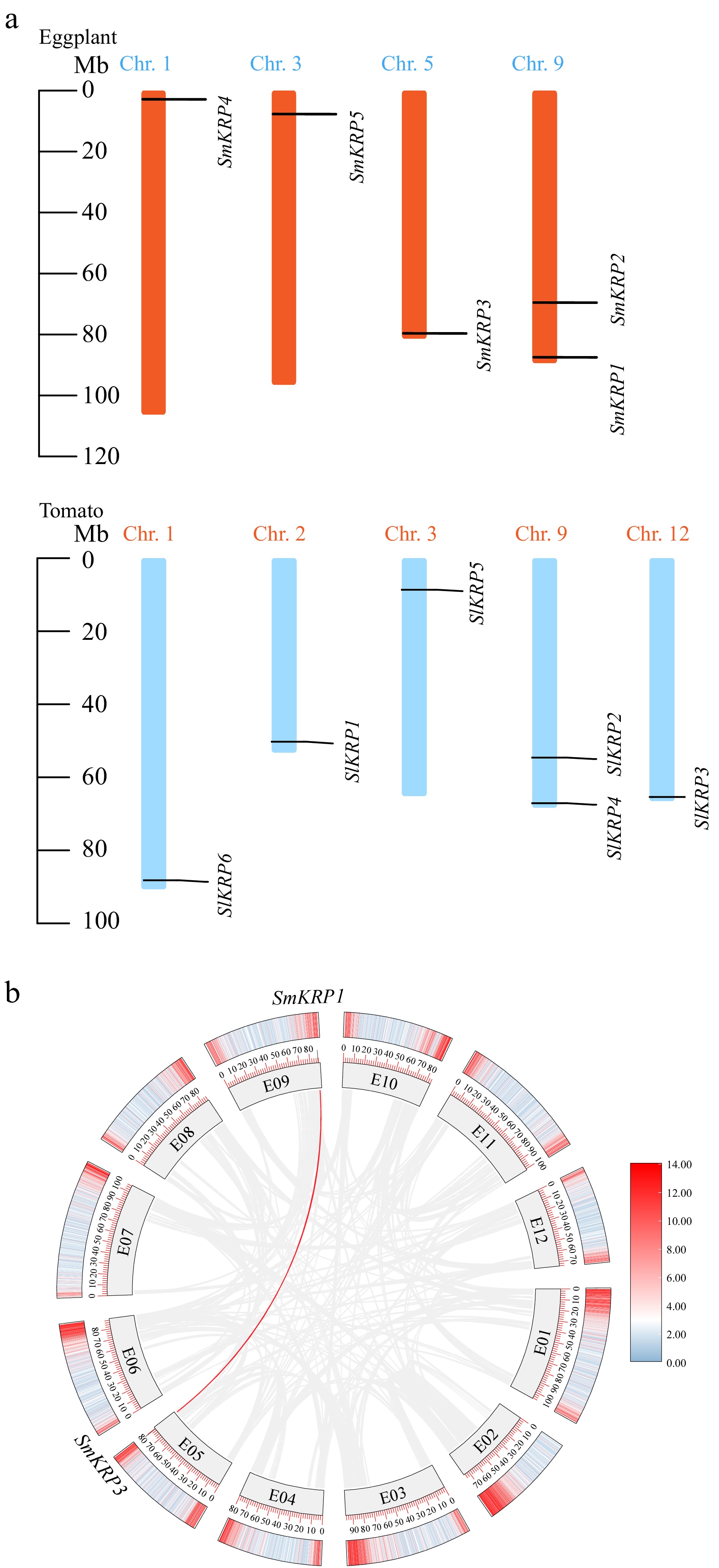
Figure 2.
(a) Chromosomal assignment of KRP members in eggplant and tomato, (b) the duplication of SmKRPs on eggplant chromosomes. The Chr. 1, Chr. 3, Chr. 5, Chr. 9, and Chr. 12 in (a) as well as E01~E12 in (b) represent the chromosome number.
The phylogenic relationship among SmKRPs were analyzed. There is a high sequence similarity between SmKRP1 and SmKRP3, and SmKRP4 and SmKRP5, respectively (Fig. 3a). The gene structures of SmKRPs were also analyzed. According to the phylogenic relationship among SmKRPs, the KRP members with higher sequence similarity have the same exon number. SmKRP1−3 have three exons, while SmKRP4 and SmKRP5 contain four exons (Fig. 3b). The conserved domain in the amino acid sequences of SmKRP proteins were analyzed by searching the SMART website and found that all the SmKRP proteins exhibited a conserved CDI domain at C terminal (Fig. 3c). Moreover, 10 conserved motifs (motif 1−10) of SmKRP proteins were identified by searching the Multiple Em for Motif Elicitation (MEME) website with amino acid sequences (Fig. 3d). The amino acid sequence length of these conserved motifs ranged from 6 to 47 aa. Motif 1 within these five SmKRPs was the CDI domain.
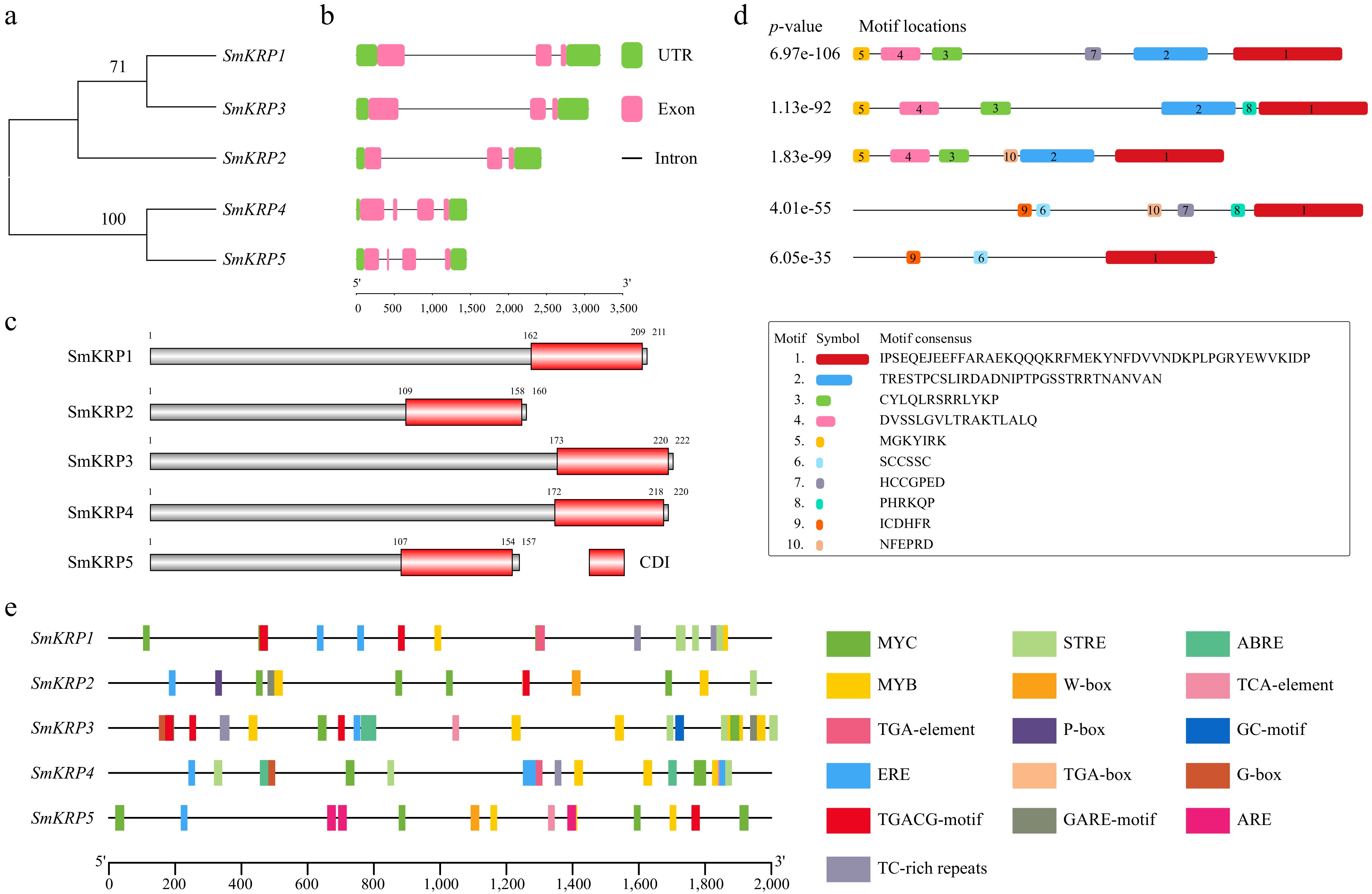
Figure 3.
Analysis of gene structure, conserved domains and motifs of SmKRPs. (a) Phylogenetic relationship analysis among SmKRPs. (b) Distribution of exon, intron, and UTR of SmKRPs. Green box, pink box and black line represent UTR, exon, and intron, respectively. (c) Conserved domains analysis of SmKRP proteins. The red box represents conserved domain CDI. (d) Distribution of conserved motifs within SmKRP proteins. The 1–10 motifs were identified by searching MEME website, and were distinguished by different color boxes. The motif 1 is CDI domain. (e) Analysis of cis-elements within the promoters of SmKRPs. The cis-elements were predicted by searching the PlantCARE website, and were differentiated by different colored boxes.
As shown in Fig. 3e, multiple cis-elements related to phytohormone response elements, transcription factor binding elements, and stress response elements were observed within these five SmKRPs promoters (Supplemental Table S5). The phytohormone response elements consisted of abscisic acid (SA) responsive element TCA-element, jasmonic acid methyl ester (MeJA) responsive element TGACG-motif, abscisic acid (ABA) responsive element ABRE, ethylene-responsive element ERE, auxin-responsive element TGA-element or TGA-box, and gibberellin responsive element GARE-motif or P-box within the promoters of SmKRPs. The transcription factor binding elements, including MYB transcription factor binding element MYB, MYC transcription factor binding element MYC, bZIP transcription factor binding element G-box, and WRKY transcription factor binding element W-box, were identified in the promoters of SmKRPs. In addition, we also found that the SmKRPs promoters contained some elements related to stress response, including the stress response element STRE, cis-acting element involved in defense and stress responsiveness TC-rich repeats, and cis-acting regulatory element essential for the anaerobic induction ARE.
Analysis of expression profiles of SmKRPs under salt and dehydration stress treatment
-
Based on the analysis of the mRNA sequencing result of eggplant roots under salt stress treatment, it was found that salt stress could only induce the up-regulation of SmKRP3 expression (Supplemental Fig. S3). To verify this result, the expression of SmKRPs in eggplant roots treated with 200 mM NaCl stress was detected by RT-qPCR assay. Salt stress could induce significant up-regulation of SmKRP3 at 6 h post-treatment. The expression of other SmKRPs exhibited a down-regulated tendency under salt stress treatment (Fig. 4a). The expression profiles of SmKRPs under dehydration stress treatment were also tested. It was found that the expression levels of all SmKRPs displayed a down-regulated trend in the condition of dehydration stress treatment (Fig. 4b), implying that SmKRPs may play a negative role in eggplant response to dehydration stress.
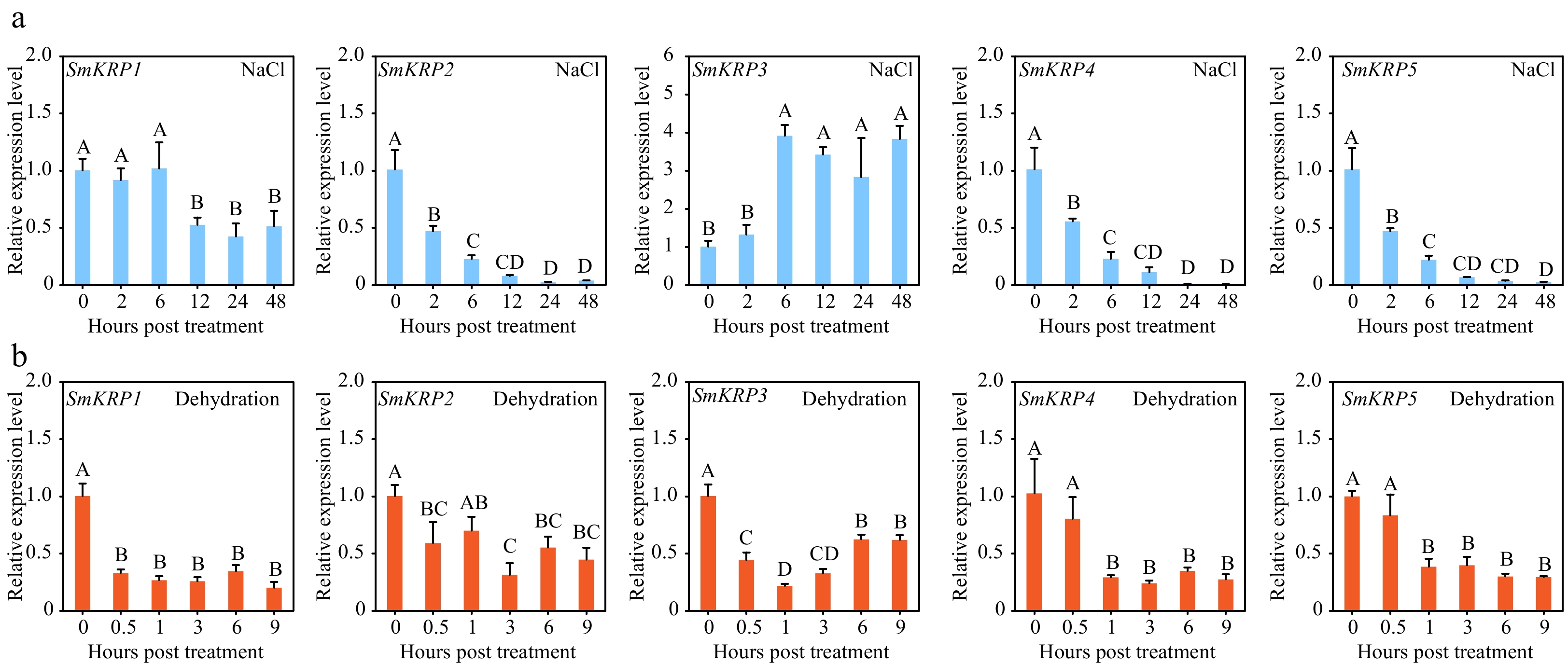
Figure 4.
Expression profile analysis of SmKRPs. Expression levels of SmKRPs were detected in eggplant roots treated with (a) NaCl or (b) dehydration stress by RT-qPCR assay. Three biological repeats were applied to calculate the mean ± standard deviation. Different upper letters represent highly significant differences, as performed by Fisher's protected LSD test (p < 0.01).
Analysis of tissues specific expression of SmKRPs
-
It is informative to understand the role of SmKRPs in different biological processes in eggplant by analyzing the tissue-specific expression of SmKRPs. An RT-qPCR assay was performed to examine the expression of SmKRPs in diverse tissues from various developmental stages of eggplants. It was observed that a higher expression of SmKRP1, SmKRP2, and SmKRP3 exhibited in the root (RT) than in the young leaf (YL) and stem (ST), while the expression of both SmKRP4 and SmKRP5 in YL were higher than that in the RT and ST in the seedlings. In mature plants, the highest expression levels of SmKRP1, SmKRP2, and SmKRP3 were recorded in the fruit (FRT) compared to the other tissues. The highest expression levels of both SmKRP4 and SmKRP5 were observed in the YL (Fig. 5).
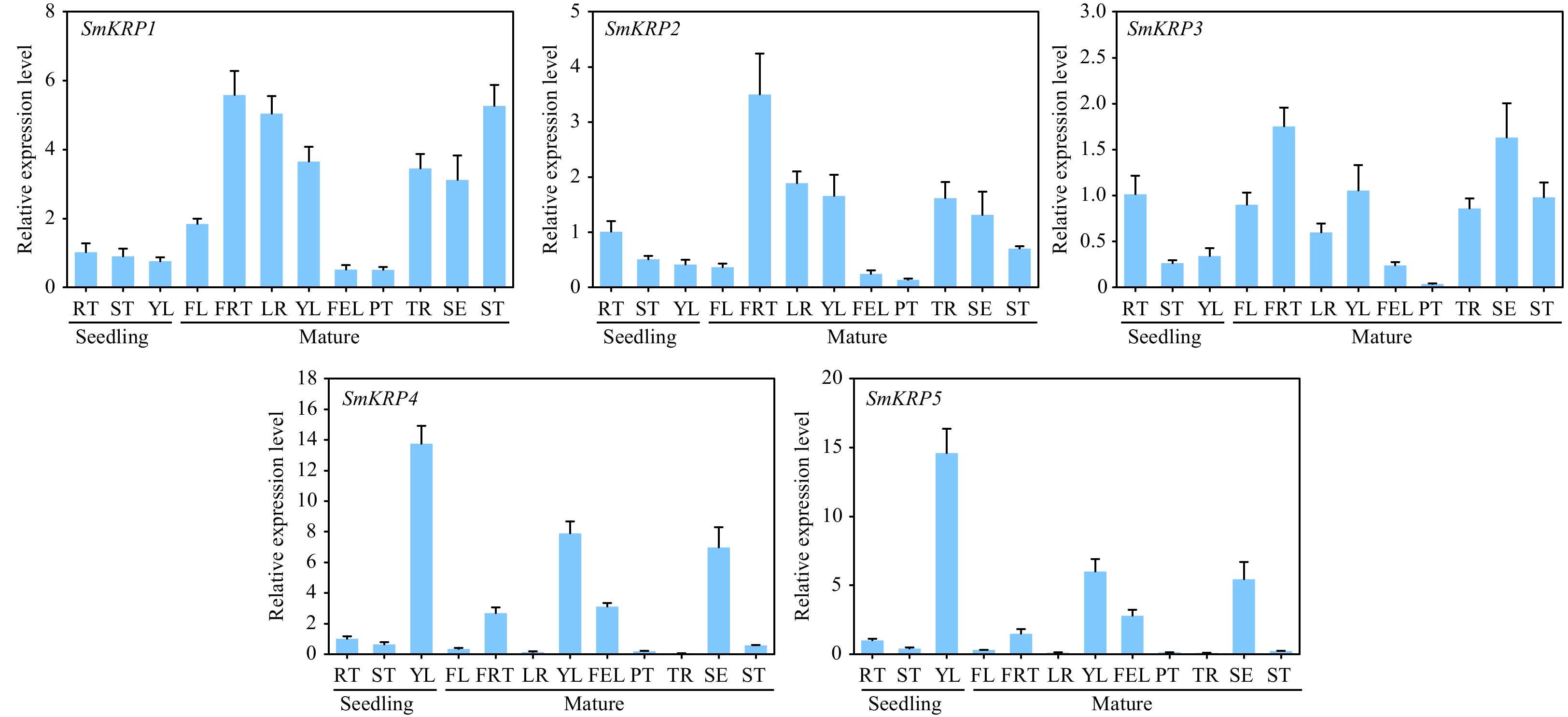
Figure 5.
Analysis of tissue specific expression of SmKRPs. The relative expression levels of SmKRPs in the different tissues from seedlings including root (RT), stem (ST), and young leaf (YL) and from mature plants including flower (FL), fruit (FRT), lateral root (LR), YL, full expand leaf (FEL), petiole (PT), taproot (TR), sepal (SE), and ST by RT-qPCR assay. Three biological repeats were applied to calculate the mean ± standard deviation. Different uppercase letters represent highly significant differences, as performed by Student's t-test (p < 0.01).
Subcellular localization analysis of SmKRP proteins
-
Next, the subcellular localization of SmKRP proteins was investigated. Firstly, SmKRP proteins' subcellular localization was predicted by searching the Plant-mPLoc (
www.csbio.sjtu.edu.cn/bioinf/plant-multi ) website with their amino acid sequences. All SmKRP proteins were predicted to be located in the nucleus (Supplemental Fig. S4a). The nuclear localization signal (NLS) within amino acid sequences of SmKRP proteins were further analyzed by searching the INSP (www.csbio.sjtu.edu.cn/bioinf/INSP ) website. Shown in Supplemental Fig. S4b, one NLS in the amino acid sequence of SmKRP2 protein was observed, while two NLS were contented in the other four SmKRP protein amino acid sequences. To further confirm whether SmKRP proteins are located in the nucleus, subcellular localization of SmKRP proteins in the epidermic cells of Nicotiana benthamiana leaves was investigated via Agrobacterium-mediated transient infiltration. The full-length CDS of SmKRPs were cloned into the destination vector pBinGFP2 (Fig. 6a). The laser scanning confocal microscope was used to observe the fluorescence signals. We observed that the green fluorescence signal of all SmKRP-GFP proteins occurred in the nucleus. The green fluorescence signal expressed by an empty vector appeared in the whole cell (Fig. 6b), suggesting that all SmKRP proteins are located in the nucleus of the epidermic cells of Nicotiana benthamiana leaves.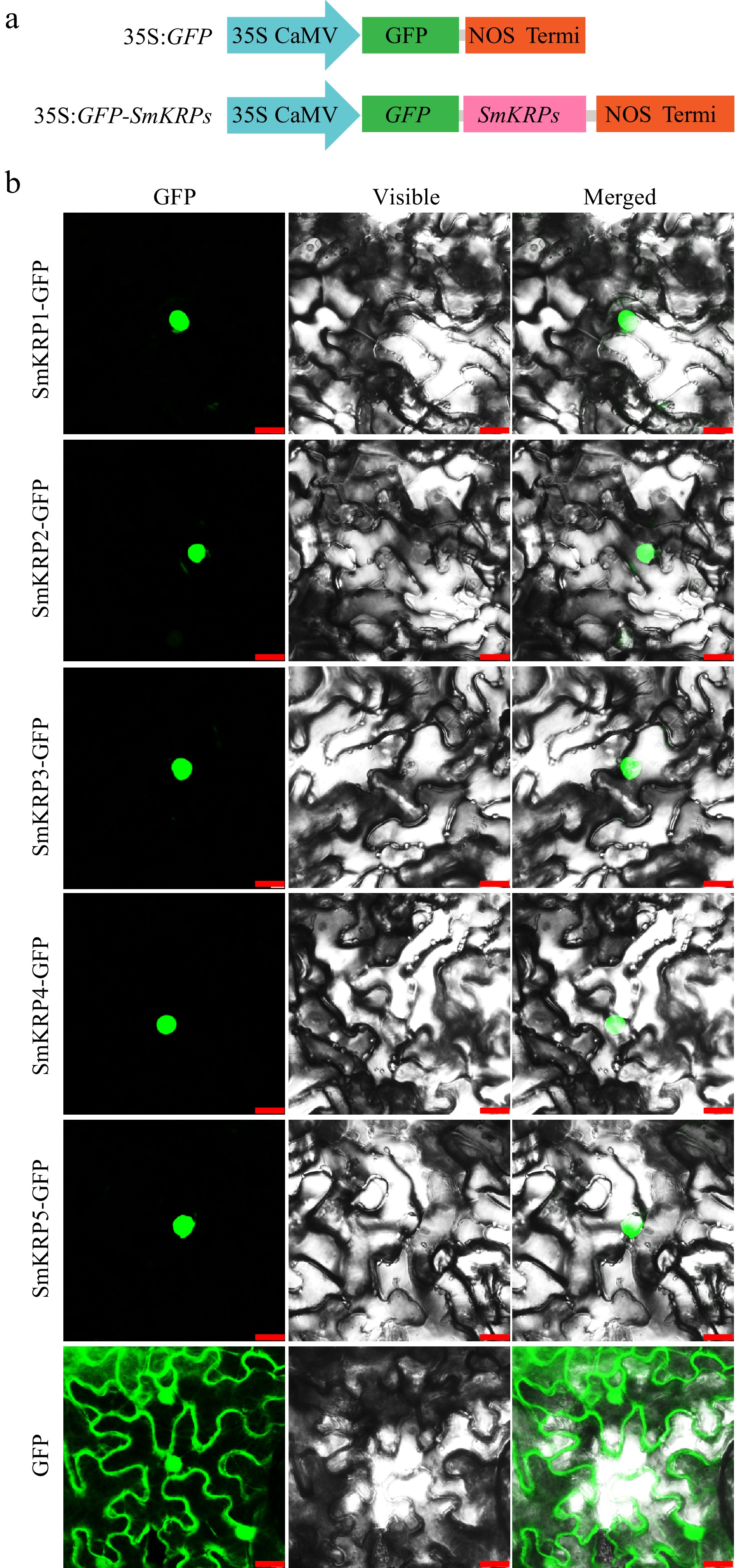
Figure 6.
Subcellular localization of SmKRP proteins. (a) Diagrammatic drawing of recombinant vector structures of 35S:GFP-SmKRPs or empty vector 35S:GFP. (b) Subcellular localization of SmKRP proteins in the epidermic cells of Nicotiana benthamiana leaves. Bar = 25 μm.
Analysis of amino acid sequence multiple alignment and phylogenetic relationship of SmKRP3
-
Due to the significant up-regulation of SmKRP3 expression under salt stress treatment, we explored the function of SmKRP3 in eggplant against salt stress. Firstly, we analyzed the multiple alignment of the SmKRP3 amino acid sequence with its homologs from the other plant species. We found that SmKRP3 amino acid sequence shares 71%, 70%, 66%, 54%, 51%, 49%, 45%, 44%, 43%, 43%, 39%, and 37% sequence similarities with its homologs from the selected plant species including Solanum tuberosum hypothetical protein KY285_009599 (KAH0733892.1), Solanum lycopersicum cyclin-dependent kinase inhibitor 4-like (NP_001304938.1), Solanum dulcamara cyclin-dependent kinase inhibitor 4-like (XP_055813395.1), Nicotiana tabacum cyclin-dependent kinase inhibitor 5-like (XP_016513903.1), Lycium ferocissimum cyclin-dependent kinase inhibitor 4-like (XP_059286725.1), Capsicum annuum cyclin-dependent kinase inhibitor 4 (XP_047259206.1), Ipomoea nil cyclin-dependent kinase inhibitor 3-like (XP_019156941.1), Gossypium hirsutum cyclin-dependent kinase inhibitor 5 (XP_016732133.1), Prunus persica cyclin-dependent kinase inhibitor 5 (ALV85618.1), Arabidopsis thaliana inhibitor/interactor with cyclin-dependent kinase (NP_199693.1), Triticum aestivum cyclin-dependent kinase inhibitor 4-like isoform X2 (XP_044449154.1), and Oryza sativa Japonica Group cyclin-dependent kinase inhibitor 4 (NP_001390949.1), respectively (Supplemental Fig. S5a). Based on the amino acid sequences of SmKRP3 homologs above, an evolutionary tree was constructed to analyze the phylogenetic relationship of these homologs. SmKRP3 exhibited a closest phylogenetic relationship with Solanum dulcamara cyclin-dependent kinase inhibitor 4-like (Supplemental Fig. S5b).
Silencing of SmKRP3 enhances the susceptibility of eggplant to salt stress
-
Additionally, the impact of SmKRP3 silencing on eggplant tolerance to salt stress was evaluated using a VIGS assay. Compared to the control eggplants, SmKRP3 expression level was significantly down-regulated in the roots of SmKRP3-silenced (TRV:SmKRP3) eggplants, and the increased regulation of SmKRP3 expression by salt stress treatment was inhibited in the SmKRP3-silenced eggplants compared to the TRV:00 control eggplants (Fig. 7a). In addition, it was also found that the SmKRP3 expression level in the leaves of SmKRP3-silenced eggplants was observably down-regulated comparing to the control plants (Supplemental Fig. S6), suggesting that silencing of SmKRP3 was successful. The SmKRP3-silenced and control eggplants were treated with 200 mM NaCl solution. After 24 h, SmKRP3-silenced eggplants exhibited more obvious wilt symptom than that of control plants (Fig. 7b). The lower survival rate was recorded in the SmKRP3-silenced eggplants comparing to the control under salt stress treatment after 4 d (Fig. 7c), revealing that silencing of SmKRP3 enhanced eggplant susceptibility to salt stress. Moreover, silencing of SmKRP3 induced overt down-regulation of expression levels of salt stress defense-related marker genes including SmGSTU10[41], SmNCED1[28], SmDHN1[30], and SmDHNX1[30] under salt stress treatment (Fig. 7d). SmKRP3-silenced and control eggplants treated with salt stress showed physiological indices, including APX and CAT enzyme activity. An obvious decrease of APX and CAT enzyme activities were observed in the roots of SmKRP3-silenced eggplants compared to the control plants (Fig. 7e). These data revealed that SmKRP3 positively regulates salt stress tolerance in eggplants.
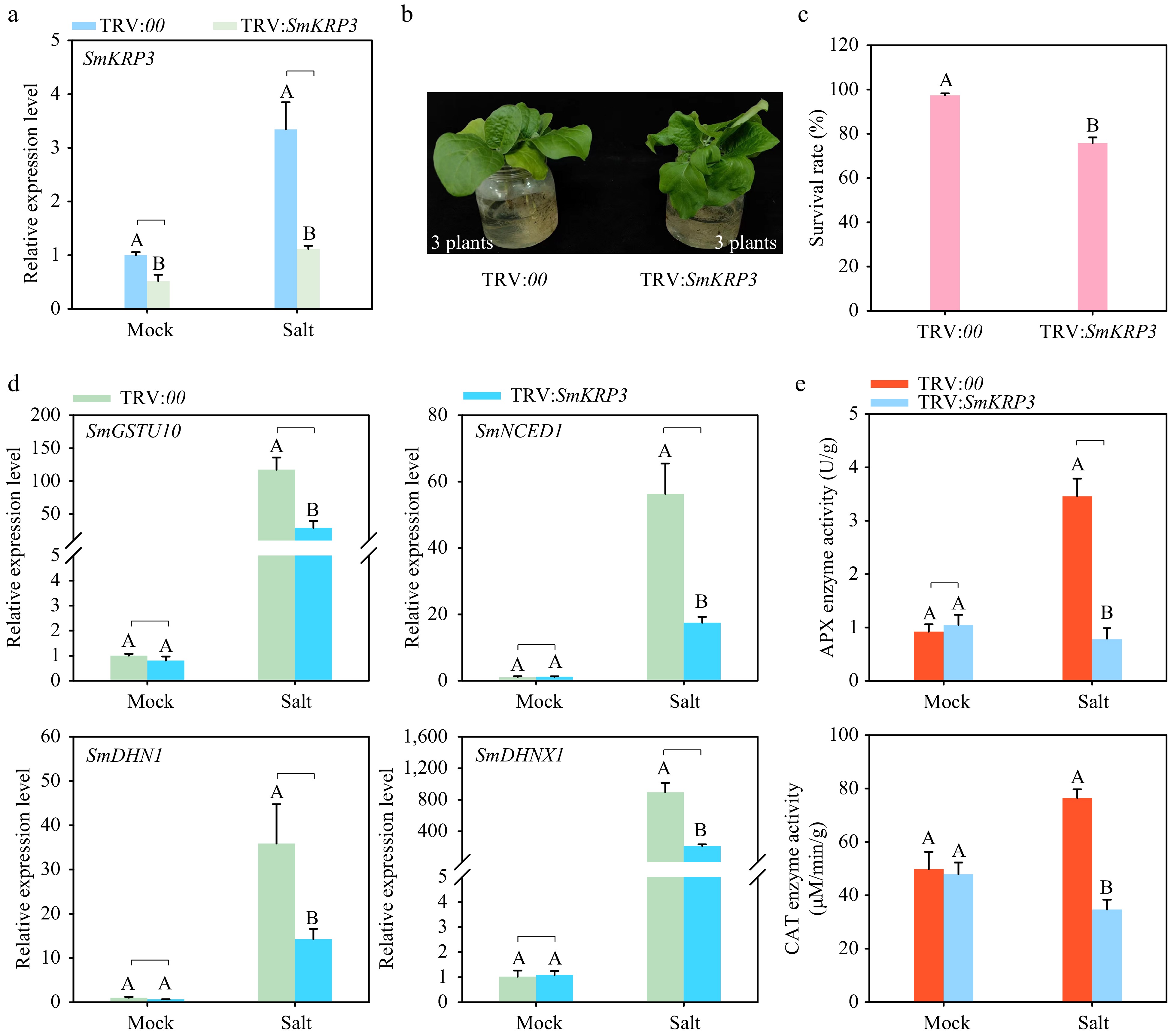
Figure 7.
Silencing of SmKRP3 enhances susceptibility of eggplant to salt stress. (a) Detection of silencing efficiency of SmKRP3. (b) Phenotype analysis of SmKRP3-silenced or control eggplants under salt stress treatment after 24 h post treatment. (c) Survival rate analysis of SmKRP3-silenced or control eggplants under salt stress treatment at 4 d post salt stress treatment. (d) Detection of expression levels of salt stress defense related marker genes SmGSTU10, SmNCED1, SmDHN1, and SmDHNX1 in the roots of SmKRP3-silenced or control eggplants at 24 h post salt stress treatment. (e) Measurements of APX and CAT enzyme activities in SmKRP3-silenced or control eggplant roots at 48 h post salt stress treatment. Three biological repeats were applied to calculate the mean ± standard deviation. Different uppercase letters represent highly significant differences, as performed by Student's t-test (p < 0.01).
-
Plant needs to create new organs via rapid cell division and differentiation to repair the damage caused by multiple abiotic and biotic stresses[42,43]. Acted as an inhibitor of CDKs activity, KRPs play a vital role in regulating the cell cycle process[44]. So far, the KRP genes have been identified in certain plant species, such as Arabidopsis thaliana[17], tomato[18], and soybean[19], and mainly function in the plant's growth and development processes. However, the information of KRP gene family members in plants and their functions in plant response to abiotic stresses were largely unknown. Herein, we identified five KRP genes in the eggplant genome, further analyzed the sequences, structures, and expression of SmKRPs, and explored the functions of SmKRP3 in eggplant response to salt stress.
In the present study, five KRP members in the eggplant HQ-1315 genome were identified, named SmKRP1−5. Interestingly, a previous study reported six KRP members in eggplant '67/3' (v3.0 and v4.1) genome[18]. Six KRP members in the genome of eggplant variety GUIQIE-1 were also identified (Supplemental Table S2). This difference in the number of KRP members is due to the distinction of amino acid sequences and conserved domain between SMEL_002g167200 or SMEL4.1_09g013240.1 protein in eggplant '67/3' (v3.0 and v4.1), EGP31387.1 protein in GUIQIE-1, and its homolog Smechr0202706 protein in eggplant HQ-1315 (Supplemental Fig. S1). The deficiency of the conserved CDI domain of the Smechr0202706 protein may be likely due to genetic evolution or mutations in eggplant HQ-1315. We also found that there was no change in the number of KRP members in Solanaceae plants such as six KRP genes in potatoes, six KRP genes in tobacco, six KRP genes in pepper, and six KRP genes in tomato (Supplemental Table S3), suggesting that KRP gene family in Solanaceae probably did not expand during evolution. We next generated an evolutionary tree of SmKRPs with its homologs from Arabidopsis thaliana, tomato, and pepper to analyze their phylogenetic relationship. Five SmKRPs were classified into three classes (Fig. 1), consistent with the Arabidopsis thaliana report[17]. In addition, it was found that SmKRP1 is more closely related to SmKRP3 among the SmKRPs by analyzing the phylogenetic relationship of SmKRPs (Fig. 3a), implying that segment duplication may occur between SmKRP1 and SmKRP3. The result of chromosomal assignment analysis revealed that SmKRPs located on four chromosomes, including Chr. 1, 3, 5, and 9 (Fig. 2a). The inter-chromosome segment duplication and tandem duplication are important ways of genetic variation and species evolution in plants[45]. The collinearity relationship between SmKRP1 and SmKRP3 was observed, suggesting that a duplicated segment indeed exhibited between SmKRP1 and SmKRP3 (Fig. 2b). Previous study showed that SlKRP1 and SlKRP3, SlKRP1 and SlKRP5, SlKRP1 and SlKRP6, as well as SlKRP3 and SlKRP4 exhibit inter-chromosomal fragment replication in tomato[18]. At the same time, only SmKRP1 and SmKRP3 occur in inter-chromosome segment duplication in eggplant, suggesting that SmKRPs may be more conservative in structures and functions in eggplant. However, we did not observe the collinearity relationship among the KRP members in eggplant, Arabidopsis thaliana, and tomato (Supplemental Fig. S2), implying that KRP genes in these three plant species may have evolved independently. The results of gene structures and conserved motifs analysis showed that the similar gene structures and motifs were observed among the closely related genes such as SmKRP1, SmKRP2, and SmKRP3 (Fig. 3b, d). Multiple cis-elements including phytohormone response, transcription factor binding, and stress response related elements were contained within the promoters of SmKRPs (Fig. 3e), suggesting that SmKRPs expression levels were regulated by multiple phytohormone and transcription factor components in different biological processes.
Previous studies revealed that KRP genes mainly function in the processes of growth and development in plants[19,21,22,46−48]. However, a few studies reported that KRP genes were involved in the plant's response to abiotic stresses. In tomato, only a few SlKRP expression levels were up-regulated by abiotic stresses. Drought and cold stresses could only up-regulate the expression of SlKRP3 and SlKRP2, respectively[18]. Salt stress could up-regulate GmKRP2b, GmKRP4, and GmKRP5 expression[19]. A similar expression pattern of KRPs in this study was observed in that only SmKRP3 expression level was up-regulated by salt stress. In contrast, a declining trend of SmKRPs expression was recorded under dehydration stress treatment (Fig. 4a, b). The result of tissues specific expression of SmKRPs in the diverse tissues from different developmental stages of eggplants revealed that SmKRP1–3 have higher expression levels in RT in the seedlings and in FRT in the mature plants, while SmKRP4 and SmKRP5 have higher expression levels in YL both in the seedlings and mature plants (Fig. 5), suggesting that SmKRPs mainly function in the growth and development in root, leaf, and fruit. Subsequently, subcellular localization of SmKRP proteins was investigated in epidermic cells of Nicotiana benthamiana leaves, and it was found that all SmKRP proteins located in the nucleus (Fig. 6b), which was similar to a previous study[18]. To investigate the function of SmKRPs in eggplant response to abiotic stresses, the function of SmKRP3 in eggplant response to salt stress was explored as SmKRP3 expression was up-regulated by salt stress treatment. Based on the VISG assay, we found that silencing of SmKRP3 enhanced the susceptibility of eggplants to salt stress (Fig. 7b) and significantly down-regulated the expression levels of salt stress defense-related genes SmGSTU10, SmNCED1, SmNDH1, and SmNDHX1 (Fig. 7d), and decreased APX and CAT enzyme activities (Fig. 7e), indicating that SmKRP3 positively functions in eggplant response to salt stress. Abiotic stresses such as salt, drought stress could cause oxidation reaction and DNA damage, which resulted in cell cycle retention[49]. The cell damage caused by abiotic stresses was alleviated by the interaction between CDK and KRP to maintain cell cycle process to generate new cells[49]. Therefore, SmKRP3 may be involved in the process of regulating the cell cycle to promote plant cell division, leading to alleviate the damage to eggplant caused by salt stress and increase tolerance of eggplant to salt stress.
-
In this study, five KRP gene family members were identified in the eggplant genome located on the four chromosomes and divided them into three classes by phylogenetic relationship analysis. A collinearity relationship between SmKRP1 and SmKRP3 was exhibited. The study on gene structures, conserved motifs and domains, and cis-elements within SmKRPs promoters revealed the evolutionally conserved function of SmKRPs. The nucleus locations of SmKRP proteins were observed, and the high expression of SmKRPs in young leaf (YL), fruit (FRT), and root (RT). In addition, except for the up-regulation of SmKRP3 under salt stress treatment, the other SmKRPs expression displayed unchanged or declining trends under salt or dehydration stress treatment. Silencing of SmKRP3 enhanced the susceptibility of eggplant to salt stress, suggesting that SmKRP3 positively regulates the salt stress tolerance in eggplants. The reported data provides new insight into the KRP gene family of eggplant and the function of SmKRPs in eggplant response to abiotic stresses.
-
The authors confirm contribution to the paper as follows: study conception and design: Shen L, Yang X; data collection: Shen L, Yang S, Xia X; analysis and interpretation of results: Shen L, Nie W; draft manuscript preparation: Shen L; manuscript revision: Shen L, Nie W, Yang X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This work was supported by the National Natural Science Foundation of China (32102385). We are very grateful to the reviewers' suggestions for improving our paper's quality. In addition, we are thankful to Rahat Sharif (College of Horticulture and Landscape Architecture, Yangzhou University, China) for revising the language of the manuscript.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Lei Shen, Shixin Yang
- Supplemental Table S1 The primer pairs sequences used in this study.
- Supplemental Table S2 The KRP amino acid sequences in the eggplant variety GUIQIE-1.
- Supplemental Table S3 The KRP amino acid sequences in Solanaceae plants including potato, tomato, pepper, and tobacco.
- Supplemental Table S4 Analysis of homology and sequence similarity of the KRP members between eggplant and tomato.
- Supplemental Table S5 The promoter sequences of KRP members in the eggplant variety HQ-1315.
- Supplemental Fig. S1 Amino acid sequences alignment (a) and conserved domain analysis (b) among Smechr0202706, SMEL_002g167200, SMEL4.1_09g013240.1, and EGP31387.1.
- Supplemental Fig. S2 Collinearity relationship analysis among the KRP members in the eggplant, Arabidopsis thaliana, and tomato.
- Supplemental Fig. S3 Expression analysis of SmKRPs in eggplant roots at 0 and 24 h post salt stress treatment according to mRNA-seq data. The heat map was generated by TBtools software.
- Supplemental Fig. S4 Prediction of subcellular localization(a) and NLS (b) of SmKRP proteins. The subcellular localization and NLS of SmKRP proteins were predicted by searching Plant-mPLoc and INSP website, respectively.
- Supplemental Fig. S5 Analysis of amino acid sequence multiple alignment (a) and phylogenetic relationship (b) of SmKRP3 with its homologs. The black and pink shadow indicate 100% and 75 ~ 100% sequences identity, respectively.
- Supplemental Fig. S6 Detection of silencing efficiency of SmKRP3 in the leaves of SmKRP3-silenced or control eggplants. Three biological repeats were applied to calculate the mean ± standard deviation. Different upper letters represent highly significant differences, as performed by Student's t-test (p < 0.01).
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Shen L, Yang S, Xia X, Nie W, Yang X. 2024. Genome-wide identification of Kip-related protein (KRP) gene family members in eggplant and the function of SmKRP3 under salt stress. Vegetable Research 4: e013 doi: 10.48130/vegres-0024-0012 

Genome-wide identification of Kip-related protein (KRP) gene family members in eggplant and the function of SmKRP3 under salt stress
- Received: 07 February 2024
- Revised: 31 March 2024
- Accepted: 07 April 2024
- Published online: 08 May 2024
Abstract: Kip-related proteins (KRPs) are essential to cell division throughout their whole process of growth and development. Research on KRPs and their roles has mostly focused on Arabidopsis thaliana. Nonetheless, details on the structure and functions of KRP gene family members in eggplant are still unclear. In this study, we have discovered and analyzed five KRP genes in the eggplant HQ-1315 genome. Our investigation focused on determining their chromosomal position, collinearity, gene structure, conserved motif, and domain, as well as cis-element. The expression levels of all KRP genes were decreased by salt and dehydration stress, except SmKRP3. The expression of SmKRP3 was significantly increased by salt stress. Transient expression assay revealed that all of the eggplant KRP proteins are located in the nucleus of epidermic cells of Nicotiana benthamiana leaves. Silencing of SmKRP3 increased the sensitivity of eggplant to salt stress. This was accompanied by a significant decrease in the expression of salt stress marker genes SmGSTU10, SmNCED1, SmDHN1, and SmDHNX1. An apparent decrease in the activity of the enzymes ascorbate peroxidase (APX) and catalase (CAT). Our findings show that SmKRP3 has a positive role in salt stress, shedding fresh light on the complete information of the KRP gene family members in eggplants.
-
Key words:
- Eggplant /
- Solanum melongena /
- SmKRP3 /
- Salt stress /
- Kip-related protein (KRP)