








-
As a major driver of biodiversity loss, invasive plant species threaten the natural environment and human health globally[1−7]. Invasive plant species not only disrupt natural community assembly but also act as one of the most significant impediments to restoring native ecosystems[7,8]. Bidens pilosa is a widespread weed in tropical, subtropical and warm temperate regions[9]. This weed is a diverse annual herb native to tropical and Central America[9] and is considered an invasive weed in Thailand[10]. Even though several studies have been conducted on fungi associated with Bidens pilosa[11−13], the comprehensive fungal diversity of Bidens pilosa is still yet to be understood.
Fungi associated with Bidens pilosa have been observed in some studies, and the reported species belong to the families Albuginaceae, Apiosporaceae, Botryosphaeriaceae, Ceratobasidiaceae, Cladosporiaceae, Diaporthaceae, Erysiphaceae, Glomerellaceae, Mycosphaerellaceae, Nectriaceae, Periconiaceae, Peronosporaceae, Phyllostictaceae, Sclerotiniaceae, Stachybotryaceae, Tetraplosphaeriaceae, and Torulaceae[11−14]. However, species of Didymosphaeriaceae have not been reported from Bidens pilosa yet. Didymosphaeriaceae, as introduced by Munk[15], comprises 33 genera[16]. Members of this family can be found as endophytes, pathogens and saprobes in soil as well as various host plants from both aquatic and terrestrial habitats[17−24]. Among them, Chromolaenicola, which was introduced by Mapook et al.[17], accommodated seven accepted species[16,25,26], including four asexual morph (Chromolaenicola chiangraiensis, C. clematidis, C. lampangensis, and C. siamensis) and three sexual morph taxa (C. nanensis, C. sapinda and C. thailandensis)[17,27]. Chromolaenicola species occur as saprobes in terrestrial habitats in China and Thailand[17,27−29]. Members of Chromolaenicola were found from different plant families, viz., Asteraceae, Bromeliaceae, Fabaceae, Ranunculaceae, and Spindaceae[17,27−30]. Some Chromolaenicola species, such as C. lampangensis and C. nanensis, showed potential antimicrobial properties[17]. The sexual morph of Chromolaenicola is characterized by its immersed to semi-immersed, solitary or scattered, coriaceous, globose to subglobose ascomata with an ostiole, 3–4 layers of textura angularis cells, cylindrical to filiform, septate, branching pseudoparaphyses, 6–8-spored, bitunicate, cylindrical, pedicellate asci with an ocular chamber, and hyaline to brown, uniseriate, ellipsoid to broadly fusiform, muriform spores without gelatinous sheath. The asexual morph is characterized by its immersed to semi-immersed, globose to obpyriform, pycnidia conidiomata with an ostiole, 2–4 layers of textura angularis cells, hyaline and unbranched, smooth, elongated, broadly filiform to ampulliform conidiogenous cells, and oblong or oval to ellipsoid, globose to subglobose, hyaline to pale brown, aseptate to 1-septate conidia. The linkage of sexual and asexual morph from Chromolaenicola has not been reported yet.
In the current study, morphological descriptions and photo plates of Chromolaenicola species collected in northern Thailand are presented. Both multi-gene analyses and morphological comparisons were used to confirm the species identification. The preliminary antibacterial screening was also conducted and presented for all Chromolaenicola species.
-
Dead stems of Bidens Pilosa were collected in March from the roadsides in Doi Pui District, Chiang Rai Province, Thailand. All specimens were kept in plastic bags labeled with collection details and taken to the laboratory. Single spore isolation was carried out on malt extract agar (MEA), following the methods by Senanayake et al.[31] and kept for 24 h at room temperature. The spore germination was observed within 24 h using a Motic SMZ 168 Series microscope. Germinated spores were transferred to new MEA plates. All the micro-morphological characters were observed using a Nikon ECLIPSE 80i compound microscope (Nikon, Japan) fitted to a Canon 550D digital camera (Canon, Japan). Tarosoft Image Framework (v 0. 9.7) was used to measure photomicrograph structures. Adobe Photoshop CS6 Extended (v 10.0.) was used to edit and prepare photo plates (Adobe Systems, USA). Forty-day-old cultures were used for molecular studies. Specimens were deposited at the herbaria of Mae Fah Luang University (Herb. MFLU) while living cultures were maintained at Mae Fah Luang University culture collection (MFLUCC). Faces of fungi (FoF) numbers and Index Fungorum (IF) numbers were obtained as instructed by Jayasiri et al.[32] and Index fungorum[25]. Moreover, the species descriptions were submitted to GMS Microfungi[33].
DNA extraction, PCR amplification and sequencing
-
Genomic DNA was extracted from forty-day-old mycelium using E.Z.N.A.® Tissue DNA Kit (Omega Biotek Inc.), following the manufacturer’s instructions. DNA amplifications were performed by polymerase chain reactions (PCR), following Mapook et al.[34]. The details of PCR primers and protocols are shown in Table 1. The quality of PCR products was confirmed on 1% gels. The PCR products were sent to a commercial sequencing provider (Solgent Co., Ltd, Thailand). The newly generated nucleotide sequences were deposited in GenBank, and accession numbers were obtained (Table 2).
Table 1. PCR conditions used in this study.
Gene Primers PCR conditions Ref. Forward Reverse Large subunit (LSU) LR0R LR5 95 °C: 3 min, (94 °C: 30 s, 56 °C: 50 s, 72 °C: 1 min) × 40 cycles 72 °C: 7 min [35] Small subunit (SSU) NS1 NS4 95 °C for 3 min, (94 °C: 30 s, 55 °C: 50 s, 72 °C :1 min) × 40 cycles 72 °C: 7 min [36] Internal transcribed spacer (ITS) ITS5 ITS4 95 °C for 3 min, (94 °C: 30 s, 55 °C: 50 s, 72 °C :1 min) × 40 cycles 72 °C: 7 min [36] Elongation factor-1 alpha (tef1- α) EF-1 983F EF1-2218R 95 °C: 3 min, (94 °C: 30 s, 55 °C: 50 s, 72 °C: 1 min) × 40 cycles 72 °C: 7 min [37] RNA polymerase II subunit (rpb2) fRPB2-5 F fRPB2-7cR 95 °C: 5 min, (95 °C :1 min, 52 °C: 2 min, 72 °C: 90 s) × 40 cycles 72 °C: 10 min [38] Table 2. List of taxa, specimens and sequences used in phylogenetic analyses.
Species Strain numbers GenBank accession numbers Ref. LSU SSU ITS tef1-α rpb2 Bimuria omanensis SQUCC 15280 NG_071257 N/A NR_173301 MT279046 N/A [39] B. novae-zelandiae CBS 107.79 MH872950 NA MH861181 NA N/A [40] Chromolaenicola ananasi MFLU 23-0167 OR438811 OR458332 OR438340 OR500305 N/A [30] C. clematidis MFLUCC 17-2075T MT310601 MT214554 MT226671 N/A N/A [29] C. chiangraiensis MFLUCC 17-1493 MN325005 MN325011 MN325017 MN335650 MN335655 [17] C. chiangraiensis MFLUCC 24-0058 PP464125 PP464129 PP464138 PP474193 PP474190 This study Chromolaenicola nanensis MFLUCC 17-1477 MN325002 MN325008 MN325014 MN335647 MN335653 [17] C. nanensis MFLUCC 17-1473 MN325003 MN325009 MN325015 MN335648 MN335653 [17] C. lampangensis MFLUCC 17-1462 MN325004 MN325010 MN325016 MN335649 MN335654 [17] C. siamensis MFLUCC 17-2527 NG_066311 N/A NR_163337 N/A N/A [28] C. siamensis MFLUCC 24-0057 PP464124 PP464128 PP464137 PP474192 PP474189 This study C. sapindi KUMCC 21-0564T OP059009 OP059058 OP058967 OP135943 N/A [27] C. sapindi KUMCC 21-0594 OP059010 OP059059 OP058968 OP135944 N/A [27] C. thailandensis MFLUCC 17-1510 MN325006 MN325012 MN325018 MN335651 N/A [17] C. thailandensis MFLUCC 17-1475 MN325007 MN325013 MN325019 MN335652 MN335656 [17] C. thailandensis MFLUCC 24-0056 PP464123 PP464127 PP464136 PP474191 PP474188 This study Deniquelata barringtoniae MFLUCC 11−0422 JX254655 JX254656 NR_111779 N/A N/A [41] D. quercina ABRIICC 10068 MH316157 MH316155 MH316153 N/A N/A [42] Didymocrea leucaenae MFLUCC 17−0896 NG_066304 MK347826 NR_164298 MK360052 N/A [28] D. sadasivanii CBS 438.65 DQ384103 DQ384066 MH870299 N/A N/A [40] Letendraea cordylinicola MFLUCC 11−0148 NG_059530 NG_068362 NR_154118 N/A N/A [41] L. helminthicola CBS 884.85 AY016362 AY016345 MK404145 MK404174 N/A [43] Montagnula acaciae MFLUCC 18−1636 ON117298 ON117267 ON117280 ON158093 N/A [44] M. acaciae NCYUCC 19−0087 ON117299 ON117268 ON117281 ON158094 N/A [44] M. aloes CPC 19671 JX069847 N/A JX069863 N/A N/A [45] M. aloes CBS 132531 NG_042676 N/A NR_111757 N/A N/A [40] Periconia pseudodigitata KT1395 AB807564 AB797274 LC014591 AB808540 N/A [46] P. pseudodigitata KT1195A AB807563 AB797273 LC014590 AB808539 N/A [46] T: Type strains; Abbreviations of culture collections: CBS: Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands, CPC: Working collection of Pedro Crous housed at CBS, KT: K. Tanaka, MFLUCC: Mae Fah Luang University Culture Collection, Chiang Rai, Thailand, NCYUCC: National Chiayi University Culture Collection, Taiwan. SQUCC: Sultan Qaboos University Culture Collection, Sultanate of Oman. ABRIICC: Agricultural Biotechnology Research Institute of Iran Culture Collection, Iran. Sequences generated in the current study are in bold. N/A: Not available. Sequence alignment and phylogenetic analyses
-
Newly generated forward and reverse sequences were assembled in the SeqMan[47]. The assembled sequences were used for BLAST searches at NCBI (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ). Based on previously published data[17,27,30] and BLAST search results, taxa were selected, and phylogenetic analyses were conducted using the combined LSU, SSU, ITS, tef1-α, and rpb2 sequence data. Sequence alignments were made with the MAFFT v. 7 online tool (http://mafft.cbrc.jp/alignment/server ; 2016). Using MEGA v. 6.0, alignments were improved where necessary, and composite sequence alignments were obtained[48].Maximum likelihood (ML) and Bayesian inference (BI) analyses were conducted using the combined dataset of LSU, SSU, ITS, tef1-α, and rpb2[49,50]. RA × ML and Bayesian analyses were carried out on the CIPRESS Science Gateway Portal (
www.phylo.org ) using the methods described by Miller et al.[51]. Maximum likelihood analysis was performed by RA × ML-HPC v.8[52] with rapid bootstrap analysis, followed by 1000 bootstrap replicates and the GTRGAMMA substitution model. MrBayes was used to perform BI analysis on XSEDE 3.2.7[53], with tree samples taken at every 100th generation during the 5M generation run of four concurrent Markov chains. The first 25% of the trees were removed as part of the burn-in phase, and calculations for the Posterior Probability were made for the remaining 75% of the trees (PP)[49,54]. The phylogenetic tree was displayed using Fig Tree v1.4.0[55] and was modified in Microsoft Office PowerPoint v. 2013.Preliminary screening for antibacterial activity
-
Preliminary screening for antimicrobial activity was carried out following the methods of Mapook et al.[17]. Antibacterial discs of Ampicillin were used as a positive control for screening[56]. Preliminary antibacterial activities were tested against Bacillus subtilis (TISTR 1248), Escherichia coli (TISTR 527), and Staphylococcus aureus (TISTR Y4b) using the agar plug diffusion method[57]. Bacterial test organisms were grown on Nutrient Agar (NA) for 24 h. After 24 h of inoculation, 2–3 loops of the bacterial test organisms were transferred to the nutrient broth. Before adding microbial suspensions to the sterile Mueller-Hinton agar media, cell counts were performed on the suspensions (6.7 × 105 cells/mL), as detailed by Mapook et al.[17]. Fungal mycelium plugs from our isolates were transferred to a solid medium plate and allowed to grow at room temperature for 24–48 h. Inhibition zones were measured and compared to the positive control.
-
Phylogenetic analysis was conducted from the combined LSU, SSU, ITS, tef1-α, and rpb2 sequence data of twenty-eight strains, of which three were newly sequenced, while the other 23 strains were obtained from BLAST search (NCBI) and recent papers[17,27,29,30]. Periconia pseudodigitata strains KT1395 and KT1195A were selected as the outgroup. The best-scoring RA×ML tree with a final likelihood value of −9277.475085 is presented (Fig. 1). The matrix had 532 distinct alignment patterns, with 15.55% of undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.235845, C = 0.255030, G = 0.269910, T = 0.239215; substitution rates: AC = 1.648158, AG = 2.210120, AT = 1.275667, CG = 1.033223, CT = 7.205216, GT = 1.000000; gamma distribution shape parameter α = 0.110629.
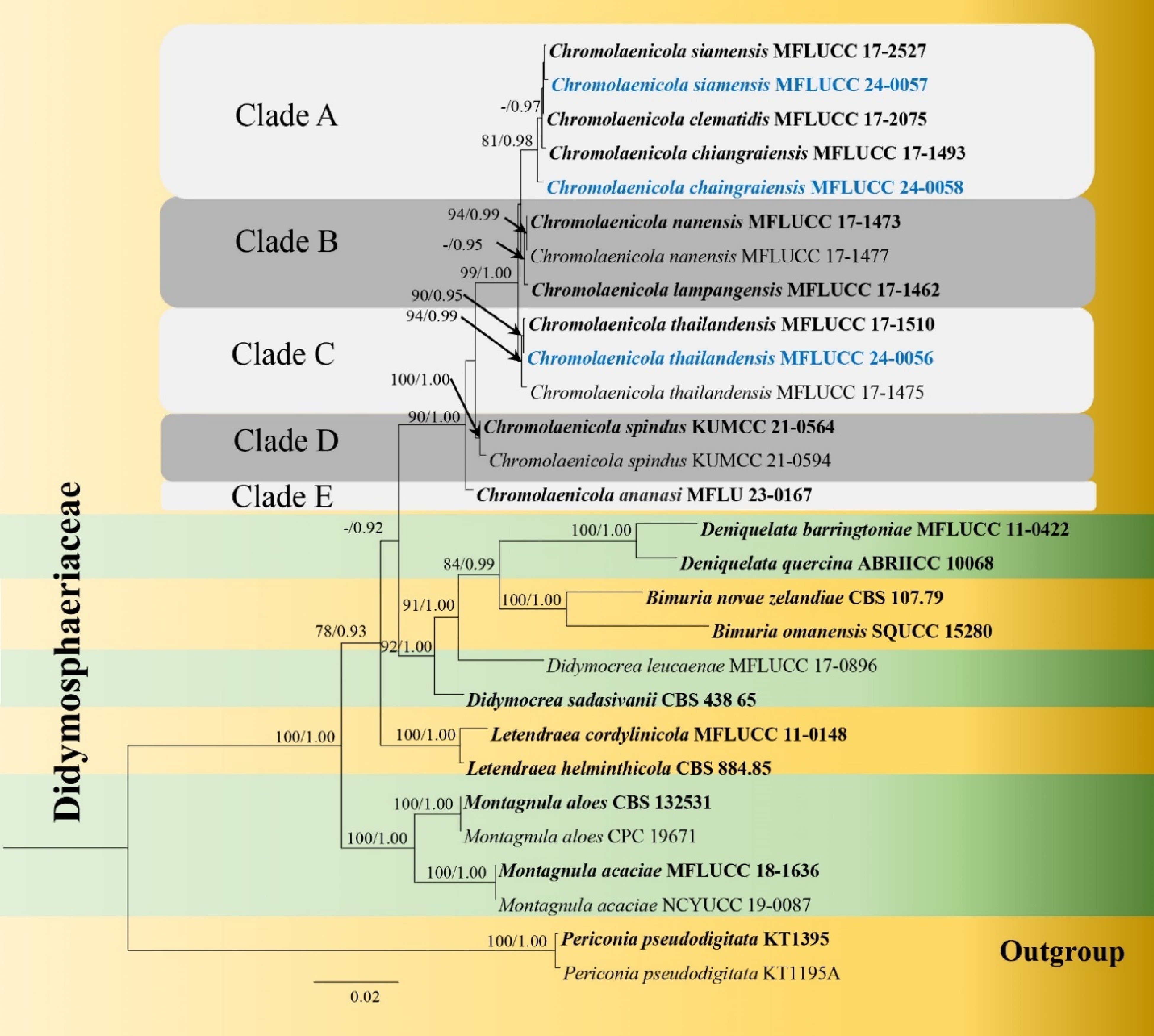
Figure 1.
Phylogram generated from maximum likelihood analysis based on the combined dataset of LSU, SSU, ITS, tef1-α and rpb2 sequence data. Bootstrap support values for ML equal to or greater than 75% and BYPP equal to or greater than 0.95 are given at the nodes. Newly generated sequences are in blue and type species are in bold.
Tree topologies of ML and BI criteria were comparable to and consistent with earlier investigations[17,27]. Chromolaenicola formed an independent topmost clade in the phylogenetic tree. Chromolaenicola species were divided into five clades (Clades A, B, C, D and E) in the phylogenetic tree. Our three strains, represented by three species, were grouped within the genus (Fig. 1). Chromolaenicola chiangraiensis (MFLUCC 24-0058) and C. siamensis (MFLUCC 24-0057) were placed in Clade A. Chromolaenicola chiangraiensis (MFLUCC 24-0058) formed a separate branch with 81% ML and 0.98 BYPP support. Our strain, Chromolaenicola siamensis (MFLUCC 24-0057) clustered with C. siamensis (MFLUCC 17-2527) with 55% ML and 0.81 BYPP. Three Chromolaenicola thailandensis strains (MFLUCC 17-1510, MFLUCC 17-1475, MFLUCC 24-0056) clustered together with 94% ML and 0.99 BYPP support and formed a distinct lineage in Clade C.
Taxonomy
-
Didymosphaeriaceae Boonmee and K.D. Hyde, Fungal Diversity 80: 462 (2016).
Chromolaenicola chiangraiensis Mapook & K.D. Hyde Fungal Diversity 101, 1–175 (2020). (Fig. 2).
Index fungorum number: IF557280, Faces of fungi number: FOF 07784

Figure 2.
Chromolaenicola chaingraiensis (MFLU 24-0030, new host record). (a), (b) Colonies on the substrate. (c)–(f) Conidia and conidiogenous cells. (g)–(j) Conidia. (k) Germinating conidia. (l) Culture on MEA. Scale bars: (a), (b) = 500 µm, (c) = 30 µm, (d)–(j) = 10 µm, (k) = 20 µm, (l) = 10 mm.
Saprobic on dead stems of Bidens pilosa. Sexual morph: Undetermined. Asexual morph: appearing as Colonies on the host substrate, superficial, scattered, gregarious, black. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 2–4 × 1–2 μm (
$\overline x $ $\overline x $ Culture characteristics: Conidia germinating on MEA within 24 h, reaching 20 mm after 10 d at room temperature, irregular, undulate, curled margin, yellow to pale brown on the surface and wrinkle and brown in reverse.
Material examined: Thailand, Chiang Rai Province, Doi Pui, on dead stems of Bidens pilosa, 14 March 2023, Zin Hnin Htet (BP-DP-10, MFLU 24-0030, new host record); living culture MFLUCC 24-0058.
Notes: Morphologically, our species, Chromolaenicola chiangraiensis (MFLUCC 24-0058) is similar to C. chiangraiensis (MFLUCC 17-1493) in having superficial, scattered, dark brown to black colonies, holoblastic, hyaline conidiogenous cells, oval to ellipsoidal, aseptate to 1-septate, reddish brown, verruculose conidia but differ in having shorter conidiogenous cells (2–4 vs 3.5–6.5 µm) (Table 3). A comparison of the tef1-α gene region of Chromolaenicola chiangraiensis (MFLUCC 24-0058) and C. chiangraiensis (MFLUCC 17-1493) reveals 13 base pair differences (1.48%) across 876 nucleotides. Therefore, our strain (MFLUCC 24-0058) is described as a new host record of Chromolaenicola chiangraiensis from Bidens pilosa (Asteraceae), which was previously recorded from Chromolaena odorata (Asteraceae) in Chiang Rai Province, Thailand[17].
Table 3. Synopsis of recorded asexual morph of Chromolaenicola species.
Species Conidiomata
(µm)Peridium
(µm)Conidiogenous cells
(µm)Conidia
(µm)Host/substrate Ref. C. ananasi (MFLU 23-0167) − − − 7–8 × 4–5 Ananas comosus (Bromeliaceae) [30] C. chiangraiensis
(MFLUCC 17-1493)− − 3.5–6.5 × 1–2 9–14 × 6–9 Chromolaena odorata (Asteraceae) [17] C. chaingraiensis
(MFLUCC 24-0058)− − 2–4 × 1–2 9–13 × 6–10 Bidens pilosa (Asteraceae) This study C. clematidis
(MFLUCC 17–2075)76–145 × 107–128 5–10 2.6–4.5 × 4–7 7–10 × 4.5–7 Clematis subumbellata (Ranunculaceae) [29] C. lampangensis
(MFLUCC 17-1462)150–230 × 170–270 10–20 – 12–15 × 4–6.5 Chromolaena odorata (Asteraceae) [17] C. siamensis
(= Cylindroaseptospora siamensis, MFLUCC 17–2527)110–165 × 140–190 15–38 6.5–7.4 × 3.2–4.7 7.2–9.4 × 5.4–6.5 Leucaena sp. (Fabaceae) [28] C. siamensis
(MFLUCC 24-0057)130–235 × 170–230 15–20 1–2.5 × 2–4 7–15 × 5–10 Bidens pilosa (Asteraceae) This study C. thailandensis
(MFLUCC 24-0056)100–150 × 110–150 13–20 1–2 × 3–4 5–11 × 4–10 Bidens pilosa (Asteraceae) This study Chromolaenicola siamensis (Jayasiri, E.B.G. Jones & K.D. Hyde) Mapook & K.D. Hyde Fungal Diversity 101, 1–175 (2020). (Fig. 3).
Index fungorum number: IF557283, Faces of fungi number: FOF 07787

Figure 3.
Chromolaenicola siamensis (MFLU 24-0029, new host record). (a), (b) Conidiomata on the substrate. (c) Section through conidiomata. (d) Peridium. (e), (f) Conidiogenous cells. (g)–(l) Conidia. (m) Germinating conidia. (n) Culture on MEA. Scale bars: (a), (b) = 500 µm, (c) = 50 µm, (d) = 20 µm, (e)–(m) = 5 µm, (n) = 10 mm.
Saprobic on dead stems of Bidens pilosa. Sexual morph: Undetermined. Asexual morph: Conidiomata 130–235 × 170–230 μm (
$\overline x $ $\overline x $ $\overline x $ Culture characteristics: Conidia germinating on MEA after 24 h, 22 mm after 10 d at room temperature, irregular, entire, curled margin, yellow to pale brown on the surface, wrinkled and brown in reverse.
Material examined: Thailand, Chiang Rai Province, Doi Pui, on dead stems of Bidens pilosa, 14 March 2023, Zin Hnin Htet (BP-DP-7, MFLU 24-0029, new host record); living culture MFLUCC 24-0057.
Notes: In our phylogenetic study, our strain (MFLUCC 24-0057) is sistered to Chromolaenicola siamensis (MFLUCC 17-1527) with 51% ML and 0.82 BYPP. When we compared the morphology, our strain (MFLUCC 24-0057) is similar to Chromolaenicola siamensis (MFLUCC 17-1527) in having pycnidial, solitary, immersed, globose to obpyriform, unilocular conidiomata, phialidic, hyaline conidiogenous cells, and hyaline to dark brown, globose to subglobose, aseptate to 1-septate conidia with similar size (7–15 × 5–10 μm vs 7.2–9.4 × 5.4–6.5 μm). Our strain (MFLUCC 24-0057) differs from C. siamensis (MFLUCC 17-1527) in having thinner peridium (15–20 µm vs 15–38 μm) and shorter conidiogenous cells (1–2.5 × 2–4 μm vs 6.5–7.4 × 3.2–4.7 μm) (Table 3). However, the comparison of base pair differences revealed no or insignificant results LSU 0% (0/851), ITS 0.2% (1/459), tef1-α 0.3% (2/740), rpb2 0.1% (1/914), which indicates that they are conspecific. Therefore, we herein report C. siamensis as a new host record from Bidens Pilosa (Asteraceae), while this species was previously recorded on the decaying pods of Leucaena sp. (Fabaceae) in Lampang Province, Thailand[28].
Chromolaenicola thailandensis Mapook & K.D. Hyde Fungal Diversity 101, 1–175 (2020). (Fig. 4).
Index fungorum number: IF557284, Faces of fungi number: FOF 07788
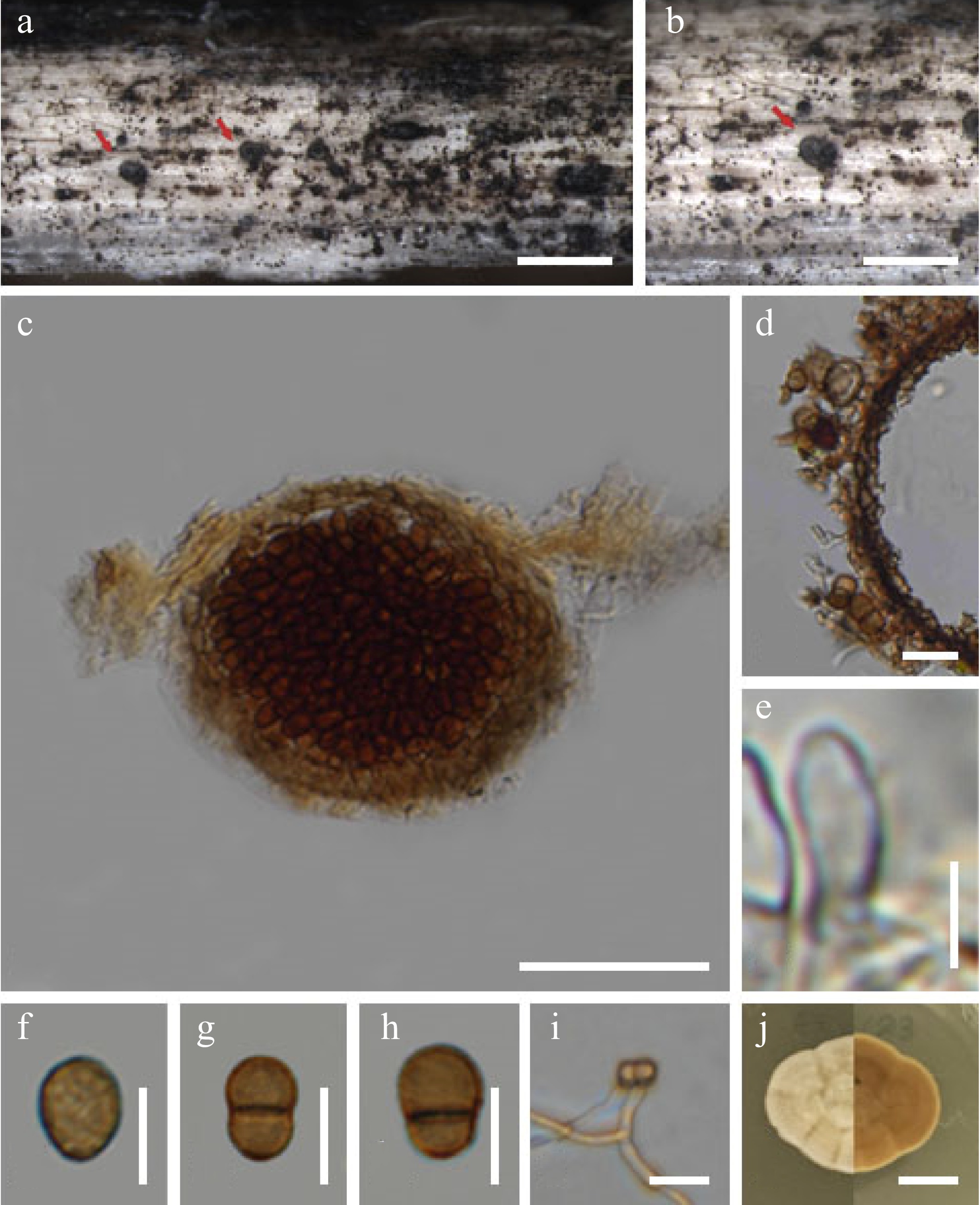
Figure 4.
Chromolaenicola thailandensis (MFLU 24-0028, new host record, first report of asexual morph). (a), (b) Colonies on substrate. (c) Section through conidiomata. (d) Peridium. (e) Conidiogenous cells. (f)–(h) Conidia. (i) Germinating conidia. (j) Culture on MEA. Scale bars: (a), (b) = 500 µm, (c) = 50 µm, (d)–(i) = 10 µm, (j) = 10 mm.
Saprobic on the dead stems of Bidens pilosa. Sexual morph: Undetermined. Asexual morph: Conidiomata 100–150 × 110–150 µm (
$\overline x $ $\overline x $ $\overline x $ Culture characteristics: Conidia germinating on MEA within 24 h, 21 mm after 10 d at room temperature, irregular, entire margin, smooth, wrinkled, pale yellow on the surface, curled and brown in reverse (Fig. 5).
Material examined: Thailand, Chiang Rai Province, Doi Pui, on dead stems of Bidens pilosa, 14 March 2023, Zin Hnin Htet (BP-DP-2, MFLU 24-0028, new asexual morph record); living culture MFLUCC 24-0056.
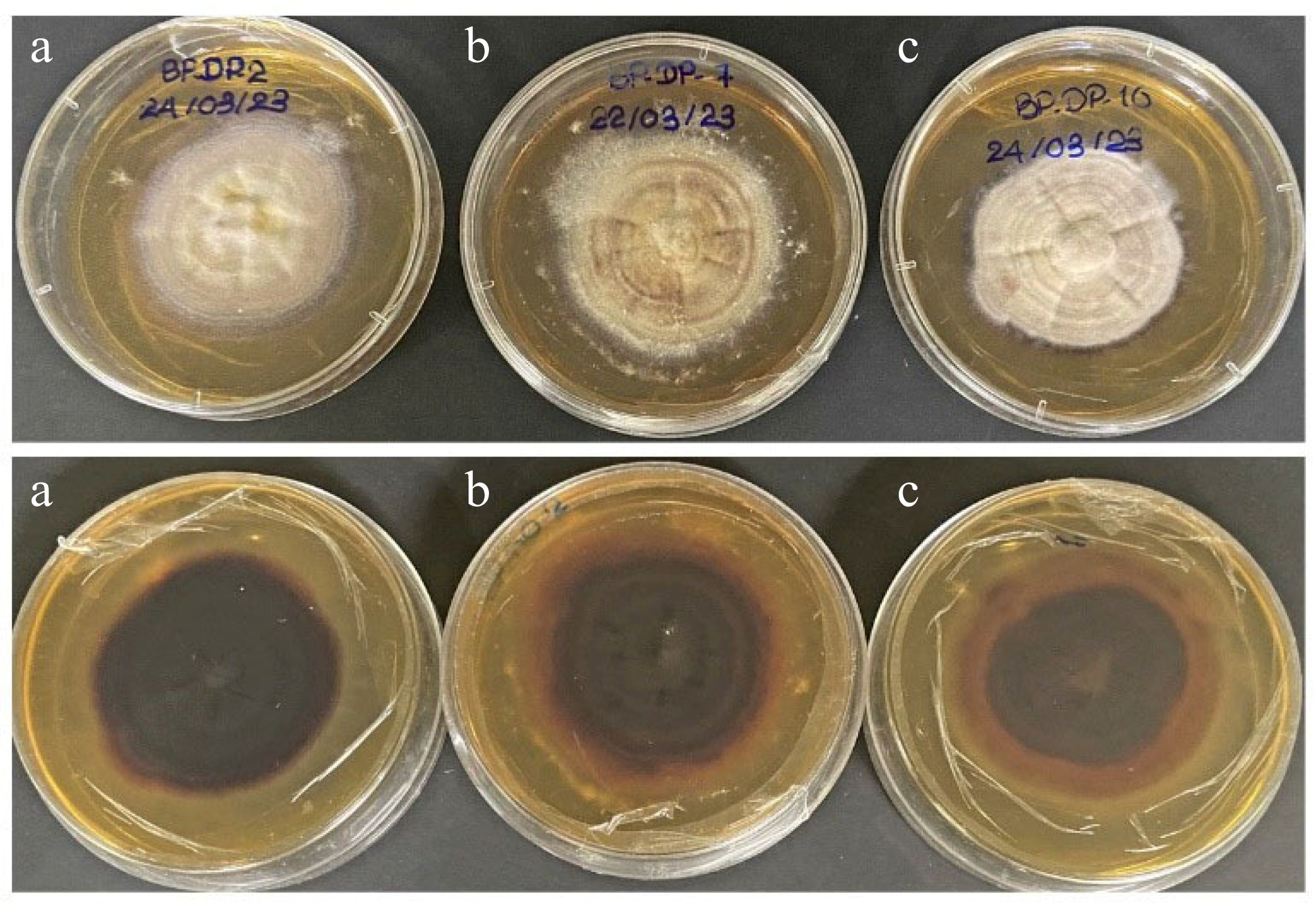
Figure 5.
Culture characteristics on MEA. (a) Chromolaenicola thailandensis (MFLUCC 24-0056), (b) Chromolaenicola siamensis (MFLUCC 24-0057), (c) Chromolaenicola chiangraiensis (MFLUCC 24-0058).
Notes: Chromolaenicola thailandensis (MFLUCC 17-1510, MFLUCC 17-1475) was found in its sexual morph in nature (Table 4)[17]. In the current study, we collected an asexual morph of C. thailandensis (MFLUCC 24-0056) on the dead stems of Bidens pilosa. However, we could not obtain its sexual morph in culture; hence failed to compare its morphology with C. thailandensis (MFLUCC 17-1510, MFLUCC 17-1475). Furthermore, there are no significant base pair differences in all five gene regions between our strain (MFLUCC 24-0056) and C. thailandensis (MFLUCC 17-1510). Therefore, we reported our strains as the new asexual morph of C. thailandensis and also the new host record from Bidens pilosa (Asteraceae) while previous strains of C. thailandensis was recorded on Chromolaena odorata (Asteraceae) in Chiang Rai Province, Thailand[17].
Table 4. Synopsis of recorded sexual morph of Chromolaenicola species.
Species Ascomata
(μm)Peridium
(μm)Asci
(μm)Ascospores
(μm)Host/substrate Ref. C. nanensis (MFLUCC 17-1473) 210–230 × 200–220 15–20 110–145 × 10–12.5 16–20 × 7.5–9 Chromolaena odorata (Asteraceae) [17] C. spindi (KUMCC 21-0564) 420–530 × 270–350 15–25 125–155 × 12–16 16–23 × 6.5–9.5 Sapindus rarak (Spindaceae) [27] C. thailandensis (MFLUCC 17-1510) 145–225 × 175–240 10–20 90–160 × 10–14 16–24 × 9–11 Chromolaena odorata (Asteraceae) [17] Preliminary screening for antibacterial activity
-
In this study, we observed the antibacterial activity of our newly isolated Chromolaenicola species against Bacillus subtilis, Escherichia coli, and Staphylococcus aureus. Among the three isolates, MFLUCC 24-0056, MFLUCC 24-0057, and MFLUCC 24-0058 showed antibacterial activity against Bacillus subtilis (17, 18, and 10 mm, respectively) and considered partial inhibition compared to the positive control. However, they did not inhibit the growth of E. coli and Staphylococcus aureus. Moreover, we also re-confirmed the preliminary antimicrobial activity result of Chromolaenicola species previously studied (Table 5).
Table 5. Preliminary antimicrobial activity result of Chromolaenicola species.
Species Zone of inhibition (mm); Ampicillin (+) Ref. Bacillus subtilis Escherichia coli Staphylococcus aureus Mucor plumbeus Chromolaenicola ananasi (MFLU 23-0167) N/A N/A N/A N/A [30] C. chaingraiensis (MFLUCC 24-0058) 10 mm inhibition no inhibition No inhibition N/A This study C. chiangraiensis (MFLUCC 17-1493) No inhibition No inhibition N/A no inhibition [17] C. clematidis (MFLUCC 17-2075) N/A N/A N/A N/A [29] C. lampangensis (MFLUCC 17-1462) No inhibition no inhibition N/A 14 mm inhibition [17] C. nanensis (MFLUCC 17-1473) No inhibition No inhibition N/A 12 mm inhibition [17] C. siamensis (MFLUCC 17-2527) N/A N/A N/A N/A [28] C. siamensis (MFLUCC 24-0057) 18 mm inhibition no inhibition no inhibition N/A This study C. spindii (KUMCC 21-0564) N/A N/A N/A N/A [27] C. thailandensis (MFLUCC 17-1510) No inhibition No inhibition N/A No inhibition [17] C. thailandensis (MFLUCC 24-0056) 17 mm inhibition No inhibition No inhibition N/A This study N/A: Not available; Positive control (+): Ampicillin. -
Chromolaenicola species are found as saprobes in terrestrial on different plant hosts[17,27−29]. In previous studies, seven Chromolaenicola species were reported from Thailand, and two from China[17,27−30]. We also provided detailed information on all Chromolaenicola species (Tables 3 & 4). Based on the morphological and phylogenetic analyses, three new host records and one new asexual morph report is described in this paper. Although the conidial color and conidiogenous cell sizes of the Chromolaenicola chiangraiensis strains MFLUCC 24-0058 and MFLUCC 17-1493 are different, the base pair difference is not significant, and the latter was reported on Chromolaena odorata (Asteraceae). Therefore, we described our strain as a new host record on Bidens pilosa (Asteraceae). Furthermore, Chromolaenicola siamensis (MFLUCC 17-1527) was recorded on Leucaena sp. (Fabaceae) and herein we reported our strain (MFLUCC 24-0057) as a new host record on Bidens pilosa (Asteraceae). This record also marks the first occurrence of C. siamensis on Asteraceae species. Moreover, we found the asexual morph of Chromolaenicola thailandensis (MFLUCC 24-0056) for the first time in our study, and this is also the new host record on Bidens pilosa (Asteraceae).
In this study, we provided the preliminary screening results of all Chromolaenicola species, including known previous studies. Three isolates of Chromolaenicola species from northern Thailand underwent preliminary screening for antibacterial activity, and all three isolates showed partial inhibition of the growth of Bacillus subtilis. (Table 5). Previously, four Chromolaenicola species, C. chaingraiensis (MFLUCC 17-1493), C. lampangensis (MFLUCC 17-1462), C. nanensis (MFLUCC 17-1473), and C. thailandensis (MFLUCC 17-1510) have been studied for their potential antimicrobial properties[17]. Among them, C. lampangensis (MFLUCC 17-1462), C. nanensis (MFLUCC 17-1473) can inhibit the growth of Mucor plumbeus. The rest of the species of this genus have not yet been explored for their potential biological properties[27−30]. In our study, the agar plug diffusion method was used to diffuse an antimicrobial drug into the agar, inhibiting bacterial growth, and then assessing the inhibition zone[17,57]. Several benefits of the agar plug diffusion method include its inexpensive cost and ability to examine a large number of fungal cultures quickly[57]. However, because it is impossible to determine the amount of antimicrobial agent diffused into the agar medium, this method is not appropriate for determining the minimum inhibitory concentration (MIC)[57]. Our study will contribute to the knowledge of the species diversity in Chromolaenicola and insight into their potential biological properties, which will be useful for further research.
-
The authors confirm contributions to the paper as follows: Fungal specimen collection and isolation, and manuscript writing: Htet ZH; fungal identification and contributed in the revision of the manuscript: Mapook A, Chethana KWT. All authors have read and agreed to the published version of the manuscript. All authors reviewed the results and approved the final version of the manuscript.
-
The data generated and analyzed during this study are available in the article. DNA sequence data are available in the GenBank database, and the accession numbers are provided in Table 1.
We would like to express our gratitude to the Center of Excellence in Fungal Research, as well as the Basic Research Fund support from the National Science, Research and Innovation Fund (Grant No. 652A01001), entitled 'Studies of fungi associated with Asteraceae and the discovery of biological properties'. Zin Hnin Htet would like to thank Mae Fah Luang University for giving her a scholarship to pursue a PhD and the re-inventing grant provided by the university for the molecular work 'Mae Fah Luang University Grant - Reinventing University System' (Grant No. 652A16049). Ausana Mapook thanks Mae Fah Luang University for funding 'Taxonomy, Phylogeny, Risk Assessment, and Potential Impact of Fungi on Siam Weed in Northern Thailand' (Grant No. 651A16029).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Htet ZH, Mapook A, Chethana KWT. 2024. Molecular taxonomy reveals new records of Chromolaenicola (Didymosphaeriaceae, Pleosporales) and potential antibacterial properties. Studies in Fungi 9: e006 doi: 10.48130/sif-0024-0006 

Molecular taxonomy reveals new records of Chromolaenicola (Didymosphaeriaceae, Pleosporales) and potential antibacterial properties
- Received: 18 March 2024
- Revised: 22 April 2024
- Accepted: 02 May 2024
- Published online: 22 May 2024
Abstract: In the present study, Chromolaenicola specimens were collected from the dead stems of Bidens pilosa in northern Thailand and compared with other Chromolaenicola species. Maximum-likelihood and Bayesian analyses were conducted on a combined LSU, SSU, ITS, tef-1, and rpb2 data set. Morphological and phylogenetic analyses revealed three species, Chromolaenicola chiangraiensis (new host record), C. siamensis (new host record), and C. thailandensis (new host and asexual morph record), respectively. A preliminary screening demonstrated the ability of Chromolaenicola chiangraiensis, C. thailandensis, and C. siamensis to partially inhibit the growth of gram-positive bacteria (Bacillus subtilis). Here, the detailed morphology, illustrations, and potential antibacterial properties of Chromolaenicola species are described. The present research will add to the body of knowledge on Chromolaenicola by revealing its possible antibacterial properties.
-
Key words:
- Antibacterial properties /
- Dothideomycetes /
- Multi-gene phylogeny /
- New host records