








-
The discovery and development of antibiotics are among the main accomplishments of the twentieth century. The prolonged, abusive use of antibiotics has fueled the emergence of drug-resistant pathogens, posing a grave threat to human health, with over 2 million challenging-to-treat infectious diseases annually, resulting in approximately 700,000 deaths[1,2]. By 2050, it is projected that this mortality rate will soar to 10 million, underscoring the urgent need for the discovery of new secondary metabolites with potent, wide-ranging antibacterial properties in response to the rise of drug-resistant bacteria[3].
Biotechnological approaches have been utilized in the production of antibiotics from natural product sources[4] such as metabolomic engineering of Streptomyces coelicolor as a generic host for combinatorial biosynthesis and mutasynthesis in an effort to produce novel analogs of metabolites[5]. Dereplication is an analytical methodology used for detecting known compounds and is employed during the first step of antibiotic screening. This method resolutes re-discovery problems, which makes use of mass spectrometry data and molecular network annotations based on public datasets such as the Global Natural Products Social (GNPS,
https://gnps.ucsd.edu , accessed on 10 July 2023)[6,7]. Other approaches include co-cultivation where two microorganisms are incubated in the same medium[8]. Diversification of secondary metabolites cannot be achieved by mono cultivation, hence, the interaction between another microorganism in the culture medium can awaken silenced biosynthetic pathways to produce novel analogs[9]. Exploring new habitats such as marine ecosystems[10], taxa inhabiting extreme conditions[11,12], difficult-to-reach habitats[13], and plant-associated endophytes and epiphytes[14] have been investigated.Among the widely used methods to induce bioactive compounds from fungi is the One Strain Many Compounds (OSMAC) strategy. This method was first developed by Zeeck in the early 2000s and is notable for its simplicity and practicality[15]. OSMAC entails altering cultivation parameters such as medium composition and physical parameters (pH and temperature) or adding biosynthetic precursors and enzyme inhibitors/inducers to activate silent biosynthetic gene clusters and expand metabolite patterns produced by endophytes[16]. One successful application of OSMAC in the induction of antimicrobial compounds in fungi are the production of the metabolites isopropylchaetominine, isoterrelumamide A, and 5′-epi-averufanin from the sponge-associated fungus Aspergillus carneus using three different culture media. Thus, it is an efficient approach in releasing nature's chemical diversity. Prior reports on the use of OSMAC illustrate coupling with mass spectrometry (MS) followed by dereplication for rapid identification of known secondary metabolites in crude extracts[17−19]. Cryptic metabolites from marine-derived microorganisms are reportedly observed through dereplication when cultivated in different culture media, where epigenetic modifiers and different chemical components[20] have been employed to awaken silent gene clusters.
With the increasing demand for antibiotic compounds that effectively kill rapidly evolving infectious pathogens, induction of antimicrobial compounds from fungi using OSMAC along with dereplication can provide an efficient approach to antibiotic drug discovery. In this paper, the induction of the antimicrobials fusaric acid and beauvericin from the endophytic fungus, Fusarium sp. (UST-UVG10) isolated from Uvaria valderammensis was investigated in three different media and resulted in differentiated antibacterial and antimycobacterial properties.
-
The fungal culture was previously isolated from healthy leaves of Uvaria valderramensis, collected in Valderrama, Antique, Panay Island, Philippines (11°00'05.60" N; 122°07'45.58" E) on April 2022. The plant parts were surface sterilized with 70% ethanol (1 min), 4% sodium hypochlorite (30 s), and sterile distilled water (SDW) (1 min) to remove any contaminants[21]. The tissue imprint method was performed to confirm the efficacy of the surface sterilization protocol[22]. The processed samples were then placed on a petri dish with potato dextrose agar (PDA) and incubated at 25 °C for a week until fungal colonies appeared. After isolation and purification, the endophytic fungi were grouped based on their colony appearance. After which, because of frequent occurrence and based on colony morphotype, the endophytic fungus assigned hitherto as UST-UVG10 was chosen for this study. The culture was duplicated and stored for long-term preservation in 50% aqueous glycerol solution at 4 °C.
Morphological characterization
-
Morphological characterizations were carried out for 14 d in the PDA plate, which involved observation of the fungal colony characteristics such as color, elevation, and margin, as well as some important microscopic features, including fungal spores' size and shape. These observations were made using the Euromex Nexius Zoom Stereo Microscope (S/N- EU 1980535) at magnifications of 1.5× to 10× and an Olympus CX23 binocular compound microscope. The fungal strain was assigned with accession number CMS903.23 by the UST-CMS.
Molecular analysis
-
The endophytic fungus UST-UVG10 was sent to Macrogen Inc., South Korea, for DNA extraction, purification, amplification, and gene sequencing. The amplification step was performed using the universal primers ITS1 (5'-TCCGTAGGTGAACCTGCGG-3') and ITS4 (5'-TCCTCCGCTTATTGATATGC-3'), as described in previous studies[23]. The obtained sequence was subjected to BLAST search analysis to determine the highest percentage identity. Parameters for BLAST search were modified to include sequences from type materials and exclude uncultured specimens. Gathered sequences were analyzed using MEGA v. 11 using the Jukes-Cantor model with Gamma distribution (G+1) model of nucleotide substitution with 1,000 rapid bootstrap replicates. The phylogenetic tree was viewed using FigTree 1.4.0 and edited by Microsoft Office PowerPoint 365. The sequences used for analyses with accession numbers are given in Table 1.
Table 1. Reference taxa used in this study with their strain number and GenBank accession numbers.
Taxa Strain no. GenBank accession numbers (ITS) Ref. Fusarium dlaminii NRRL 13164T NR 182425 [24] Fusarium dlaminii NRRL 13164 U34572 [25] Fusarium acutatum NRRL 13308 U34573 [25] Fusarium acutatum CBS 402.97 MH862652 [26] Fusarium acutatum CBS 739.97 MH862669 [26] Fusarium acutatum CBS 402.97T NR_111142 [27] Fusarium phyllophilum NRRL 13617T NR_182426 [25] Fusarium phyllophilum NRRL 13617 U34574 [25] Fusarium hostae NRRL 29888T NR 171109 [28] Fusarium fujikuroi CBS 221.76 AB725607 [29] Fusarium fujikuroi CBS 221.76T NR_111889 [27] Fusarium fujikuroi CBS 221.76 MW827608 [30] Fusarium hechiense LC13644T NR_182846 [31] Fusarium hechiense LC13644 MW016454 [31] Fusarium elaeagni LC13627T NR_182845 [31] Fusarium elaeagni LC13627 MW016426 [31] Fusarium sp. NRRL28852 AF158304 [32] Fusarium sp. NRRL26793 AF158312 [32] Fusarium nygamai CBS 749.97 MH862671 [26] Fusarium nygamai NRRL 13448T NR 130698 [25] Fusarium nygamai CBS 749.97 MT740750 [33]* Fusarium aquaticum LC7502T NR 182844 [31] Fusarium aquaticum LC7502 MW016408 [31] Fusarium panlongense LC13656T NR 182847 [31] Fusarium panlongense LC13656 MW016470 [31] T Type specimens. * The species Fusarium nygamai (CBS 749.97) with the accession number - MT740750, was not yet published. www.ncbi.nlm.nih.gov/nuccore/2100342893 .Small scale fermentation and extraction
-
The pure fungal strain was inoculated in a 250 mL Erlenmeyer flask containing either 100 mL of Q6 ½ medium (D-glucose 0.25%, glycerine 1%, cottonseed flour 0.5%, demineralized water, pH 7.2), YM 6.3 medium (yeast extract 0.4%, malt extract 1%, glucose 0.4%, demineralized water, pH 6.3), or ZM ½ (molasses 0.5%, oat flour 0.5%, sucrose 0.4%, mannitol 0.4%, glucose 0.15%, calcium carbonate 0.15%, edamine 0.05%, ammonium sulfate 0.05%, demineralized water, pH 7.2). For this purpose, one-week-old cultures of Fusarium sp. (UST-UVG10) isolated on PDA were cut into small block pieces and transferred to Erlenmeyer flasks containing each medium. Cultivation was performed at room temperature, under static conditions, and in ambient light. Cultures on liquid media were incubated for 2−3 weeks, depending on the fungal growth. The glucose and pH levels were monitored daily using a glucose strip test and pH strip test, respectively. Fermentation was stopped when the glucose was depleted, and the pH value reached at 7.
Gravity filtration was used to separate the mycelia and supernatant of each culture flask. The mycelia were then soaked in acetone under ultrasonic conditions. The combined organic extracts were evaporated to dryness using a rotary evaporator (40 °C), distilled water was added to the crude product, and extracted again with the same volume of ethyl acetate (EtOAc). The aqueous layer was discarded, while the organic layer was treated with anhydrous sodium sulfate (Na2SO4) and evaporated using a rotary evaporator to obtain the crude EtOAc extract from the mycelia. In contrast, the supernatant (or broth) was treated three times with equal volumes of EtOAc. The combined EtOAc extracts from the broth were treated with anhydrous Na2SO4 and evaporated using a rotary evaporator to obtain crude EtOAc extracts. There were six crude EtOAc extracts from the three liquid media for this fungal strain.
Antimicrobial activity screening
-
Kirby-Bauer method was followed for disk diffusion assay. In vitro antimicrobial activity was screened using MHA (Mueller Hinton Agar). The plates were prepared by pouring 15 mL of molten media into sterile petri plates. The plates were allowed to solidify for 5 min. After which 0.1% v/v Staphylococcus aureus suspension was swabbed and allowed to dry for 5 min. The discs loaded with 1 mg/mL of fungal extract were placed on the surface of the medium, and the plates were incubated at 37 °C for 24 h. Sterile distilled water was used as a negative control, and methanol was used as a solvent control. Streptomycin was used as a positive control. At the end of incubation, inhibition zones formed around the discs were measured with a vernier caliper. These were performed in triplicates.
Minimum inhibitory concentration (MIC) determination
-
Minimum inhibitory concentration (MIC) as previously described with slight modification[34]. Plates were prepared aseptic, and the sterile 96-well plate was labeled. A volume of 100 μL of 0.1mg/mL of test material in 10% (v/v) DMSO was pipetted into the first row of the plate. All other wells contained 50 μL of nutrient broth. Two-fold serial dilutions were performed using a micropipette, and tips were discarded after use so that each well had 50 μL of the test material in serially descending concentrations. Next, 10 μL of resazurin indicator solution was added to each well. Finally, 10 μL of Staphylococcus aureus suspension (1.5 × 108 cfu/mL) was added to each well. A total of eight test concentrations were used which ranged from 1.3 to 166 ppm. Each plate was wrapped loosely in cling wrap to prevent bacteria from becoming dehydrated. Further, the plates had a set of controls, including a column with streptomycin as a positive control, a column with all solutions except the test compound, and a column with all solutions except the bacterial solution, which was replaced with 10 μL of nutrient broth. The plates were prepared in triplicate and placed in an incubator at 37 °C for 18−24 h. The color change was then evaluated visually. Any color changes from purple to pink or colorless were recorded as inhibited growth. The MIC value was determined as the lowest concentration at which color change occurred. Finally, MIC for the test material and the bacterial strain was calculated as the average of three values.
Microplate Alamar Blue Assay (MABA)
-
The antitubercular activity of the most potent compound against Mycobacterium tuberculosis H37Rv (American Type Culture Collection, Rockville, MD, USA) was evaluated using Microplate Alamar Blue Assay (MABA) and Low-Oxygen Recovery Assay (LORA). The test concentrations used ranged from 0.5 to 64 µg/mL. MIC90 was determined as the lowest concentration inhibiting fluorescence by 90% compared to the untreated bacterial control. As positive controls, the standard antituberculosis drugs isoniazid, streptomycin, and rifampin were used.
Metabolomic profiling of fungal crude extracts
-
Samples were prepared in methanol (100 μg/mL) and analyzed in triplicate using a Thermo Fisher Q ExactiveTM Plus mass spectrometer (Thermo Fisher Scientific, Waltham, MA) outfitted with an electrospray ionization (ESI) source and a Waters Acquity ultra-performance liquid chromatography (Waters Corporation, Milford, MA). Each sample was injected and eluted in a 3 L volume using a reversed-phase column (BEH C18, 1.7 m, 2.1 mm × 50 mm, Waters Corporation) using a binary solvent system consisting of water with 0.1% formic acid (solvent A) and acetonitrile with 0.1% formic acid (solvent B). 10-minute gradient elution began with 10% solvent B for 0.5 min, climbed to 100% B for 8 min, and then returned to starting conditions in the final 1.5 min. Analysis was carried out throughout full scan acquisition, gathering profile data while alternating between positive and negative polarity. The scan range was 120 to 1,500 m/z, and the scan time was 200 ms. The AGC target was set at 1,106, with a capillary voltage and temperature of −0.7 V and 310 °C, respectively; the S-lens RF level was set to 80.00, the spray voltage was 3.7 kV, and the sheath and auxiliary gas flows were 50.15 and 15.16, respectively.
MSConvert tool of MZMine was used to convert RAW data to mzXML format. The dataset was uploaded on GNPS and SIRIUS, which enabled the comparison of sample data to public spectral libraries of reference natural product compounds. Parameters for library matching include precursor ion mass tolerance of 0.01 Da, fragment ion mass tolerance of 0.05 Da, and minimum similarity (cosine) score of 0.70.
The parameters for SIRIUS tools were set as follows: molecular formula candidates retained; molecular formula database, (Bio Database, COCONUT, GNPS & Natural Product); maximum precursor ion m/z computer (10); profile, orbitrap; m/z maximum deviation; ions annotated with MZmine were prioritized and other ions considered (that is [M]+, [M+H]+, [M+K]+, [M+Na]+, [M+H-H2O]+, [M+NH4]+); ZODIAC was not utilized for data mining.
-
Morphological observation revealed isolate UST-UVG10 grown on PDA for 14 days appeared off-white, powdery, semi-submerge scarce to moderate mycelia, entire filamentous margin; reverse pale yellow (Fig. 1). The colony reached a diameter between 40−90 mm in diameter. Macroconidia subhyaline, falcate, (24−46 µm × 4−6 µm) (
$\overline {\rm x} $ $\overline {\rm x} $ 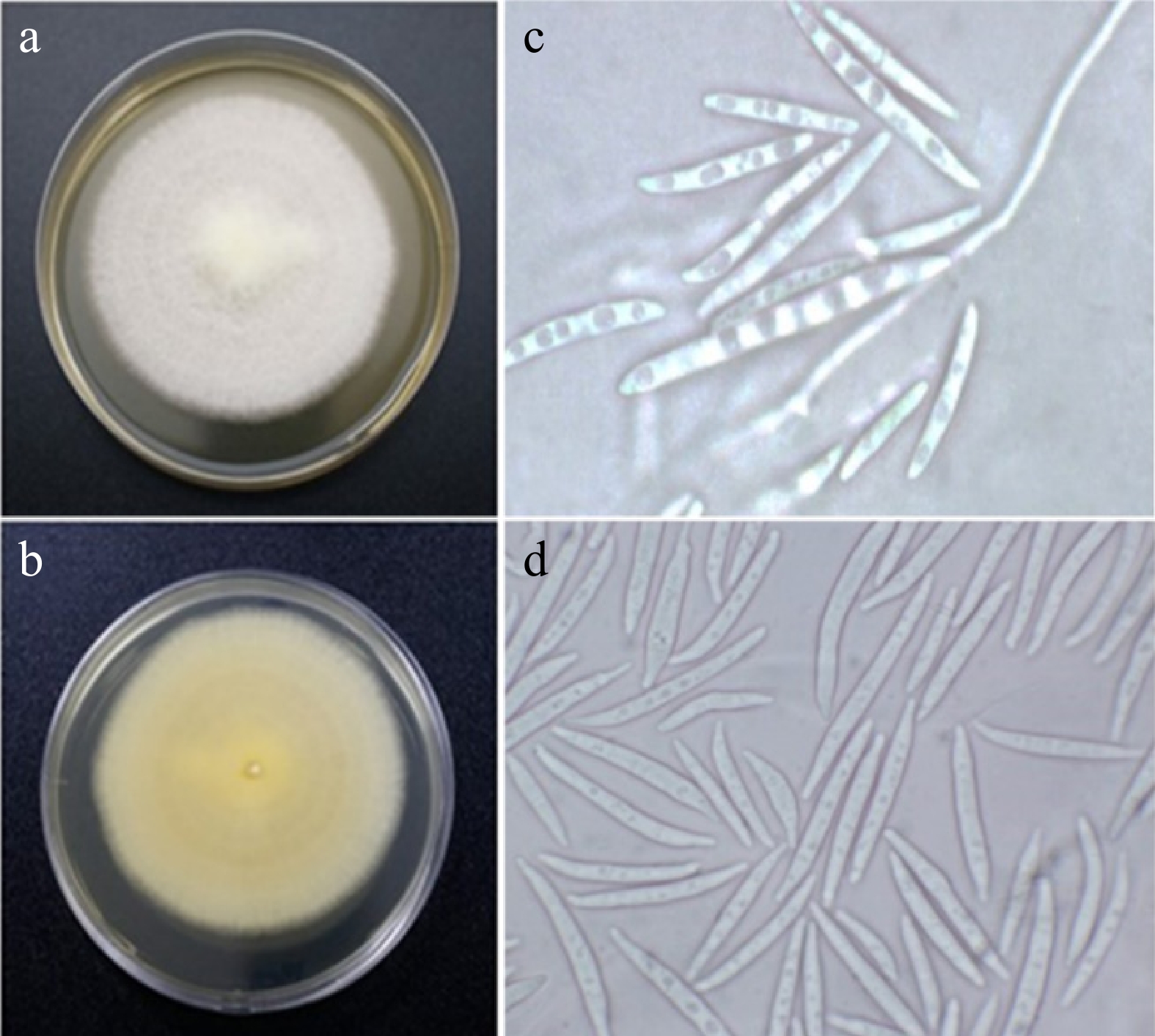
Figure 1.
Fusarium sp. (UST-UVG10) isolated from the leaves of U. valderramensis. (a), (b) Observed and reverse view of the colony on PDA plates after 14 d of incubation. (c), (d) Micrographs; macro- and microconidia.
For the phylogenetic tree, NCBI blast parameters included sequences from type material and excluded the uncultured samples. Based on our results, it yielded a range of Fusarium species with sequences in this study showing 100%−97% identity and 0.0 e value compared to (UST-UVG10). Evolutionary analyses were conducted in MEGA11. The tree was inferred using ML through the Jukes-Cantor model with Gamma distribution (five categories (+G, parameter = 0.1000)) (Fig. 2). The tree with the highest log likelihood (−1839.24) is shown. The analysis includes 26 nucleotide sequences with a total of 1925 positions in the final dataset. The phylogenetic analysis of ITS sequences identified the isolate to genus level, within the Fusarium complex, due to species complexity. Based on the tree, it formed a clade with F. phyllophilum, with a minimal branch length. Therefore, UST-UVG10 was given the name Fusarium sp. Additional gene markers for full validation are recommended.
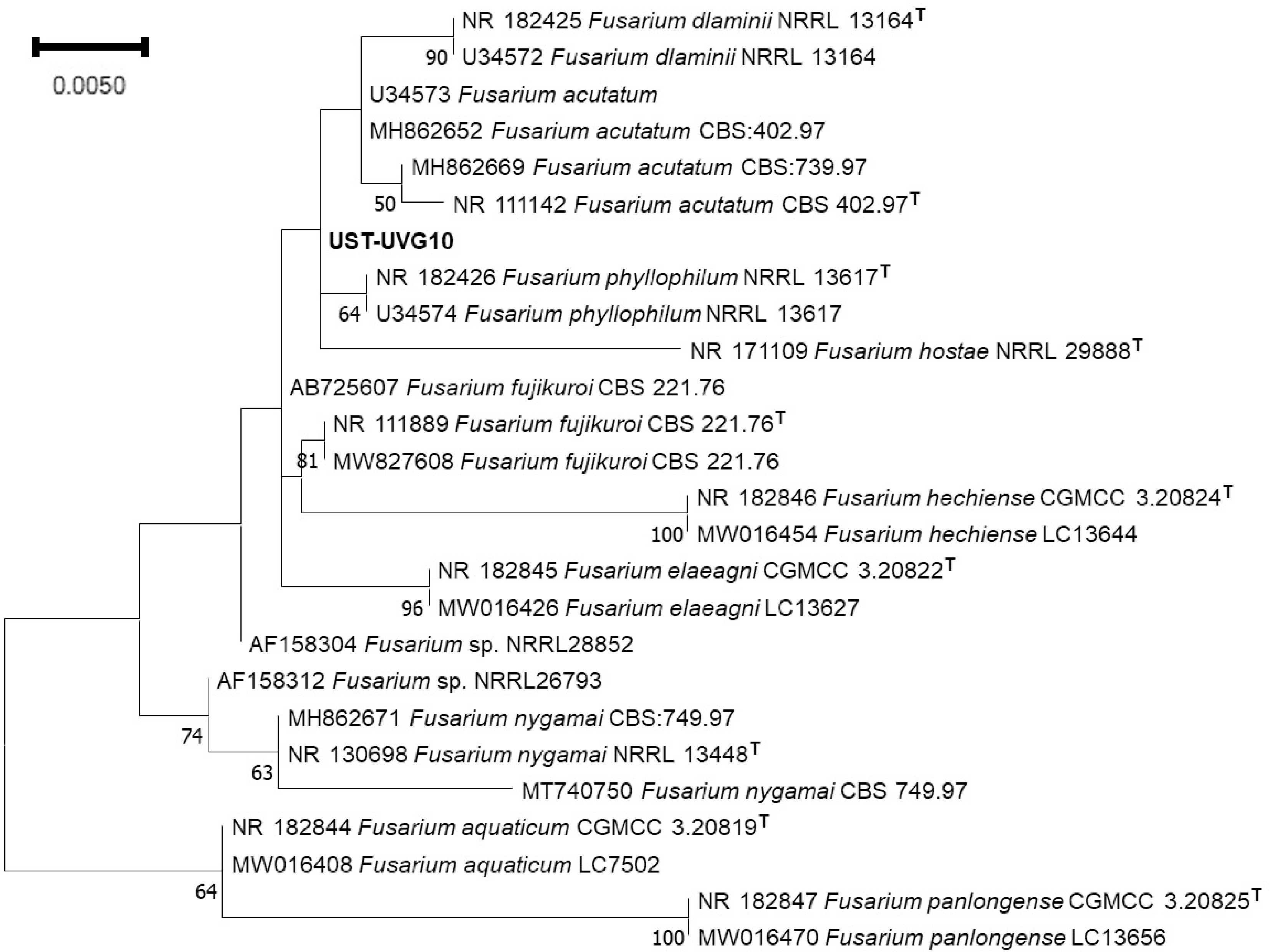
Figure 2.
The phylogenetic tree of isolated UST-UVG10, based on ITS region amplification, reveals genus-level identification due to its species complexity. Numbers in the node indicate the bootstrap values.
Antimicrobial activity screening
-
Fusarium sp. (UST-UVG10) was subjected to small-scale fermentation in three liquid media namely YM 6.3, Q6 ½, and ZM ½. Preliminary screening of the fungal crude extracts revealed that only the crude extract of Fusarium sp. (UST-UVG10) grown in ZM ½ medium showed potent antibacterial activity against Staphylococcus aureus (zone of inhibition (ZOI) = 13.85 mm, broth extract; zone of inhibition (ZOI) = 8.11 mm, mycelium extract) (Table 2).
Table 2. Antibacterial activity of the fungal crude extracts against Staphylococcus aureus.
Extract Zone of inhibition (mm) YM 6.3 Culture media 6.56 Mycelia 6 Q6 ½ Culture media 6 Mycelia 6 ZM ½ Culture media 13.85 Mycelia 8.11 Streptomycin 31.84 Minimum inhibitory concentration (MIC) vs Staphylococcus aureus and Mycobacterium tuberculosis H37Rv
-
During the test for TB susceptibility using M. tb H37Rv as the test pathogen and colorimetric MABA as screening method, extract YM 6.3 showed weak inhibition (MABA MIC = 62 μg/mL) while the rest of the extracts are negative.
Metabolomic profiling of fungal crude extracts
-
The constituents present in the fungal crude extracts were profiled using LC-HRESIMS and dereplicated using SIRIUS and GNPS (Fig. 3) to determine the putative metabolites present in the active extracts. In extracts YM6.3 and Q6 ½, the most abundant antibiotic compound is the depsipeptide mycotoxin, beauvericin (2) while pronounced in ZM ½ is the polyketide-derived picolinic acid, fusaric acid (1). Compound 1 is found in minor amounts in the YM 6.3 extract (Fig. 4).
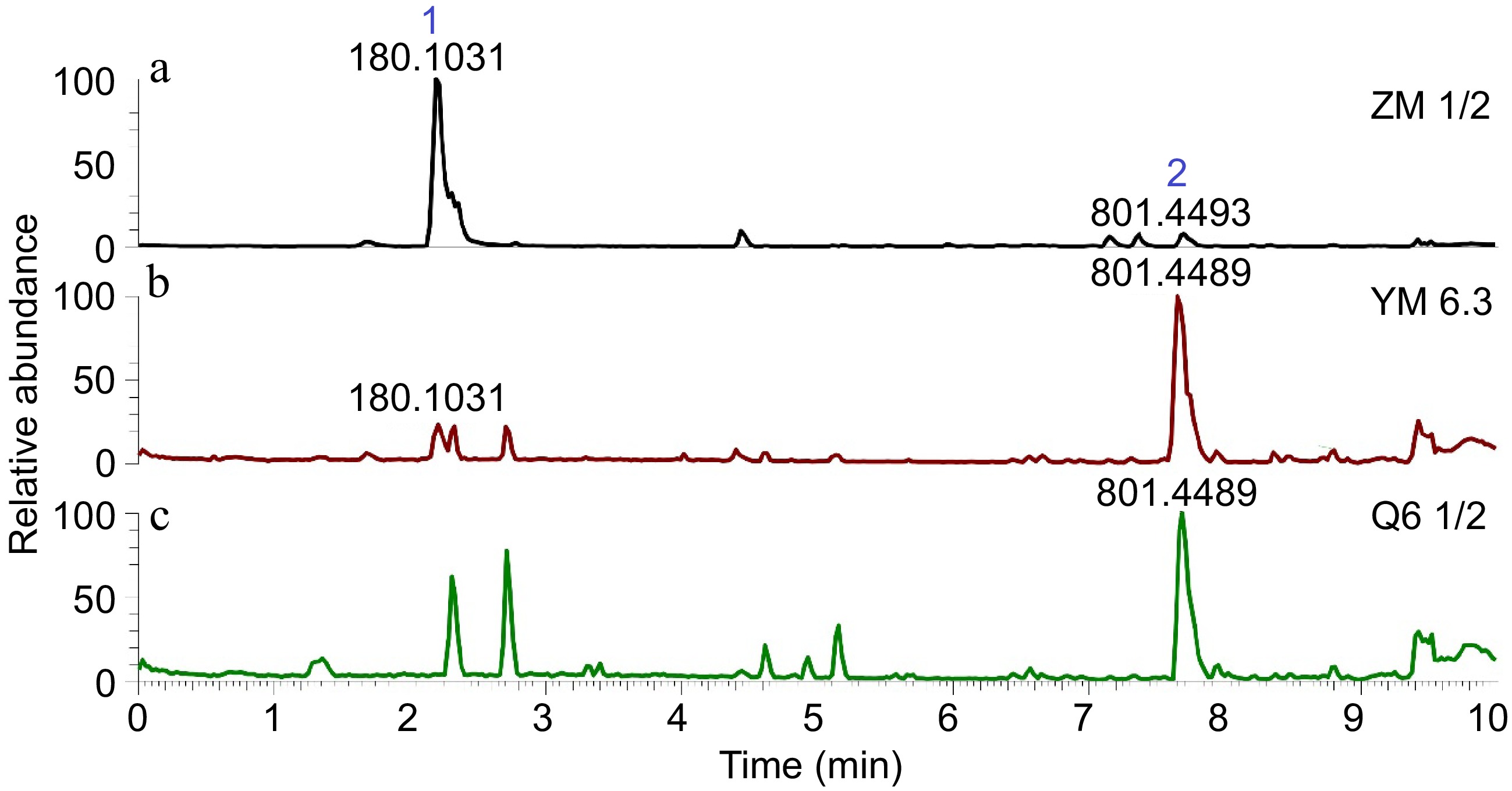
Figure 3.
LC-HRESIMS chromatograms of the EtOAc extracts of Fusarium sp. (UST-UVG10). (a) ZM ½, (b) YM 6.3, (c) Q6 ½.
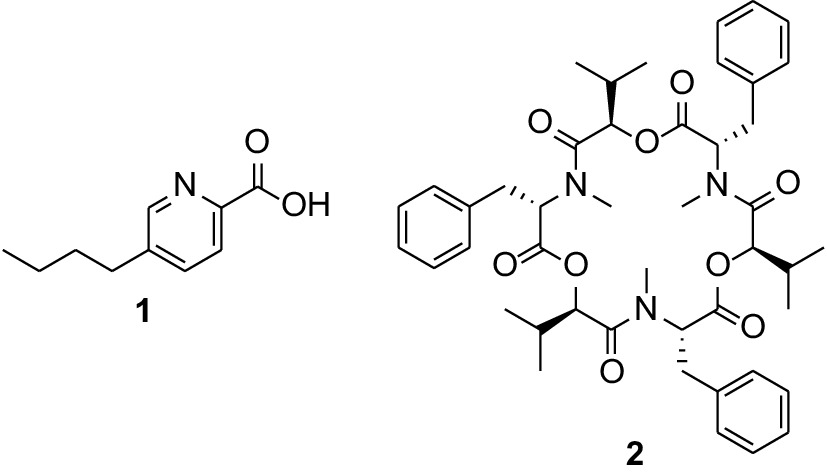
Figure 4.
Structures of the major antibiotic compounds fusaric acid (1) and beauvericin (2) identified from Fusarium sp. (UST-UVG10) extracts.
The major antibiotic compound 1 from the ZM ½ extract was purified by repeated preparative HPLC, identified spectroscopically, and screened for its inhibitory activity against S. aureus and M. tuberculosis H37Rv. Thus, fusaric acid exhibited potent inhibition against S. aureus (MIC = 1.30 μg/mL) and is comparable with the MIC of the positive drug antibiotic streptomycin. However, against M. tuberculosis H37Rv it showed antimycobacterial activity (MABA MIC = 52.4 μg/mL) (Table 3).
Table 3. Minimum Inhibitory Concentrations (MICs) of the fungal extracts against Staphylococcus aureus and Mycobacterium tuberculosis H37Rv.
Extract/Compound MIC vs S. aureus (μg/mL) MABA MIC vs M. tb H37Rv (μg/mL) YM 6.3 − 62 Q6 ½ − >64 ZM ½ − >64 Fusaric acid (1) 1.30 52.4 Streptomycin 1.30 − Isoniazid − 0.25 Rifampin − 0.12 Fusaric acid, 1: white solid; UV (MeOH) max 225, 269 nm.1H NMR (MeOH-d4, 600 MHz): 0.93 (3H, m, H-10), 1.36 (2H, m, H-9), 1.63 (2H, m, H-8), 2.73 (2H, t, J = 7.7 Hz, H-7), 7.89 (1H, d, J = 7.9 Hz, H-4), 8.09 (1H, d, J = 8.0 Hz, H-3), 8.49 (1H, s, H-6).13C NMR (MeOH-d4, 150 MHz): 12.8 (CH3, C-10), 21.9 (CH2, C-9), 32.1 (CH2, C-7), 32.8 (CH2, C-8), 124.8 (CH, C-3), 138.5 (CH, C-4), 143.0 (C, C-5), 145.5 (C, C-2), 149.7 (CH, C-6), 166.8 (C, C-11); HRESIMS m/z 180.1014 [M + H]+ (calcd for C10H14NO2, 180.1019).
-
The importance of selecting plants growing in certain locations as they often harbor secondary metabolites exhibiting diverse biological activities cannot be overemphasized[35]. This provides a unique breeding ground for microorganisms such as endophytic fungi found inside plant tissues. The host plant U. valderramensis is known to harbor and produce bioactive secondary metabolites[36−40]. This has prompted studies related to its associated endophytic fungi. The isolated endophytic fungus, Fusarium sp. (UST-UVG10) was chosen due to its frequent occurrence in the leaves of U. valderramensis. It was identified through morphological and molecular analysis and fermented in three different culture media resulting to extracts analyzed for metabolic components and assessed for antibacterial/antimycobacterial activity.
The induction of antimicrobial compounds using the one strain many compounds (OSMAC) approach is an efficient strategy for producing new secondary metabolites, and enhancing production of a specific metabolite[41]. This method alters fungal culture conditions and induces silent biogenetic clusters to produce different fungal natural products[42]. Secondary metabolites are produced at specific conditions, wherein their biosynthesis is diversely regulated[43]. The varying macro- and micronutrient components of the culture medium influence the production of microbial metabolites. Limiting a single nutrient’s availability affects the gene clusters involved in controlling S.M.s[44]. Mainly, carbon and nitrogen sources in the medium affect microbial secondary metabolism, including the C/N ratio also having a role in fermentation products[45−47]. Thus, metabolomic profiling on YM 6.3, Q6 ½, and ZM ½ using LC-HRESIMS experiments followed by analysis with GNPS and SIRIUS to determine putative secondary metabolites, revealed two major peaks identified as the known antibiotic compounds, fusaric acid (1) and beauvericin (2). Fusaric acid (1) is the abundant and major compound present in the ZM ½ medium with a peak area of 9.03 × 107, whereas on the other hand, beauvericin (2) was the major peak detected in the YM 6.3 and Q6 ½ extracts with a peak area of 1.40 x 107 and 9.28 × 106, respectively. Interestingly, fusaric acid (1) was not observed in the Q6 ½ LC chromatogram. Based on the inhibition data against S. aureus, extracts containing high amounts of 1 potently displayed antibacterial and antimycobacterial activity compared to the extracts that contain 2 as the principal peak component. This was supported by the MIC value of 1 (MIC = 1.30 µg/mL). Based on the peak area and intensity of the base peak of the three chromatograms, the presence of both fusaric acid (1) and beauvericin (2) led to significant antibacterial activity illustrating synergistic activity. The present study also reports the inhibitory activity of 1 against the virulent TB strain, M. tuberculosis H37Rv.
The ZM ½ medium is mainly composed of different types of sugars as carbon sources and uses ammonium sulfate as a nitrogen source. Comparing the varying components of each growth medium, ZM ½ is the only medium that contains a nitrogen source and a relatively high amount of sugar compared to YM 6.3 and Q6 ½. Amino nitrogen and high concentrations of sugar increase the yield of antibacterial compounds produced by Fusarium species[48]. Thus, we noted that ZM ½ medium was able to induce the production of the antimicrobial compound, fusaric acid (1) compared to other culture media based on the analysis of the LC-HRESIMS data. The use of complex carbon sources may also affect antibiotic production in microorganisms, having starch as one of the enhancer of carbon source[49]. Building blocks of starch such as glucose, a regulator for catabolite repression, enables and disables specific pathways including secondary metabolite production that heavily depends on the level of glucose present[50].
Fusaric acid (FA) and its derivatives exhibit antibacterial effects and are labeled as a non-specific toxin that has antibiotic effects on microorganisms[51−53]. It is a picolinic acid derivative and is known to be produced by several Fusarium species, for example F. oxysporum, F. fujikoroi, F. proliferatum, and F. verticillioides[54−58]. The proposed biosynthetic route for fusaric acid requires aspartate or oxaloacetate and three acetate units, cascading to the production of primary metabolites L-aspartic acid paired with malonyl-CoA derived from acetyl-CoA, which is consequently biocatalyzed by an enzyme complex to form fusaric acid[59]. To produce the secondary metabolite fusaric acid, the fungus must be cultured in an environment with a rich nitrogen source and sugar. This condition fuels glycolysis and tricarboxylic acid pathway and their respective precursors for the continuous synthesis of fusaric acid. Hence, using ZM ½ aides fusaric acid production. On the other hand, beauvericin (BEA) is a cyclic hexadepsipeptide mycotoxin produced by different Fusarium species specifically F. oxysporum, F. subglutinans, F. sacchari, and F. tardicrescens[60−62]. Biosynthesis of beauvericin is executed through rapid catalysis by nonribosomal peptide synthetases (NRPSs), beauvericin synthetase, much like any non-ribosomal peptides (NRPs). The metabolites required are L-phenylalanine, D-hydroxyisovaleric acid, ATP/Mg2+, and S-adenosyl-methionine (AdoMet)[63,64]. Non-ribosomal peptides are structurally diverse antibiotics developed and produced for a wide range of pharmacological uses[65]. The concentration of BEA in the three medium extracts is comparable due to probable substrate requirement for NRPS - from fatty acids, alpha-hyroxy acids, and alpha-keto acids, to heterocycles, which are mainly produced if a stable carbon source is present[66]. Beauvericin also has strong antibacterial activity against various gram-positive and gram-negative pathogenic bacteria including M. tuberculosis[64].
-
In the present study, the endophytic fungus Fusarium sp. (UST-UVG10) associated with Uvaria valderramensis was subjected to the OSMAC approach. Metabolomic profiling through LC-HRESIMS followed by dereplication revealed that the antibiotic fusaric acid is highly abundant in the extract obtained from the ZM ½ medium, while the depsipeptide beauvericin remains present in YM6.3 and Q6 ½. Thus, the ZM ½ extract exhibited potent antibacterial and antitubercular activities. A relationship between culture medium components and relative abundance of the antibiotic compounds fusaric acid and beauvericin was established as a potential platform for targeted metabolite production and antimicrobial activity enhancement.
-
The authors confirm contribution to the paper as follows: study conception and design: Macabeo APG; data collection: Drapisa HJ, Llames LC, Abellanosa EA, Vidar W; analysis and interpretation of results: Drapisa HJ, Llames LC, Abellanosa EA, Vidar W; draft manuscript preparation: Drapisa HJ, Abellanosa EA, Macabeo APG. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available in the GenBank repository. The ITS sequences of the fungus were recorded and deposited in GenBank with the accession number OQ873338. These data were derived from the following resources available in the public domain:
www.ncbi.nlm.nih.gov/genbank .Copies of NMR and HRMS spectroscopic data of the compounds are available on request from the corresponding author (APGM).
Funds and large equipment grant from the Alexander von Humboldt Foundation especially through its Digital Cooperation Fellowship (PHL 1163345 GF-P) during the COVID-19 pandemic is greatly acknowledged. This research was partially funded by the DOST Grants-in-Aide-National Research Council of the Philippines (NRCP). Finally, the DOST-ASTHRDP for supporting the graduate study of HJ Drapisa.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Drapisa HJC, Llames LC, Abellanosa EA, Vidar WS, Macabeo APG. 2024. Differential induction of fusaric acid in the endophytic fungus, Fusarium sp. (UST-UVG10) by three different media results to enhanced, variable anti-staphylococcal and antimycobacterial activity. Studies in Fungi 9: e007 doi: 10.48130/sif-0024-0007 

Differential induction of fusaric acid in the endophytic fungus, Fusarium sp. (UST-UVG10) by three different media results to enhanced, variable anti-staphylococcal and antimycobacterial activity
- Received: 25 January 2024
- Revised: 01 May 2024
- Accepted: 21 May 2024
- Published online: 18 June 2024
Abstract: The rise of drug-resistant pathogenic bacteria has enabled the discovery and innovative biotechnological production of antimicrobial substances. Among the widely used strategies to induce antimicrobial substances from fungi is the One Strain Many Compounds (OSMAC) approach where alteration of cultivation parameters are explored. Herein, we report a comparative study on the metabolic profile and antibacterial/antimycobacterial activity of the extracts obtained from the endophytic fungus Fusarium sp. (UST-UVG10) associated with the Philippine endemic Annonaceae plant Uvaria valderramensis using the OSMAC approach. Fusarium sp. (UST-UVG10) was fermented on three different media (Q6 ½, ZM ½, and YM 6.3) to obtain crude extracts for chemical profiling and antimicrobial activity assessments. Among the three extracts, only the ZM ½ extract exhibited strong inhibition against Staphylococcus aureus (ZOI = 13.85 mm). Evaluation of antituberculosis activity using the Microplate Alamar Blue Assay (MABA) showed the YM 6.3 extracts to exhibit inhibition against the fast-growing Mycobacterium tuberculosis H37Rv (MIC = 62.0 μg/mL). Metabolic profiling using liquid chromatography-high resolution electrospray ionization mass spectrometry followed by dereplication with Global Natural Product Social Molecular Networking (GNPS) and SIRIUS databases illustrated induction of the mycotoxin picolinic acid derivative, fusaric acid in ZM ½ and the fungal antibiotic cyclodepsipeptide beauvericin in YM6.3 extracts. Fusaric acid was isolated, characterized, and exhibited profound antibacterial activity against S. aureus (MIC = 1.30 μg/mL) with moderate antimycobacterial activity against M. tuberculosis H37Rv (MIC = 52.4 μg/mL). Overall, the present study demonstrated the correlative influence of culture medium components and the relative abundance of specific antimicrobial compounds, establishing a potential platform for targeted metabolite production.
-
Key words:
- Antibacterial /
- Antitubercular /
- Fusaric acid /
- Endophytic fungi /
- Fusarium proliferatum /
- OSMAC