










-
The compound ferulic acid (FA) is derived from the resin of asafetida, a perennial herb belonging to the Umbelliferaceae family. Within the plant kingdom, FA serves as a prevalent phenolic acid with diverse health benefits, encompassing its role as an efficacious scavenger of free radicals, antithrombotic and antibacterial agent, modulator of inflammation, among other effects[1]. The physiological functions of FA as an antioxidant are numerous, although its mechanism is not fully understood compared to other antioxidants.
The redox potential value serves as a macroscopic manifestation of intricate reactions occurring within the intestinal environment. Fluctuations in redox potential are intricately associated with the multifaceted response of intestinal bacteria and overall gastrointestinal health[2]. Diarrhea frequently co-occurs with inflammation, resulting in alterations in the microbial composition of the gut as bacteria flourish in an inflammatory and oxygen-rich environment. Intestinal dysbiosis is characterized by enhanced respiratory metabolism of aerobic bacteria, elevated redox potential[3]. This ecological imbalance provides electron acceptors for cellular metabolism, thereby prompting microorganisms to prefer metabolic reactions with higher thermodynamic redox potential energy[4]. Conversely, due to the anaerobic environment, metabolic reactions with lower oxidation-reduction potential energy are favored in thermodynamics within a healthy organism[5]. Therefore, it is crucial to regulate the intestinal chemical environment to reduce redox potential and promote stability within the intestinal microecology while reducing the occurrence of diarrhea. Research has substantiated that dietary consumption of phenolic acids can augment gut functionality through their anti-inflammatory properties on the gastrointestinal tract and modulation of the gut microbiome[6]. The multifaceted regulatory functions of the intestinal microbiota primarily stem from its metabolic capacity and the array of metabolites it generates[7]. These microbial metabolites regulate host homeostasis through various mechanisms, while polyphenols modulate the composition and metabolism of the intestinal microbiome[8,9]. Consumption of antioxidant-rich foods could serve as an effective strategy for modulating the oxygen environment within the gastrointestinal tract[10,11].
The magnesium citrate diarrhea model involves administering a hypertonic solution of magnesium citrate to animals, which is poorly absorbed by the body and leads to the development of chronic diarrhea in rats[12]. Nutritional chronic diarrhea commonly manifests in weaned piglets during clinical production. Diarrhea often coincides with intestinal Enterobacter dilatation, and the heightened oxygen content within the intestine leads to an elevation in intestinal redox potential, thereby facilitating intestinal Escherichia coli dilatation[13]. Given the association between intestinal redox potential and host susceptibility to pathogenic bacteria, it is worth investigating whether low redox potential correlates with nutritional diarrhea. Therefore, studying the pathogenesis, drug efficacy, and nutritional support of chronic diarrhea holds significant clinical value. As an antioxidant, FA possesses a diverse range of physiological functions. However, the precise mechanism underlying its antioxidative properties remains incompletely understood. Antioxidants can modulate the redox potential within the gastrointestinal tract. Despite limited research on mammals, it is worth noting that apart from its antioxidant effects, FA may exert influence on intestinal redox potential through the regulation of gut microflora and metabolites. Notably, there existed a correlation between redox potential and intestinal health (e.g., diarrhea) in animals. Consequently, further investigation is warranted to explore whether FA can alleviate diarrhea by mitigating redox potential. The current study will provide new insights for modulating intestinal redox potential and alleviating chronic diarrhea.
-
The assessment of antioxidant activity in vitro was conducted using various concentrations of antioxidants, as outlined in the references[14]. Antioxidants including chemical classes [Ethoxyquin (EQ, HPLC, 94.7%) and butylated hydroxytoluene (BHT, HPLC, 98.1%)], polyphenols (FA, Supplementary Information 1, HPLC, 99.9% and resveratrol, HPLC, 99.8%), natural pigments (anthocyanin, HPLC, 95.1% and carotene, HPLC, 99.3%), vitamins [vitamin C (VC, HPLC, 95.2%) and vitamin E (VE, HPLC, 93.6%)] and rutin (HPLC, 96.3%) were evaluated 2,2'-diphenyl-1-picrylhydrazyl (DPPH), 2,2'-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) and ferric-reducing antioxidant power (FRAP) activity. All antioxidants were purchased from Shanghai Yuanye Biotechnology Co., Ltd. (Shanghai, China). The DPPH scavenging activity of antioxidants was assessed using the DPPH scavenging assay kit (Solarbio, Beijing, China). Briefly, a 10 µL sample was combined with 190 µL of DPPH working solution and incubated in darkness at room temperature for 30 min. Subsequently, absorbance was measured at 515 nm. The percentage of DPPH free radical scavenging rate (%) was calculated using the formula: [As0 − (As1− As2)] / As0 × 100, where As0 represents the absorbance of the control group (dH2O replaces the sample solution), As1 represents the absorbance of the sample mixed with DPPH solution, and As2 represents the absorbance with only the sample solution. The ABTS clearance assay kit was employed to evaluate the ABTS scavenging activity of nine antioxidants[15]. Briefly, a 10 µL sample was added to 190 µL of ABTS working solution at ambient temperature and incubated in the absence of light for 6 min. Subsequently, the absorbance was measured at 405 nm. The percentage scavenging rate of ABTS free radicals can be calculated using the formula (%) = [As0 − (As1 − As2)] / As0 × 100, where As0 represents the absorbance of the control group (replacing the sample solution with dH2O), As1 represents the absorbance of the sample mixed with ABTS, and As2 represents the absorbance of an individual sample solution. Ferric-reducing activity of antioxidants was evaluated using the FRAP assay kit (Yuanye, Shanghai, China) according to the manual instructions. Briefly, 264 µL FRAP working solution was added to 30 µL of sample and incubated at 37 °C for 30 min. The absorbance was measured at 593 nm. The results were expressed as Fe2+ quivalents.
Intervention experiments of different antioxidants in vivo
-
EQ, FA, rutin, and resveratrol antioxidants were screened based on FRAP analysis. Six-week old male sprague-dawley (SD) rats (average initial weight 150 g) were purchased from Cavens Biogle (Suzhou, China) and allowed to acclimatize to the animal facility environment for a week before the experiments. All rats had ad libitum access to food and water. Thirty rats were randomly divided into five groups, i.e., the control (n = 6) group, EQ (n = 6), FA (n = 6), rutin (n = 6), and resveratrol (n = 6) groups. The experimental rats were individually housed in a single cage with an iron frame at the bottom. Refer to Supplementary Information 2 and Supplementary Information 3 for an inventory of various antioxidant dietary formulations. In detail, low and high concentrations of antioxidants were supplemented into the daily ration of rats in the EQ, FA, rutin, and resveratrol groups. After the end of the low concentration of antioxidants intervention period, the high concentration of antioxidants intervention began after feeding the antioxidant-free diet for one week. All interventions continued for 15 d. During the experiment, fresh fecal samples were collected daily for redox potential measurements. Finally, all rats were euthanized by CO2 asphyxiation, ileum and colonic tissues, and digesta were collected. Samples were immediately snap-frozen and transferred for storage at −80 °C until further 16S rRNA gene sequencing and metabolite analysis.
Rat diarrhea model induced with magnesium citrate
-
A magnesium citrate-induced diarrhea model[12] was established using 6-week-old SD rats. Thirty rats were randomly divided into three groups: Con (no treatment; n = 10); ConMg (Chronic Mg-induced diarrhea; n = 10), and FAMg (a diet supplemented with FA in chronic Mg-induced diarrhea; n = 10). Throughout the experiment, rats in Con and ConMg groups were provided with a standard diet and had ad libitum access to feed and water. Rats in the FAMg group were given a diet enriched with FA antioxidant along with ad libitum access to feed and water. The experimental rats were individually housed in a cage with an iron frame at the bottom. After 10 d of feeding, the corresponding chronic diarrhea model was established. The weight, water intake, redox potential, and diarrhea index were measured at consistent time points. The fecal samples of days 7, 17, and 24 were immediately snap-frozen and transferred for storage at −80 °C for further 16S rRNA gene sequencing analysis.
Redox potential measurement
-
Fresh feces and intestinal digesta were collected in a pre-filled zip-lock bag with CO2, ensuring that each sampling point was controlled within a half-hour timeframe. After sample collection, it was promptly sealed and mixed before determining the redox potential. Subsequently, the samples were collected using freeze-storage tubes free from dnases and rnases to preserve their microbial composition and antioxidant capacity. All samples were initially flash-frozen in liquid nitrogen and then transferred to a −80 °C freezer. In accordance with our previous methodology[13], the redox potential of feces and intestinal digesta was measured using a direct method. The fecal redox potential in the diarrhea model was determined through a dilution method.
Observation of diarrhea
-
The characteristics and color of the feces were observed. Each rat's loose feces and formed feces were counted daily, and the diameter of each loose feces was measured using a divider and scale. Diarrhea rate and diarrhea index[12] were primary indicators to evaluate diarrhea severity. Loose feces rate = Number of loose feces per animal ÷ Total number of feces per animal. The grading system for watery feces consistency is based on diameter: < 1 cm is grade 1, ~1.9 cm is grade 2, 2~3 cm is grade 3, and > 3 cm is grade 4. Average watery feces consistency = Total volume of watery feces ÷ Number of watery feces.
Determination of T-AOC, SOD, ROS, and MDA in intestine
-
Colon and ileal tissue were excised, rinsed with ice-cold saline, and dried with filter paper. Then, the samples were homogenized with 0.1 g/mL wet weight of ice-cold physiological saline in a glass Teflon homogenizer for 3 min at 4 °C[16], which was followed by centrifugation at 2,000× g for 20 min at 4 °C. The supernatant was collected and used for the determination of total antioxidant capacity (T-AOC), superoxide dismutase (SOD), reactive oxygen species (ROS), and malondialdehyde (MDA) using commercial kits (Solarbio, Beijing, China). Please refer to Supplementary Information 4 for detailed procedures.
16S rRNA gene sequencing and processing
-
The microbial DNA was extracted from frozen intestinal digesta and fecal samples using the E.Z.N.A.® Stool DNA Kit (Omega Bio-tek, Norcross, GA, USA) following the manufacturer's recommended protocols. DNA concentration and integrity were measured with Nanodrop 2000 (Thermo Fisher Scientific, USA) and agarose gel electrophoresis. Extracted DNA was stored at −20 °C until further processing. The extracted DNA was used as a template for PCR amplification of bacteria. 16S rRNA genes with the barcoded primers and takara ex taq (Takara). For bacterial diversity analysis[17], V3−V4 variable regions of 16S rRNA genes was amplified with universal primers 343F (5'-TACGGRAGGCAGCAG-3') and 798R (5'-AGGGTATCTAATCCT-3') for V3−V4 regions. The unweighted unifrac distance matrix performed by R package was used for unweighted unifrac principal coordinates analysis to estimate the beta diversity[18]. Then the R package was used to analyze the significant differences between different groups using the wilcoxon statistical test. The linear discriminant analysis effect size (LEfSe) method was used to compare the taxonomy abundance spectrum[19].
Metabolome analysis
-
The metabolomic analysis was performed by Shanghai Luming Biological Technology Co., Ltd (Shanghai, China). An qcquity uplc I-class plus (Waters Corporation, Milford, USA) fitted with q-exactive mass spectrometer equipped with heated electrospray ionization (ESI) source (Thermo Fisher Scientific, Waltham, MA, USA) was used to analyze the metabolic profiling in both ESI positive and ESI negative ion modes. The original LC-MS data were processed by software progenesis QI V2.3 (Nonlinear, Dynamics, Newcastle, UK) for baseline filtering, peak identification, integral, retention time correction, peak alignment, and normalization. Compound identification was based on precise mass-to-charge ratio (M/z), secondary fragments, and isotopic distribution using the human metabolome database (HMDB), lipidmaps (V2.3), metlin, and self-built databases. The extracted data were then further processed by removing any peaks with a missing value (ion intensity = 0) in more than 50% in groups, by replacing zero value with half of the minimum value, and by screening according to the qualitative results of the compound. The matrix was imported in R to carry out principal component analysis (PCoA) to observe the overall distribution among the samples and the stability of the whole analysis process. A two-tailed student’s t-test was further used to verify whether the metabolites of difference between groups were significant.
Data calculation and statistical analyses
-
Statistically significant differences were performed using SPSS Statistics 23.0 software and p < 0.05 presented the statistical significance of differences. All data are expressed as mean ± SD. Correlations between datasets were calculated using Pearman rank correlation. Besides, specific details of the statistical analyses for all experiments were displayed in the figure legends and results section.
-
All types of antioxidants exhibited high DPPH clearance (Supplementary Information 5, Fig. 1a), and different antioxidants of the same type also displayed differences in the clearance of ABTS (Supplementary Information 6, Fig. 1b). Both VE and carotene had poor clearances of DPPH and ABTS. FRAP assay involved the reduction of Fe3+ to Fe2+ by the antioxidant in the sample, resulting in the formation of a stable orange-red complex with phenolines and exhibiting a characteristic absorption peak at a wavelength of 520 nm (Supplementary Information 7, Fig. 1c). Based on FRAP analysis, EQ, FA, rutin, and resveratrol were selected for in vivo testing. The effect of different antioxidants on redox potential was analyzed via a one-way analysis of variance. Then, the dynamic changes of redox potential were not significant under the intervention of low concentration of FA (Fig. 1d). However, at high concentration, FA showed a significant reduction in fecal redox potential. The subsequent reduction of the redox potential was observed with resveratrol and rutin, followed by EQ (Fig. 1e). The ratio of Δredox potential to FRAP in high-concentration intervention test could be utilized for assessing the reduction rate of antioxidants on redox potential. The results showed that FA exhibited the highest ratio of Δredox potential to FRAP (Fig. 1f). Furthermore, compared to other antioxidants, FA also displayed the capacity to reduce the redox potential of ileal (Fig. 1g) and colonic (Fig. 1h) digesta.
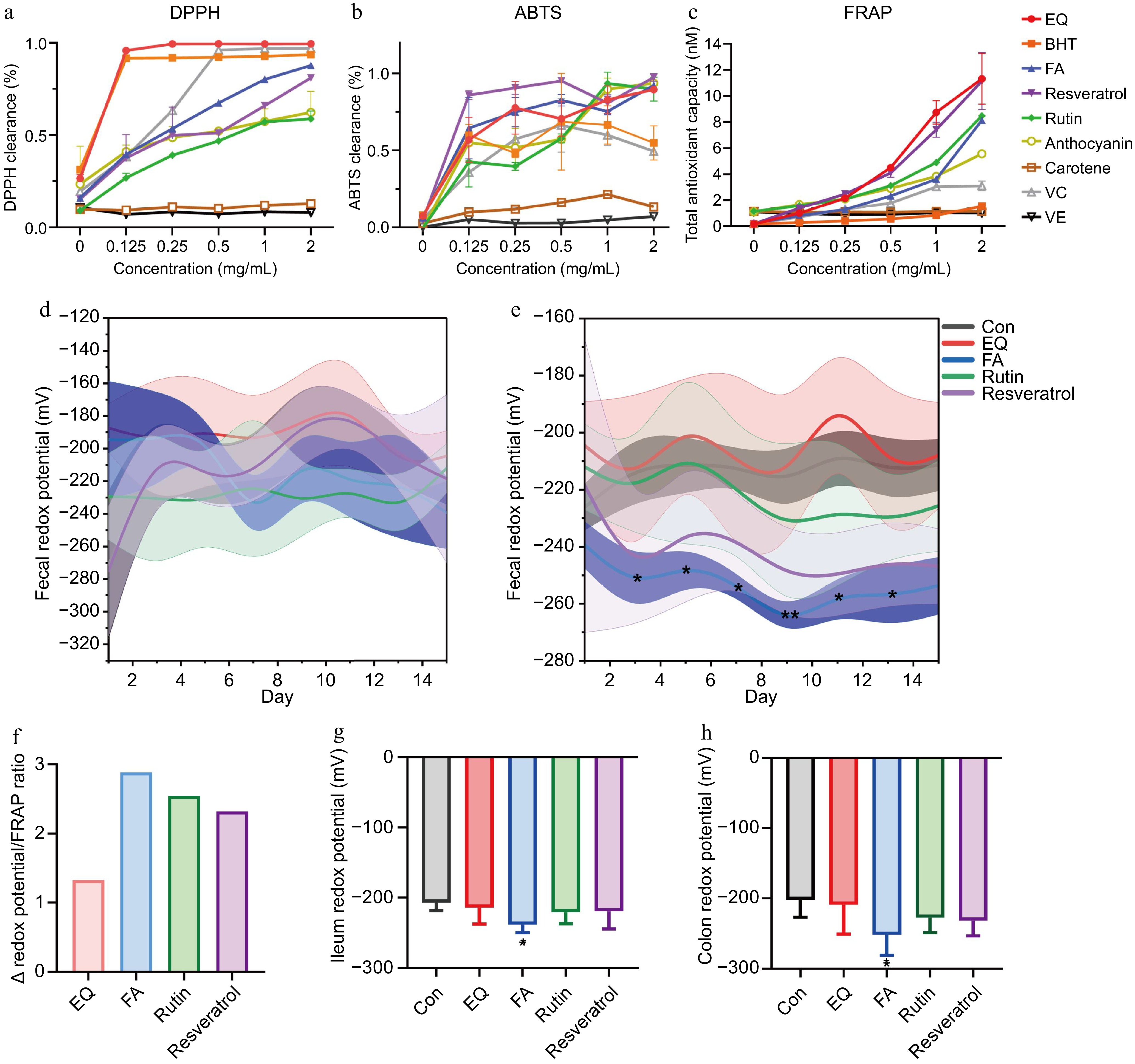
Figure 1.
Comparison of regulating redox potential ability of different antioxidants. (a), (b) The DPPH and ABTS clearance of different kinds of antioxidants in vitro. (c) Determination of the FRAP activity of different kinds of antioxidants in vitro. (d), (e) Dynamic changes of redox potential at diverse antioxidant intervention in vivo. (f) Ratio of Δ redox potential to FRAP. (g), (h) Effects of EQ, FA, rutin and resveratrol on regulating redox potential of ileal and colonic digesta.
Effects of FA supplementation on gut microbiota in rats
-
Based on the observed effects of FA on fecal redox potential, the impact of FA on intestinal microbial communities was further investigated. According to the Venn diagram (Fig. 2a), 2,311 ASVs were clustered in colonic digesta, in which 427 and 428 ASVs were unique in the Con and FA groups, respectively. As shown in Fig. 2b, compared with the Con group, FA supplementation had no significant effect on α-diversity (Chao1, Ace) of ileal and colonic microbiota (p > 0.05). Moreover, a plot of principal coordinate analysis (Fig. 2c) showed that there was no separation of colonic microbiota between the two groups (p > 0.05). However, FA significantly decreased colon redox potential and increased the abundance of Actinobacteriota (Fig. 2d; p < 0.05). At the genus level, FA significantly decreased the abundance of Prevotella and Rodentibacter and enhanced the abundance of Corynebacterium, Christensenellaceae_R-7_group in the colon (Fig. 2e; p < 0.05). LEfSe analysis was performed for diverse taxa compared with the integration of ileum and colon microflora. Eubacterium sraeum_group was the only distinctive genus in the FA ileum microbiome. Actinomycetes, Christensenellaceae_R-7_group, Erysipelatoclostridium and Paludicola were enriched by FA treatment in the colon (Fig. 2f). Prevotella and Lachnospiraceae_NK4A136_group were enriched in the colon of the Con group.
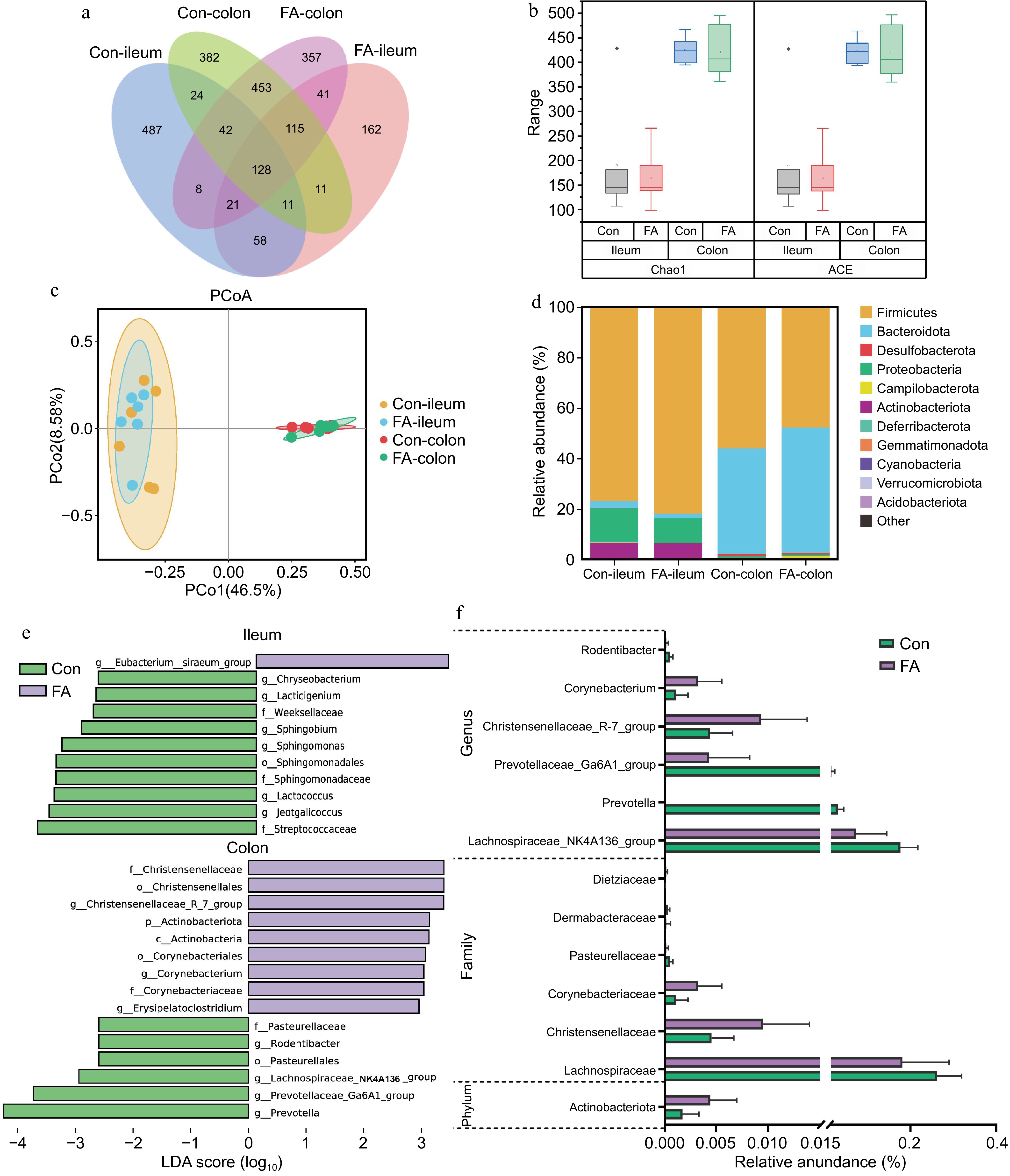
Figure 2.
Effects of FA supplementation on intestinal microbes at different taxonomic levels of rats. (a) Venn diagrams show the numbers of unique and shared ASVs between the four groups. (b) The alpha diversity of ileal and colonic microbiota. (c) Unweighted principal coordinate analysis of ileal and colonic microbiota. (d) Composition of ileal and colonic microbiota at the phylum level. (e) LDA score plot generated from LEfSe analysis used to evaluate the differentially abundant. Taxa from phylum to genus of ileal and colonic microbiota (LDA Score > 2, p < 0.05). Con = Control group; FA = FA supplementation group; ASV = Amplicon sequence variant; LDA = Linear discriminant analysis; LEfSe = Linear discriminant analysis effect size. (f) The relative abundance of colonic microbes with significant differences.
Effects of FA supplementation on metabolites in the gut of rats
-
The volcano plot showed that 48 differentially metabolites (15 up-regulated and 33 downregulated) were identified in ileal digesta (Fig. 3a) and 255 differentially metabolites (111 up-regulated and 144 down-regulated) were identified in colonic digesta (Fig. 3c). Overall, the most significant downregulated metabolite was gluconic acid in ileal digesta (Fig. 3b; p < 0.05) and alpha-muricholic acid in colonic digesta. Whereas the most significant up-regulated metabolite was tic10 in ileal digesta and azelaic acid in colonic digesta (Fig. 3d; p < 0.05). The chemical properties of differential metabolites were classified based on Funmeta (
https://funmeta.oebiotech.com/ ). The results showed that FA increased the proportion of reductive metabolites in up-regulated metabolites and decreased the proportion of oxidizing metabolites in down-regulated metabolites (Fig. 3e & f). Furthermore, D-fructose was found to be a reductive metabolite that was significantly up-regulated in the ileum (Fig. 3g; p < 0.05) but down-regulated in the colon (Fig. 3h; p < 0.05). In the ileum (Fig. 3i), the differential metabolites were predominantly enriched in lipoic acid metabolism and pentose phosphate pathway, as well as in galactose metabolism and amino sugar and nucleotide sugar metabolism. On the other hand, the differential metabolites in the colon (Fig. 3j) were significantly enriched in metabolic pathways such as linoleic acid metabolism, fructose and mannose metabolism, and taurine and hypotaurine metabolism.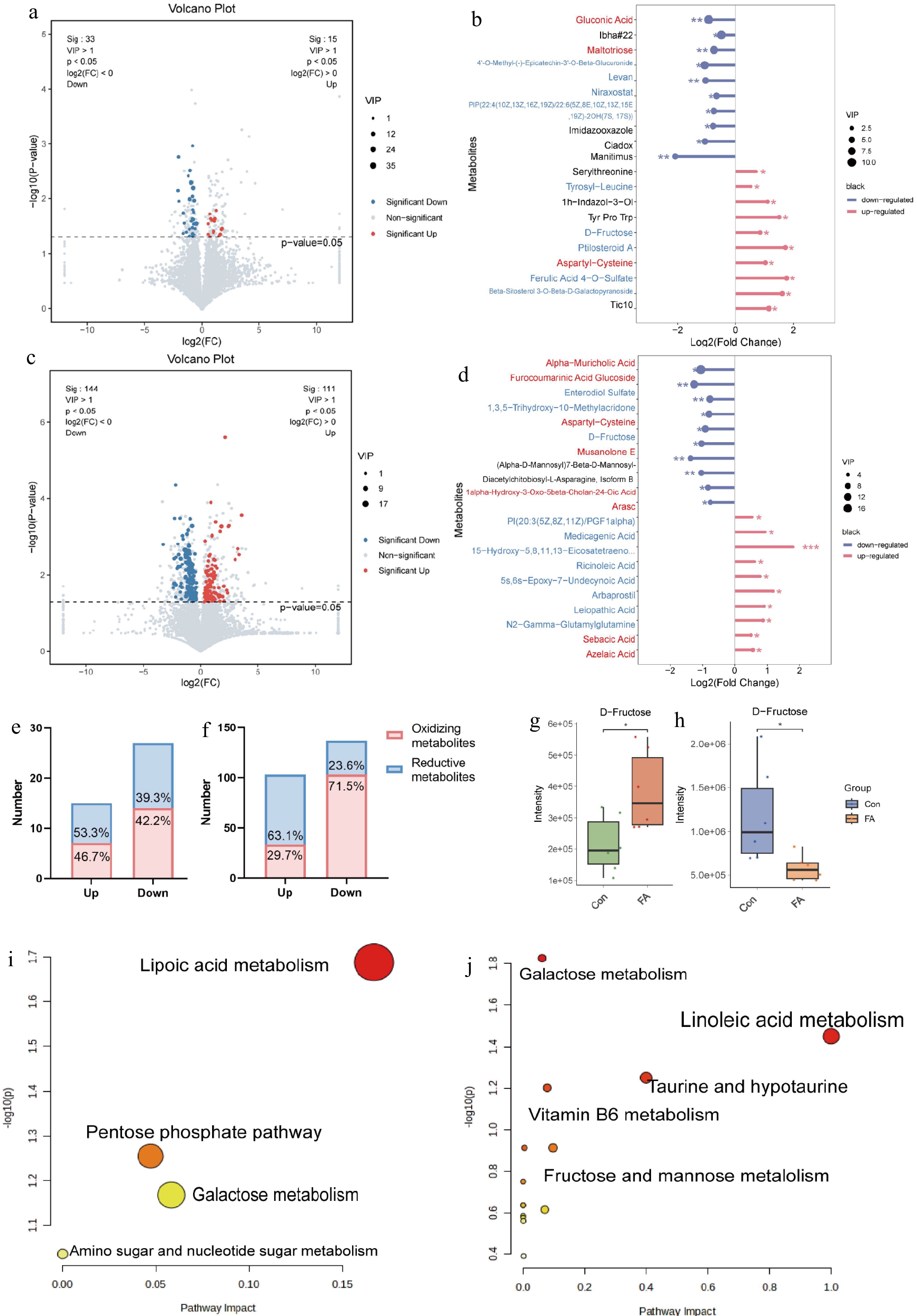
Figure 3.
Effects of FA supplementation on intestinal metabolism of rats. (a), (c) The volcano plot displays the distribution of differentially expressed metabolites of ileal and colonic digesta. Fold change (FC) was calculated by comparing with the Con group. Metabolites over the dashed line have a significant difference (p < 0.05, VIP > 1 and FC < 1). The red dots represent significantly up-regulated differential metabolites (p < 0.05, VIP > 1 and FC > 1), while the blue dots indicate significantly down-regulated differential metabolites (p < 0.05, VIP > 1 and FC < 1). (b), (d) Lolipopmaps of ileal and colonic digesta showing metabolites and their log2 (FoldChange) values. Red font indicates antioxidant; blue font indicates oxidant. An asterisk indicates the significance of differential metabolism (* indicates significance < 0.05, > 0.01; ** indicates significance < 0.01, > 0.001; *** indicates significance < 0.001, > 0.0001; **** means significance < 0.0001), and the dot size is determined by the VIP value. (e), (f) Statistical analysises of redox metabolites in ileal and colonic digesta. (g), (h) Differential expression analysises of D-Fructose in ileal and colonic digesta. (i), (j) KEGG pathway enrichment analysis on the differential metabolites identified in the ileum and colon.
Improvement of intestinal antioxidant activity by FA supplementation
-
The intestinal redox potential is closely associated with the activity of antioxidants in the intestine. FA significantly increased T-AOC level (Fig. 4a) and decreased MDA level (Fig. 4d) in ileal and colonic tissue (p < 0.05). In addition, FA significantly increased SOD (Fig. 4b) and ROS levels (Fig. 4c) in ileal tissue (p < 0.05). Pearman correlation analysis showed that the redox potential of ileum and colon were both negatively correlated with T-AOC (Fig. 4e & i). Additionally, a negative correlation between colonic redox potential and MDA level was observed (Fig. 4l), but no significant correlation in ileal was found (Fig. 4h). Furthermore, the SOD (Fig. 4f & j) and ROS (Fig. 4g & k) levels had no significant association with redox potential in both the ileum and colon. Correlation heat map analysis showed that metabolites including l-phenylalanyl-l-proline, aspartyl-cysteine, and D-fructose were positively correlated with redox potential. On the other hand, alpha-muricholic acid glucoside and furocoumarinic acid glucoside were negatively correlated with T-AOC (Fig. 4m).
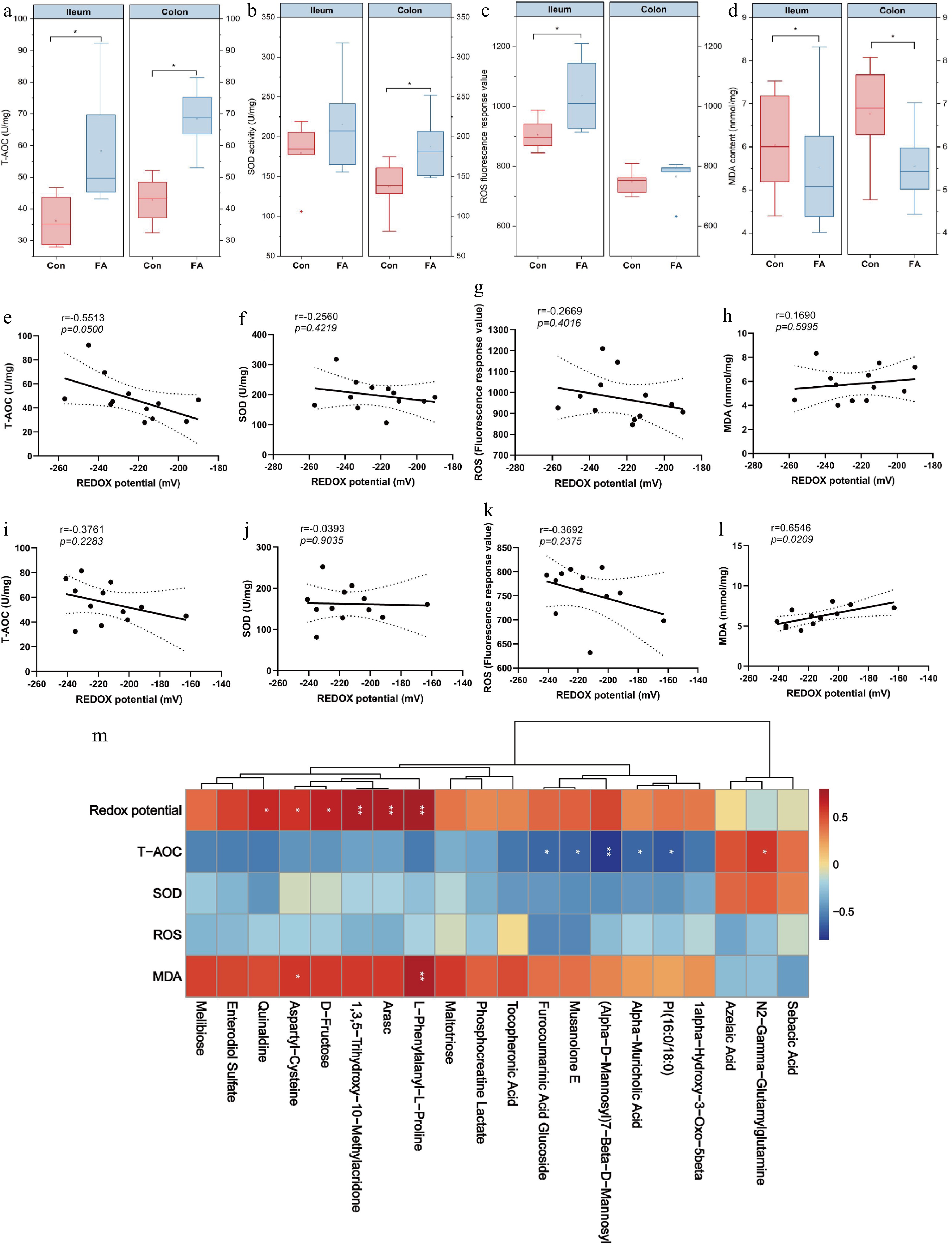
Figure 4.
Improvement of FA supplementation on intestinal antioxidant activity. (a)−(d) The levels of T-AOC, SOD, ROS, and MDA activities. (e)−(l) Correlation analysis of redox potential of ileal and colonic digesta with different antioxidant indexes. (m) Correlations between the colonic microbiota and redox potential, T-AOC, SOD, ROS, and MDA activities. The analysis is based on Pearson's correlation coefficient. T-AOC = Total antioxidant capacity; SOD = Superoxide dismutase; ROS = Reactive oxygen species; MDA = Malondialdehyde. * = p < 0.05; ** = p < 0.01.
Alleviation of diarrhea by FA supplementation in a diarrhea model
-
During the balance period (days 0−7), FA failed to show a weight gain advantage due to the limited intervention time (Fig. 5a & b). The FAMg group did not exhibit a significant increase in body weight compared to the ConMg group on both days 17 and 24 (Fig. 5a; p > 0.05). There was no significant difference in water intake among all groups throughout the experiment (Fig. 5c & d; p > 0.05). While the diarrhea index was consistently maintained at a low level and was significantly reduced on days 13 and 14 in the FA group (Fig. 5e & f; p < 0.05). The diarrhea index gradually declined to zero on days 18 to 24 following the cessation of the diarrhea model. On day 7, FA significantly reduced redox potential (Fig. 5g; p < 0.05). The redox potential of the FA group also remained at a low level during the diarrhea period (days 8-17) and decreased significantly on days 13, 14, 16, and 17 (Fig. 5h; p < 0.05). Furthermore, FA demonstrated its ability to sustain a lower redox potential during the recovery period (Fig. 5i). The correlation analysis indicated that redox potential was positively correlated with the diarrhea index (Fig. 5j; R = 0.6976; p = 0.0006).
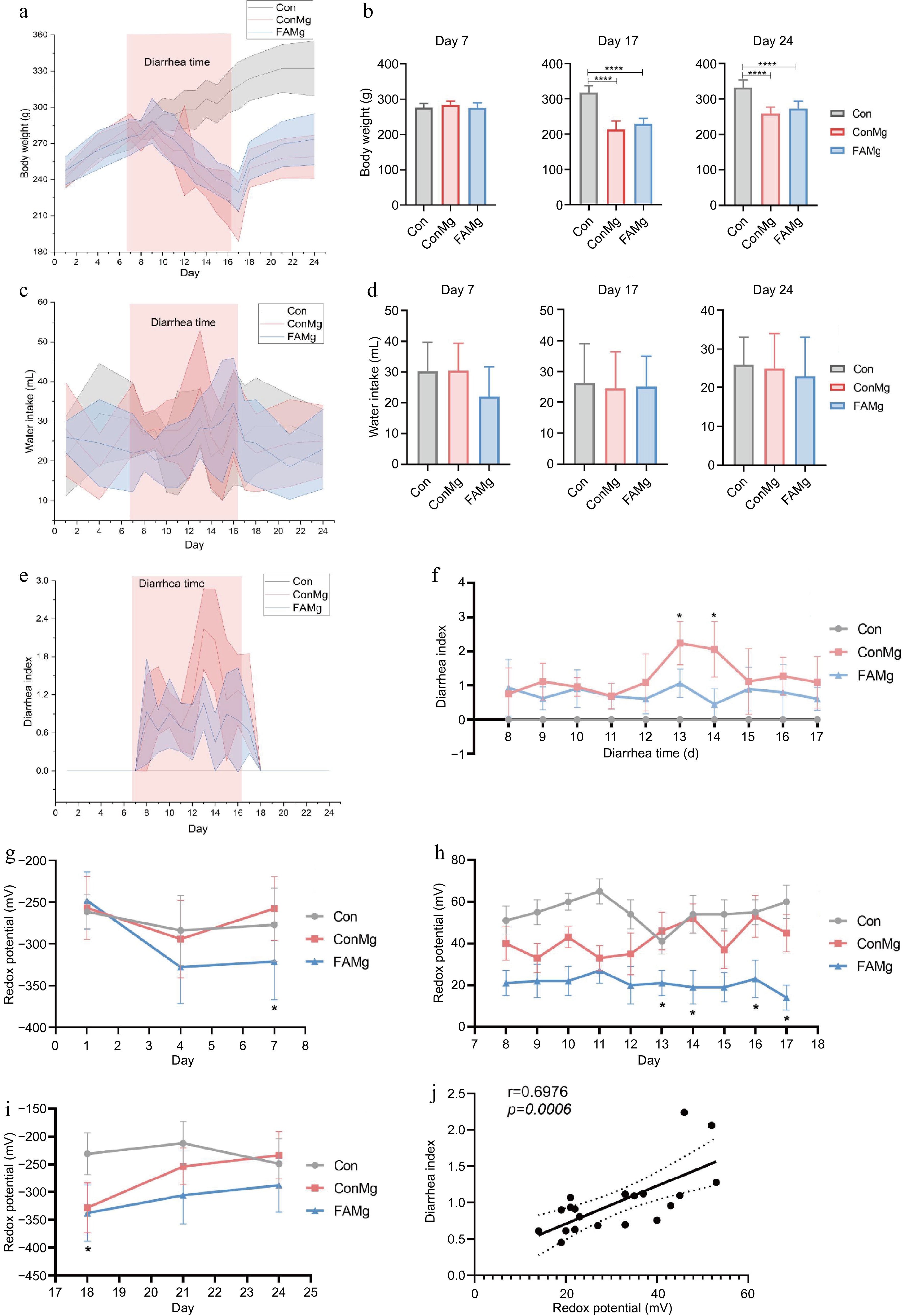
Figure 5.
Alleviation of FA supplementation on diarrhea in diarrhea model. (a) Dynamic changes in body weight. (b) Comparison of body weight changes in different days. (c) Dynamic changes in water intake. (d) Comparison of water intake change in different days. (e) Dynamic changes in diarrhea index. (f) Statistical analysis of diarrhea index. (g) Dynamic changes in redox potential in the balance period. (h) Statistical analysis of redox potential in diarrhea period. (i) Dynamic changes in redox potential in the recovery period. (j) Correlation analysis of redox potential and diarrhea index. The analysis is based on Pearson's correlation coefficient. * = p < 0.05.
Effects of FA on fecal microbiota in diarrhea model
-
FA had no significant effect on the α-diversity (Chao1 and ACE indices) on day 7 (Fig. 6a & c; p > 0.05). However, the Chao1 and ACE indices in FAMg group increased significantly compared with ConMg on day 17 (Fig. 6b & d; p < 0.05). Furthermore, from the structural composition of the microbiota (Fig. 6e), FA had no significant effect on Firmicutes but increased the abundance of Actinobacteriota on day 7 (p < 0.05). However, on day 17, the abundances of Firmicutes in ConMg and FAMg groups were significantly decreased compared to Con group (p < 0.05). Conversely, Bacteroidota expanded significantly in the ConMg group compared to the Con group on day 17 (Fig. 6f; p < 0.05).
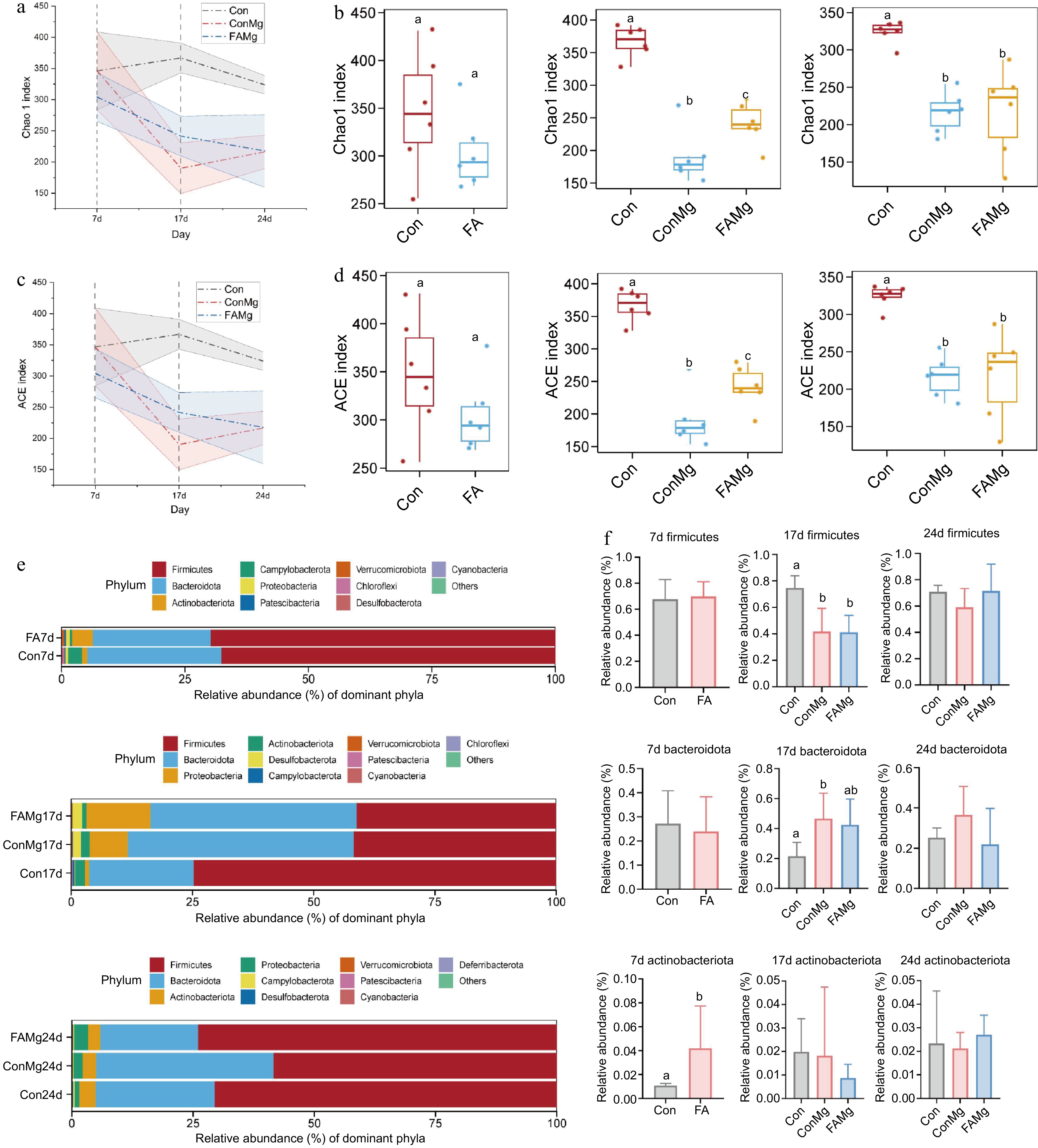
Figure 6.
Effects of FA on fecal microbiota at defferent periods of rats. (a), (c) Dynamic changes of Chao 1 and ACE indexes in each group of diarrhea model. (b), (d) Comparison of the change of Chao 1 and ACE indexes in the balance, diarrhea and recovery periods. (e) Composition of fecal microbiota at the phylum level. (f) Statistical analysis of abundances of Firmicutes, Bacteroidota and Actinobacteria in different periods.
At the genus level, the abundances of Lachnospiraceae_NK4A136_group, Helicobacter, Eubacterium siraeum group, and Eubacterium xylanophilum group were decreased significantly in the FA group (Fig. 7a; p < 0.05). During day 17, both FAMg and ConMg significantly increased the abundance of Bacteroides, while decreasing the abundance of Lachnospiraceae_NK4A136_group, Lactobacillus, and Limosilactobacillus (p < 0.05). Importantly, FAMg showed an ability to significantly increase Blautia abundance compared to ConMg (Fig. 7b; p < 0.05). During day 24, FAMg significantly reduced the abundance of Bifidobacterium, Flavonifractor and Ruminococcus torques group compared to ConMg (Fig. 7c; p < 0.05). The presence of numerous bacterial genera was identified at the genus level, followed by the prediction of influential variables using a random forest model. GCA-900066575 and Lachnospiraceae_NK4A136_group in the database displayed a higher value of contribution to enhance the redox potential and had a significant positive correlation with redox potential (Fig. 7d & e). Moreover, Negativibacillus, Adlercreutzia, UCG.005, and Ralstonia contributed substantially to reducing the redox potential.
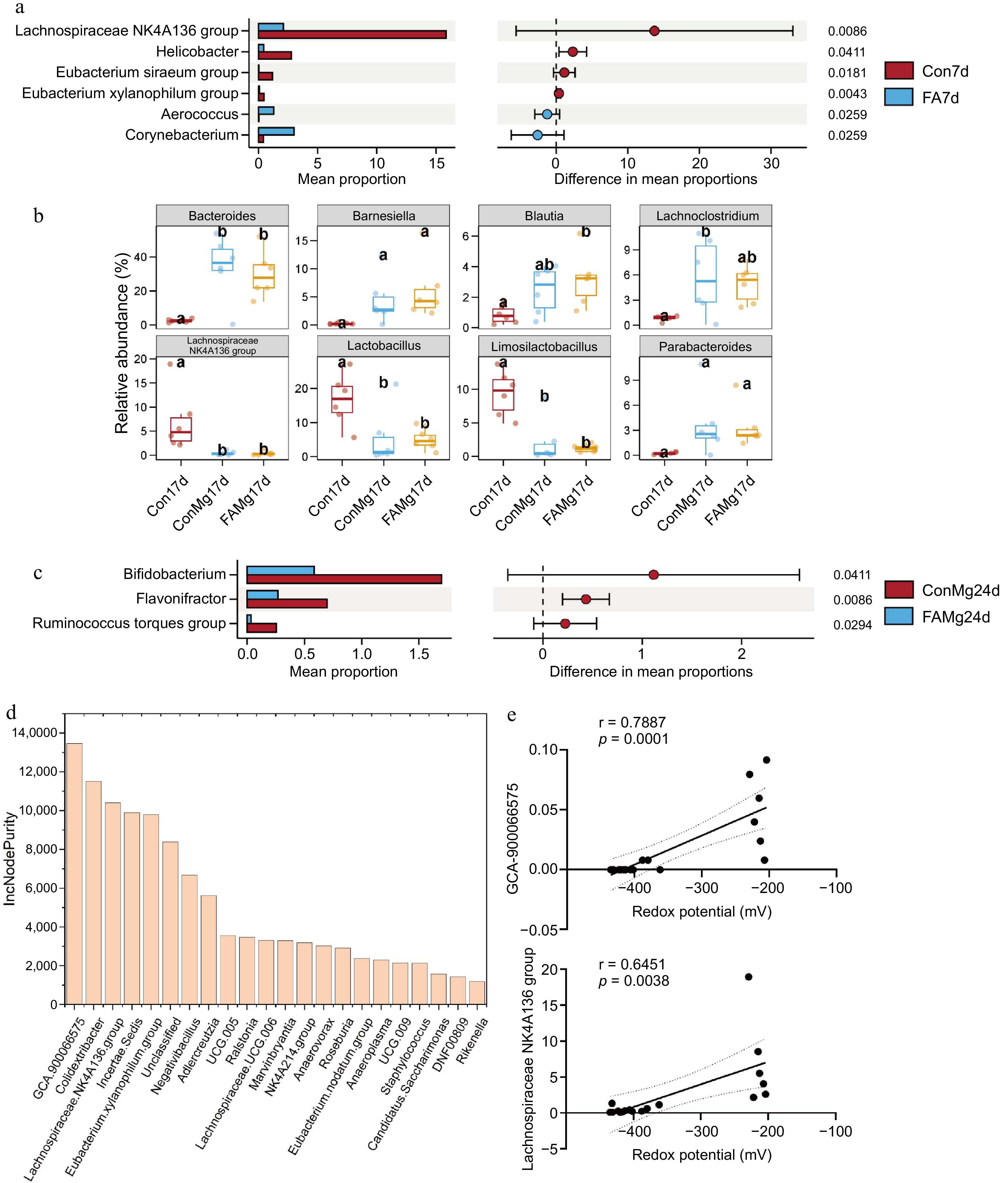
Figure 7.
Effects of FA on fecal microbiota at genus level by regulating redox potential. (a) Differential microbiota was analyzed using the Wilcoxon test in the balance period. (b) Statistical analysis on the top eight microbiota in the diarrhea period. Different letters on the column indicate significant differences, while the same letters indicate non-significant differences. (c) Differential microbiota was analyzed using the Wilcoxon test in the recovery period. (d) Significantly different microbiota was ranked in descending order of importance to redox potential prediction using random forest regression. (e) The correlation between GCA-900066575, Lachnospiraceae_NK4A136_group and redox potential. The random forest regression was calculated using the hmisc R package (v.4.5.0). Pearman's rho with asymptotic measure-specific P value between microbiota and redox potential and p < 0.05 was used to identify a significant correlation.
-
Diarrhea is a major global health problem. Studies have reported that diarrhea can cause a series of disorders such as malnutrition, vitamin deficiency, anemia, and reduced body resistance, which seriously damage body health. Some studies have found that intestinal imbalance is one of the main pathogenic factors of diarrhea[20]. Notably, in our previous study[13], a significant increase in intestinal redox potential was observed during diarrhea. Therefore, it is of great significance to reduce the redox potential by regulating the intestinal chemical environment to promote intestinal microecological stability and further reduce the occurrence of diarrhea. Thus, in the current experiment, a rat model of diarrhea was established to investigate the potential regulatory effects of antioxidant supplementation on the intestinal chemical environment to reduce the occurrence of diarrhea. In this study, the findings demonstrated that fecal administration could enhance the structure of redox metabolites in rats by regulating intestinal microflora, thereby reducing intestinal redox potential. Furthermore, preventing bacterial overgrowth during diarrhea expedited the reestablishment of intestinal microbiome composition and function after diarrhea.
A fish study has found that the addition of antioxidants in vitro can effectively decrease the intestinal redox potential, thereby inhibiting the colonization of pathogens in the intestine[11]. Any substance or compound that scavenges oxygen free radicals or inhibits cellular oxidation processes is considered an antioxidant[21]. Antioxidants play a crucial role in human health by inhibiting or delaying adverse oxidative reactions, thereby preventing the development of diseases associated with oxidative stress[22]. However, assessing antioxidant capacity is a complex task, as no single method can fully capture the natural reactions occurring in the body[23]. Various methods have been employed to evaluate antioxidant activity, including those based on scavenging stable free radicals (such as DPPH, ABTS, and FRAP)[24]. Although both DPPH and ABTS methods are relatively simple and feasible, their chemical properties limit their applicability in biological environments. On the other hand, FRAP and ABTS methods demonstrated good correlation and consistency in results[25].
Over the past few decades, extensive studies have been conducted on the antioxidant activity of polyphenols. Numerous studies have been published demonstrating their protective effects against various diseases caused by oxidative stress, including neurodegenerative diseases[26], diabetes[27], cardiovascuar disease[28], and allergies[29]. Recent investigations have also unveiled other mechanisms by which polyphenol antioxidants regulate the gut microbiome[30]. However, these protective effects are primarily attributed to their ability to directly scavenge free radicals such as oxygen and nitrogen species, which is dependent on the presence of intramolecular phenolic hydroxyl groups that generate stable phenoxyl radicals and their spatial arrangement i.e., electronic structure[31,32]. Polyphenol antioxidants, such as FA and resveratrol, exhibited significant efficacy in reducing redox potential, with comparatively higher FRAP-based in vitro antioxidant activity. The association between FARP and redox potential was assessed according to Miliciac et al.[33], and the results demonstrated that the ratio of FA-induced changes in redox potential to FRAP was the highest, indicating that FA had a significant effect on reducing intestinal redox potential.
Further assessment of ileum and colonic digesta confirmed that FA exhibited superior efficacy in reducing redox potential. Intestinal redox potential serves as a crucial internal environmental factor. To comprehend the effect of FA on intestinal redox potential, 16S rRNA gene sequencing analysis was conducted to investigate the dynamics of microbial community reestablishment in rat feces across ileal, colonic digesta, and diarrhea models following FA treatment. In the absence of diarrhea, FA supplementation did not significantly affect the α-diversity of intestinal microbiota. Similarly, previous studies have reported that although FA does not significantly affect the α-diversity of intestinal microbiota, it does reshape its composition[34]. Furthermore, the present results demonstrate a significant alteration in the β-diversity of intestinal microbiota induced by FA. FA supplementation significantly reduced the abundance of Lachnospiraceae_NK4A136_group, a member of the Lachnospiraceae family, while increasing the prevalence of Christensenellaceae_R-7_group, a genus in the Christensenellaceae family. Lachnospiraceae has been reported to be effective against age-induced oxidative stress and inflammation[35], whereas Christensenellaceae_R-7_group is considered a reliable biomarker for overall health in organisms[36].
Metabolome sequencing of ileal colon contents was then utilized to evaluate the impact of FA supplementation on intestinal redox status. Gut microbes communicate through metabolites, but the exact effect of these metabolites on gut redox potential remains largely unexplored[37]. However, the redox activity exhibited by these microbial metabolites directly reflects the redox state of the gut[38]. The present findings suggest that FA supplementation significantly increases the proportion of reduced metabolites in both the colon and ileum, thereby maintaining a lower intestinal redox potential. These results are consistent with the findings of previous studies[39]. To further understand the changes in oxidative and reductive metabolism, the analysis of KEGG pathway enrichment of differential metabolites demonstrated that the differential metabolites were predominantly enriched in lipoic acid metabolism and pentose phosphate pathway, as well as galactose metabolism and amino sugar and nucleotide sugar metabolism in the ileum. On the other hand, the differential metabolites in the colon were significantly enriched in metabolic pathways such as linoleic acid metabolism, fructose and mannose metabolism, and taurine and hypotaurine metabolism. Furthermore, they enhance the reductive metabolism in the intestine by increasing intestinal metabolism[40]. However, the causal effect of FA on host response and reduced redox potential remains unclear, even though the reduced gut redox potential has been associated with gut microbiota remodeling and metabolite changes due to FA supplementation. It is known that intestinal redox potential is often influenced by host-related factors[41]. For instance, during periods of stress, intestinal antioxidant active substances are significantly reduced, thereby decreasing intestinal resistance[42]. This study focused on examining the correlation between FA supplementation and antioxidant status as well as redox potential. Additionally, the relationship between metabolites and redox potential was investigated. FA supplementation can enhance intestinal antioxidant activity and is associated with intestinal redox potential. Interestingly, it was discovered that metabolites linked to both redox potential and total antioxidant capacity in the gut did not overlap, suggesting that host-related factors were not primarily responsible for reducing FA-induced redox potential.
In the case of diarrhea, FAMg slowed down the decline in gut microbiota α-diversity and facilitated a faster recovery from significantly reduced levels compared to ConMg. To investigate the examined process of the disrupted gut microbiome during diarrhea, both its composition and function were examined[43]. The average abundance of Lactobacillus and Limosilactobacillus in the FAMg group was more similar to that in the Con group. Limosilactobacillus has been demonstrated to have a potent antioxidant mechanism against oxidative stress and its associated chronic ailments[44]. In the meantime, empirical studies have demonstrated that Limosilactobacillus can effectively mitigate the symptoms of diarrhea and colon inflammation induced by antibiotics and concurrently facilitate the normal expression of colon immune factors[45], indicating that the gut microflora can cascade with the body's immune system to fight against the disease in the case of diarrhea. Importantly, FAMg demonstrated the ability to significantly enhance Blautia abundance compared to ConMg. Studies have shown that Blautia, one of the most prevalent and crucial acetogenic bacteria in the gut, could alleviate depression and accelerate the progression of breast cancer[46]. Consequently, a decrease in gut redox potential leads to an increase in the abundance of key probiotics in the gut during diarrhea. To elucidate the microbial contributors and their associated functions in diarrhea following FA supplementation, the bacterial genera responsible for redox potential regulation was identified. Notably, Lachnospiraceae_NK4A136_group made a significantly higher contribution to redox potential regulation. Interestingly, the abundance of Lachnospiraceae_NK4A136_group was found to be markedly reduced in the FA-treated group during in vivo screening and subsequent recovery period after diarrhea. Lachnospiraceae_NK4A136_group belongs to sporogenic anaerobic bacteria of the trichospirillaceae family. Lachnospiraceae_NK4A136_group produces SCFAs through fermentation of dietary polysaccharides, which is negatively associated with a variety of metabolic diseases and chronic inflammation[47]. The abundance of Lachnospiraceae_NK4A136_group was positively correlated with redox potential. Therefore, Lachnospiraceae_NK4A136_group may represent a bacterial genus that potentially regulates the redox potential in the intestinal environment. However, the important role of Lachnospiraceae_NK4A136_group in regulating intestinal redox potential and its mechanism still needs to be further studied.
-
In summary, the present study demonstrated that FA supplementation reduced diarrhea by decreasing the intestinal redox potential. Furthermore, it has been observed that FA-regulated gut microbiota and metabolites contributed to a more rapid reduction in intestinal redox potential and enhanced microbial reductive metabolism in rats after diarrhea. This study uncovered the potential of FA in alleviating diarrhea and highlights the interplay between FA-regulated gut microbiota and changes in redox potential during post-diarrhea processes. The present findings provide novel insights into the mechanisms by which FA regulates intestinal redox potential and identify potential strategies for mitigating diarrhea.
This research was supported by the National Natural Science Foundation of China (32272891) and the National Key R&D Program of China (2022YFD1300402).
-
All procedures were reviewed and preapproved by the Animal Care and Use Committee of Nanjing Agricultural University, identification number: SYXK2019-0066, approval date: 2023-05-09, and implemented based on the standard of Experimental Animal Care and Use Guidelines of China, identification number: EACUGC2018-01. The research followed the 'Replacement, Reduction, and Refinement' principles to minimize harm to animals. This article provides details on the housing conditions, care, and pain management for the animals, ensuring that the impact on the animals is minimized during the experiment.
-
The authors confirm contribution to the paper as follows: data curation: Feng N, You J, Wang D, Li L; formal analysis: Feng N, Wang D; writing - original draft: Feng N, You J; writing - review: Feng N; writing - editing: Feng N, You J, Su Y, Feng X; investigation: You J; resources: Xu R; validation: Xu R, Chen L; conceptualization, supervision: Su Y, Feng X; project administration, funding acquisition: Su Y. All authors reviewed the results and approved the final version of the manuscript.
-
The raw data of 16S rRNA gene sequencing in the study have been deposited in the repository of the Sequence Read Archive, with number SUB14680623. The data of metabolome analysis have been included as part of the supplementary material.
-
The authors declare that they have no conflict of interest. Lian Li and Yong Su are the Editorial Board members of Animal Advances who were blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of these Editorial Board members and the research groups.
-
# Authors contributed equally: Ni Feng, Jinwei You
- Supplementary Information 1 Photos of three Pulsatilla specimens.
- Supplementary Information 2 Superimposed diagram of total ion current (TIC) diagram of mass spectrometry detection of QC samples. (a: positive ion mode. b: negative ion mode.
- Supplementary Information 3 CV distribution diagram of samples in each group.
- Supplementary Information 4 Schematic diagram of mass spectrometry multiple reaction monitoring mode.
- Supplementary Information 5 Total ion current chromatogram of mass spectrometry analysis of mixed samples (a: positive ion mode. b: negative ion mode).
- Supplementary Information 6 Multi-pea k chromatogram of metabolite detection in multiple reaction monitoring (MRM) mode (a: negative ion mode. b: positive ion mode).
- Supplementary Information 7 Metabolite quantitative analysis integration correction diagram (Randomly select 24 metabolites.).
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Feng N, You J, Xu R, Chen L, Wang D, et al. 2024. Ferulic acid reduces gut redox potential and alleviates chronic Mg-induced diarrhea by modulating gut microbiota and metabolites. Animal Advances 1: e004 doi: 10.48130/animadv-0024-0004 

Ferulic acid reduces gut redox potential and alleviates chronic Mg-induced diarrhea by modulating gut microbiota and metabolites
- Received: 10 September 2024
- Revised: 10 October 2024
- Accepted: 11 October 2024
- Published online: 08 November 2024
Abstract: Ferulic acid, a phenolic antioxidant, is widely utilized for its antioxidative properties. The roles of ferulic acid in regulating intestinal redox potential and alleviating chronic diarrhea were investigated in this study. Through in vitro antioxidant activity and in vivo tests, ferulic acid exhibited a more pronounced capacity in reducing the intestinal redox potential of rats than other common antioxidants. Meanwhile, ferulic acid significantly enriched the abundance of Actinobacteriota while reducing the Lachnospiraceae_NK4A136_group in the colon. Metabolomics analysis revealed that ferulic acid treatment resulted in a two-fold increase of reductive metabolites in the colon, thereby contributing to the maintenance of a lower intestinal redox potential. A diarrheal rat model was induced by magnesium citrate to validate the roles of ferulic acid in alleviating chronic diarrhea. Ferulic acid effectively reduces the diarrhea index to prevent the expansion of Bacteroidota by maintaining a low intestinal redox potential. Simultaneously, ferulic acid significantly enhanced the abundance of Blautia during diarrhea, facilitating a more rapid recovery of intestinal microbiota. Lachnospiraceae_NK4A136_group made a notable contribution to the high redox potential. The findings suggest that redox potential is a promising target for alleviating diarrhea, and provides novel insights into the mechanism by which ferulic acid regulates the health of animals or humans.
-
Key words:
- Diarrhea /
- Ferulic acid /
- Gut microbiota /
- Redox potential