








-
The increasing global population, climate change, and environmental pollution pose significant challenges to agricultural production[1−3]. Concurrently, the public's demand for safe and high-quality agricultural products continues to rise. Therefore, it is of great urgency to seek environmentally friendly methods for crop pests and diseases control, while also increasing yields, to establish a sustainable agricultural production system. As sessile organisms, plants have evolved sophisticated processes that provide protection against herbivory and infection, allow access to limited resources, and promote growth and development in rapidly changing environments. JA and SA are two well-known defense-related phytohormones[4−7]. Plant pathogens exist as either biotrophs or necrotrophs, where biotrophs feed on living cells and necrotrophs of necrotic cellular debris[8]. JA plays a critical role in necrotrophic infection as well as herbivory[9,10], while SA is required for resistance to biotrophic pathogens, pattern-triggered immunity (PTI), and systemic acquired resistance (SAR)[8,11]. With more investigations work on the signaling pathways of these hormones, it has become clear that light influence defense responses such as systemic acquired resistance (SAR). For instance, research demonstrates that SAR is completely lost in the dark, while SAR occurs under both moderate and intense light conditions in Arabidopsis[12].
Light not only provides the energy necessary for photosynthesis but also serves as a signal, transmitting environmental information that enables plants to better adapt to dynamic surroundings[13]. Key components of light perception are phytochromes which are photoreceptors that perceive red (R) and far-red (FR) light. Inactive phytochromes are present in the cytosol (Pr form) but move into the nucleus when active (Pfr form)[14]. Light perception can also mediate JA and SA biosynthesis and signal transduction. It mediates the JA-SA pathway, encompassing both its biosynthesis and signal transduction. Mechanistic studies have indicated photoreceptors, particularly phytochrome B (phyB)[15−17], pivotal transcription hubs like ELONGATED HYPOCOTYL 5 (HY5)[18], PHYTOCHROME INTERACTING FACTORS (PIFs)[19], and FHY3 (FAR-RED ELONGATED HYPOCOTYL3)/FAR1 (FAR-RED IMPAIRED RESPONSE 1)[20] and a few other players in the light signal networks[21−23] as participants in the modulation of stress responses, growth, development, and the regulation of JA and SA biosynthesis and signal transduction. Consequently, the manipulation of the light environments in cultivated crops has the potential to optimize crop production[24] and enhance stress resistance[25].
JA and SA have been extensively studied for their pivotal roles in plant defense and response to abiotic stress. Recent studies have revealed that various light signaling components, including photoreceptors and fundamental transcriptional hubs, play a role in JA/SA-regulated stress responses. Key components of the JA pathway, CORONATINE-INSENSITIVE (COI), JASMONATE ZIM-DOMAIN (JAZ) proteins, MYELOCYTOMATOSIS 2 (MYC2), and JASMONIC ACID-INSENSITIVE1 (JAI1), as well as NPR1 (NONEXPRESSOR OF PATHOGENESIS-RELATED GENES 1), SID2 (SA INDUCTION-DEFICIENT2)/ICS1 (ISOCHORISMATE SYNTHASE 1) in the SA pathway, play roles in light signaling pathways. Both JA and SA biosynthesis are initiated in the chloroplast, an organelle whose function is light-dependent, thus alluding to a complex interplay between light and JA/SA signaling during plant stress.
Recent advances have shown that light signaling participates in JA and SA biosynthesis and transduction pathways in response to both biotic and abiotic stress. Additionally, studies regarding light-modulation of JA/SA pathways in plant growth, and development have emphasized the delicate balance between plant growth, resource competition, and defense. In this review, recent advances in the multifaceted light-mediated role of JA/SA pathways during plant adaptation are highlighted. Additionally, the opportunities to apply these fundamental theoretical insights into agriculture production are discussed. This includes the use of LED-based greenhouse light manipulation[26,27], intercropping[28,29], rational close planting[30], and other agronomic techniques, as well as crop ideal architecture breeding[31] to optimize the growth and development of horticultural crops thus improving plant productivity.
-
JA and SA are critical regulators of plant immune responses[32]. Significant research over the decades has worked to elucidate the complex cross-talk that exists between JA and SA in the context of defense. More recently, an additional focus has been given to the light's role in hormonal regulation (Fig. 1). Plants have evolved a dynamic allocation known as the 'growth-defense trade-off'[10], which leads to the careful allocation of energy resources. Light provides the currency that plants must meticulously divide between metabolically expensive defense processes and growth[8−10,33−36]. A deeper understanding of how light regulates the JA/SA pathways during plant defense is critical for modern agriculture, as it could allow for precise manipulation of crop traits under the constraints of environmental stress and challenges.
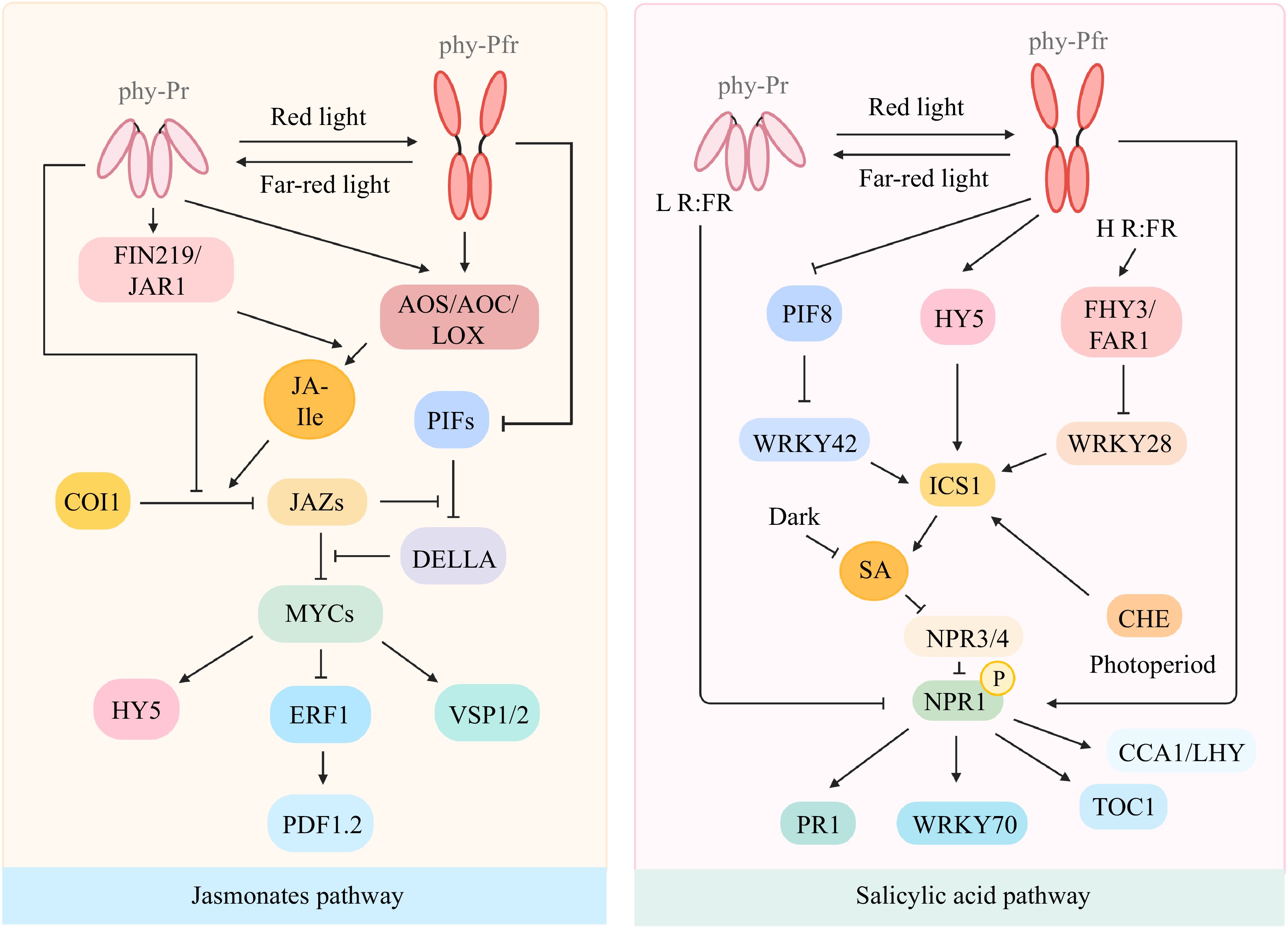
Figure 1.
Crosstalk of light and JA/SA during plant defense and development. Key components in light signal transduction such as phyB, PIFs, and HY5 play roles in JA/SA signaling through manipulating AOS/AOC/LOX, JAZs, MYCs (fundamental elements in JA biosynthesis or signaling) and ICS1, NPR1, WRKYs (key players in SA biosynthesis or signaling). Created with
BioRender.com .As an important plant hormone, JA has been widely documented for its role during biotic stress. Recently, the interaction between light and JA has garnered increased interest in various plants, including Arabidopsis, soybean, and tomato (Table 1). In Arabidopsis, it has been found that FR light can suppress genes involved in fungal defense, such as PLANT DEFENSIN 1.2 (PDF1.2), which is a classical marker for JA production. This light-mediated suppression is due to the inhibition of the JA-responsive AP2/ERF transcription factor, ETHYLENE RESPONSE FACTOR 1 (ERF1)[37]. In contrast, FR light has been shown to activate the transcription of genes related to insect defense, such as VEGETATIVE STORAGE PROTEIN 1 (VSP1) and VEGETATIVE STORAGE PROTEIN 2 (VSP2), through the activation of a key JA transcription factor, MYC2[31,32]. In addition to FR light, red light also can influence the defense response of Arabidopsis against fungal infections. Studies have shown that both red and white light can enhance the expression of various genes involved in JA biosynthesis through the photoreceptor phyB, resulting in an increased accumulation of the metabolically active JA hormone, jasmonyl-isoleucine (JA-Ile). The accumulation of JA-Ile was found to be due to the light-induced degradation of the JA-repressor, JAZ9, in a COI-dependent manner[33]. This suggests that phyB-mediated light signaling may play a role in Arabidopsis' defense against pathogens like Botrytis cinerea, by modulating JA biosynthesis and metabolism, thereby influencing JA levels and the stability of JAZ proteins. Intriguingly, parallel findings from a recent study on tomato indicate that phyB signaling enhances resistance to Botrytis cinerea, through the JA-dependent regulation of soluble sugars[34]. Additionally, blue light enhances soybean defenses against the Soybean mosaic virus (SMV) by more effectively activating JA signaling and upregulating GmMYC2 and GmERF genes compared to white light[37]. Conversely, under FR light treatment, most key genes within the JA signaling pathway are downregulated in SMV-infested soybean plants.
Table 1. Effect of light treatment on JA-modulated plant defense.
Species Pathogens/insects Mechanism Light treatment and
key light elementArabidopsis Botrytis cinerea Red light can regulate JA biosynthesis and metabolism through
phyB signaling, affecting the stability of JAZ9 protein[38].Red light treatment,
phyB signalingArabidopsis Diamondback moth
(Plutella xylostella)UV-B decreases the attractiveness of Arabidopsis plants for the diamondback moth in a JA signaling-dependent manner[39]. UV-B treatment Soybean Soybean mosaic virus (SMV) Blue light treatment can induce JA signaling pathway and increase
the expression of GmMYC2 and GmERFs[40].Blue light treatment Soybean Fusarium verticillioides JA can promote resistance to infection by promoting the accumulation
of isoflavone in soybean pods; vegetative stage shading can promote isoflavone accumulation and improve pod resistance to Fusarium verticillioides[41].Vegetative stage
shading treatmentTomato Botrytis cinerea FR light inhibits phyB signaling, thus reducing JA response, and
resulting in elevated leaf glucose and fructose levels, and enhancing
tomato sensitivity to disease caused by Botrytis cinerea[42].FR light treatment,
phyB signalingTomato Thrips (Frankliniella occidentalis) High photosynthetically active radiation (PAR) increased thrips
resistance against thrips in tomato by inducing the expression of
JA-responsive defense-related genes (such as PROTEINASE INHIBITOR-IIf
(PI-IIf), THREONINE DEAMINASE-2 (TD-2) and JASMONATE INDUCIBLE
PROTEIN-21J(JIP-21))[43].Light intensity Broccoli Pieris brassicae Supplementary levels of moderate UV-B on broccoli sprouts increased the expression of JA signaling genes, while negatively affecting the performance of Pieris brassicae caterpillars[44]. UV-B treatment Watermelon Root knot nematode (Meloidogyne incognita) Red light can significantly increase the expression of JA biosynthesis genes (AOS and LOX), and JA content in roots, triggering plant
defenses against nematodes[45].Red light treatment Interactions between light and JA are not confined to Arabidopsis, with extensive documentation in a variety of other plants. In the case of watermelon plants infected with the root-knot nematode, Meloidogyne incognita (RKN), it has been observed that treatment with red light can reduce the number of galls in the roots and suppress nematode incidence. Additionally, the foliar application of methyl jasmonate (MeJA) has been shown to significantly decrease RKN infection. Further research has revealed that under red light treatment, the expression of JA biosynthesis genes, specifically ALLENE OXIDE SYNTHASE (AOS) and LIPOXYGENASE (LOX), as well as JA content in the leaves and roots are significantly increased compared to those treated with white light[35]. This increase in JA accumulation in both leaves and roots due to red light may be facilitated by the translocation of JA from the leaves to roots through the vascular tissues[36], thereby triggering the defense mechanisms against nematodes.
Currently, research on Arabidopsis has thoroughly investigated the intricate signaling pathways through which light signals regulate JA. However, for other plant species, studies predominantly concentrate on the alterations in plant hormone levels following light exposure. Consequently, for horticultural crops like tomato, soybean, and watermelon, there remains considerable scope for exploring the specific role of light on regulatory pathways involved in JA signaling.
-
Over the past decades, research has consistently demonstrated the crucial role that SA plays in plant defense against pathogens with a biotrophic lifestyle[46]. The mediation of the SA pathway by light has been investigated in response to various pathogens (Table 2), including fungi (e.g. powdery mildew[47]), bacteria (e.g. Pseudomonas syringae pv. tomato DC3000 (Pst DC3000)[48]), oomycetes (e.g. Phytophthora capsici[18]), viruses (e.g. SMV[40]), and nematodes (e.g. RKN[45]). The underlying mechanisms have been deeply explored in the model plant Arabidopsis, as well as crops such as tomato[48] and soybean[40]. Research indicates that light directly influences the accumulation of SA, likely due to one of the SA biosynthesis pathways, known as the isochorismate (ICS) pathway, initiating in the chloroplast[49]. Additionally, there is evidence that continuous light exposure can elicit resistance against Pst DC3000, an effect that is SA-dependent. This phenomenon occurs when constant light stimulates SA production, leading to increased stomatal opening and a reduction in the bacterial-induced aqueous microenvironments which causes water-soaked lesions during infection[50]. In Arabidopsis, it has also been observed that the spontaneous cell death phenotype observed in bak1 bkkk1 double mutants is observed only in the light due to the overproduction of SA[51]. The SA overaccumulation is also able to enhance resistance to Pst DC3000 in a mechanism distinct from PTI.
Table 2. Effect of light treatments on SA-mediated plant defense.
Species Pathogens Mechanism Light treatment and
key light elementCucumber Powdery mildew Compared with white light, Red light increases the expression of SA signaling marker genes (PR1, WRKY30, and WRKY6) and improves disease resistance[52]. Red light treatment Oriental melon Powdery mildew Red light promotes SA biosynthesis and resistance against powdery mildew through the PIF8-WRKY42-ICS module. PIF8 serves as a negative regulator of WRKY42, thereby inhibiting transcriptional activation of downstream ICS[47]. Red light treatment Arabidopsis Pseudomonas syringae
pv. tomato DC3000Constant light induces the production of SA, which counters effector-induced stomatal closure by Pst DC3000, thus allowing for transpiration and inducing SA-related disease resistance[50]. Constant light treatment Tomato Pseudomonas syringae
pv. tomato DC3000A 12-hour red light exposure at night enhanced tomato resistance, significantly upregulating transcription factors including WRKY18, WRKY53, WRKY60, and WRKY70, while NPR1 silencing partly reduced Pst DC3000 resistance induced by red light[48]. Red light treatment
during the nightPepper Phytophthora capsici Red light induces SA accumulation through HY5 to enhance
resistance against Phytophthora capsici[17].Red light treatment; HY5 Soybean Soybean mosaic virus Blue light triggers soybean resistance to SMV by orchestrating
SA and JA defense pathways[40].Blue light treatment Furthermore, the light signaling cascade encompasses photoreceptors, transcription factors, and various downstream functional elements. Investigations regarding the role of phytochromes in SA-mediated plant defense have revealed that mutations in phyA and phyB heighten the sensitivity of Nicotiana tabacum to Cucumber mosaic virus (CMV) and Chili veinal mottle virus (ChiVMV) by inhibiting the SA-mediated defense pathway in tobacco plants[53,54]. Research has shown that the deactivation of phyB results in a reduction in SA-mediated defense mechanisms[55]. In Arabidopsis, the SA signaling transduction pathway is significantly compromised in phyA and phyB mutants, concurrent with a marked decrease in the expression of the PATHOGENESIS-RELATED 1 (PR1) gene[56]. Extensive research has shown that the deactivation of phyB results in a reduction in SA-mediated defense mechanisms[55].
In recent years, emerging research has indicated that various transcription factors involved in light signaling participate in the SA pathways. Two pivotal transcription factors in light signal transduction, PIFs, and HY5, have garnered particular attention. In oriental melon, red light has been shown to facilitate SA biosynthesis through the PIF8-WRKY42 regulatory module, thereby bolstering the plant’s resistance to powdery mildew[47]. Phenotypic evidence suggests that following immune activation, PIFs can enhance the resistance against Pst DC3000 via the SA pathway as well[50]. In pepper, red light is known to trigger the accumulation of SA through HY5, which in turn strengthens resistance against Phytophthora capsici[18]. This enhancement is attributed to the induction of CaHY5 expression by red light, leading to the activation of SA biosynthesis genes CaPHENYLALANINE AMMONIA-LYASE 3 (CaPAL3) and CaPAL7. The activation of these genes results in SA accumulation and the upregulation of SA response genes CaPR1 and CaPR1L, thereby increasing the plant's resistance to the pathogen[18].
In addition to capturing light for photosynthesis, the quality, intensity, period, and duration of light are critical factors that influence plant responses to stress and environmental changes. Historically, research on light quality has predominantly concentrated on the effects of red, far-red, blue, and ultraviolet (UV) light, with relatively less attention given to green light. However, a study has demonstrated that a variety of light treatments, including purple, blue, green, yellow, and red light, all effectively suppress the growth of the pathogen Pst DC3000 on tomato leaves when compared to the dark control[48]. More recent investigations into the light-mediated SA pathway in plant defense have particularly highlighted the benefits of red light treatment[18,47] or low-red to far-red (R: FR) light ratio conditions[57]. Red light has been identified as a positive regulator of the SA pathway in various plants, including Arabidopsis, tomato[48], pepper[18], oriental melon[47], and soybean[40], enhancing their defenses against a range of biotic stresses. While the specific mechanisms differ among plant and pathogen species, the consensus among numerous studies is that red light markedly stimulates SA biosynthesis and signaling transduction during the defense processes. The impact of red light treatment during fungal infections, such as powdery mildew, has been particularly examined in cucumber[52] and oriental melon plants[47], with findings suggesting that SA is a key component in the defense mechanism modulated by red light. Under red light, the expression levels of SA-associated genes such as PR1, WRKY30, and WRKY6 are significantly higher compared to those treated with white light[52]. Moreover, red light treatment has also been found to enhance plant resistance to bacterial invasion. For example, RNA sequencing data following a 12-h red light treatment, especially during the night, has been shown to bolster tomato plants' resistance. RNA sequencing data revealed the involvement of SA in red light-induced resistance in tomatoes challenged with Pst DC3000. The transcription levels of several defense-related transcription factors, including WRKY18, WRKY53, WRKY60, and WRKY70, are notably increased by red light exposure. Furthermore, silencing the SA receptor NPR1 partially diminishes the red light-induced resistance in tomato plants[45,48]. In the case of watermelon, red light has been shown to enhance resistance to RKN, at least partially through the promotion of SA-dependent pathways[45]. These findings suggest that red light stimulates downstream SA transduction cassette, thus changing SA biosynthesis genes at the transcriptional level, activating certain kinases, and ultimately leading to increased SA accumulation, thereby fortifying the plant's defense capabilities of the treated plants.
Under high plant density, the composition of light reaching the vegetation canopy changes, leading to a set of developmental responses known as the shade-avoidance syndrome (SAS). Traditionally, SAS is associated with a low R: FR light ratio, while a high R: FR ratio indicates the opposite condition. Research suggests that the quality of light can influence plant defense mechanisms by modulating the SA pathway[15]. In Arabidopsis, it has been observed that a low R: FR ratio diminishes pathogen resistance, with some underlying mechanisms already identified. At the transcriptional level, a low R: FR ratio results in decreased SA-related gene expression, and repressed SA-activated kinases[15]. Additionally, the phosphorylation of NPR1 protein is reduced in the nucleus, thus weakening the pathogen defenses that depend on SA[15]. In soybean, FR light has also been shown to inhibit SA-dependent genes in plants infected with SMV[40].
Compared to red light, the impact of blue light signaling on the SA pathway has not been as thoroughly explored. However, it is intriguing to note that blue light has been shown to enhance soybean resistance to the SMV by coordinating both SA and JA defense pathways. In addition to blue light, ultraviolet light also exerts a significant influence on SA signaling. UV light has been observed to elevate SA content, leading to an upregulation in the expression of SA-associated genes in tomatoes and tobacco leaves[27,58].
In addition to various light-quality treatments, research indicates that photoperiod also plays a role in regulating SA production and defense against pathogenic attacks[59]. The central circadian clock oscillator, CCA1 HIKING EXPEDITION (CHE), is essential for the synthesis of SA in response to pathogens[60]. Moreover, the SA-related immune regulator NPR1 regulating both the morning-phased CIRCADIAN CLOCK ASSOCIATED 1 (CCA1)/LATE ELONGATED HYPOCOTYL (LHY) and the evening-phased TIMING OF CAB2 EXPRESSION 1 (TOC1), enables plants to regulate their immune responses in the morning and minimize growth costs at night[59]. However, numerous facets of the control that photoperiod and circadian rhythm contribute to plant growth and stress responses remain unclear.
Light quality may also influence the crosstalk between JA and SA. Some indirect evidence suggests that red and blue light can induce resistance to SMV in soybean by coordinating the defense pathway of SA and JA, but this hypothesis requires further testing[40]. UV radiation has been observed to strongly activate SA-related defense responses in JA-deficient genotypes after thrips infection. Further research indicates that UV radiation may enhance tomato resistance to Pst DC3000 in JA-deficient genotypes through the activation of SA defense[61]. However, this evidence is indirect and circumstantial.
-
The interaction between light and JA has a significant impact on abiotic stress responses (Table 3). Research by Wang et al. indicated that FR light and a low R/FR light positively influence the cold tolerance of tomato plants[17]. Under FR light, phyA activates abscisic acid (ABA) signaling, which in turn induces the expression of downstream JA signaling components, leading to increased expression of C-REPEATBINDINGFACTOR1 (CBF1) and enhances cold tolerance in tomatoes[17]. In contrast, red light was found to have a negative effect on the cold tolerance of these plants. The involvement of phytochromes, specifically phyA and phyB, in SAS, is also well-established. PhyB suppresses SAS under high R: FR conditions, while phyA does so under low R: FR conditions. Studies have revealed that COI1, a key player in JA signaling, is crucial for phyA-mediated inhibition of SAS. Under low R: FR conditions, phyA regulates SAS by engaging JA signaling and promoting the degradation of JAZ1, which is downstream of COI1[31]. Further work has shown that in this pathway, transcription factors such as MYC2, MYC3, and MYC4 are degraded by JAZ1[62]. JA and phyA signaling stabilizes these transcription factors, thereby altering plant SAS. JA signaling also participates in phosphate responses under shade conditions. JAZ directly interacts with PHOSPHATE STARVATION RESPONSE1 (PHR1) to repress the transcriptional activity of phosphate starvation-induced genes. Moreover, FHY3 and FAR1 directly bind to the promoters of NITRATE-INDUCIBLE, GARP-TYPE TRANSCRIPTIONAL REPRESSOR1.1 (NIGT1.1) and NITRATE-INDUCIBLE, GARP-TYPE TRANSCRIPTIONAL REPRESSOR1.2 (NIGT1.2) to activate their expression while JAZs suppress these processes[63], enriching our understanding of the relationship between light and nutrient intake regulated by JA.
Table 3. Effect of light treatments on JA/SA-mediated abiotic stress.
Phytohormone
typeSpecies Abiotic stress Mechanism Light treatment and
key light elementJA Tomato Cold stress PhyA induces the expression of JA signaling components, increases the expression of CBF1, thus enhances cold tolerance in tomatoes[17]. Low R/FR light treatment Arabidopsis Heat and
high light stressCombined high light and heat stress will increase the levels of JA and JA-Ile, as well as the expression of transcripts related to JA biosynthesis[64]. Additionally, a JA-deficient mutant (aos) is more sensitive to heat stress[64]. High light treatment Arabidopsis High light/
UV-B stressTCP4 interacts with UVR8, activating the transcription of the JA synthesis gene LOX2, which subsequently improves UV tolerance[21]. High light treatment SA Barley Cold acclimation SA levels were lowered under WFR and WFRB light conditions compared to W light[65]. Blue and FR light supplementation to white light (WFRB), white light enriched with FR (WFR) Barley Cold acclimation FR light represses SA levels at low temperatures in Barley leaves. This phenomenon may exhibit similarities to the SAS[66]. Far-red light supplementation Tomato Chilling stress The SA biosynthesis gene SlPAL5 alleviates photosystem II damage under chilling stress[67]. / Arabidopsis High light stress High light conditions have been found to increase SA content[62]. Exogenous SA application can alleviate photoinhibition and improve photoprotection from high light in Arabidopsis[68]. High light treatment Rice High light stress High light significantly increases SA levels[62]. Endogenous SA protects rice from oxidative damage caused by high light[63]. High light treatment Similarly, the interaction between JA and light can affect heat stress responses. Combined high light and heat stress were found to increase the levels of JA and JA-Ile, as well as the expression of over 2,200 transcripts, including those related to JA biosynthesis[67]. Additionally, a JA-deficient mutant (aos) exhibited increased sensitivity to combined light and heat stress treatments, highlighting the positive regulatory role of JA in plant responses to such stress conditions[67]. In nature, high light is often accompanied by high UV-B. In Arabidopsis, the interplay between JA and UV-B signaling is modulated when TCP FAMILY TRANSCRIPTION FACTOR 4 (TCP4) interacts with UV RESISTANCE LOCUS 8 (UVR8), activating the transcription of the JA synthesis gene LOX2, which subsequently improves UV tolerance[21].
-
Light conditions can influence SA production and alter resistance to abiotic stress (Table 3). During cold stress, white light enriched with FR (WFR) and blue light (WFRB) has been observed to inhibit SA levels, a response akin to SAS in barley leaves[65]. There is evidence that SA also contributes to photoprotection in rice and Arabidopsis. Recent research has concentrated on both the impact of exogenous SA application and the activation of endogenous SA pathways. In tomatoes, the SA biosynthesis gene SlPAL5 mediates two SA response pathways that are involved in the protection of photosystem II under chilling stress[67]. High light conditions have also been found to elevate SA content in Arabidopsis and rice[62]. Endogenous SA can mitigate photoinhibition, indicating that SA may have played a role in the adaptation to increased light exposure following the terrestrialization of plants[62]. Furthermore, SA has been shown to protect rice plants from oxidative damage induced by intense sunlight[63]. Both phenotypic and protein-level evidence supports the idea that the application of exogenous SA can reduce photoinhibition and safeguard photosystem II from high light intensity in Arabidopsis[68].
-
Recent studies indicate that JA can influence various developmental processes in a light-dependent manner, including the secretion of extrafloral nectar[69], the elongation of hypocotyls[70,71], the accumulation of anthocyanin and chlorophyll, and so on. In lima beans, it has been observed that JA stimulates the secretion of extrafloral nectar (EFN) in a light-dependent fashion[69]. JA decreases EFN secretion in the dark but induces it under light conditions. This regulatory effect is also modulated by the R: FR light ratio, where the light environment can control the biosynthesis of JA-Ile, thereby affecting subsequent EFN secretion.
In Arabidopsis, the interplay between light and JA has a multifaceted impact on photomorphogenesis. Research has uncovered that phytochrome A (phyA) negatively regulates the expression of JA-responsive genes in a FIN219/JAR1-dependent process. Significantly, FAR-RED INSENSITIVE 219 (FIN219)/JASMONATE RESISTANT1 (JAR1) can bind directly to phyA, reducing its levels and activity. Additional studies have demonstrated that FR light and MeJA can increase the binding of FIN219/JAR1 to the active Pfr form of phyA, which inhibits hypocotyl elongation and promotes the accumulation of anthocyanin and chlorophyll[70]. Moreover, FIN219/JAR1 facilitates the nuclear accumulation of HY5 by modulating CONSTITUTIVELY PHOTOMORPHOGENIC 1 (COP1) exclusion, leading to hypocotyl elongation[71−73]. Additionally, it is found that the long hypocotyl phenotype of the Arabidopsis phyB mutant is partially suppressed by the overexpression of MYC2. The study further points out that MYCs directly bind to the promoter of HY5 and regulate its expression[74]. However, under blue light, MYC2 physically interacts with HY5, and at the transcriptional level, MYC2 negatively regulates the expression of HY5, while HY5 also suppresses the expression of MYC2[74,75]. Evidence has also emerged that JAZ3 interacts with the light-responsive transcription factor PIF4, enhancing the liquid-liquid phase separation (LLPS) of PIF4 during Arabidopsis photomorphogenesis[19]. Currently, research into how JA regulates plant photomorphogenesis is limited, primarily focusing on hypocotyl elongation in Arabidopsis and rice. Beyond model plants, investigating how light and JA interact to modulate photomorphogenesis in horticultural crops is an area of interest. For instance, it is worth exploring whether light can modify plant architecture through hormone-related pathways, thereby facilitating dense planting and improving space utilization. Beyond photomorphogenesis, there are also several related studies on the role of light-regulated JA signals in controlling traits such as fruit quality. For instance, in eggplant, SmJAZ5/10 interacts with SmMYB5 synergistically activating the expression of SmF3H (FLAVANONE 3-HYDROXYLASE) and SmANS (ANTHOCYANIDIN SYNTHASE), thereby promoting anthocyanin synthesis in eggplant fruit peel[23]. Other secondary metabolites, such as artemisinin, are also regulated by JA-light interactions, where AaMYB108 interact with AaCOP1 and AaJAZ8 respectively, upregulating artemisinin biosynthesis in Artemisia annua[22].
Compared to the well-studied interactions between light and JA during plant development, the interplay between light and SA remains relatively unexplored. Recent findings have primarily concentrated on how light influences stomatal development[76], senescence[19], and the effects of vegetable quality[77]. SA is integral to the plant hormone signaling network during senescence, and maintaining the SA pathway can help inhibit plant aging under stress[78,79]. Several studies have implied that light can modulate the SA pathway during senescence. Direct evidence in Arabidopsis has shown that light impacts SA-mediated senescence, with high R: FR light partially repressing leaf senescence through the FHY3-WRKY28 transcriptional module[21]. ICS1 is a direct target of WRKY28; FHY3 negatively regulates SA signaling by binding to the promoter and repressing WRKY28, thus inhibiting ICS1-mediated SA biosynthesis. Light also influences SA-related stomatal regulation, with studies suggesting that SA biosynthesis is involved in the recovery of stomatal conductance after periods of darkness[76]. In contrast, SA is thought not to be implicated in darkness-induced stomatal closure since SA synthesis is light-dependent.
Research has also indicated that certain light qualities can modulate gene expression and promote the accumulation of bioactive compounds. In Broccoli (Brassica oleracea var. italica), it was found, compared with white light, blue light significantly inhibits the expression of SA synthesis gene BoPAL and BoBENZOIC ACID 2-HYDROXYLASE (BA2H)[80]. Since SA inhibits the production of the glucosinolate-based secondary metabolite, sulforaphane (SFN), blue light was able to reduce SA accumulation and promote SFN production.
Polyamines are low molecular weight organic compounds that regulate plant growth, development, and stress response. Studies have also shown that the application of exogenous SA results in polyamine metabolism that is dependent on light in tomatoes[77]. There may be practical value in combining light treatment with exogenous SA applications to maximize development and growth alteration on commercial crops.
-
Plants have evolved a sophisticated energy allocation system to balance defense against pathogens and herbivores while competing for sunlight in resource-limited environments[57,81]. Within this system, the interplay between light, JA, and SA is crucial for striking a balance between growth, competitive ability, and defense mechanisms. Over time, significant progress has been made in understanding the crosstalk between JA and SA with key components of light signaling, especially in the context of shade avoidance and defense responses[82].
Light plays a significant role in the equilibrium between plant growth and defense. In Arabidopsis, MYC2-mediated SA biosynthesis and signaling, which is essential for resistance against bacterial pathogens, are partially regulated by light[83]. Evidence suggests that under low R: FR light conditions, plants may prioritize light foraging for rapid growth over defense[15,84]. Furthermore, low R: FR light ratio can inhibit both SA-dependent and JA-dependent resistance to diseases[15]. For example, under such conditions, phyB is predominantly in the inactive Pr form, allowing PIF4 to activate the transcription of the sulfotransferase gene STa (SULFOTRANSFERASE a). STa catalyzes the sulfation of OH-JA, reducing its availability for the JA-Ile pathway in Arabidopsis[85]. Concurrently, the phosphorylation of NPR1 is significantly reduced, impairing SA-mediated resistance. An intriguing discovery is that under suboptimal light, MYC2 is necessary for the microbiota-root-shoot circuit in Arabidopsis to combat Pst DC3000 and Botrytis cinerea[86]. Low R: FR ratios inactivate the photoreceptor phyB, rapidly reducing DELLA protein abundance, and the inhibitory effect of FR on jasmonate signaling is impairing in gai-1 mutants, thus promoting JAZ10 stability, urging the plants for rapid growth[87]. Interestingly, a study shows in rice and Arabidopsis, JAZ9 could interrupt RGA (a DELLA protein)-PIF3 interaction[88]. During SAS, plants may aggressively compete with their neighbors, potentially sacrificing JA/SA-dependent defense abilities. While these mechanisms have been primarily discovered in Arabidopsis, some research has also been conducted on crops, particularly tomatoes[61].
These findings align with the observation that high planting densities in agriculture often correspond with increased pests and disease indices. This underscores the urgency of breeding crops with ideal architectures for high-density planting, as well as exploring supplementary lighting and rational close planting strategies. The influence of plant light sensing on microbial root commensals may necessitate a reevaluation of soil microbiome management strategies. However, the dependency of these phenomena on light intensity or disease indices requires further investigation. Future research could delve into how light mediates JA and SA pathways to drive a trade-off between growth, competition, and stress resistance or tolerance in vegetable plants such as tomatoes. Additionally, whether this trade-off[56,61] varies among species demands more detailed research. Understanding how plants adapt to various light conditions could inform strategies for optimizing agricultural resource configuration through practices like intercropping, relay cropping, and light supplementation. The interaction of well-known hormone pathways, including gibberellins (GA), abscisic acid (ABA), strigolactones (SL), and RALF (RAPID ALKALINIZATION FACTOR) under diverse light conditions remains an enigma. In the natural environment, where resources are limited, the growth-defense trade-off means that plants cannot simultaneously exhibit high resistance and robust growth phenotypes. Nevertheless, there may be workarounds that allow for a reevaluation of strategies in agricultural production. Further elucidation of these trade-offs and potential bypasses could aid in the development of disease-resistant plants without compromising growth abilities[89].
-
Delving into the light-mediation of the JA-SA pathways is crucial for enhancing our comprehension of how plants balance defense and growth in response to fluctuating environments. This understanding provides a theoretical foundation for utilizing light to regulate crop development, reduce reliance on biocides, improve the quality of crops and ensure food safety. Light and JA/SA interactions modulate a delicate equilibrium for growth and defense. Additionally, these interactions highlight that light signaling is tightly regulated by numerous transcription factors, reflecting the evolutionary development of diverse regulatory pathways that enable plants to adapt to complex and dynamic environments.
Further investigation into the components of light signal transduction, including photoreceptors such as phytochromes phyB and phyA, cryptochromes (CRYs), genes involved in UV resistance such as UVR8, and light-responsive transcription factors (e.g. PIFs, HY5, FHY3/FAR1, BBXs, MYBs), could help identify key regulators in light-mediated survival and reproduction of plants within the JA/SA pathway. Understanding the involvement of JA and SA biosynthesis genes and pivotal players in signal transduction (JAZs, MYCs, and COI in the JA pathway as well as NPR1, ICS1, and WRKYs in the SA pathway) in light-mediated plant growth and defense is essential for a comprehensive grasp of how crops perceive and respond to environmental cues. Moreover, exploring the potential interactions between light/JA/SA with REACTIVE OXYGEN SPECIES (ROS), calcium signaling, and proteins that interact with light signaling components, such as the RECEPTOR-LIKE KINASE FERONIA (FER)[90] and CALCIUM-DEPENDENT PROTEIN KINASE (CPK)[91], warrants further research.
Integrating bioinformatic approaches to employ multi-omics data[92] such as transcriptomics, proteomics, metabolomics, and utilizing CRISPR/cas9 genome editing technology[93,94], will offer significant opportunities for bridging basic research with agricultural applications. Light quality, photoperiod, intensity, and duration modulate the defense-related hormones JA and SA through both biosynthesis and signaling, thus highlighting the importance of light condition alterations. Interestingly, under low R: FR conditions, evidence suggests that plants may forgo resistance to compete for light, indicating the operation of a sophisticated and precise energy allocation system. This natural phenomenon is in contrast to modern intensive cropping practices, making it essential to advance crop architecture breeding[31], light manipulation, and appropriate agronomic measures to reduce disease occurrence in intensive agricultural settings.
LEDs have emerged as an innovative and efficient light source. For example, employing LED-derived red light during the night[48] and managing greenhouse light environments can enhance resistance against biotrophic microbial pathogens in horticultural crops[74,95]. Future research focusing on the integration of light treatments combined with exogenous JA/SA applications may present new opportunities for the crop industry. Furthermore, investigating key genes in the light signal-mediated JA-SA pathway that enhance plant stress resistance could provide insights for breeding crops with superior stress resistance and growth qualities. Utilizing techniques such as the exogenous application of plant physiological active substances, plant growth regulators such as MeJA[70], SA[96], and light condition regulation are considered environmentally friendly and effective methods for improving the adaptability of vegetable crops to various adversities and to enhance the quality of vegetable products. Such advancements are essential in meeting the current and future demand for sustainable and healthy horticultural strategies.
-
The authors confirm their contribution to the paper as follows: literature collection: Hong J, Meng K; tables and models design: Hong J, Meng K; manuscript layout and format: Zhou Y, Kang H, Hong J, Meng K; draft manuscript preparation: Zhou Y, Kang H, Hong J, Meng K, Thomas HR, Yang Y, Williams B; manuscript modification and review: Zhou Y, Kang H, Thomas H, Yang Y, Williams B. All authors reviewed the results and approved the final version of the manuscript.
-
The data generated and analyzed during the current study are available from the corresponding authors upon reasonable request.
This work was supported by grants from the National Natural Science Foundation of China (32330094; U21A20233) and the Starry Night Science Fund of Zhejiang University Shanghai Institute for Advanced Study (SN-ZJU-SIAS-0011).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Jiachen Hong, Kaiwei Meng
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Hong J, Meng K, Thomas HR, Yang Y, Williams B, et al. 2024. Reframing agriculture by light: the role of light-mediated jasmonates/salicylic acid regulation in plant defense, development and beyond. Vegetable Research 4: e027 doi: 10.48130/vegres-0024-0026 

Reframing agriculture by light: the role of light-mediated jasmonates/salicylic acid regulation in plant defense, development and beyond
- Received: 25 April 2024
- Revised: 10 June 2024
- Accepted: 12 June 2024
- Published online: 09 August 2024
Abstract: Light serves not only as essential energy required for plant growth but as a signal, offering plants vital environmental information. Hormone signaling plays a pivotal role in the sophisticated resource allocation system which allows plants to adapt to ever-changing surroundings. Jasmonates (JA) and Salicylic acid (SA) are known as key defensive phytohormones in plants. Numerous physiological studies indicate light treatment impacts plant defense, growth, and development via pathways involving JA and SA. Recent molecular evidence suggests that photoreceptors and various transcription factors in phototransduction are implicated in the signaling of these defensive hormones. Thus, the growth-defense trade-off, which is modulated by light, is partially mediated through JA and SA signaling pathways. In this review, we highlight the recent advances toward understanding the interplay between light signaling and the JA/SA pathways. The mechanisms by which plants respond to biotic stress and abiotic stress in various species such as Arabidopsis, tomato, soybean, and cucumber are reviewed in depth. Furthermore, we discuss the potential opportunities to utilize these basic insights in practical agriculture applications, such as strategic manipulations of artificial lights for flexible and environmentally friendly approaches to enhancing crop growth and managing disease in greenhouses.
-
Key words:
- Light /
- Jasmonates /
- Salicylic acid /
- Plants defense /
- Biotic/abiotic stress