








-
In 2022, the tomato cultivated area in China reached 1.1466 million hm² (FAOSTAT, 2024) (
www.fao.org/faostat ). The total amount of tomato residues that can be used as resources reached 19.37 million tons. At present, the main treatment methods for vegetable residues in China include discarding them as waste, burning them in the field, returning them to the field after composting, or directly crushing them and returning them to the field[1,2]. Although incineration or disposal saves time and labor, it may result in serious environmental pollution and lead to the wasting of agricultural resources[3,4]. The compost facility for straw treatment is simple and cheap, but the transportation cost is high[5]. Return field directly in situ, including the crushing of vegetable residues and returning the residues to the field, which efficiently dissolves a large amount of vegetable residues[6]. On a dry substance basis, vegetable residues contain abundant mineral elements needed for plant growth, with a C/N ratio of approximately 7.0~21.3, with N, P, and K contents of approximately 1.2%~3.1%, 1.2%~1.9% and 3.0%~3.9%, respectively[7]. The contents of Ca, Me, Fe, Be, and Zn among the trace elements are approximately 1.6%~8.7%, the content of organic substances is approximately 42.8%~67.4%[8], the contents of sugar and cellulose are as high as 75% and the contents of hemicellulose and lignin are approximately 9% and 5%, respectively[9]. How to effectively utilize the remaining large amount of vegetable residues is a problem that should be explored.To date, China accounts for 7% of the global arable land but uses more than 33% of chemical fertilizers (FAOSTAT, 2022). The extensive use of chemical fertilizers not only result in high cost but also poses serious harm to the ecological environment[10]. The input of organic materials into the tillage system is conducive to the stimulation of soil microbial activity, the increase in soil microbial abundance and the number of flora, and the promotion of the recycling of mineral elements in the soil[11−13]. Long-term straw return with chemical fertilizers effectively increases the contents of soil organic carbon, available phosphorus, and total nitrogen[14], which can improve the C/N ratio of soil, promote the decomposition of organic carbon, and effectively increase the soil nutrition content of the tillage layer, thus increasing the yield[15,16]. Vegetable residues contain a large amount of mineral elements and organic substances, which can be used for the transformation of mineral elements after direct return to the field, thereby reducing the amount of chemical fertilizers needed and promoting organic growth of plants[17]. Because most countries have shifted from a high-speed development model to a high-quality development model, the sustainable development of agriculture and the efficient use of resources are particularly important. However, it is unknown what kind of consequences will be caused by continuing to plant tomatoes after tomato residues are returned to the field.
Bacillus subtilis is an environmentally friendly and efficient broad-spectrum biocontrol bacterium. It not only can secrete antibacterial substances in the host and induce systemic resistance in plants[18,19], but directly antagonize microbial pathogens by competing for space and nutrients and effectively suppress a variety of plant diseases[20]. Additionally, it also has the natural potential to treat soil pollutants to reduce the use concentrations and residues of agrochemicals in farmland[21,22]. It was found that the synergistic effect of B. subtilis NCD-2 and broccoli residues could increase potato yield by 32% compared to the untreated control[23]. Combined application of B. cereus and corn stover increased soil microbial diversity and soil organic matter content and promoted the growth of Begonia seedlings[24]. Of course, the synergy effect of applying B. subtilis and tomato residues to the soil on the continuous cultivation of tomatoes is unknown.
Based on the above problems and research status, in this experiment, the application of B. subtilis in the soil with tomato residues returned to the field were combined to study the effect on soil physicochemical properties, enzyme activity, soil bacterial community structure, and tomato yield and quality to provide a theoretical basis and technical guidance for tomato plant residue resource utilization and the sustainable cultivation of facility tomatoes.
-
The field experiment was carried out in the research farm of Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing, China (latitude 39°57' N, longitude 116°19' E and altitude 50 m above sea level). The tomato variety 'Galaguo' was used, tomato plant residues were taken from the last tomato plants after tomato harvesting in the solar greenhouse the previous year. The residue nutrients were measured as follows: dry substance (14.06%), total nitrogen (30.20 g·kg−1), total phosphorus (7.10 g·kg−1) and total potassium (48.50 g·kg−1). The effective viable count of the completely water-soluble B. subtilis dry powder used was 10 × 1010 g−1. Additionally, the physicochemical properties of the original surface soil (0~20 cm) in the solar greenhouse were as follows: bulk density 1.31 g·cm−3, organic matter 1.88%, pH 7.11, alkaline hydrolysis nitrogen 172.67 mg·kg−1, available phosphorus 76.34 mg·kg−1, and available potassium 210 mg·kg−1.
Experimental design
-
Before planting tomato, 9,000 kg·hm−2 of organic fertilizer (N, P, and K contents of 5.11, 7.25, and 22.21 g·kg−1, respectively) and 450 kg·hm−2 ternary compound fertilizer (N : P : K = 15:15:15) were applied in advance. After mixing evenly, the tomato residue and B. subtilis were applied to the soil as treatments. To investigate the effects of tomato residues and B. subtilis on the soil environment, the following six treatments were used: 1. control (CK), 2. soil add tomato residues (TC), 3. soil add B. subtilis (BS), 4. soil add tomato residues and B. subtilis (TC-BS), 5. double tomato residues (2TC), and 6. double tomato residues and B. subtilis (2TC-BS). Each treatment had five replicated plots (each plot about 8.4 m2). The treatment amount of tomato residue was 24,000 kg·hm−2 (TC) or 48,000 kg·hm−2 (2TC) which were covered thinly with soil, 225 kg·hm−2 B. subtilis was evenly applied in the relevant treatments. Other greenhouse management practices were performed according to conventional planting management methods.
The collection of samples was one day before the tomato plant uprooting, six soil samplings for each treatment were taken with soil sampling at five points, and the soil of 0~20 cm depth near the stem base of the plant was taken. After mixing, the soil was divided into four parts by the quartering method. One sample was stored in a −80 °C freezer as soon as possible for DNA extraction and sequencing. The remaining soil was placed in a cool place to air dry naturally for the determination of soil nutrient content and enzyme activity.
When the third truss tomato ripened, nine tomato fruits were selected from each plot for fruit measurement and nutrient quality analysis. The horizontal and vertical diameter and individual fruit weight of the tomato fruit were first measured, then the hardness of the fruit was measured. Cutting off 1/6 of the fruit for measuring the content of soluble solids and the remaining samples were ground using a wall breaking machine to determine the quality of tomatoes.
Determination of the growth, physiological index, quality, and yield of tomato
-
During the fruit-setting period, the plant height (the distance from the base of the stem to the growing point) was measured using a tape measure and the stem thickness at approximately 1 cm above the ground was measured using a vernier caliper. Chlorophyll was determined by the 95% ethanol extraction method[25]. The net photosynthetic rate (Pn) of functional leaves (the leaves opposite the first panicle and fruit) was measured using a Li-6400 portable photosynthetic analyzer (LI-COR, USA).
The content of lycopene was determined by the methanol extraction colorimetric method[26]. The content of vitamin C (VC), soluble sugar, titratable acid, and soluble protein were determined by the xylene extraction colorimetric method, the antrone colorimetric method, the acid-base titration, and the Coomassie brilliant blue G-250 staining method, respectively[25]. The content of soluble solids was measured using a hand-held sugar meter (PAL-1, ATAGO Corporation, Japan). The fruit hardness was measured using a fruit hardness tester (GY-3). The individual yield of three rows of tomato were randomly selected for each treatment, which was converted to the yield per mu to calculate the total yield.
Determination of soil nutrient content, and enzyme activity
-
Soil bulk density and porosity were measured using a three-phase soil analyzer (DIK-1150, Daiki Rika Kogyo Co., Saitama, Japan)[27]. The EC value was measured using a conductivity meter (ISPRING, TDS-Ph-EC). The measurment of organic substance, total nitrogen, and alkaline hydrolysis nitrogen were carried out using the dichromate oxidation-external heating method, the semimicro Kjeldahl method, and the alkaline hydrolysis-diffusion method, respectively[28]. The total carbon was measured by an elemental analyzer (Vario Macro, Elementar Analysensysteme GnbH, Germany).
Soil sucrase and cellulase activity were determined using the 3,5-dinitrosalicylic acid colorimetric method, while the soil-neutral phosphatase activity was determined using the disodium phosphate colorimetric method and soil urease activity was determined using the indophenol blue colorimetric method[29].
DNA extraction and 16S rDNA sequencing in soil
-
Total microbial genomic DNA was extracted using the DNeasy PowerSoil kit according to the manufacturer's instructions. The concentration and quality of the extracted DNA were checked using a NanoDrop ND-1000 spectrophotometer and agarose gel electrophoresis, respectively. The extracted DNA was used as a template to amplify soil bacterial sequences and the V3-V4 regions of the bacterial 16S rDNA gene were amplified by PCR using the forward primer 338-F (5'-ACTCCTACGGGAGGCAGCA-3') and the reverse primer 806-R (5'-GGACTACHVGGGTWTCTAAT-3'). The amplification products were purified using Vazyme vhtstm DNA cleaning beads (Vazyme, Nanjing, China) and quantified using the Quant-iT PicoGreen dsDNA kit (Invitrogen, Carlsbad, CA, USA). Sequencing was performed on the Illumina NovaSeq platform using the NovaSeq-PE250 by Shanghai Passeno Biotechnology Co., Ltd (Shanghai, China).
Bioinformatics analysis
-
High-quality (> 97% similarity level) sequences were analyzed using the Parsons Gene Cloud platform (
www.genescloud.cn/home ) for subsequent analysis. QIIME 2 software was used for taxonomic annotation and composition analysis of the obtained sequences. Chao1, Shannon and Simpson index was calculated using the OTU table[30].Data analysis
-
Microsoft Excel 2019 was used to collate the data. DPS 18.0 was used to perform analysis of variance (ANOVA). Multiple comparisons among different treatments were performed using Duncan's new multiple range method (α = 0.05). GraphPad Prism 8.4.2 and Adobe Illustrator 2019 were used for the graphs.
-
To explore the effects of returning tomato residues to the field combined with the application of B. subtilis on soil production, the effects of the different treatments were first analyzed on the soil physicochemical properties (Fig. 1a−d) and enzyme activities (Fig. 1e−h). Compared with CK, the 2TC-BS treatment significantly reduced the soil bulk density (Fig. 1a) and with a gradual decrease with increasing amounts of tomato residues. While the TC-BS and 2TC-BS treatments significantly increased the total soil pores (Fig. 1 Trichoderma B) and EC value (Fig. 1c) compared to CK, leading to the indefinitely neutral soil pH (Fig. 1d). In addition, no significant change in the TC or BS treatments compared to CK were found. Analysis of soil enzyme activities under the different treatments showed that compared with CK, the urease (Fig. 1e), sucrase (Fig. 1f), neutral phosphatase (Fig. 1g) and cellulase (Fig. 1h) activities in soils treated with TC, TC-BS, 2TC and 2TC-BS were significantly increased. The cellulase activity increased with increasing amounts of tomato residues returned to the field. These results demonstrated that the combined application of tomato residues and B. subtilis was more conducive to the improvement in soil physicochemical properties and soil enzyme activities.
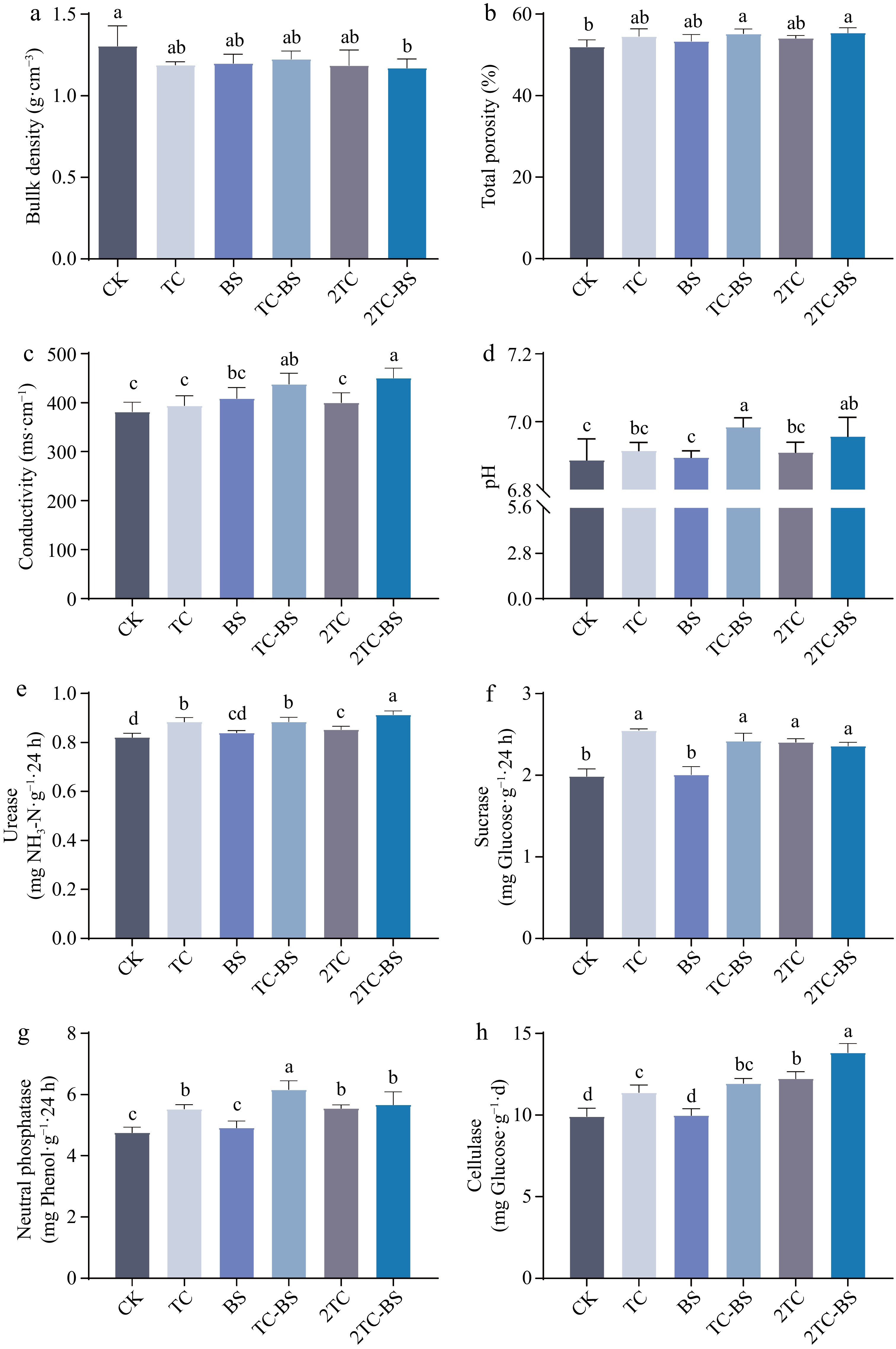
Figure 1.
Effects of different treatments on soil physical and chemical properties and soil enzyme activity. The different letters indicated significant differences as assessed by the Duncan test (p < 0.05; n ≥ 3). Conventional fertilization (CK), tomato residues (TC), B. subtilis (BS), tomato residues + B. subtilis (TC-BS), double tomato residues (2TC), and double tomato residues + B. subtilis (2TC-BS).
Soil nutrient content
-
The effects of the return of tomato residues combined with the application of B. subtilis on soil nutrient contents were measured (Table 1). Compared with CK, the TC and 2TC significantly increased the content of the organic substances and total carbon and the C/N ratio of the soil; moreover, the TC-BS and 2TC-BS treatments significantly increased the total carbon content and the C/N. While the 2TC, TC-BS, and 2TC-BS decreased the total N content of the soil. In addition, the BS treatment led to no significant changes in soil nutrient contents compared with CK. These results suggested that combined application with tomato residues was important for improving the soil nutrient content.
Table 1. Effects of the different treatments on soil nutrient content.
Treaments Organic substance
(g·kg−1)Total sarbon
(g·kg−1)Total nitrogen
(g·kg−1)Alkali-hydro nitrogen
(mg·kg−1)C/N CK 23.29 ± 1.87 b 23.08 ± 1.44 c 2.88 ± 0.22 ab 265.97 ± 13.55 ab 7.96 ± 0.28 c TC 25.95 ± 1.01 a 27.03 ± 1.17 a 3.02 ± 0.12 a 281.51 ± 10.32 a 8.94 ± 0.24 b BS 24.45 ± 0.84 ab 22.63 ± 1.20 c 2.84 ± 0.13 ab 257.63 ± 9.68 b 7.98 ± 0.31 c TC-BS 24.67 ± 2.04 ab 26.17 ± 0.69 ab 2.63 ± 0.13 bc 264.23 ± 11.30 ab 9.98 ± 0.66 a 2TC 25.88 ± 1.59 a 25.09 ± 1.14 b 2.42 ± 0.21 b 268.42 ± 20.54 ab 10.41 ± 1.02 a 2TC-BS 25.05 ± 1.66 ab 25.15 ± 1.24 b 2.64 ± 0.21 bc 281.93 ± 9.46 a 9.60 ± 0.94 ab The different letters indicate significant differences as assessed by the Duncan test (p < 0.05; n ≥ 3), conventional fertilization (CK), tomato residues (TC), B. subtilis (BS), tomato residues + B. subtilis (TC-BS), double tomato residues (2TC), and double tomato residues + B. subtilis (2TC-BS). Soil microbial community structure
-
The Chao1 index, Goods_Coverage index, the Shannon and Simpson indices were used to characterize the abundance, the coverage, and the diversity of the microbial communities, respectively (Table 2). The diversity of soil bacteria were analyzed and counted. The Chao1 index of the TC-BS and 2TC treatments was significantly higher than that of the CK and the differences between the Goods_Coverage index and Shannon index among the treatments were not significant. While the Simpson index of all treatments except for BS was significantly higher than that of the CK. These results showed that the addition of tomato residues was also important for the enrichment of microbial communities in the soil.
Table 2. Effects of the different treatments on the alpha diversity of the soil bacteria.
Treatments Chao1 Goods_coverage Shannon Simpson CK 6,981.83 ± 202.57 bc 0.97 ± 0.01 ab 10.68 ± 0.18 a 0.99 ± 0.00 b TC 7,131.05 ± 378.01 abc 0.98 ± 0.01 ab 10.59 ± 0.30 a 1.00 ± 0.00 a BS 6,752.89 ± 333.69 c 0.98 ± 0.00 a 10.08 ± 0.64 a 0.99 ± 0.01 b TC-BS 7,456.27 ± 277.17 a 0.97 ± 0.00 b 10.48 ± 0.31 a 1.00 ± 0.00 a 2TC 7,515.53 ± 198.99 a 0.97 ± 0.00 b 10.67 ± 0.16 a 1.00 ± 0.00 a 2TC-BS 7,216.68 ± 296.77 ab 0.97 ± 0.00 b 10.64 ± 0.18 a 1.00 ± 0.00 a The different letters indicate significant differences as assessed by the Duncan test (p < 0.05; n ≥ 3), conventional fertilization (CK), tomato residues (TC), B. subtilis (BS), tomato residues + B. subtilis (TC-BS), double tomato residues (2TC), and double tomato residues + B. subtilis (2TC-BS). The relative abundance of bacteria in the soil at the phylum and genus levels were also studied. Thirty-one bacterial phyla were identified through taxonomic composition analysis of high-throughput OTU sequences (Fig. 2a). The dominant bacterial phyla in the soil were Actinobacteria, Proteobacteria, Firmicutes, Chloroflexi, Acidobacteriota, and Bacteroides, while Actinobacteria, Proteobacteria, Firmicutes, and Bacteriodetes were the main dominant phylum of the tomato root microbial community[31]. Here there was no significant difference in the relative abundance of the Proteobacteria, Firmicutes, Bacteroidetes, and Gemmatimonadetes in the soils of the various treatments. Furthermore, the relative abundance of the Planctomycetes phylum under the BS, TC, 2TC, and 2TC-BS treatments was significantly increased compared to the CK. While the relative abundance of Deinococcus-Thermus among the BS, TC, TC-BS, 2TC, and 2TC-BS treatments were significantly reduced.
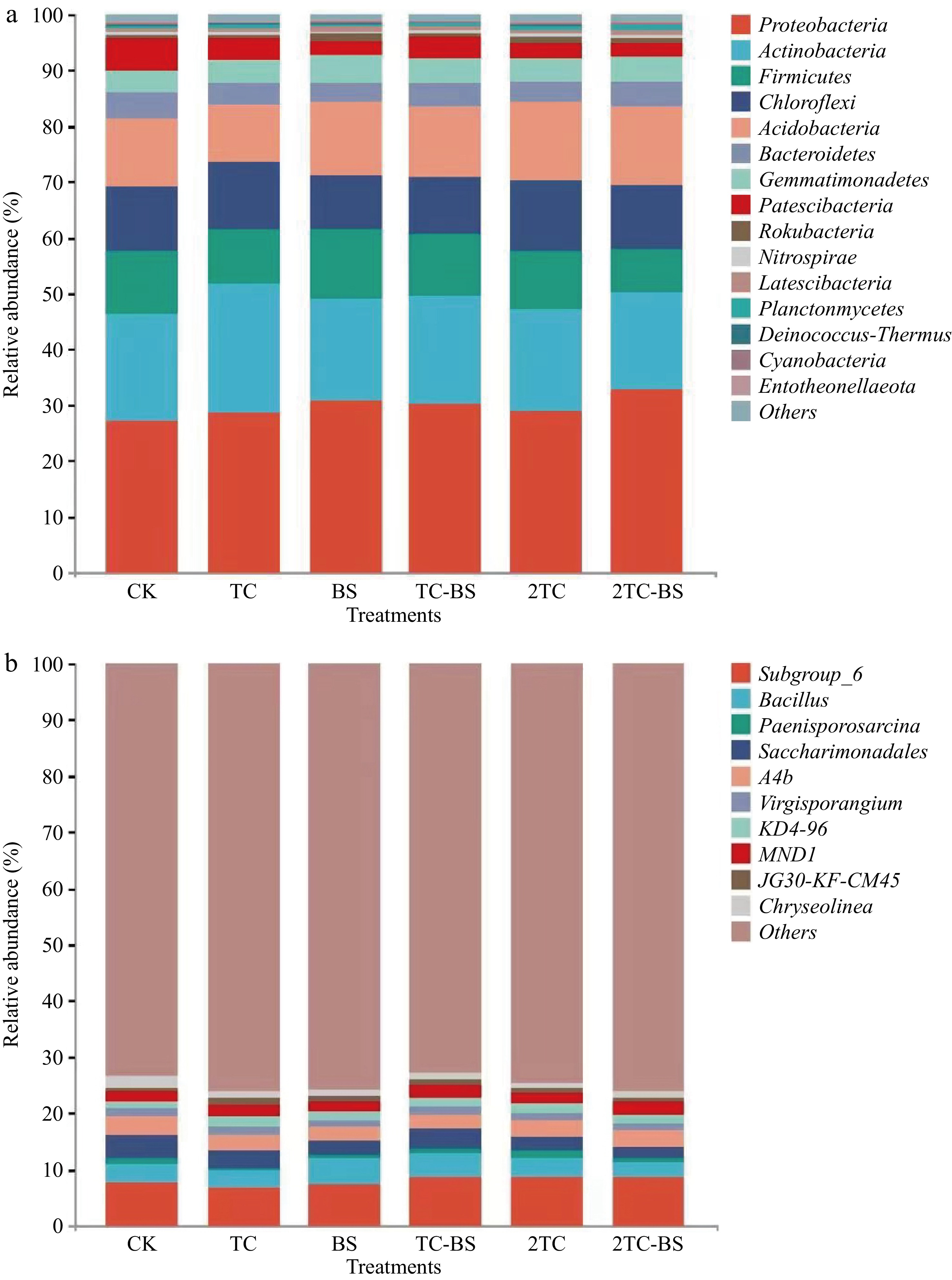
Figure 2.
Effects of the treatments on relative abundance of bacteria in soil at phyla and genera level. Conventional fertilization (CK), tomato residues (TC), B. subtilis (BS), tomato residues + B. subtilis (TC-BS), double tomato residues (2TC), and double tomato residues + B. subtilis (2TC-BS).
In addition, a total of 483 bacterial genera were also identified (Fig. 2b). Compared with CK, BS treatment significantly increased the relative abundance of Bacteroidetes, SBR1031, KD4-96, MND1, and Lysobacter but decreased that of Paenisporosarcina and A4b. Studies have shown that four species of the Lysobacter, Lysobacter enzymogenes, Lysobacter antibioticus, Lysobacter brunescens, and Lysobacter gummosus have inhibitory effects on various microorganisms and nematode activities[32,33]. TC treatment significantly increased the relative abundance of KD4-96 and MND1 and decreased the relative abundance of Bacteroidetes. TC-BS treatment significantly increased the relative abundance of Subgroup-6, Paenisporosarcina, Lysobacter, KD4-96, and MND1 and decreased the relative abundance of A4b and JG30-KF-CM45. These results indicated that the combined application of both tomato residues and B. subtilis helped improve the diversity and the relative abundance of bacteria at the phylum and genus levels in the soil.
Growth, quality, and yield of tomato
-
It was found that the addition of tomato residues and combined application with B. subtilis led to significant effects on soil indicators, thus, whether it also affected the aboveground growth in tomato was explored. The results indicated that compared with CK, the TC and TC-BS treatments significantly increased the plant height at the flowering stage (Fig. 3a), but the difference in thick stem among the treatments was not significant (Fig. 3b). The root activity of TC-BS and 2TC-BS was also higher (Fig. 3c). In addition, the total chlorophyll content of the TC, TC-BS, 2TC, and 2TC-BS treatments was significantly increased compared with that of the CK (Fig. 3d). Furthermore, the net photosynthetic rate was significantly improved in the TC-BS treatment and even the 2TC-BS treatment was improved more significantly compared to the other treatments (Fig. 3e).
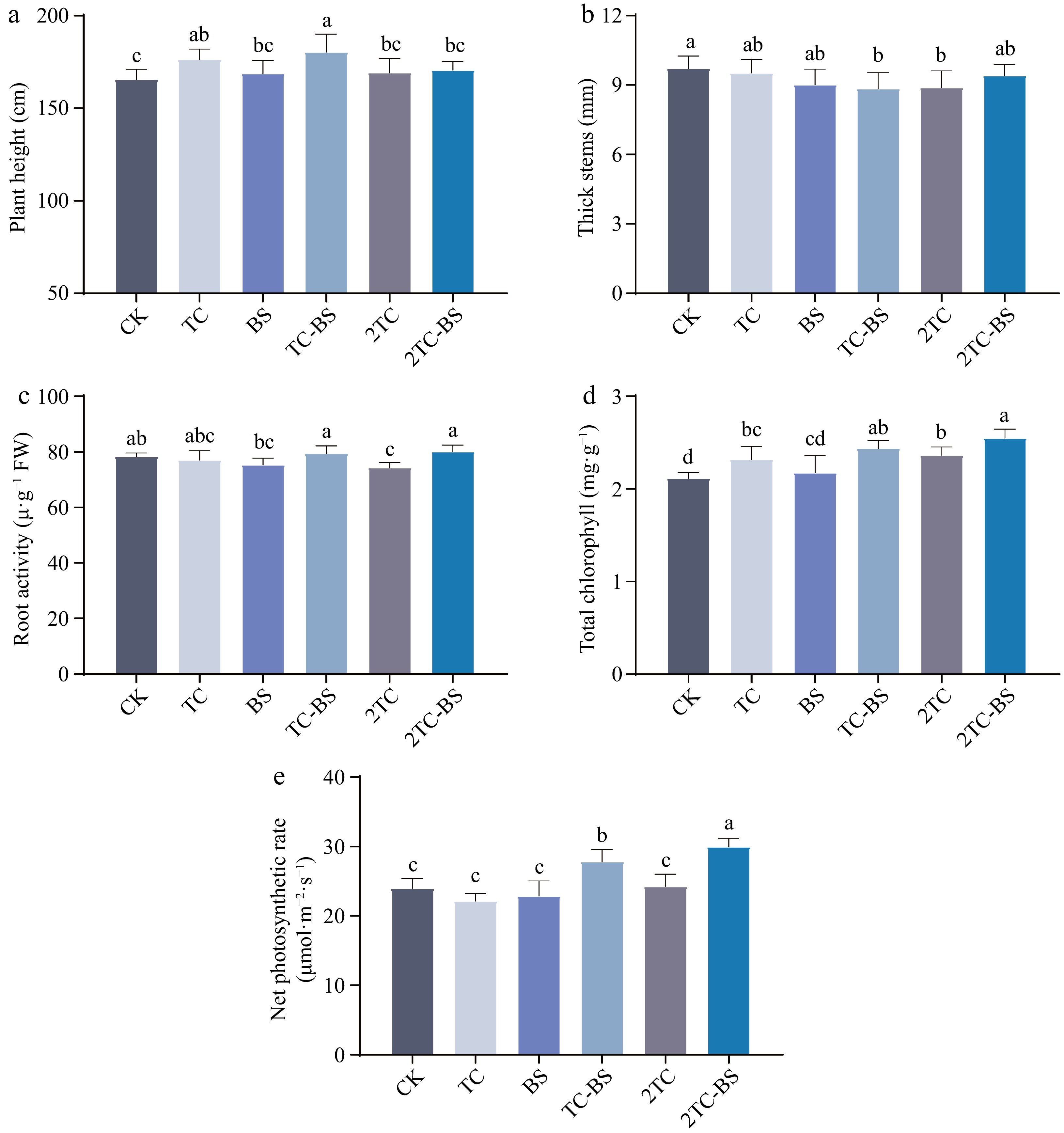
Figure 3.
Effects of the different treatments on the growth and physiology of the tomato. The different letters indicated significant differences as assessed by the Duncan test (p < 0.05; n ≥ 3), conventional fertilization (CK), tomato residues (TC), B. subtilis (BS), tomato residues + B. subtilis (TC-BS), double tomato residues (2TC), and double tomato residues + B. subtilis (2TC-BS).
The combined application of tomato plant residue with B. subtilis also affected the tomato fruit quality (Table 3). The present analysis revealed that the soluble solids and VC contents of tomato fruits in the TC treatment and the VC content in the BS treatment were significantly higher than those in the CK treatment. While the TC, TC-BS, and 2TC-BS treatments significantly increased the soluble sugar, sugar/acid ratio, soluble protein, and lycopene contents. Additionally, the BS, TC-BS, and 2TC treatments significantly reduced the content of titratable acid.
Table 3. Effect of the different treatments on fruit quality of tomato.
Treatments Soluble solids
(%)Soluble sugars
(%)Titratable acid
(%)Sugar/acid Soluble protein
(mg·g−1)Vc
(ug·g−1)Lycopene
(ug·g−1)CK 6.46 ± 0.37 bc 6.86 ± 0.18 cd 0.62 ± 0.02 a 11.13 ± 0.44 c 1.17 ± 0.07 d 115.39 ± 4.59 c 22.06 ± 1.27 cd TC 7.14 ± 0.57 a 7.89 ± 0.78 b 0.60 ± 0.03 a 13.11 ± 1.89 b 1.72 ± 0.24 b 121.86 ± 4.38 b 23.32 ± 0.41 bc BS 5.90 ± 0.33 c 6.29 ± 0.73 d 0.57 ± 0.01 b 11.05 ± 1.17 c 1.01 ± 0.14 d 135.24 ± 3.82 a 20.76 ± 0.34 de TC-BS 6.64 ± 0.49 ab 8.10 ± 0.94 b 0.58 ± 0.01 b 14.00 ± 1.64 ab 1.96 ± 0.12 a 121.33 ± 4.37 bc 26.49 ± 0.68 a 2TC 6.72 ± 0.59 ab 7.75 ± 0.65 bc 0.58 ± 0.01 b 13.33 ± 0.96 ab 1.05 ± 0.01 d 123.78 ± 3.94 b 20.35 ± 1.17 e 2TC-BS 7.04 ± 0.59 ab 9.23 ± 0.62 a 0.61 ± 0.01 a 15.05 ± 0.96 a 1.43 ± 0.18 c 118.75 ± 1.99 bc 24.29 ± 1.17 b The different letters indicate significant differences as assessed by the Duncan test (p < 0.05; n ≥ 3), conventional fertilization (CK), tomato residues (TC), B. subtilis (BS), tomato residues + B. subtilis (TC-BS), double tomato residues (2TC), and double tomato residues + B. subtilis (2TC-BS). Moreover, the statistical analysis of tomato fruit yield (Table 4) demonstrated that compared with CK, the 2TC treatment significantly increased the weight per fruit. And the yield per plant and total yield of the TC, BS, TC-BS, 2TC, and 2TC-BS treatments were also significantly higher than those of CK. In terms of the rate of increase in the total yield, TC-BS treatment showed the highest increase of 10.98%, followed by that of 2TC-BS treatment at 10.95%; and 2TC treatment showed a relative increase of 7.09%. BS and TC treatments increased by 5.92% and 3.44%, respectively.
Table 4. Effects of the different treatments on tomato fruit yield.
Treatments Single fruit weight
(g·single fruit−1)Yield per plant
(kg·plant−1)Increased
productivityCK 97.60 ± 10.63 bc 2.05 ± 0.05 d − TC 90.62 ± 9.32 c 2.12 ± 0.05 c 3.44% BS 93.70 ± 11.38 bc 2.20 ± 0.03 bc 5.92% TC-BS 100.83 ± 10.32 abc 2.28 ± 0.07 a 10.98% 2TC 116.70 ± 7.40 a 2.25 ± 0.04 b 7.09% 2TC-BS 113.26 ± 11.76 ab 2.28 ± 0.04 a 10.95% The different letters indicate significant differences as assessed by the Duncan test (p < 0.05; n ≥ 3), conventional fertilization (CK), tomato residues (TC), B. subtilis (BS), tomato residues + B. subtilis (TC-BS), double tomato residues (2TC), and double tomato residues + B. subtilis (2TC-BS). -
Vegetable residues contained a large number of cellulolytic bacteria, which can decompose the residues into nutrients that are easily absorbed by plants during the decomposition process after being returned to the field[34]. B. subtilis can assist plants in obtaining nutrients through a variety of mechanisms, including increasing the surface area and nitrogen fixation capacity of plant roots, increasing phosphorus solubilization, and producing indole-3-acetic acid (IAA)[35]. Returning the tomato residues which are rich in nitrogen and carbon to the field was conducive to the mineralization of soil nitrogen and increased the content of alkaline nitrogen in the soil[36] promoting the synthesis of leaf photosynthetic pigments and thereby enhancing plant photosynthesis[37]. In this study, single and double application of tomato residues increased the contents of tomato soil organic substance, total carbon and total nitrogen; when adding B. subtilis, only the total carbon content of the soil significantly increased (Table 1). Moreover, the application of B. subtilis alone had a nonsignificant effect on the soil organic substance while significantly increasing it after combining with tomato residues (Table 1), because the growth and propagation of B. subtilis consumes organic substances in the soil. Therefore, the synergistic effect of B. subtilis and tomato residues were more beneficial for increasing the nutrient content of the soil.
Soil enzyme activity is one of the most important indicators for evaluating soil fertility and can effectively reflect the intensity and direction of various biochemical metabolic processes in soil[38,39]. Numerous studies have reported that straw return can improve the activities of soil urease, acid phosphatase, catalase, and invertase[11]. Studies also have found that the yield and quality of tomato fruits were positively correlated with soil enzyme activities, where the influence of soil invertase was most prominent[40]. Asghar & Kataoka[41] found that inoculation with Trichoderma RW309 in organic compost significantly increased the phosphatase activity of ginger soil, mainly because the strain increased the diversity of ginger rhizosphere microbes and the inducible compounds secreted by these microbes improved the soil enzyme activity[42]. The results of this study demonstrated that the synergistic effect of returning tomato residues to the field combined with applying B. subtilis significantly increased the urease, neutral phosphatase, sucrase, and cellulase activities in the soil, where the increase of cellulase activity was the largest (Fig. 1). Similarly, as mentioned earlier, the increase of cellulase activity can enhance the absorption of soil nutrients.
When tomato residues were returned to the field along with B. subtilis, a large amount of organic energy and abundant microorganisms were input into the soil. The cellulose in the residues were decomposed by cellulase into sugars, furfurals, and organic acids[43], followed by an invertase-catalyzed reaction to produce glucose, fructose, etc., thereby providing energy for microbial activities. Previous studies have revealed that the soil microbial community is determined by trophic factors and is affected by soil nutrients, temperature, and humidity. In particular, the soil pH has been demonstrated in multiple studies to be the strongest factor in the formation of the microbial community structure[44,45]. In this study, compared with conventional fertilization (CK), the application of tomato plant residue in combination with B. subtilis significantly increased the soil pH toward neutrality (Fig. 1) and significantly increased the Chao1 and Simpson indices (Table 2). The Simpson index reflected the diversity of the community, which is influenced by the species richness and evenness in the sample community, laying the foundation for the changes in the structure of bacterial communities in the tomato soil. Additionally, Wei et al.[46] showed that returning the fruit and vegetable residues to the field in solar greenhouses significantly increased the relative abundance of Gemmatimonadetes, Planctomycetes, and Saccharomycota. Xie et al.[30] indicated that the application of B. subtilis in ginger fields treated with tomato plant residue compost significantly increased the soil bacteria diversity, and the relative abundance of the Beneficial Proteobacteria, Acidobacteria, Chloroflexi, and Bacillus were also significantly increased. In this study, returning tomato residues to the field combined with B. subtilis treatment significantly increased the relative abundances of Actinobacteria, Acidobacteria, Sphingomonas, and Lysobacter. Studies have reported that bacteria such as Actinobacteria determine the extent and speed of the degradation of organic substances like straw[47]. Sphingomonas can degrade varieties of harmful organic compounds[48,49]. The Lysobacter spp. (Lysobacter enzymogenes, Lysobacter antibioticus, Lysobacter brunescens, and Lysobacter gummosus) can inhibit the activity of nematodes[32,33]. The increase in the abundance of these beneficial microorganisms can help degrade the harmful organic and inorganic compounds in the soil and prevent and control nematodes, thereby achieving the effect of soil improvement. Therefore, returning tomato residues to the field combined with the application of B. subtilis was also of great significance in improving the structure of soil microbial communities and thus improving soil structure and nutrient content.
Returning vegetable residues to the field has good potential for promoting plant growth and improving fruit quality[50]. Bacillus had been used previously in the formulation of biopesticides, and biofertilizers with plant growth-promoting factors[51]. Studies have demonstrated that sugar content and acidity are two important aspects that affect the flavor and quality of tomato fruit. The indicators for evaluating the taste of tomato fruits can be represented by soluble solids, soluble sugars, organic acids, and sugar-to-acid ratio[52,53]. In this study it was found that the single application of tomato residues or the single application of B. subtilis resulted in a soluble sugar content of ≥ 7.75% and a sugar-acid ratio of 13.11 or above in tomato. Moreover, lycopene is the main pigment in tomato fruit with strong antioxidant ability[54], which can inhibit cell proliferation, and invasion and prevent cell apoptosis[55]. The present study showed that both single application of tomato residues and single application of B. subtilis significantly increased the content of lycopene and vitamin C in tomato. Therefore, both returning tomato residues to the field or the application of B. subtilis individually have a significant promoting effect on improving the fruit quality of tomato.
Gupta et al.[56] found that applying two strains of B. subtilis in the soil could replace 20% of the mineral fertilizer and achieve the optimal yield of chili peppers. The use of Pseudomonas putida can reduce 25% of inorganic fertilizers, which can improve the growth and productivity of pepper[57]. The active substances secreted by B. subtilis can stimulate an increase of crop growth-type endogenous hormones such as IAA and GA and a decrease in ABA, helping to increase the yield and income[35]. The results of this study also indicated that the application of B. subtilis in combination with returning single or double tomato residues significantly increased the plant height, root vitality, chlorophyll content, and net photosynthetic rate of tomato.
-
Returning tomato residues to the field combined with the application of B. subtilis significantly increased the relative abundance of beneficial microorganisms and increased soil enzyme activities and nutrient content in the soil for tomato growth, thereby promoting tomato yield and improving fruit quality. Moreover, it also realizes the resource utilization of tomato residues, providing a theoretical basis and technical guidance for resource utilization and high-quality production of tomato residues.
In summary, the two treatments, TC-BS and 2TC-BS, combining the addition of tomato residues and B. subtilis to the field were more conducive to the aboveground growth of tomato.
-
The authors confirm contribution to the paper as follows: study design and supervision: He C; experiment performing and draft manuscript writing: Shi A, Di Q; data analysis: Shi A, Xie K; study discussion and manuscript revision: Feng X, Wang X, Li Y, Sun M, Wang J, Han L, Yu X. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article, and are available from the corresponding author on reasonable request.
This research was financially supported by the National key R&D project of China (2022YFD1602403). The Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-IVFCAAS). This research acknowledges the support of The Key Laboratory of Horticultural Crop Biology and Germplasm Innovation, The Ministry of Agriculture, China.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Aokun Shi, Qinghua Di
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Shi A, Di Q, Xie K, Feng X, Wang X, et al. 2024. Tomato plant residues and Bacillus subtilis application can improve the soil environment and increase tomato yield and quality. Technology in Horticulture 4: e024 doi: 10.48130/tihort-0024-0021 

Tomato plant residues and Bacillus subtilis application can improve the soil environment and increase tomato yield and quality
- Received: 16 May 2024
- Revised: 15 August 2024
- Accepted: 27 August 2024
- Published online: 09 October 2024
Abstract: The maintenance of vegetable yield, the improvement of vegetable quality, and the utilization of vegetable residue are the top priorities for developing sustainable facility agriculture. To explore the residue resource utilization and the high quality and yield of tomatoes in solar greenhouses, conventional fertilization was used as the control (CK) in this study. To explore the effects of cultivation with tomato residues and B. subtilis (BS) five treatments were carried out as follows: 1. tomato residues (TC); 2. B. subtilis (BS); 3. tomato residues + B. subtilis (TC-BS); 4. double tomato residues (2TC) and 5. double tomato residues + B. subtilis (2TC-BS). The tomato growth, physiology, yield, quality, soil physicochemical traits, nutrient content, enzyme activity, and bacterial diversity were measured. The results showed that compared with CK, the synergy of TC and BS was important in improving tomato yield and soil quality. TC-BS and 2TC-BS significantly increased the plant height, root activity, total chlorophyll content, and net photosynthetic rate with up to 10.98% and 10.95% increases in yield, respectively. The tomato fruit quality such as the lycopene, soluble sugar, soluble protein, and vitamin C (VC) contents were also significantly increased. Additionally, 16S rDNA sequencing results indicated that the TC-BS and 2TC-BS significantly increased the diversity of soil bacteria. In conclusion, increasing the application of B. subtilis along with incorporating tomato residues can significantly improve the soil environment, enrich the beneficial microorganisms, and promote tomato growth, its yield, and quality.
-
Key words:
- Tomato /
- Tomato residues /
- Bacillus subtilis /
- Soil nutrient /
- Yield and quality