










-
Tea is a valuable economic crop in China that requires adequate fertilization to support its optimal growth and development[1]. Since the mid-twentieth century, the use of chemical fertilizers has significantly enhanced tea plantation production and profitability[2]. However, the expansion of tea gardens and subsequent increase in production have resulted in a significant escalation in fertilizer usage, leading to more severe consequences[3]. According to an estimate, approximately 30% of tea plantations in China are experiencing excessive use of chemical fertilizers, with 80% opting for compound fertilizers with an equal nutrient ratio, highlighting the increasing resilience of P fertilizers[1]. Studies have shown that soil P levels decrease under no P fertilization but increase after the application of P fertilizers[4]. However, excessive utilization of chemical fertilizers can result in soil compaction and environmental contamination[5]. On the other hand, organic fertilizers offer significant benefits to soil fertility by decreasing bulk density and enhancing soil texture[6]. Additionally, it is evident that the inclusion of organic fertilizer along with chemical sources improves crop quality, as observed in tea[7]. Some studies have also stated that the combined use of organic and compound fertilizers can lead to a substantial increase in soil total N, alkali-hydrolyzed N, available P, and available K[8]. Excessive active P in high-P conditions can accumulate in the soil[9], elevating P levels in the surface and lower layers[4]. This accumulation alters the soil's physical and chemical properties, thus affecting water and nutrient movement and transformation[10]. For tea plantations, it has been well reported that long-term excessive utilization of phosphate fertilizer does not effectively increase the soil's available P content. Instead, it leads to the inefficient use of fertilizer resources and exacerbates soil environmental pollution. Thus, alternative approaches to enhance fertilization use efficiency in tea plantation systems and the achievement of sustainable development goals are needed.
Phosphorus, an indispensable nutrient for plant growth and development[11], offer numerous crucial roles in metabolic processes, including photosynthesis, respiration[12], and mineral metabolism[13]. Many studies have shown that P application can increase antioxidant enzyme activities in P-deficient plants, thus protecting them from photooxidation damage[14]. Previous studies have also observed that adequate P application significantly enhances various the growth and development of various crops. Working with strawberries, Zheng et al. reported that adequate P supplementation can elevate plant height, petiole length, and leaf area while boosting the activities of antioxidative enzymes[15]. In rice, P fertilizer has been reported to improve grain quality traits, such as starch content[16]. Recently, Yan et al. documented improved wheat yield under P application, which was associated with high P accumulation and distribution in plant organs[17,18]. However, excessive P fertilizer can negatively affect plant growth and developmental processes. Higher P application results in fewer tillers, which, in turn, can harm crop yield in rice[19]. P application above the optimum level results in stunted root growth[20]. In addition, phosphorus deficiency in the soil can impact chlorophyll synthesis, hinder dry matter accumulation, and reduce sugar content in plant leaf[21]. Although the content of P is low in the soil of tea plantations, many physiological processes, such as photosynthesis and respiration, are highly dependent on P in tea plant. P affects the decomposition and metabolism of minerals and metabolites, thereby affecting the yield and quality of tea[22]. Both P-deficiency and excess reduced the syntheses of flavonoids and phosphorylated metabolites[13,23]. Thus, the judicious application of P at an agronomic optimum is urgently needed to promote plant performance and improve crop quality.
Given the current research status, although published studies have focused on the detrimental impacts of soil P deficiency on tea quality, further in-depth exploration is needed on the optimal application of P fertilizer in high-P environments, particularly when combined with organic fertilizers. Moreover, the effects of combined chemical and organic P application on soil properties, plant health, tea yield, and quality are yet to be fully explored. Here, we hypothesized that a low dose of chemical P, combined with an organic source, would be beneficial in improving soil properties, nutrient acquisition, growth, quality, and yield of tea. This study was conducted to determine the impact of varying levels of P fertilizer application, using both organic and inorganic sources, on soil physicochemical properties, enzymatic activities, plant growth, physiology, stress resistance ability, and quality of tea crop. This study would offer more precise guidance on P application for sustainable tea gardens. Additionally, it would also help prevent the negative outcomes of excessive or inadequate P application and achieve sustainable developmental goals for ecological restoration.
-
The experiment utilized the tea cultivar 'Baiye 1' and was conducted in Daping village, Shangnan County, Shangluo City, Shaanxi Province, China (Fig. 1). Fertilization experiments were carried out under suitable weather conditions. Soil samples were collected under the 0−20 cm layer. The soil was placed in a ventilated, dry place to avoid direct light and dried, and then the soil nutrient content and enzyme activity were analyzed through 16, 60, and 100 mesh screens[24]. Fresh tea leaves (including bud head, first leaf, and second leaf position) were collected and crushed with a plant crusher, and then vacuum freeze-dried for the determination of the biochemical quality of tea[9]. Hydroponic tea plants were maintained in the laboratory of Northwest A&F University in Shaanxi Province. Daytime and nighttime temperature of 25 °C/18 °C, with a photoperiod of 14 h of light and 10 h of darkness, and a relative humidity of 75% ± 5%.
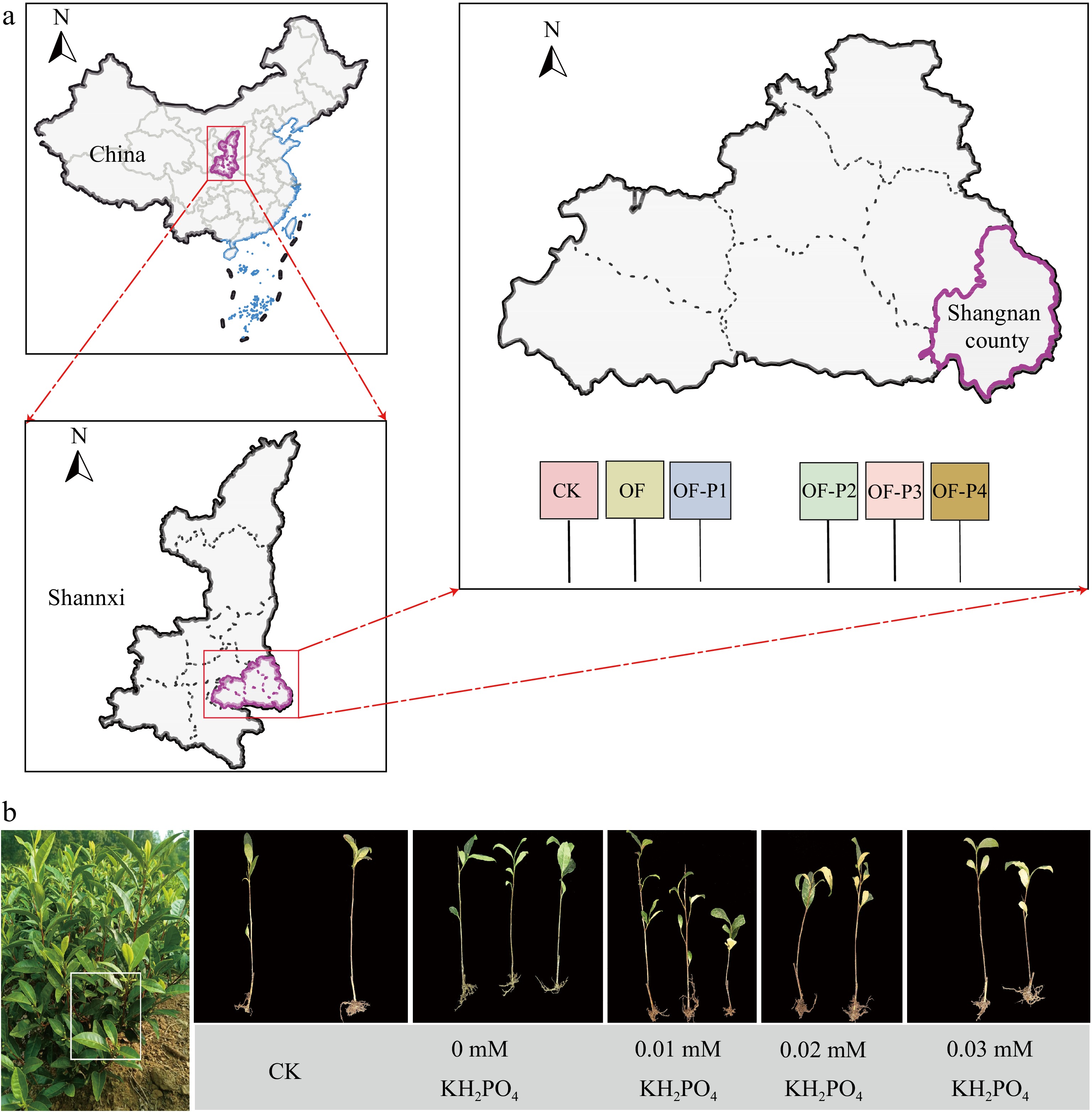
Figure 1.
(a) Location of the study area, and (b) test design site.
A randomized block design was used and different P management practices viz. unfertilized (control), sole organic fertilizer (OF) application, OF + N application at 300 kg/ha (N300) (OF-P1), OF + N300 + P application at 45 kg/ha (OF-P2), OF + N300 + P application at 90 kg/ha (OF-P3), and OF + N300 + P application at 135 kg/ha (OF-P4) were maintained. Each treatment was replicated three times. Each plot size had a 25 m2 area. The application rate for organic fertilizer was consistent across treatments at organic fertilizer 7,500 kg/ha. Sixty percent of the N fertilizer was applied as a basal dressing, with the remaining 40% applied as topdressing. Phosphorus, P fertilizer, and organic fertilizer were combined and applied as basal dressing.
Changes in hydroponic root growth of tea plant
-
Tea plants were initially cultured in deionized water for 3 d, followed by 1/2 nutrient solution for 1 week, and then transitioned to full strength nutrient solution for further cultivation. The concentrations of KH2PO4 and K2SO4 in the medium were adjusted to create P concentration gradients of 0, 0.01, 0.02, and 0.03 mM according to Wang's method in two tea cultivars 'Longjing 43' and 'Shuchazao'[25]. The nutrient solution was refreshed weekly. Photographs of the tea seedling were taken and stored for subsequent observation of root development[26].
Determination of physical and chemical properties and enzyme activity of soil in tea plantation
-
Soil moisture content and bulk density were determined using the drying method[27]. Total porosity was calculated based on bulk density. Total phosphorus and total potassium contents in the soil were determined using resistance colorimetry and flame spectrophotometry, respectively. Total nitrogen content was analyzed using a continuous flow analyzer[28]. Available phosphorus content in the soil was determined using the hydrochloric acid-ammonium fluoride method[29]. Trace elements aluminum and calcium were determined using inductively coupled plasma mass spectrometry. Acid phosphatase activity was assessed using p-nitrobenzene disodium phosphate colorimetry, while phytase activity was determined using the molybdenum blue method. Phosphomonesterase activity was measured using the p-nitrobenzene phosphate method.
Determination of chlorophyll and tea quality-related content
-
The relative chlorophyll content of leaves was determined using the ethanol extraction method. Chlorophyll fluorescence was observed using the hexagon imaging PAM (WALZ)[30]. The contents of water extract, tea polyphenol, free amino acid, and caffeine in the fresh parts of tea tree (bud head, first leaf, and second leaf) were determined, with five replicates for each sample. The content of water extract was determined by boiling water reflux method. The content of tea polyphenols was determined by the ferrous tartrate colorimetric method. The content of free amino acids was determined by ninhydrin colorimetry. Caffeine is easily soluble in water, first extracted by boiling water reflux, and then determined after removing the interfering substances[31].
Determination of reactive oxygen species and antioxidant enzymes
-
The determination of superoxide anion contents followed the methods outlined by Tan et al.[32] and Sui et al.[33]. Hydrogen peroxide contents were estimated using the method described by Moloi & van der Westhuizen[34]. Additionally, antioxidant enzyme activities were determined using the modified method as outlines by Chen et al.[35].
Statistical analysis
-
All data was processed using Origin 2021 and SPSS Statistics 25.0. All drawings were made using GraphPad prism 8.0 and the chiplot website (
www.chiplot.online ), including principal component analysis and correlation analysis. All data were measured three times under each treatment. -
Soil water content and porosity increased first and then decreased with increasing P rates. However, these treatments showed no significant effect (p > 0.05) on soil bulk density. Compared with CK, all treatments increased soil water retention capacity; however, the maximum values were recorded for OF-P2. A similar trend was observed for soil total N, total P, and total K, where values increased initially and then decreased with increasing P rates. The maximum values were recorded for the OF-P2 treatment, which increased total N, total P, and total K content by 30.5%, 45.2%, and 1.6%, respectively, over the control. There was no significant (p > 0.05) change in available P content between the unfertilized control and sole organic fertilizer (OF) application. Additionally, OF-P2 increased available aluminum content by 45.6% higher than the control, whereas there was a significant decrease in exchangeable calcium content under the same treatment. The activity of phosphatase in the soil directly influences the conversion of organic P and enhances the absorption of inorganic P. There were significant differences in phosphatase activity among different treatments, with maximum values recorded for the OF-P2 treatment, whereas the CK treatment depicted significantly lower values than other treatments. Phytase activity increased significantly with increasing P rate. Compared with the CK treatment, the effect of low P on soil enzyme activity was small. There was no significant difference in phosphomonesterase activity among the treatments (Supplemental Fig. S1). Correlation analysis showed that total soil P had a significant effect on water content, bulk density, and total porosity (p < 0.01) (Fig. 2), whereas it had a highly significant effect on soil aluminum content (p < 0.001). The content of inorganic P in the soil was affected by available P and available K (p < 0.05). Linear analysis of soil phosphatase activity and total P and available P contents showed that the correlation reached 0.5395 and 0.6057, respectively (Fig. 2b−e). Soil phytase activity was linearly correlated with total P and available P contents, reaching 0.4002 and 0.523. The results of soil physical and chemical properties of tea garden with different P application rates showed that the suitable P rate could accelerate the decomposition of P and enhance the absorption and utilization of soil nutrients by tea seedlings.
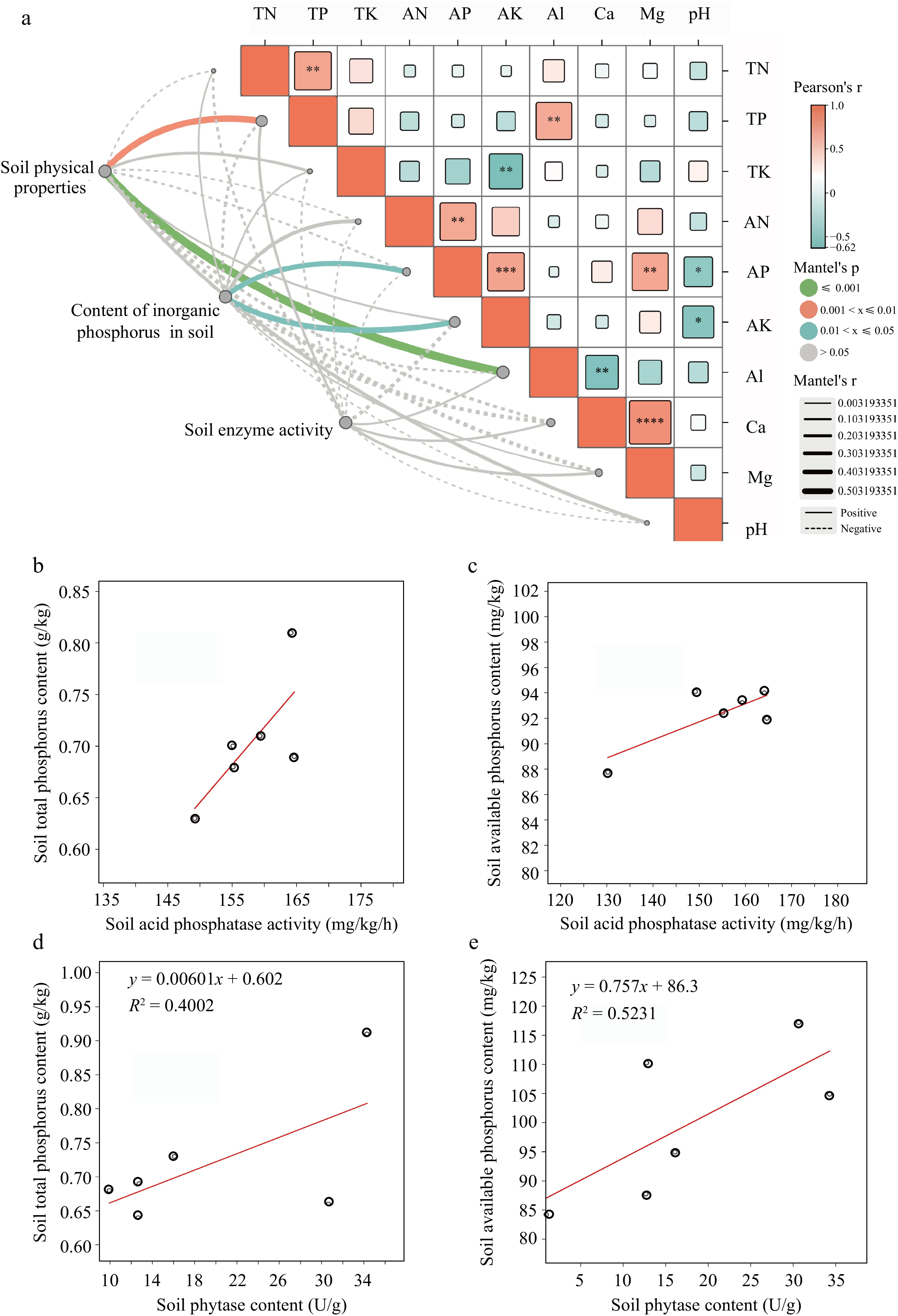
Figure 2.
(a) Correlation analysis among soil physical and chemical properties. (b) Linear relationship between acid phosphatase and total phosphorus, (c) acid phosphatase and available phosphorus, (d) phytase and total phosphorus, (e) phytase and available phosphorus. TN: total nitrogen, TP: total phosphorus, TK: total potassium, AN: available nitrogen, AP: available phosphorus, AK: available potassium. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
Effects of different phosphorus application rates on 100-bud weight and bud density of tea plant
-
The P management treatments significantly affected the 100-bud weight of one bud (Fig 3a), with the OF-P2 showing significantly (p < 0.05) higher values (15.3% higher than the CK) than other treatments. For the 100-bud weight of one bud, the treatments were ordered as OF-P2 > OF-P3 > OF-P1 > CK > OF > OF-P4. Bud and leaf density also differed significantly among the treatments, with OF-P1, OF-P2, OF-P3, and OF-P4 demonstrating an increase of 35.4%, 50.2%, 48.3%, and 44.2%, respectively, over the CK. For bud density per 0.04 m2, these treatments were ordered as OF-P2 > OF-P3 > OF-P4 > OF-P1 > OF > CK. Overall, OF-P2 outperformed the other treatments in improving bud density per 0.04 m2 and 100-bud weight of one bud.
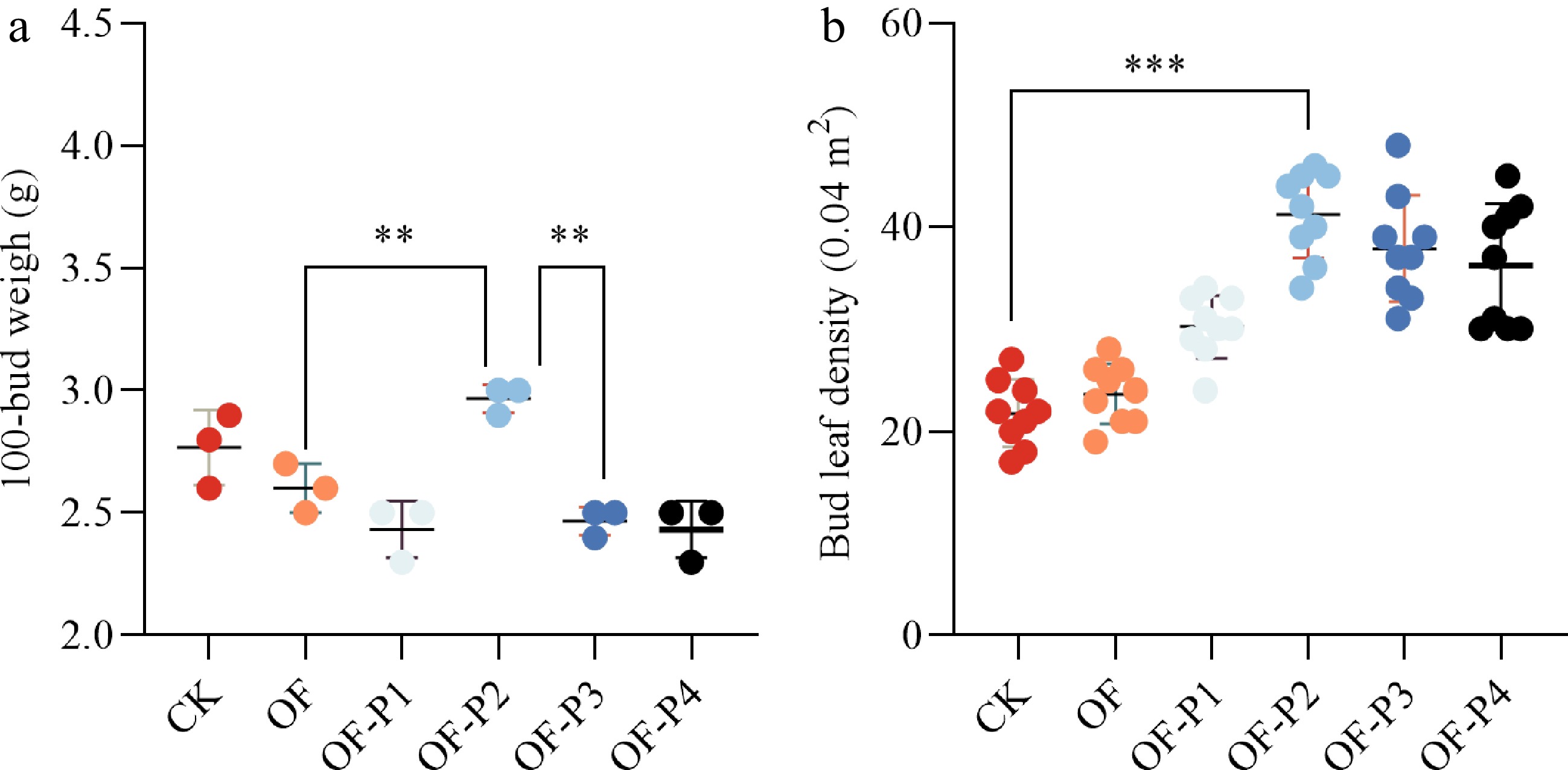
Figure 3.
(a) Effect of phosphorus based on organic fertilizer on 100-bud weight of one bud of tea plant and (b) bud density per 0.04 m2. Different * indicates the level of difference between treatments: * p < 0.05, ** p < 0.01, *** p < 0.001.
Effects of different P application rates on root growth
-
Different P rates had a significant effect on the root growth of tea (Fig. 4). Compared with the treatment without P (0 mM KH2PO4), the root length of tea trees treated with 0.01 mM KH2PO4 was significantly increased and reached the maximum for a P concentration of 0.01 mM KH2PO4. These hydroponics results showed that low P promoted root growth, whereas high rates showed inhibitory effects on plant roots.

Figure 4.
Effect of phosphorus on root growth in Camellia sinensis. (a) Root phenotype before phosphorus treatment, (b) root phenotype after phosphorus treatment.
Effect of different amounts of phosphorus application on fresh tea leaves quality
-
There were significant changes in the contents of polyphenols and free amino acids under different treatments (p < 0.05) (Fig. 5a & b), but the effect on water extract and caffeine content was nonsignificant (p > 0.05) (Fig. 5c & d). The polyphenol content of tea in different organs showed a decreasing trend with increasing P rate, with maximum reduction recorded for the OF-P2 treatment, which decreased the values by 42.3%, 45.6%, and 25.7% in the bud, first leaf, and second leaf, respectively, compared to the CK. The changes in polyphenol content in tea under different organs were as follows: OF-P2 > OF-P1 > OF-P3 > OF > CK > OF-P4. An opposite trend was observed for free amino acids, where values increased with increasing P rates. The OF-P2 treatment recorded significantly higher values by 33.5%, 40.1%, and 31.4% in the bud, first leaf, and second leaf, respectively, compared to the CK. For free amino acid contents, the treatments were arranged as OF-P2 > OF-P1 > OF-P3 > OF-P4 > CK > OF.
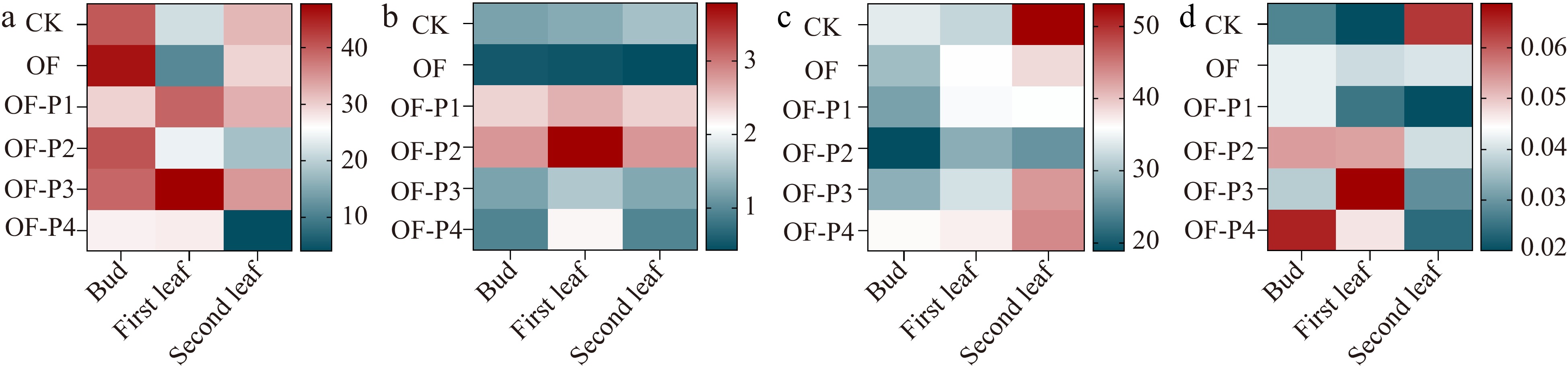
Figure 5.
Effect of phosphorus based on organic fertilizer on metabolites of different leaf positions in C. sinensis. (a) Water extract, (b) free amino acid, (c) total polyphenols, (d) caffeine.
Effects of different phosphorus application rates on chlorophyll content
-
There were significant differences in chlorophyll (Chl) content among different treatments. The Chl a, b, and carotenoid contents under all treatments were significantly higher than those in the control (CK) (Fig. 6a). The Chl content of the first leaf increased first and then decreased with increasing P rates, reaching the maximum values in the OF-P3 treatment (Supplemental Fig. S2). The Chl content in the second leaf followed the same trend as the first leaf and bud. A similar trend was observed for chlorophyll fluorescence. Correlation analysis showed that Chl content in the bud exhibited a trend of first increasing and then decreasing with increasing P rates (Fig. 6b). However, the relationship between Chl content and soil P content was not significant for the first leaf and second leaf (Fig. 6c & d). The results indicated that the bud head was more responsive to P content compared to the first-leaf and second-leaf. Overall, the Chl content after P application was higher than that of the CK, and the Chl content under low P conditions was higher than that under high P conditions.
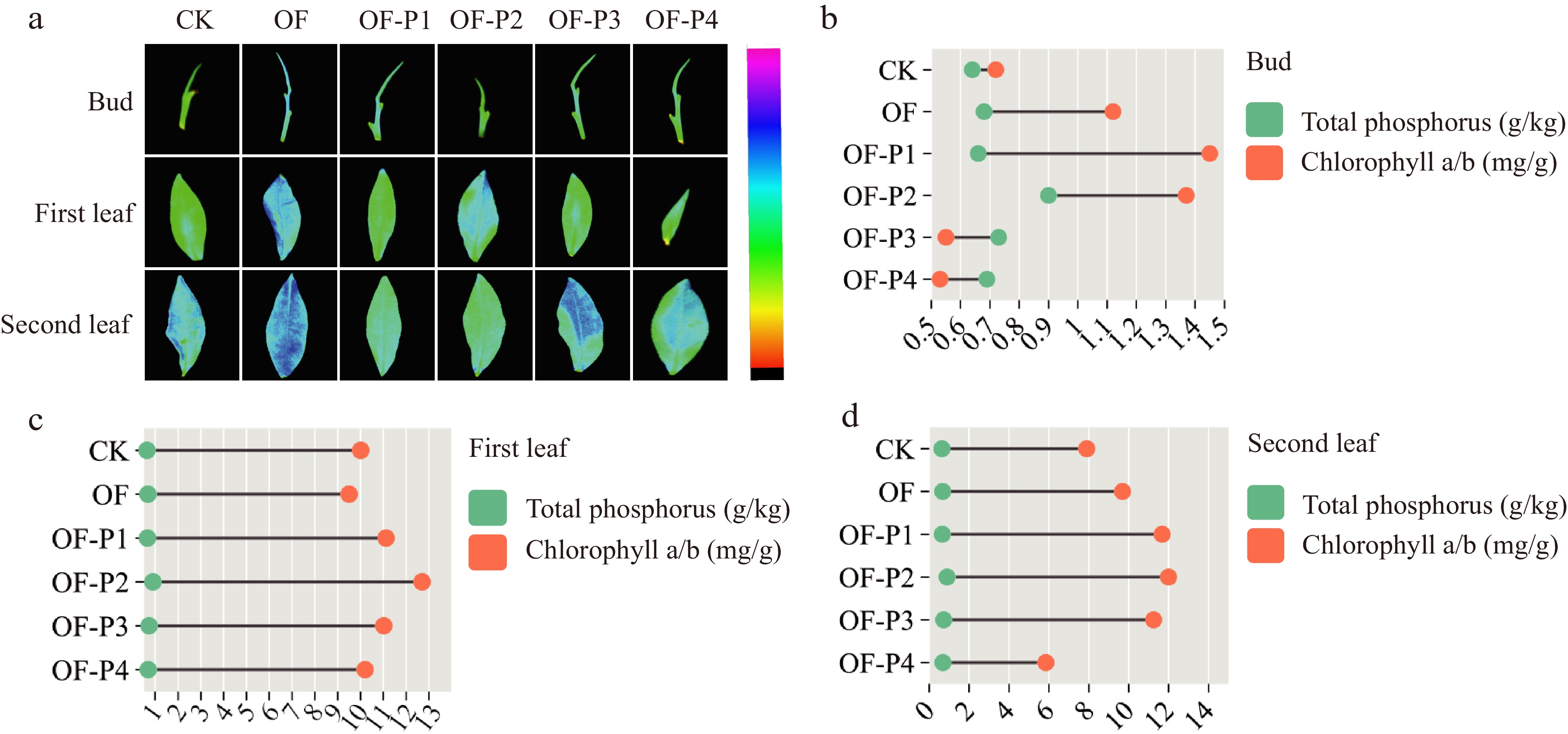
Figure 6.
(a) Effect of phosphorus based on organic fertilizer on chlorophyll content in C. sinensis chlorophyll fluorescence of different leaf positions in C. sinensis. Relationship between total phosphorus and chlorophyll a/b content on (b) bud, (c) first leaf and (d) second leaf in C. sinensis.
Effects of different phosphorus application rates on reactive oxygen species and antioxidant enzymes
-
Compared with the control, P treatments significantly decreased the H2O2 content. In the bud and second leaf, the H2O2 contents first decreased and then increased with increasing P rates (Fig. 7a−c). The lowest values were recorded for the OF-P2 treatment. The results of DAB staining of the first leaf (Fig. 7d) showed that damage caused by applying P fertilizer to the tea plant was minimal. Compared with the CK, the O2− content in the first and second leaf decreased under different P fertilizer treatments. In the bud, first and second leaf, O2− content showed a decreasing trend with increasing P rates. The maximum reduction was recorded for the OF-P2 treatment, which decreased the values by 50.1%, 42.8%, and 14.3% in the bud, first and second leaf, respectively, lower than the control (Fig. 7e−g). A leaf stained with NBT also showed the same trend (Fig. 7h).
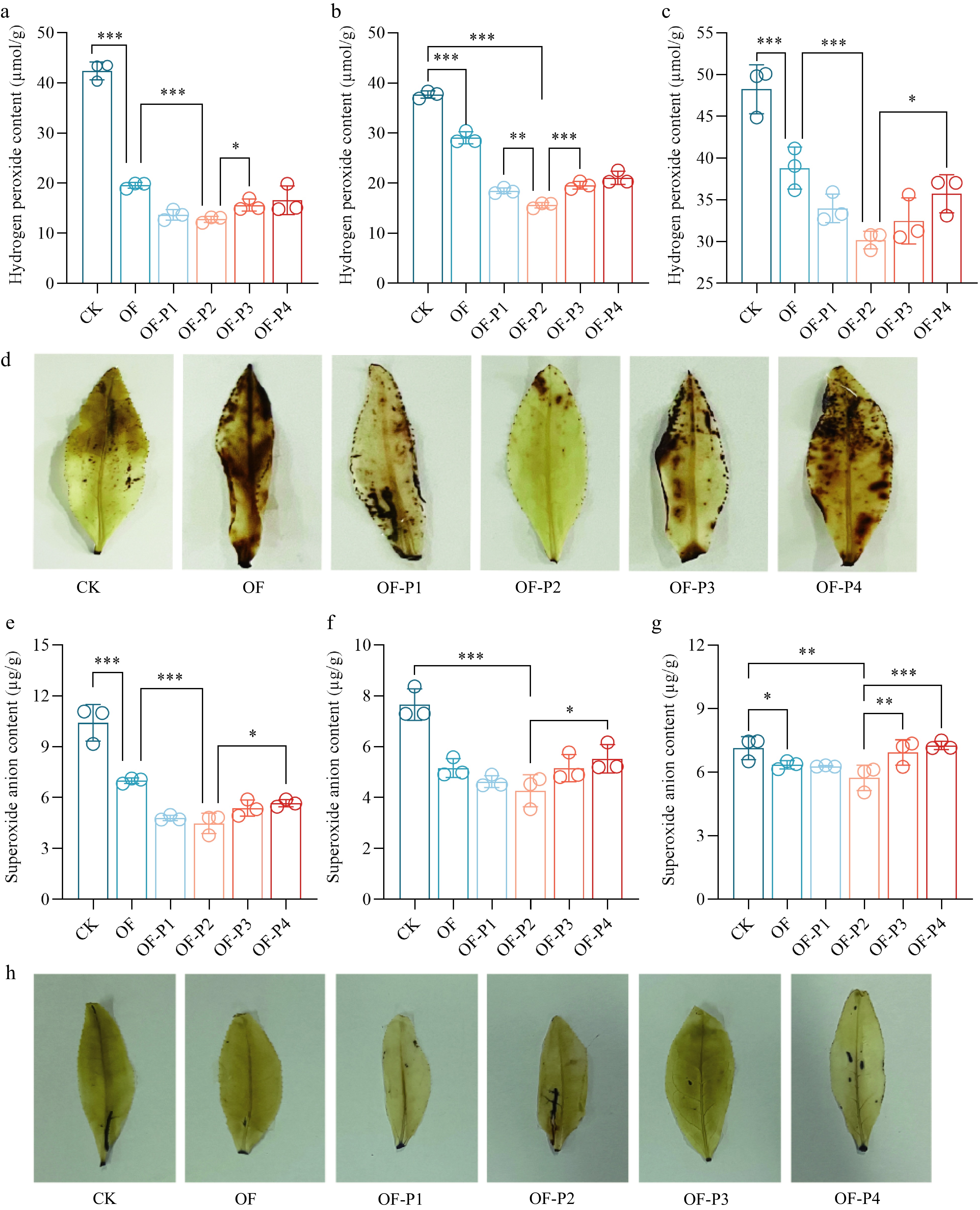
Figure 7.
Effect of phosphorus based on organic fertilizer on hydrogen peroxide and superoxide anion contents in C. sinensis. hydrogen peroxide content on (a) bud, (b) first leaf, and (c) second leaf, (d) DAB staining in first leaf, (e) superoxide anion content on bud, (f) first leaf, and (g) second leaf, (h) NBT staining in first leaf. Different * indicates the level of difference between treatments: * p < 0.05, ** p < 0.01, *** p < 0.001.
The experimental results showed that, except for the CK, the SOD activity in different leaves was significantly increased under different P amendments (Fig. 8). Maximum values for SOD were recorded for the OF-P2 treatment. POD activity in different leaves increased initially and then decreased with increasing P rates. Compared with the CK, low P treatment significantly increased POD activity compared to the higher rates. APX activity was significantly increased under different treatments compared with the control (p < 0.05). Maximum CAT activity in the first and second leaf was recorded for the OF-P2 treatment. However, the difference between the first and second leaf was not significant for CAT activity. The OF-P2 treatment increased CAT activity in the bud head, first leaf, and second leaf by 16.6%, 23.5%, and 22.9%, respectively, higher than the CK. Further PCA analysis showed that the control and OF-P2 were significantly different in the three different leaves (p < 0.05).
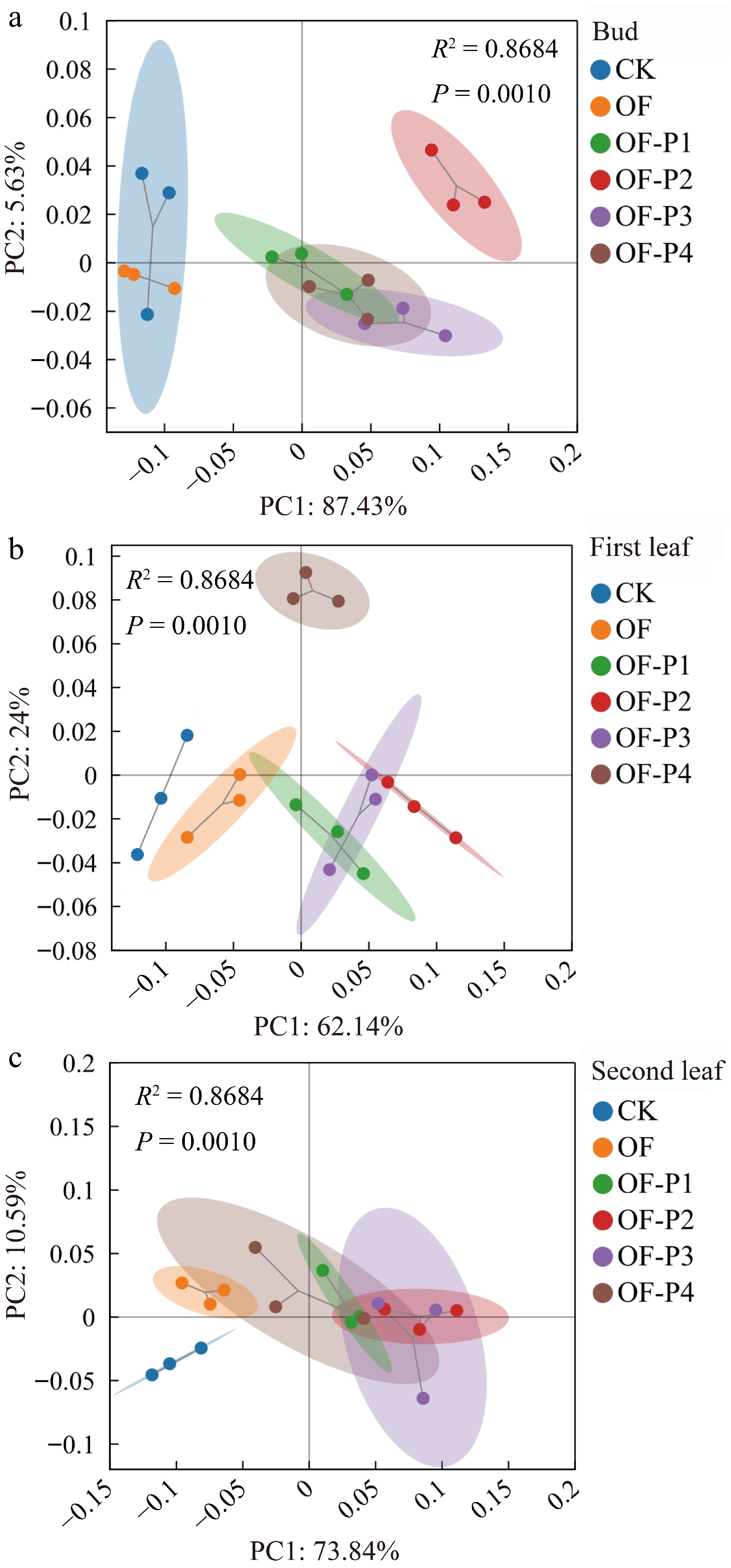
Figure 8.
Principal components analysis of phosphorus based on organic fertilizer on antioxidant enzymes of different leaf positions in C. sinensis.
-
The use of organic fertilizer can significantly stimulate and increase soil P availability[36]. The present findings demonstrated that combining organic fertilizer with low P increases total N, P, and K in the soil. These findings are in line with the research by Vu et al., which reported a positive effect of organic fertilizer on the availability of soil P[37]. Moreover, in this study, adding low P also increased aluminum levels in the soil, which demonstrated a significant improvement in aluminum availability. We also noted that low P facilitated soil enzymatic activity, including acid phosphatase and phytase, aiding in organophosphorus breakdown and enhancing plants' P absorption. Additionally, the combined application of chemical and organic P was also reported to enhance phosphatase activity, speed up the transformation of soil P, and increase soil available P and nutrient contents. Excessive P application wastes soil P resources and reduces the availability of available P, as reported previously[38]. The combined use of organic fertilizer and low chemical P markedly enhanced tea bud weight, bud density per 0.04 m2, and promoted root growth. Similar results were reported by Kirchgesser and colleagues, who demonstrated that P deficiency leads to smaller root systems, whereas adequate P application substantially improved crop performance[39]. In contrast to this study, some authors have also reported that high P inhibits the root system, possibly due to Polygonum's strong P tolerance compared to weak-tolerant crop species, including tea. In another study, similar results were reported, where peanut exhibited increased total root length, root volume, and surface area under appropriate P fertilization[18]. Furthermore, Chen and colleagues also revealed that optimal P levels significantly improved tea root vitality, while excess P had inhibitory effects[40]. The positive influences of combined organic fertilizer and chemical P on tea quality are well-documented. P deficiency results in decreased tea water extract, free amino acids, and caffeine contents[14]. Meanwhile, a sufficient supply of P has been reported to improve quality traits, for example in peanut[18]. In conclusion, combined organic and chemical P fertilizer in appropriate amounts guarantees meeting the tea plants' growth requirements and optimizing tea quality characteristics.
Under low P conditions, chlorophyll content in tea plants significantly increased compared to the control and other treatment groups. Simultaneously, heightened antioxidant enzyme activity assisted in eliminating excess free radicals in tea plants, leading to decreased hydrogen peroxide and superoxide anion levels, thereby improving plant resistance. These results align with previous studies where phosphate fertilizer application initially boosted chlorophyll content and antioxidant enzyme activity, followed by a decline[41]. Examination of chlorophyll and fluorescence parameters in Chinese cabbage under varied P levels revealed that with P fertilizer addition, the content initially stabilized, then peaked, and eventually decreased[42]. In the study of the combined application of P fertilizer on citrus, it was found that a high concentration of P would reduce the antioxidant capacity of leaves and roots, whereas leaves and roots have a stronger antioxidant capacity when P is low. Proper concentration of phosphorus can improve the physiological characteristics of citrus[43].
Therefore, it is crucial to implement effective strategies to reduce P inputs and minimize environmental losses comprehensively[9]. The present research has revealed that maintaining a low P level can enhance the available P content in the soil of tea plantation. This enhancement boosts the activity of phosphatase and phytase, thereby improving phosphorus uptake by the tea plants. In situations of low P levels, the growth of tea plant roots are stimulated, leading to an increased 100-bud weight of one bud and bud density per 0.04 m2, reduced polyphenol content in the tea, higher levels of free amino acids, and an overall improvement in tea quality. Furthermore, the increase in antioxidant enzyme activity plays a vital role in combating the adverse effects of reactive oxygen species production in plants, ultimately strengthening the resilience of tea plants.
-
The study explored the relationship between soil nutrient status and above-ground performance of tea seedlings under different P fertilizer rates from both organic and inorganic sources. Results showed that low P application (45 kg/ha) improved soil nutrient levels, leading to significant improvements in the 100-bud weight of one bud, bud density per 0.04 m2, plant resistance, and overall tea quality. This research provides valuable insights into the proper use of P fertilizers in southern Shaanxi, emphasizing the importance of determining the adequate dosage for optimal fertilizer application.
-
The authors confirm contribution to the paper as follows: study conception and design: Bai J, Wang Y, Shi R, Deng S, Wang H, Wang C, Gong C; experiments performed: Wang Y, Shi R, Deng S, Wang H; data analysis: Wang Y, Shi R; draft manuscript preparation: Wang Y, Bai J, Gong C, Hussain S. All authors reviewed the results and approved the final version of the manuscript.
-
Due to administrative requirements, the original data of the experiments during the research period of the project are not available to the public, but available from the corresponding author or the first author upon reasonable request.
This research was supported by the Key Research and Development Project of Shaanxi Province (No: 2022NY-167).
-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Effect of phosphorus based on organic fertilize on basic physical and chemical properties of tea plantation soil. water content (a), bulk density (b), total porosity (c), total nitrogen (d), total phosphorus (e), total potassium (f), available phosphorus (g), available aluminum (h), exchangeable calcium (i), soil acid phosphatase activity (j), phytase activity (k), phosphomonesterase activity contents (l).
- Supplemental Fig. S2 Effect of phosphorus based on organic fertilize on chlorophyll content in different leaf positions in Camellia sinensis. Chlorophyll a (a), chlorophyll b (b), carotenoid content (c) in bud, chlorophyll a (d), chlorophyll b (e), carotenoid content (f) in first leaf, chlorophyll a (g), chlorophyll b (h), carotenoid content (i) in second leaf.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Y, Shi R, Deng S, Wang H, Hussain S, et al. 2024. Application of low phosphorus on the basis of organic fertilizer can effectively improve yield and quality of tea plants. Beverage Plant Research 4: e037 doi: 10.48130/bpr-0024-0027 

Application of low phosphorus on the basis of organic fertilizer can effectively improve yield and quality of tea plants
- Received: 12 May 2024
- Revised: 22 June 2024
- Accepted: 01 July 2024
- Published online: 10 October 2024
Abstract: The availability of soil phosphorus (P) is essential for crop cultivation and production. However, agronomic P management for tea crops remains unexplored. Herein, the effect of different P management practices viz. unfertilized (control), organic fertilizer (OF) application, OF + N application at 300 kg/ha (N300) (OF-P1), OF + N300 + P application at 45 kg/ha (OF-P2), OF + N300 + P application at 90 kg/ha (OF-P3), and OF + N300 + P application at 135 kg/ha (OF-P4) on soil nutrient acquisition, enzymatic activities, and physio-biochemical, and quality traits of tea plants are investigated in yield. The results showed that OF-P2 treatment had significantly higher soil N (30.5%), P (42.2%), and potash (1.6%) concentrations above the control. P concentrations had a linear positive correlation with the activities of acid-phosphatase and phytase. OF-P2 had the greatest effects on plant growth, chlorophyll and carotenoid contents, and antioxidative enzyme activities than other treatments. OF-P2 treatment had a two-fold decrease in hydrogen peroxide and dioxygen (singlet) compared to the control. It was further found that OF-P2 significantly increased amino acid content by 33.5%, 40.1%, and 31.9%, and decreased polyphenol content by 42.3%, 45.6%, and 25.7% in bud, first, and second leaf, respectively, above the control. Overall, the present findings suggest that low P application (OF-P2) can increase nutrient availability, bud quality, and yield by improving soil enzymatic activities, pigment contents, and antioxidative activities. Establishing this mode of low P application may provide an optimum strategy for enhancing crop performance in regions where unreasonable P application practices are common.
-
Key words:
- Camellia sinensis /
- Phosphate fertilizer /
- Soil properties /
- Tea quality