









-
Given that plants are inherently sessile[1], plants acquired mechanisms for sensing environmental changes in response to complex environments during plant evolution[2−4]. These mechanisms have significant implications for plant production. As the reproductive organ of plants, flowers are the prerequisite for fruit production and determine the production of seeds. The transition to reproductive growth at the right time is of great significance to plant reproduction[5,6]. Plant flowering time is influenced by various environmental factors[7]. Under adversity stress, plants will adjust the flowering time to achieve the best adaptive state, thereby obtaining maximum productivity[8]. Plants have complex regulatory mechanisms to determine whether to switch from vegetative growth to reproductive growth[9], and there are two outcomes in which flowering is either promoted or inhibited. In response to abiotic stresses, some plants choose to inhibit flowering leading to the growth stagnation and a slowdown in the metabolic rate. Others may directly speed up the flowering process through self-regulation, and quickly complete the life cycle to escape adversity[10,11], which usually depends on the intensity and type of abiotic stress and plant species[12]. Stress-induced flowering is beneficial to the preservation of species, and it has important biological significance like other pathways to induce flowering[13].
About 1,000 hectares of newly added saline-alkali land in the world are abandoned every year due to improper farming practices (statistics from the United States Department of Agriculture
https://fdc.nal.usda.gov/index.html )[14]. Salinity will cause up to 50% of land loss by the mid-21st century[15]. Salinity is one of the serious abiotic stress factors[16], which seriously hinders the growth and development of plants and reduces crop yields[17,18]. The intensity of influences of salt stress on plant growth and development is mostly determined by the amount of salts in the soil and therefore the effect of salt stress on plants also has a dose effect[19]. Phenotypes of salt-stressed plants including ion toxicity and difficulty in root water absorption often lead to cell metabolism disturbance[20,21], which can in turn affect enzyme activity and protein stability in plant cells. Plants slow down transpiration and protect photosynthetic apparatus via changing their own morphology and physiological responses[22,23]. Meanwhile, plants can also change biochemical reactions and accumulate more osmotic substances in cells to deal with the harm caused by high osmotic pressure[24].Salt stress can also disrupt the flowering timing and diminish the fertility of plants[25]. Across various plant species, the impacts of salt stress on flowering exhibit variations due to the intensity of the salt stress applied. Intense salt stress significantly postpones the flowering time of Arabidopsis[26]. Salt stress causes delayed flowering, hampered fruit ripening, and decreased fruit quality in pepper[27]. Similarly, in chickpea, salt stress has been linked to delayed flowering and reduced fertility[28]. Studies on cotton have revealed that salt stress delays flowering and results in reduced yield[29]. Conversely, in many annual ornamental plants, salt stress has been found to advance and shorten the flowering period[30]. However, Iris plants have exhibited delayed flowering under salt stress condition[31]. The impact of salt stress on rice has resulted in reduced fertility and even failure of seed setting[32]. These instances further highlight the comprehensive influences of salt stress on flower development, fertility, and the overall plant productivity which should be probed case by case.
Currently, It has been ascertained that pathways including vernalization, photoperiod, circadian clock, sugar, age, thermal perception, autonomic response, and gibberellin (GA) signaling pathways all contribute to the regulation of plant flowering[33,34]. In tomato, there are two flowering signal integrators, FA and SFT, corresponding to the LFY and FT genes in Arabidopsis, playing parallel roles in the regulation of tomato flowering[35]. Previous studies have shown that LEAFY can regulate the transformation of meristematic tissue into floral organs and integrate flowering signals to control the flowering time in Arabidopsis[36,37]. The tomato FA gene is a homolog of the FLORICAULA gene in snapdragon (Antirrhinum majus) and the LEAFY gene in Arabidopsis. The homologous FA gene in tomato plays a crucial role in integrating flowering signals, controlling the transformation of meristematic tissue into floral organs, and regulating the timing of flowering, making it an important flowering signal integrator[38]. Another flowering signal integrator is FLOWERING LOCUS T (FT). It is transported via vascular bundles to the shoot apical meristem (SAM) and promotes the transition of plants from vegetative to reproductive growth.[39,40] Homologous genes of FT have been identified in various species as integral components of the flowering regulatory pathway, responsible for orchestrating plant flowering. However, distinct functional variations have been observed among different plant species[41]. As a diurnal neutral plant, tomato flowering is not regulated by photoperiod[42]. Research shows that SINGLE-FLOWER TRUSS (SFT), the tomato homolog of FLOWERING LOCUS T (FT), encoded in tomato and tobacco, induces flowering in day-neutral plants[43]. Experiments on tomato SFT overexpression plants grafted with wild-type plants proved that SFT is a direct factor in the induction of shoot apical flower bud differentiation through long-distance transport of vascular bundles in tomato and tobacco[44]. Under nutrient stress, petunia induces flowering through the induction of PnFT2. These studies imply that FT and its homologous genes maybe involved in the process of plant flowering under abiotic stress[45].
Plant growth is a coordinated process of mitosis and cell division, which is known as the cell cycle progress[46]. In plants, cell cycle is divided into four phases: G1, S, G2, and M, regulated by different cyclin (CYC) proteins and cyclin-dependent kinases (CDKs) working in cooperation[47,48]. The G1/S and G2/M transitions are two crucial checkpoints in the cell cycle that ensure normal cell cycle, holding significant importance[49,50]. Salt stress can affect the transcription and protein activity of cyclins and CDKs, which in turn can regulate the cell cycle process, influence cell division in meristematic tissues and consequently impact organ morphology. In previous studies, it has been observed that salt stress can directly reduce the activity of CDKs and CycB1;2 promoter in Arabidopsis[51] . Under moderate salt stress conditions, the expression levels of regulatory genes for the G2/M checkpoint in Arabidopsis, such as CycB1;2 and CycB1;4, are significantly reduced[52].
Tomato (Solanum lycopersicum) is recognized as a popular vegetable crop with great cash values worldwide. Studies have shown that salt stress has negative effects on tomato growth and yield but sometimes it can also improve tomato quality[53]. Nonetheless, the way tomatoes adapt their transition from vegetative growth to reproductive growth under varying degrees of salt stress remains a mystery. Comprehending the implications of salt stress on the growth and development of tomato, strategically harnessing salt stress to manage tomato flowering, and enhancing the quality of tomato fruits hold crucial importance within the realm of tomato industry practices. Consequently, delving into the mechanisms governing tomato flower development under salt stress assumes paramount significance in advancing tomato production.
In this study, alterations in flower development in the salt-tolerant tomato cultivar 'Micro Tom' treated with varying levels of salt stress were investigated. The overall developmental progression of tomato flower organs was hindered by salt treatment. Relative to the control group, with the increase in the concentrations of NaCl solutions, tomato plants produced lesser amounts of flowers. We further analyzed plants treated with 150 mM NaCl and confirmed that salt stress significantly repressed flower sizes, which in turn led to the yield reduction. We also noticed that salt stress inhibited the transcriptions of SlCYCA2;1, SlCYCB1;2 and SlCYCB1;4, leading to slightly increased levels of endocycle in sepals and stamen. Furthermore, noteworthy shifts were observed in the flowering transition of tomatoes. Tomato seedlings subjected to NaCl solutions of 50, 100 and 150 mM respectively displayed distinct instances of accelerated flowering. Remarkably, the group treated with a NaCl concentration of 100 mM showed the most pronounced early flowering phenomenon in comparison to the control group. Simultaneously, through cluster analysis and principal component analysis of physiological data and flowering parameters of salt-treated tomato seedlings and RT-qPCR data of tomato leaves and data collected from the SlSFT silencing plants, we found that the mechanism driving the salt-induced early flowering in tomato was orchestrated through the SlSFT. In summary, distinct levels of salt stress reduced biomass, and influenced the flower development of tomato seedlings which could be related to the disturbed cell cycle. Tomato seedlings under low-intensity salt stress respond by upregulating the expression of SFT, thereby triggering an earlier flowering stage, and hastening the overall lifecycle progression.
-
Tomato cultivar 'Micro-Tom' seeds were obtained from Tomato Genetics Resource Center (
https://tgrc.ucdavis.edu/ ) and propagated in a growth chamber. Seeds were sterilized in 75% alcohol for 1 min and 20% NaClO solution for 15 min, washed with autoclaved water for three times, and planted on plain or full-strength Murashige and Skoog (MS) medium containing 0.7% agar. When the cotyledons of the tomato seedlings were fully expanded, the seedlings were transplanted to cylindrical pots with a diameter of 8 cm and cultured in a matrix of vermiculite, peat, and perlite at the ratio of 1:3:1. All seedlings were grown in a growth chamber at 22 °C with 8-h dark and 25 °C with 16-h light condition under the light intensity of 15,000 lux.Treatment with NaCl
-
A NaCl stock solution (4 M) was prepared by distilling 233.76 g of NaCl into 1 L of deionized water. The NaCl stock solution was diluted to concentrations of 50, 100, 150, and 200 mM with deionized water before use. At the three-leaf stage, for each experimental group, 100 mL or so NaCl solution at specific concentration was applied to each tomato seedling and the treatment was conducted three times over 5 d. For the control, approximately 100 mL ddH2O was applied to individual plants three times in 5 d. Each experimental group had six tomato seedlings as six biological replicates.
Gene express analysis
-
Total RNA from leaves was extracted by RNAiso Plus (TaKaRa, Otsu, Japan, cat. #9109) following the manufacturer's instruction. The first strand complementary DNA (cDNA) synthesis using 2 μg total RNA was performed following instructions provided with HiScript III RT Super Mix kit with gDNA wiper (Vazyme, cat. #R323–01) for RT-qPCR analysis and HiScript III 1st Strand cDNA Synthesis Kit with gDNA wiper (Vazyme, cat. #R711–01) for full-length gene cloning. The actin-encoding gene SlACTIN (Solyc03g078400) was used as a reference gene for reverse transcription-quantitative polymerase chain reaction (RT-qPCR) data normalization. Four microliters of cDNA templates were used for RT-qPCR analysis, respectively. All primers were designed by NCBI Primer-BLAST (
www.ncbi.nlm.nih.gov/tools/primer-blast ) and listed in Supplemental Table S1.Measurement of plant height and stem diameter
-
Tomato seedlings after treatments were photographed first and ImageJ software (
https://imagej.nih.gov/ij/ ) was then used to measure the plant height and stem diameter.Leaf photosynthesis measurement
-
The net photosynthetic rate of the fifth leaf of tomato was measured via Portable Photosynthesis System (CIRAS-3, PP-Systems) from 9:00 to 12:00 on a sunny day. The light intensity and CO2 concentration in the leaf chamber of CIRAS-3 were set up at 1,000 μmol·m−2·s−1 and 400 ppm, respectively. Six biological replicates were measured for each treatment.
Chlorophyll fluorescence determination
-
From 9:00 to 12:00 on a sunny day, a Pulse Modulated Fluorometer (FMS-2, Hansatech-instruments) was used to measure the effective photochemical quantum yield of PSII(ΦPSII) and the maximum photochemical quantum yield of PSII (Fv/Fm). We first measured the effective photochemical quantum yield of PSII using the PSII-800 mode. After the measurement was completed, the plants were reacted in the dark for 30 min and then the maximum photochemical quantum yield of PSII was measured using the Fv/Fm mode.
Chlorophyll measurements
-
A portable chlorophyll meter (SPAD–502, Konica Minolta Sensing) was used to measure leaf greenness of the tomato seedlings at three weeks after treatment. For each plant, measurements were taken in six locations on the fifth leaf, two on each side of the midrib on all fully expanded leaves and then averaged[54].
Nuclear DNA ploidy analyses
-
After a three-week treatment with a 150 mM NaCl solution, tomato floral organs were dissected into four tissues including sepals, petals, stamens, and pistils. Samples were subjected to Nuclear DNA ploidy analysis using flow cytometry[55]. The experimental steps are outlined as follows: a) Chop plant tissues thoroughly in lysis buffer with a razor blade to release the nuclei; b) Filter the lysate through a 48 μm mesh sieve; c) Add 50 mg·mL−1 propidium iodide (PI) staining solution to the filtrate before conducting flow cytometry analysis. Endocycle index (EI) was calculated to indicate the level of endocycle in various plant tissues.
Virus-induced gene silencing (VIGS) of SlSFT
-
VIGS experiments were conducted using the pTRV1 and pTRV2GW vectors with appropriate modifications[56]. Including the following steps: a) Analyze the gene CDS sequence using the VIGS tool on the Sol Genomics Network to match it with suitable VIGS sequences[57]; b) After transforming Ptrv1 and Ptrv2GW-genes into Agrobacterium GV3101, positive single clones were screened. The transformed Agrobacterium were cultured in LB liquid medium containing antibiotics. After centrifugation of the bacterial suspension, the pellet was resuspended in infiltration solution containing 100 Mm MES, 100 Mm MgCl2, and 100 Mm AS. The Ptrv1 and Ptrv2GW-gene cultures were mixed in a 1:1 ratio and incubated in the dark for 2 h; c) Inoculate the mixture of Agrobacterium cultures into tomato seedlings using a vacuum pump for vacuum infiltration. After 2-d dark incubation at 18 °C, transfer the seedlings to normal light conditions for cultivation. After 10 d, use qRT-PCR to detect gene expression. SlSFT silencing plants were treated with 150 mM NaCl, and analyzed for salt-stressed phenotypes.
Data analysis
-
All data were recorded in excel, and each set of data has at least six biological replicates. Data were analyzed using the least significant difference (LSD) with Data Processing System (DPS). The p value of statistical difference between the experimental groups was less than 0.05. All statistical results were performed in GraphPad Prism (
www.graphpad-prism.cn ).Clustering analysis and principal component analysis (PCA) were performed in R Studio Version 1.1.456 (
www.rstudio.com ) with R version 3.5.2 using gplots (www.cran.r-project.org/web/packages/gplots ) and factoextra (www.cran.r-project.org/web/packages/factoextra ) packages, respectively. -
To explore the effect of salt treatment on tomato flowering time, the days to the first flowering of tomato seedlings after NaCl solution treatment were counted and it was found that the flowering time of tomato plants was altered under salt stress (Fig. 1a & b). Due to the dose-dependent nature of salt stress, the impact of salt solutions of different concentrations on the flowering time of tomatoes varies. The current research indicates that treatment with NaCl solutions in the range of 50−150 mM resulted in a noticeable early flowering phenotype in tomatoes. However, once the NaCl solution concentration reaches 200 mM, there is a significant inhibition of flowering in tomato seedlings. The number of flowers (Fig. 1c), and the number of sprays (Fig. 1d) of each tomato seedling were then counted. Among them, the flowering parameters of the tomato seedlings treated with 50, 100, and 150 mM NaCl solutions were significantly different from those control group and 200 mM group. The imposition of high-intensity salt stress significantly disrupted the physiological processes within tomato seedlings, thereby severely impeding their overall growth and developmental progress. Furthermore, due to the influence of salt stress on various physiological activities of tomato seedlings, the accumulation of biomass in tomato plants was reduced. Consequently, this reduction in biomass leads to a decrease in the number of floral primordia and the number of branches in tomato plants (Fig. 1 c & d).
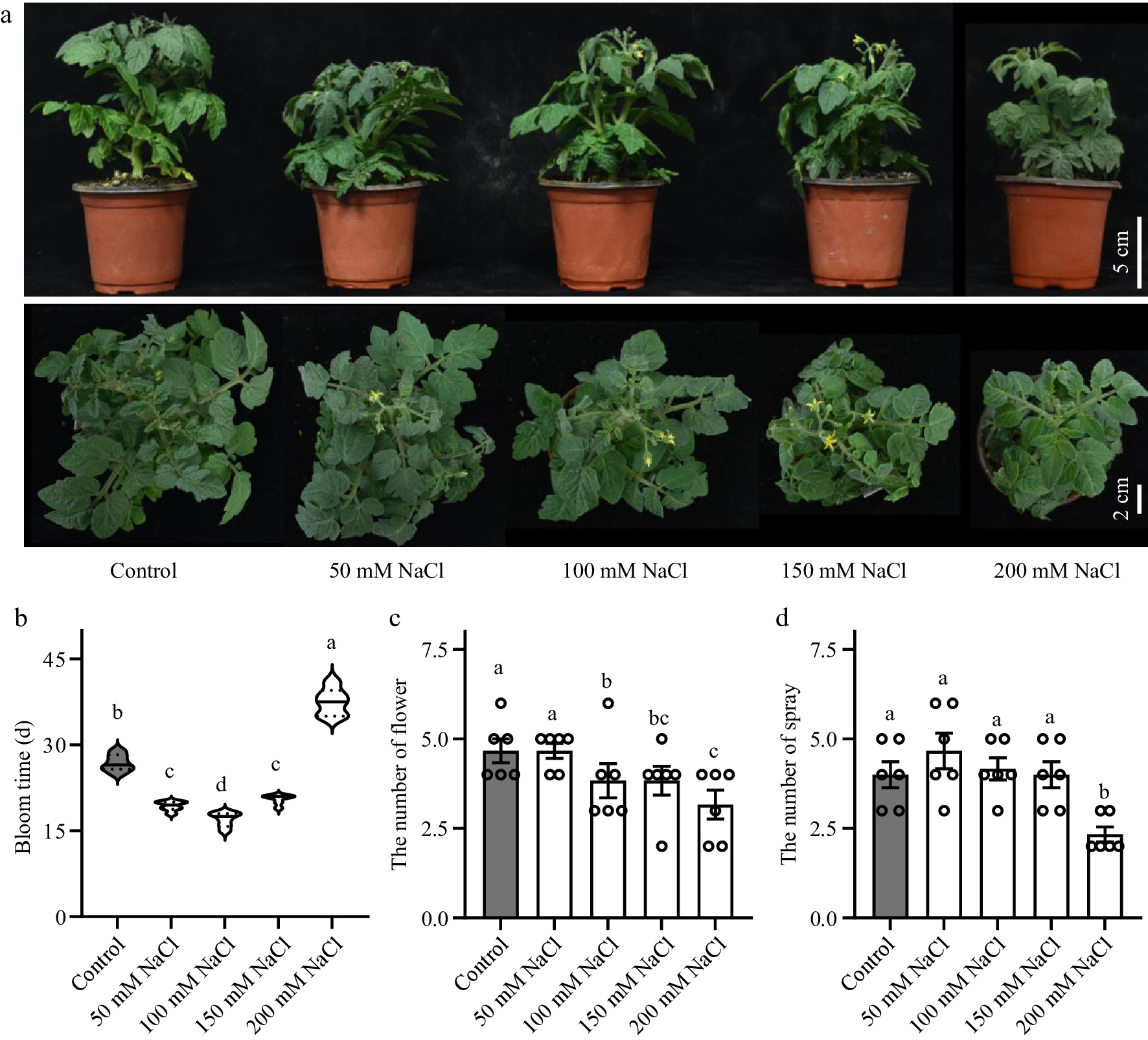
Figure 1.
Effects of salt stress on tomato flower development. (a) Phenotypes of tomato seedlings treated with NaCl solutions at 50, 100, 150 and 200 mM NaCl. (b) Flowering time of the tomato seedlings. (c) Number of flowers in tomato seedlings. (d) Number of spray in tomato seedlings. Tomato seedlings were treated with NaCl solutions for 3 weeks. Data were represented as average values of six biological replicates, with error bars indicating the standard error. Statistical analysis was performed using DPS software, and differences between treatments were considered statistically significant when denoted by distinct letters above the bars, as determined by the LSD Test (p < 0.05).
SFT is involved in the early flowering of tomato induced by salt stress
-
SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1(SOC1), LEAFY (LFY) and FLOWERING LOCUS T (FT) genes can integrate signals of various flowering pathways as a downstream signaling protein to promote flowering in Arabidopsis[36,39,40,58]. In tomato, there are only two flowering signal integrators, SlFA and SlSFT, which function in parallel in the integration of flowering signals. SlFA is a homolog of LEAFY, while SlSFT is a homolog of FT. In tomato, double mutants of slfa and slsft did not produce floral organs during their year-long growth[43].
It is speculated that the SlSFT gene or SlFA gene may be involved in the salt-stress-induced early flowering in tomato. To verify this hypothesis, we took the second true leaf (fully opened and mature) from 3-leaves-age and non-flowering tomato seedlings which were treated with NaCl solutions for 12 h, and used reverse transcription-quantitative polymerase chain reaction (RT-qPCR) to verify gene transcription level. The results showed that salt stress significantly changes the expression of SFT and FA (Fig. 2a, b). The changes in the expression levels of SlSFT were consistent with the flowering time of tomato seedlings treated with salt stress. The expression levels of SlFA were inhibited as the concentration of the NaCl solution increased, which did not correspond to the flowering phenotype. As AP1 is a direct target of SFT, the expression level of SlAP1 in the shoot apex was also measured (Fig. 2c). The expression of AP1 coincided with the expression of SFT. This further confirms that the SFT-AP1 pathway integrates the early flowering signal induced by salt stress, promoting early flowering in tomato plants.
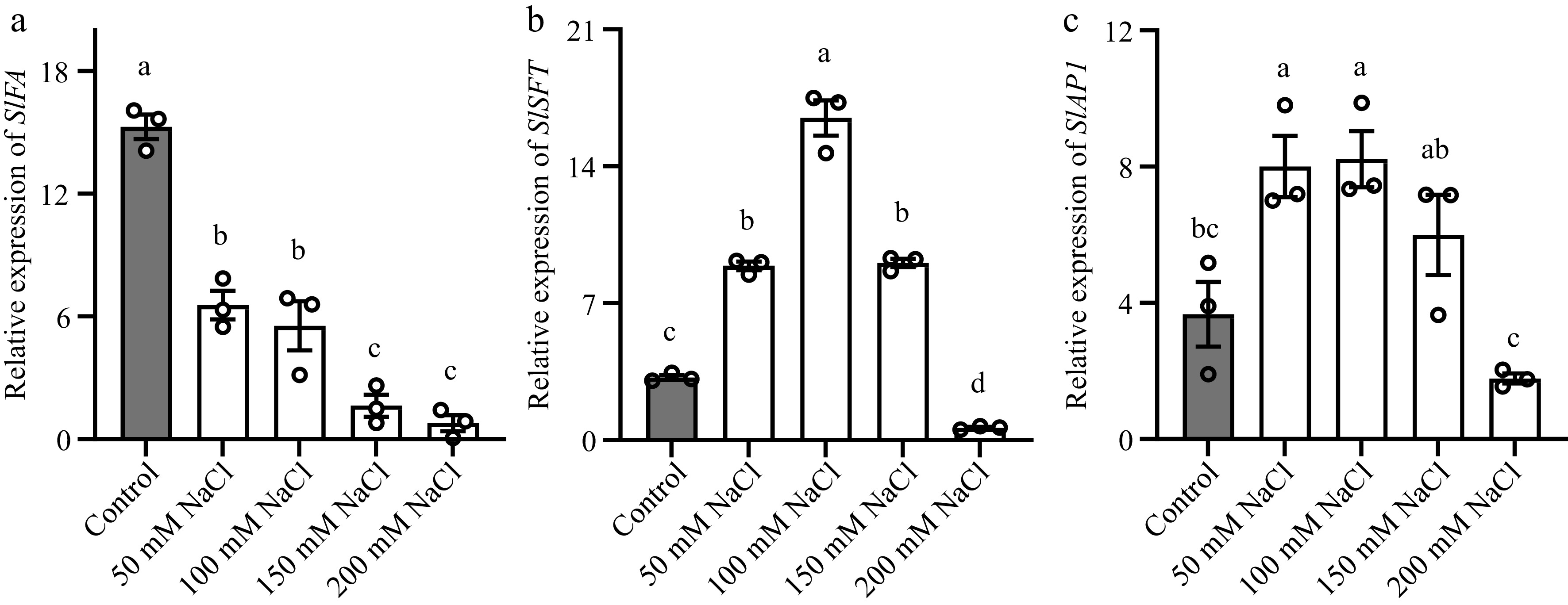
Figure 2.
Salt stress altered the transcription of SlSFT, SlFA and SlAP1 in tomato stem apex or leaves. (a), (b) Abundance of SlSFT and SlFA in the third leaf. (c) Abundance of SlAP1 in stem apex. The reference gene used for normalization was SlACTIN. Tomato seedlings were treated with NaCl solutions for 12 h. Data represents the average values from three biological replicates, with error bars indicating the standard error. Statistical analysis was performed using DPS software, and differences between treatments were considered statistically significant when denoted by distinct letters above the bars, as determined by the LSD Test (p < 0.05).
SlSFT silencing plants showed the late flowering phenotype
-
SlSFT silencing plants (TRV2-SlSFT) were generated via VIGS (Fig. 3a), along with empty vector control (TRV2), and validated the expression of SlSFT using qRT-PCR (Fig. 3b). Under normal growth conditions, TRV2-SlSFT exhibited a delayed flowering phenotype compared to TRV2. We then subjected TRV2-SlSFT plants to salt treatment and collected relevant flowering data. Through statistical analysis of flowering times, we found that the TRV2-SlSFT plants did not exhibit early flowering after salt treatment, whereas the TRV2 plants showed significant early flowering after salt treatment (Fig. 3c). This further indicates that low concentrations of salt stress induce early flowering in tomatoes by promoting the transcription of SFT.
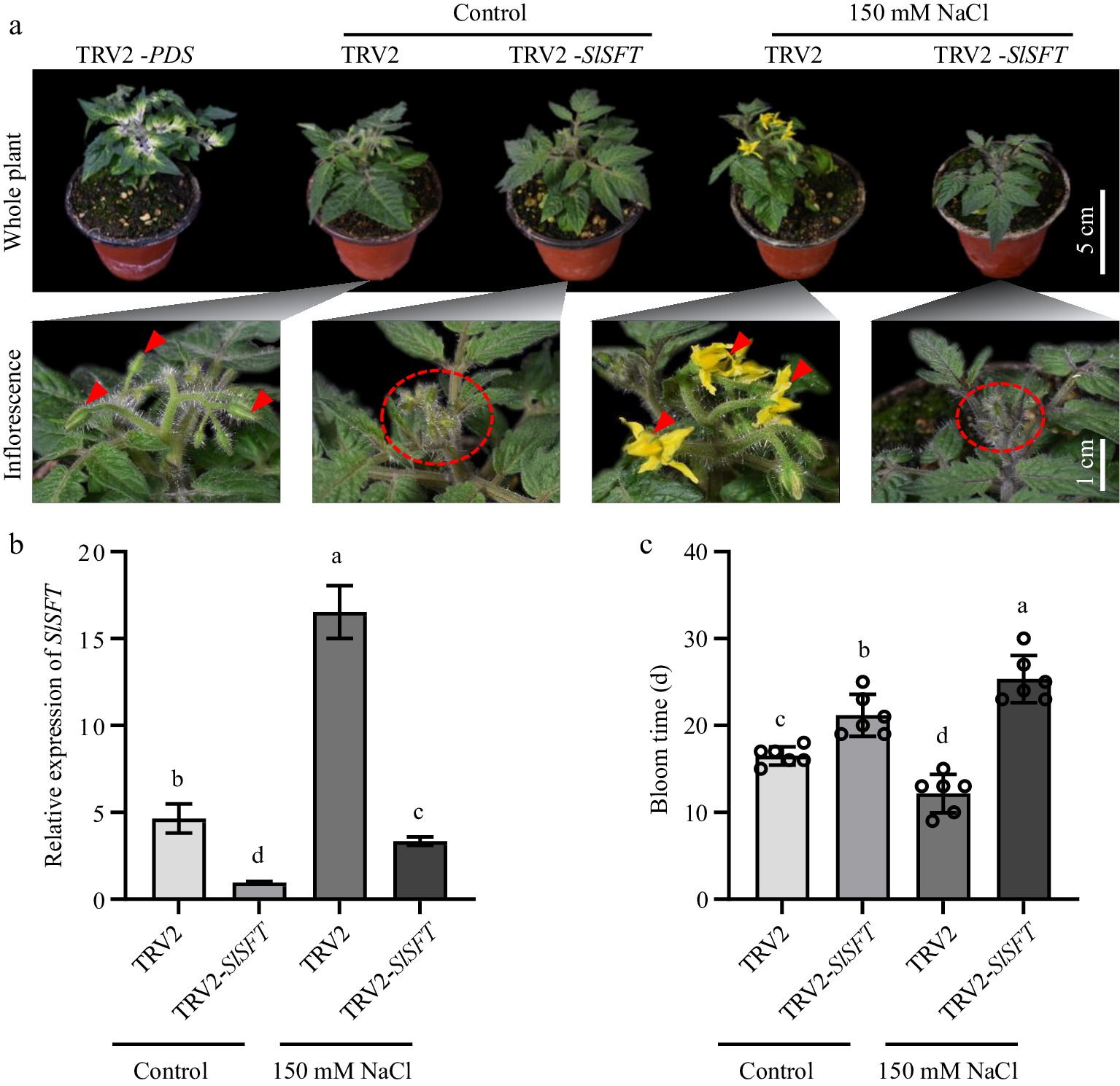
Figure 3.
SlSFT silencing plants did not exhibit early flowering under salt stress. (a) Flowering status of TRV2 and TRV2-SlSFT under salt treatment and control conditions. TRV2-PDS served as a positive control for the VIGS experiment. Red triangles and ovals indicate the inflorescences. (b) Expression levels of SlSFT in TRV2 and TRV2-SlSFT under salt stress and control conditions. (c) Bloom time of TRV2 and TRV2-SlSFT. The present data represents the average values from at least six biological replicates, with error bars indicating the standard error. Statistical analysis was performed using DPS software, and differences between treatments were considered statistically significant when denoted by distinct letters above the bars, as determined by the LSD Test (p < 0.05).
Salt stress affected the cell cycle progression of sepals and petals
-
The progress of the cell cycle in plants affects the division and expansion of cells, thereby affecting the size of plant organs[59]. Studies have shown that osmotic stress can reduce cell division rate and cell number in meristem[60]. In the present study, significant changes in the size of tomato floral organs were observed in 150 mM NaCl-treated plants relative to the control (Fig. 4a). Salt stress dramatically repressed the growth of flowers leading to shorter pistils and smaller sepals, petals, and stamens (Fig. 4b). Meanwhile, the fresh weight of floral organ also reduced in salt-stressed plants (Fig. 4c). Nuclear DNA ploidy distribution in pistils, sepals, petals, and stamens was conducted. Surprisingly, it was found that salt stress had a greater impact on tomato sepals and petals, with an increased content of 4C nuclei and a significantly higher value of endocycle index (EI), which indicated that salt stress might promote endoreduplication (Fig. 4d & e). On the other hand, salt stress had trivial effects on the nuclear DNA ploidy distribution of stamens and pistils, and their EI values did not change significantly. To be noted, the stamens of tomato flower did undergo an active endoreduplication under normal growth conditions indicated by higher relative abundances of endoreduplicated nuclei such as 8C, 16C, and 32C. However, after salt stress, the endoreduplication level in the stamen did not change, nor did the pistil. These data suggested that the cell cycle progression in various floral tissues responded differently to salt stress.
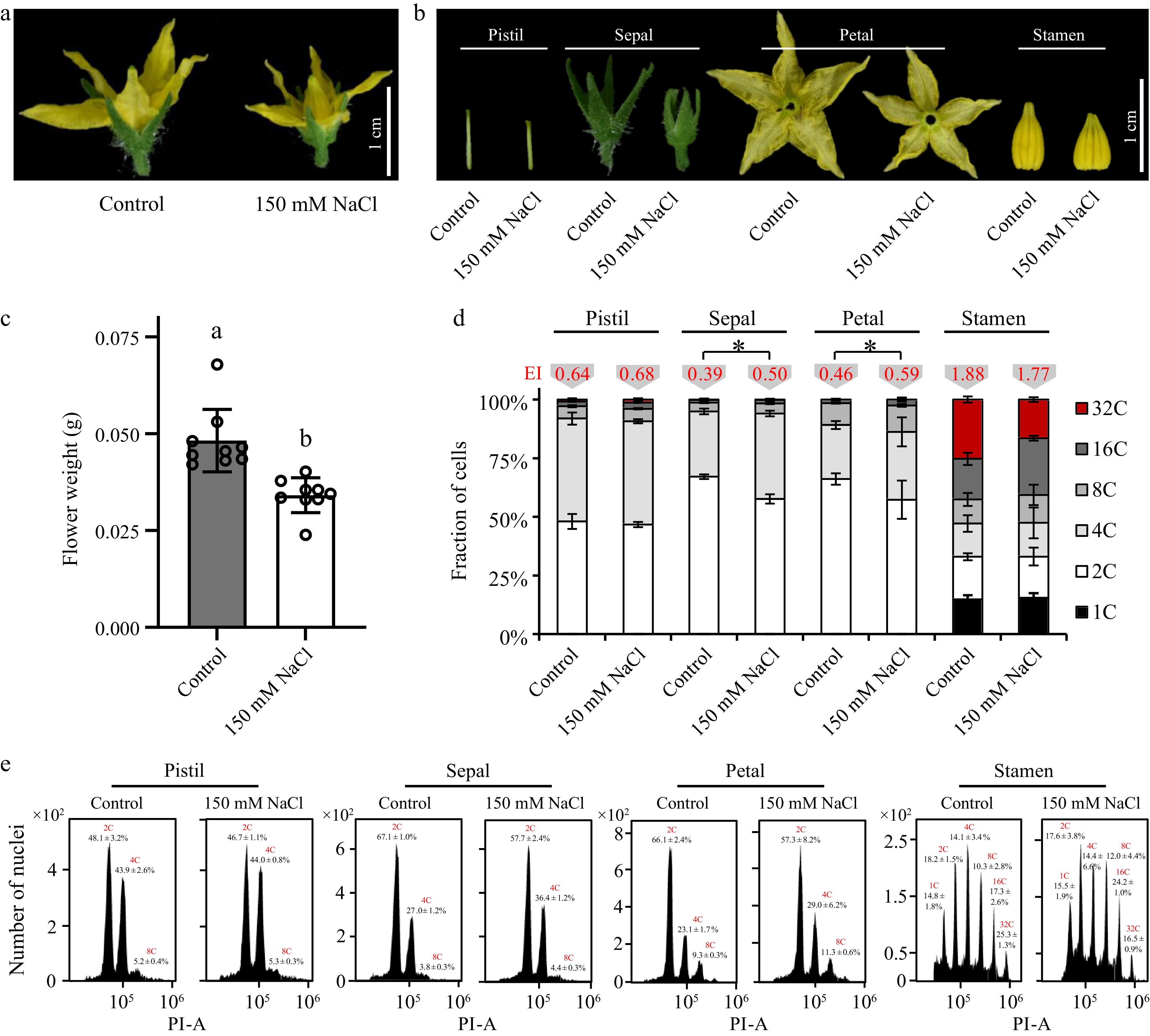
Figure 4.
Effects of salt stress on the cell cycle progress in various tomato floral tissues. Phenotypes of (a) the whole tomato floral organ and (b) various floral tissues treated with 150 mM NaCl. (c) Fresh weight of tomato floral organs treated with 150 mM NaCl. (d) Nuclear DNA ploidy distribution in sepals, petals, pistil and stamen of tomato flower. 4C, 8C, 16C, and stand for the endoreduplicated nuclei. Endocycle index (EI) values in red are located on the top of the columns. The present data represents the average values from at least three biological replicates, with error bars indicating the standard error. Statistical analysis was performed using DPS software, and differences between treatments were considered statistically significant when denoted by distinct letters above the bars, as determined by the LSD Test (p < 0.05). (e) Flow-cytometric analysis of endopolyploidy in various floral tissues.
Salt stress disrupts the regular floral cell cycle by interfering with the transcription of cyclins
-
Previous research indicates that cyclins and CDKs play crucial roles in adaptation to salt stress[60]. Under salt stress conditions, the promoter activity of CycA2;1, CycB1;1, CycB1;2, and CycB1;4 was reduced, leading to a decrease in their transcription levels, which in turn resulted in a deceleration of plant growth[51,61−63]. Correspondingly, cyclins play vital roles in re-configuring plant morphology under salt stress. Gene expression analyses were conducted to verify the relationship between the cell cycle alterations in petals and sepals and cyclin proteins (Fig. 5). Data indicated that the expression levels of SlCycA2;1 and SlCycB1;2 in the tomato floral organs were reduced to varying degrees (Fig. 5a & c). CycA and CycB are key proteins that influence the G2/M checkpoint, and salt stress hinders the transition at the G2/M checkpoint. This resulted in an increase in the 4C content in petals and sepals, leading to significant reductions in the size of petal and sepal tissues and substantial changes in the morphology of the floral organs.
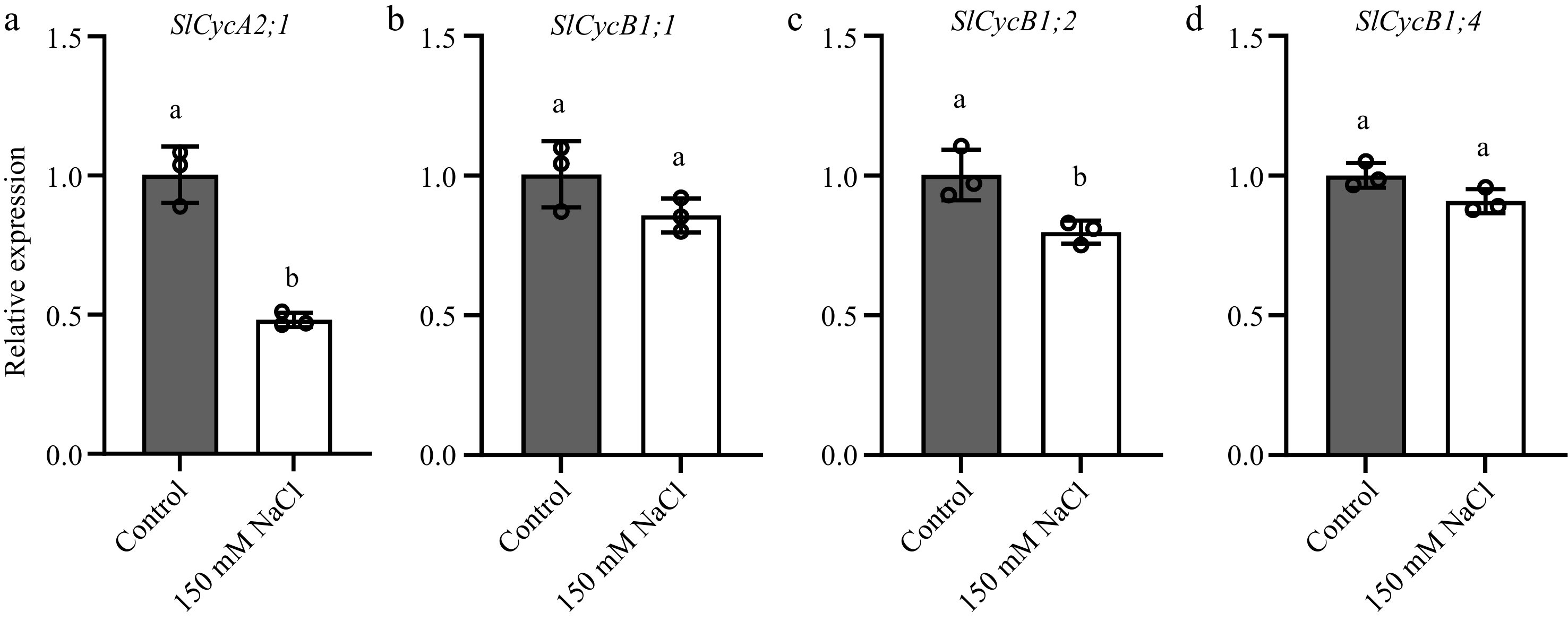
Figure 5.
Salt stress reduces the transcription of SlCycA2;1, SlCycB1;1, SlCycB1;2 and SlCycB1;4 in tomato floral organs. Transcript abundance of (a) SlCycA2;1, (b) SlCycB1;1, (c) SlCycB1;2 and (d) SlCycB1;4 in tomato floral organs. The present data represents the average values from three biological replicates, with error bars indicating the standard error. Statistical analysis was performed using DPS software, and differences between treatments were considered statistically significant when denoted by distinct letters above the bars, as determined by the LSD Test (p < 0.05).
Salt stress caused yield reduction
-
Whether the salt-stress induced early flowering could affect the development of tomato fruits was further investigated (Fig. 6). Salt stress significantly reduced the yield of tomato (Fig. 6a & b). Given that salt stress did not change the number of total fruits (Fig. 6c), the present results implied the reduction in yield was mainly caused by much smaller fruits (Fig. 6d−f). Moreover, the fact that salt stress did not influence the total fruit number of tomato (Fig. 6c) indicated that mild salt stress did not suppress the fruit set of tomato; rather, it only affected fruit development by influencing the accumulation of biomass.
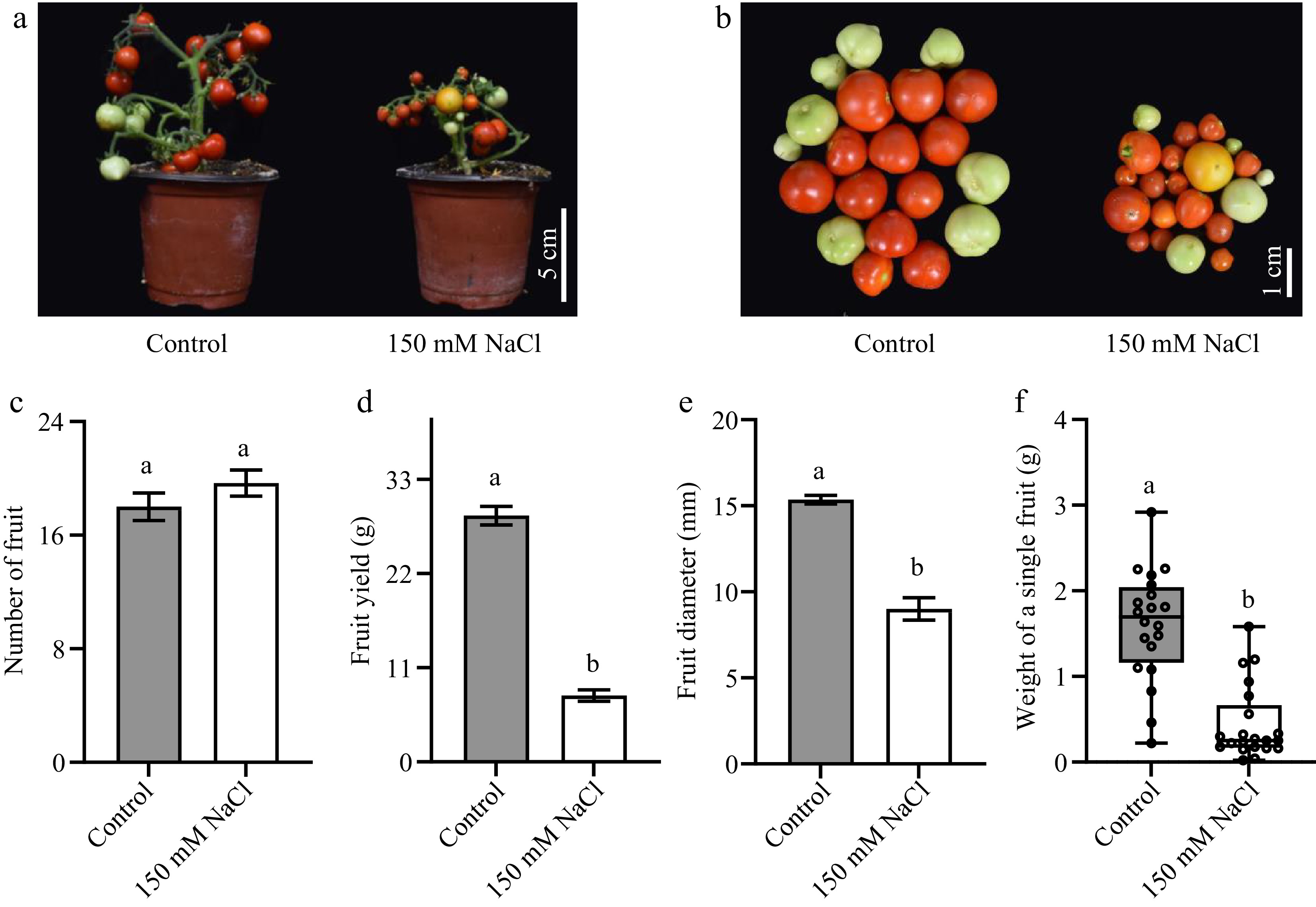
Figure 6.
The impacts of salt stress on tomato fruit yield. (a), (b) Photos of tomato fruits under normal growth conditions and salt stress. (c) The total fruit number of a single tomato seedling. (d) The total yield of a single tomato seedling. (e) The cross-sectional diameter of tomato fruits. (f) The weight of a single fruit. The present data represents the average values from at least six biological replicates, with error bars indicating the standard error. Statistical analysis was performed using DPS software, and differences between treatments were considered statistically significant when denoted by distinct letters above the bars, as determined by the LSD Test (p < 0.05).
To better understand the flowering time of tomato seedlings under salinity stress, clustering analysis and principal component analysis (PCA) were performed based on all the phenotypic data collected. Control and mild salt stress treatments were grouped together indicating that the physiological changes of tomato plants in responding to salt stress were dosage dependent (Fig. 7a). Moreover, according to PCA (Fig. 7b), the separation between the 200 mM NaCl treatment and other treatments was driven by bloom time (BT), indicating that salt stress could regulate the responses of plants to salt stress via affecting flowering time.
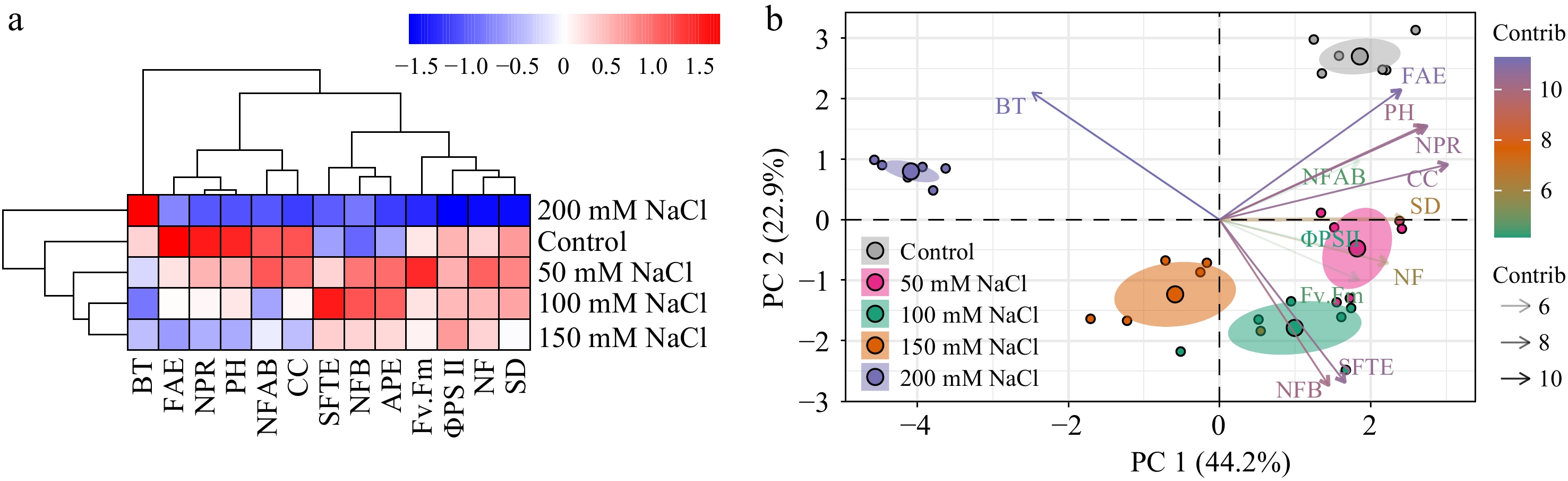
Figure 7.
Clustering analysis and PCA of physiological parameters in tomato seedlings under salt stress. (a) Clustering analysis and (b) PCA were employed to analyze the phenotypic data collected from tomato seedlings treated with NaCl solutions for 3 weeks. NF, Number of flowers; NFAB, The number of flowers on a branch; NFB, Number of flowering branches; CC, chlorophyll content; NPR, net photosynthetic rate; AWC, aboveground water content; PH, plant height; SD, stem diameter; BT, bloom time; ΦPSII, the effective photochemical quantum yield of PSII; Fv.Fm, the maximum photochemical quantum yield of PSII; FAE, relative expression of SlFA; SFTE, relative expression of SlSFT; APE, relative expression of SlAP1. Data analysis was carried out via R programming.
-
With the deterioration of the global environment, the increasing extreme climate, and the increasingly harsh soil conditions, plants will encounter various abiotic stresses in their life cycle. Due to the sessile nature of plants, plants can only cope with the adverse effects of abiotic stress by regulating physiological and biochemical reactions and metabolic regulation in vivo. Under abiotic stress, plants can avoid abiotic stress by regulating their transition from vegetative growth to reproductive growth, producing offspring as soon as possible, thereby shortening their life cycle. At the same time, when the intensity of stress surpasses the threshold that plants can tolerate, they will either experience stagnation or succumb to direct mortality. In previous studies, the regulation of reproductive growth transitions adopted by various plants in the face of abiotic stresses has been reported. Under nutrient deficiency, the vegetative growth of petunia is suppressed and the expression of PnFT2 is induced to promote flowering and produce fertile seeds[45]. Starvation stress promotes the early flowering of Lemna paucicostata[64]. Under drought stress, wheat avoids abiotic stress by flowering early through the mechanism of drought escape (DE)[65]. Under cold stress, Arabidopsis induces excessive flowering by promoting miR156[66]. Here, through the salt treatment of tomato seedlings, we found that low-intensity salt stress inhibited the growth and normal physiological and biochemical processes of tomato, and promoted the expression of SFT to make tomato flower early.
Under various levels of salt stress, the tomato's vegetative growth was hindered. Reproductive growth, being a pivotal aspect of the plant's life cycle, is also subject to salt stress. The transformation of plants to reproductive growth depends on the normal vegetative growth process. Salt stress affects the balance of hormones and transpiration making plants suffer from serious ion poisoning. All of these affect the normal vegetative growth process of plants, damage the photosynthetic mechanism and weaken photosynthesis, making it difficult for plants to produce and accumulate carbohydrates, and change the metabolic process of plants, which are all reflected in the process of plant growth and development.
It was noticed that the delayed flowering phenotype of SLSFT silencing plants was more severe under salt stress. Meanwhile, plants treated with 200 mM NaCl had much later flowering time and significantly repressed expressions of SlSFT and SlAP1. These data combined suggest that salt stress has a systematic impact on tomatoes. Under high concentrations of salt stress, the increased osmotic potential in the soil makes it difficult for plants to absorb water and nutrients, thereby affecting the overall physiological and metabolic activities which can lead to changes in gene expression. As mentioned above, plants only make the transition from vegetative to reproductive growth when there's sufficient energy. High concentration of salt stress inhibits normal physiological processes, preventing plants from accumulating enough energy. All of these disruptions combined with the altered expressions of flowering-related genes will ultimately change the plant's development as well as the flowering process.
To analyze the specific effects of salt stress on the development of each part of tomato flower organs, we dissected tomato flower organs and nuclear DNA ploidy analyses each part of tomato flower organs by flow cytometry. Under the treatment of tomato seedlings with 150 mM NaCl solution, the morphology of tomato flower was fully developed, but the size of flower organs changed significantly, and under higher concentrations of salt stress, the flower organs were deformed. To observe the effect of salt stress on the development of tomato floral organs from a microscopic point of view, we measured the cell cycle of each part of tomato floral organs by flow cytometry to reflect the cell division of floral organ tissues. From the analysis of the experimental results, we found that salt stress did not affect the normal cell division of tomato stamens and pistils, which indicates that tomato will preserve the normal DNA replication and cell division process of reproductive organs under salt stress to ensure that plants can produce fertile seeds. Unlike pistil and stamen, petals and sepals experienced a significant increase in the endoreduplication, interfering with normal cell division. This may directly affect the accumulation of floral organ biomass. However, the development of floral organs after treatment with high concentration NaCl solution was directly inhibited, and the normal morphology of floral organs could not be formed, which also indicated that the ability of tomato to tolerate salt stress to ensure normal floral organ development was limited.
The cell division and mitosis that make up the cell cycle in plant tissues play a crucial role in shaping the morphology of plants to adapt to their environment. Within the cell cycle, two key checkpoints, G1/S and G2/M, are of paramount importance, and cyclins and CDKs play a critical role in the proper progression of these checkpoints. In previous studies, it has been observed that the transcription of cyclins and CDKs is influenced by salt stress, which hinders the normal cell cycle progression. Based on the present analysis of the various parts of the floral organs under salt stress, a significant increase in content of 4C nuclei in the petals and sepals have been found compared to the control group. This indicates that salt stress affects the G2/M checkpoint in sepals and petals, a critical phase of the cell cycle. Salt stress slows down the cell division in these tissues while DNA replication continues, resulting in increased 4C content and EI values. However, the cell cycle progression in two sexual organs including pistils and stamens at this stage did not respond to salt stress. This indicates that under low-concentration salt stress, plants preserve normal cell division in reproductive organs to ensure the normal progression of sexual reproduction. In the investigation of CycA2;1, CycB1;1, CycB1;2, and CycB1;4 whose transcriptions are affected by salt stress, a reduction in promoter activity for CycA2;1 and CycB1;2 in the floral organs was observed. This further substantiates that salt stress impacts the normal cell cycle progression in the floral organs by affecting the transcription of CycA2;1 and CycB1;2, leading to significant alterations in the size of petal and sepal tissues.
The regulation of the SFT gene expression in tomatoes involves multiple pathways, and it is worth noting that tomato's flowering response is not significantly influenced by variations in light duration due to its diurnal nature. When tomato seedlings reach the stage of flowering maturity, salt stress prompts the increased expression of SFT, consequently resulting in the early onset of tomato flowering. However, this enhancement occurs exclusively under conditions of low-intensity salt stress. Interestingly, the expression of SFT is suppressed when confronted with higher levels of salt stress. Due to the complex salt stress-responsive mechanism in plants, we believe that there may be more complex molecular mechanisms that coordinate the regulation of the effects of salt stress on tomato flowering time.
As the expanse of saline-alkali land continues to grow, salt stress has emerged as a pivotal abiotic stress factor with global implications, impacting plant growth, development, and subsequently diminishing agricultural yields. The utilization of saline-alkali land has thus evolved into a pressing concern that necessitates thoughtful consideration and solutions. Mild salt stress impeded the growth and development of tomatoes, yet paradoxically expedited their flowering period and enhanced the fruit quality[53]. In this study, a molecular mechanism governing the shift in flowering timing in tomatoes when exposed to salt stress was unveiled. The current experiments demonstrated that under mild salt stress, tomato exhibited early flowering through the upregulation of SFT expression. This resulted in a shortened tomato life cycle while maintaining the unperturbed cell division of pistils and stamens, thereby ensuring successful seed production. Hence, here it is suggested that SFT, in conjunction with other molecular regulatory mechanisms, collectively contributes to the orchestration of tomato flowering time in response to salt stress. Therefore, manipulating the expression of SFT in tomatoes holds the potential to alter their flowering time under salt stress conditions. This has considerable implications for enhancing tomato production on saline-alkaline lands.
Salt stress affects the development of floral organs and the transition to flowering, both of which are crucial for plant yield. However, studies on tomato floral transition and flower development under salt stress conditions are limited. In the present research, a notable discovery has been made that salt stress influences tomato flowering transition by altering the transcription of SFT. Moreover, we found that mild salt stress accelerates the flowering process in tomatoes, a finding that has significant implications for agricultural production. Notably, the application of low levels of salt stress does not disrupt the cell cycle processes of both the pistil and stamen, ensuring the normality of sexual reproduction. Salt stress, however, results in a reduction in the biomass of tomato floral organs and significantly diminishes the size of sepals and petals through its impact on CycA2;1 and CycB1;2 transcription. This study contributes to our comprehension of the molecular mechanisms governing flowering transition and floral organ development in tomatoes under salt stress and holds substantial importance for tomato cultivation in saline-alkaline soil.
-
The authors confirm contribution to the paper as follows: experiments conception and design: Ma F, Bao Z, Sun F; performing the experiments: Sun F; data analysis: Sun F, Wang Y, Sun M, Liu G, Fang D, Ma F, Bao Z; writing the manuscript: Sun F, Ma F, Bao Z. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
We gratefully acknowledge the support of Taishan Scholar Foundation of Shandong Province (tsqntsqn201909073 to FM; tsqn201812034 to ZB) and Tai-an city improved seed project (2022NYLZ05 to FM). We thank College of Horticulture Science and Engineering at Shandong Agricultural University for supporting the Experimental Core Facilities where we performed phenotypic analysis and plant culture.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 The primer sequences used in this study.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Sun F, Wang Y, Liu G, Fang D, Sun M, et al. 2024. Salt stress induces SFT expression to promote early flowering and inhibits floral organ development by disturbing cell cycle in tomato. Vegetable Research 4: e019 doi: 10.48130/vegres-0024-0017 

Salt stress induces SFT expression to promote early flowering and inhibits floral organ development by disturbing cell cycle in tomato
- Received: 25 November 2023
- Revised: 02 April 2024
- Accepted: 30 April 2024
- Published online: 27 June 2024
Abstract: Excessive application of fertilizers and continuous cropping result in a large amount of salt residues remaining in the soil, leading to salt stress in the following crop cultivation. Salt stress jeopardizes the growth and development of plants and poses a great challenge to tomato production. Floral transition is a critical step in determining the crop yield. Here the impact of salt stress on tomato (Solanum lycopersicum) floral transition using various concentrations of NaCl solutions is investigated. Salt stress significantly reduced plant biomass compared to the control. We observed an early flowering phenotype and smaller flowers in tomato plants treated with 50−150 mM NaCl solution relative to the control. RT-qPCR analyses of genes involved in floral transition revealed that the SFT gene was upregulated under salt stress suggesting the activation of SFT (SINGLE-FLOWER TRUSS)-AP1(APETALA 1) pathway-mediated floral transition. Salt treatment led to a decrease in tomato seedlings biomass, particularly in floral organs. Meanwhile, plants treated with 150 Mm NaCl also produced much smaller fruits than the control leading to a significant decrease in yield. Nuclear DNA ploidy analysis of various floral tissues in tomato has revealed significant impacts of salt stress on the intranuclear replication within sepals and petals. We indicate that the cell cycle in floral organs is disrupted by salt stress, primarily through its influence on the transcription of CycA2;1 and CycB1;2. Our study elucidates that salt stress promotes early flowering by inducing SFT expression and retard tomato floral organ development via perturbation of the cell cycle.
-
Key words:
- Tomato /
- Flowering time /
- Cell cycle /
- Salt stress /
- Flower organ development