









-
In plant establishment and developmental processes, sugars have diverse functions, including serving as carbon skeletons, acting as substrates for respiratory reactions, and playing important roles in osmotic regulation processes and signal transduction[1,2]. Moreover, they also contribute to stress resistance activities, transient energy storage, and molecular transport[1−3]. Leaves synthesize sugars to cater to the needs of nonphotosynthetic organs such as fruits, seeds, and roots for adequate plant growth[4,5]. The transport and dissemination of fructose, sucrose, and glucose require the engagement of sugar transporters. These transporters mediate the transfer of sugars across source–sink organs, which is crucial for sugar homeostasis and the cellular exchange of sugars in organisms[6−8]. The last categories of sugar transporters are separated into three groups: sucrose transporters (SUTs), monosaccharide transporters (MSTs), and sugars. Sugars are eventually exported from transporter (SWEET) families, which execute and supply sucrose to targeted plant tissues and organs via transport from plant leaves. Then, they break up into monosaccharides, which are subsequently transported by monosaccharide transporter proteins[5,9,10]. Moreover, the major facilitator superfamily transporters include SUTs and MSTs and initially engage in the influx of sugar into the cytosol. Furthermore, vacuolar glucose transporter (vGTs) and tonoplast sugar transporter (TST), which are MSTs, are engaged in providing sugars to vacuoles from the cytosol and act as H+/sugar antiporters[11]. MSTs are also localized in the membranes of different organelles, such as vacuoles, chloroplasts, the Golgi, and plasma membranes[12,13]. In addition, MSTs are further subdivided into seven groups: vacuolar glucose transporter (VGT), plastidic glucose transporter (GlcT)/suppressor of G protein beta 1 (SGB1), sugar transporter protein (STP), polyol transporter (PLT), tonoplast monosaccharide transporter (TMT), early response to dehydration-6-like (ERD6L) and inositol transporter (INT). These STPs are the most well-characterized subfamily and function in transporting hexose from the apoplastic space into the cell[14,15]. Additionally, SUTs and MSTs contain 12 transmembrane α-helices and mediate the membrane transport of various sugars[1,16,17]. Unlike MSTs and SUTs, SWEETs are pH-independent transpopters. These sugar transporters play significant roles in phloem transport and work as bidirectional transmembrane transporters of sugars along a concentration gradient[7].
Sucrose is the major form of assimilated carbon that is generated during photosynthesis in both monocot and dicot plant species[10]. Long-range sucrose distribution from source tissues (leaves) to sink tissues is mediated through the phloem. Two groups of sucrose transporters, the SWEET and SUC families, play central roles. Since SWEET-promoting factors excrete sugar, the active load is dependent on SUC, and SUC utilizes the proton motive force of the plasma membrane to promote sucrose uptake and accumulation, with 1.3 molar concentrations in the phloem[18−20]. This SUC activity causes phloem sap movement through high turgor pressure. Moreover, fruits and seeds, as sink organs, accumulate high sugar concentrations at developmental stages through SUC activity[21,22]. Even though SUCs are essential for many basic processes in plant physiology, their exact method of action has yet to be determined. However, how SUCs detect sucrose and how proton coupling occurs during transport are not well understood. The sucrose binding site needs to have unique features to release sucrose into the environment at high sucrose concentrations; in fact, it needs to have tight and well-coordinated coupling to the driving proton gradient[23].
Notably, the SWEET proteins play a role in various sugar efflux-related processes, including nectar secretion, phloem loading, symbionts, and maternal efflux for filial tissue development[24]. In addition, SWEETs can be hijacked via pathogens to obtain nutrients from hosts[25]. SWEETs have been demonstrated to impact different physiological mechanisms, including the modulation of gibberellin responses[26], cold tolerance[27], nectar secretion[28], seed and fruit development[29], pollen development[30], and senescence[31]. The SWEET genes in plants are expressed in response to stress conditions, which illustrates the possible association of these plant genes with defense mechanisms under stress conditions[8,9]. The first identified gene encoding a glucose transporter was AtSWEET1, whose function has been characterized[19]. A phylogenetic tree of plant SWEETs revealed that SWEET members were grouped into four clades (clade I, clade II, clade III, and clade IV) with distinct characteristics. SWEETs in clade I and clade II preferentially transport hexoses, members of clade III mainly prefer sucrose, and members of clade IV are efficient fructose transporters[2,6,19,32]. For instance, Clade II AtSWEET8 is associated with pollen viability, and Clade III AtSWEET15 is associated with leaf senescence[7]. OsSWEET4/11 and ZmSWEET4 have been shown to play significant roles in filling grains in rice and maize, respectively[29,33]. SWEETs (also known as the PQ-loop repeats) are a recent type of sugar transporter that is a member of the MtN3/saliva family[19,34]. Thus far, SWEET genes have been reported in various plant species, including Glycine max[35], Citrus sinensis[36], Arabidopsis thaliana[19], Vitis vinifera[37], Oryza sativa[38], Malus pumila[39], Eriobotrya japonica[40], Lycopersicon esculentum[41], and Litchi chinensis[42], besides, Hemerocallis fulva, Triticum aestivum, Camellia sinensis, Medicago truncatula, Vitis vinifera, and Brassica rapa. The SWEET homologs contain seven α-helical transmembrane domains (TMs) and belong to the MtN3/saliva family: a tandem repeat of three transmembrane domains (TMs) connected with a linker-inversion TM[7,43]. Currently, genome-wide studies of SWEET genes have been carried out in various plant species, including Punica granatum[44], A. thaliana[19], Oryza sativa[38], and Litchi chinensis Sonn[42].
Sugar beet (Beta vulgaris L.) is in the family Chenopodiaceae. The chromosome number of cultivated sugar beet was 2n = 2x = 18[45]. Sugar beet is a biennial root crop with a prominent ability to produce sugar when used as a pure sugar feedstock. Sugar beet cultivation is suitable in temperate climate regions[46,47]. Various biotic stresses, such as fungal, nematode, bacterial, and viral stresses, threaten the productivity of sugar beets[48−50]. The sequencing of the Beta vulgaris genome was conducted in 2013[51], during which great molecular information, such as transporter information, was obtained. In addition to the development of molecular tools, molecular breeding methods have been employed to improve resistant genotypes, which leads to enhanced production while significantly decreasing costs, time, and effort. A wide range of genomic resources for sugar beet have been generated by sequence-based genetic methods. High-resolution genetic and physical maps have been constructed using single nucleotide polymorphisms (SNPs)[52,53]; moreover, to detect stress-responsive genes and reveal important metabolic pathways, transcriptome profiles have been studied[46,54]. Moreover, various sugar beet genomes, such as chromosome-level assemblies of the double-haploid line RefBeet, have been assembled[51], and genome-wide characterization has been carried out for the study of several functional genes, such as R2R3-MYB genes, the BZR transcription factor family and Na+/H+ antiporter (NHX) genes[55−57]. Despite the substantial progress in molecular and structural studies associated with sugar transporters, the story is imperfect: genome-wide studies and new approaches and tools are needed to increase sugar accumulation in target organs. This work provides insight into the evolution, structural characteristics, expression, and regulation of all sugar transporter gene families in Beta vulgaris. The goal in this study was to comprehensively identify and characterize all sugar transporters in B. vulgaris and compare them based on sequence structure and evolution. This analysis uncovers an initial step towards exploring the function of the B. vulgaris SUTR gene family further.
-
The protein sequences of sugar transporters from the model plant Arabidopsis thaliana were used as queries in the BLASTp program against the genome of Beta vulgaris via the Enesmbl Plant database[58]. In addition, sugar transporter family members were identified in S. spontaneum using the BLASTp program of the NCBI database, similar to previous methods. The presence of sugar transporter domains was checked for in all the collected proteins using the CDD tool. The physicochemical properties of SUTRs, such as molecular weight (MW), isoelectric point (pI), GRAVY, instability, and protein length, were evaluated via the ProtParam tool[59].
Evolutionary analysis of SUTRs
-
The SUTRs of proteins from Beta vulgaris (BvSUTR), Arabidopsis thaliana (AtSUTR), and Saccharum spontaneum were aligned using Clustal Omega[60]. The phylogenetic analysis was accomplished based on the maximum likelihood method and bootstrap test was adjusted to 1,000 times using the IQ tree online tool[61]. Finally, the phylogenetic tree was constructed with the iTOL online tool[62]. Duplication events between BvSUTRs were predicted based on cDNA similarity between paired genes using MEGA X[63]. In addition, Ka and Ks values of duplicated genes were calculated using TBtools[64]. In addition, the divergence time of duplicated BvSUTRs was predicted by the following equation: T = (Ks/2λ) × 10−6, (λ = 6.5 × 10−9).
Protein structure analysis of SUTR family members
-
Predicting the three-dimensional structure of proteins can be effective in comparing proteins and determining differences. In this study, the three-dimensional structure of BvSUTR proteins was estimated using the Phyre2[65] database based on maximum homology with known models. In addition, the binding sites of each protein were predicted by the Phyre investigator tool Phyre2. In addition, transmembrane structural domains of BvSUTR proteins were predicted using the TMHMM online tool[66].
Expression profile of SUTRs
-
The expression profiles of the BvSUTR genes were investigated based on RNA-seq data. The expression patterns of these genes were identified based on their orthologs in Arabidopsis using the eFP Browther (
http://bar.utoronto.ca/efp//cgi-bin/efpWeb.cgi ). In this study, the expression patterns of genes in response to abiotic stresses such as cold (continuously at 4 °C), salinity (150 mM NaCl), drought (rafts were exposed to the air stream for 150 min), wounding and heat (for 3 h at 38 °C) in shoot and root tissues were investigated. Additionally, expression data related to biotic stresses, such as treatment with Botrytis cinerea (B.c), Pseudomonas syringae (P.s), Phytophthora infestans (P.i), Erysiphe orontii (E.o), and flagellin 22 (FLG22; as an elicitor), were analyzed. The extracted data were based on FPKM (fragments per kilobase of exon model per million mapped reads) values, and log2-fold changes (treated/control) were calculated.Promoter analysis of SUTR genes
-
Analysis of promoter regions provides a good understanding of the regulatory system of each gene in response to upstream signaling elements. In the present study, the region 1,500 bp before the start codon was screened as a promoter site to identify cis-regulatory elements using the Plant CARE tool[67]. The putative cis-regulatory elements were classified based on their function.
Interaction network of SUTRs
-
The study of interaction networks can reveal information related to the functions of proteins in possible interactions and involved cellular pathways. In the present study, an interaction network of BvSUTRs was constructed using the String database based on available data from the model plant Arabidopsis[68]. In this study, the first layer was fixed on five nodes, and the second layer was adjusted to less than 10 nodes. Moreover, the significant GO (gene ontology) terms (FDR ≤ 0.05) related to interaction nodes were identified in terms of biological processes, molecular function, and cellular components.
-
In the present study, a total of 61 SUTR genes were identified in the genome of Beta vulgaris and named BvSUTR1−BvSUTR61. The length of the BvSUTR family members ranged from 171 (BvSUTR03) to 748 amino acid (aa) (BvSUTR31). The isoelectric point (pI) varied between 4.93 (BvSUTR43) and 9.84 (BvSUTR61). Based on the pI values, most of the BvSUTRs (82%) were predicted to be basic proteins, with a pI ≥ 7.0 (Supplementary Table S1). The instability index for 43 (70%) BvSUTRs was less than 40, indicating that most BvSUTRs are stable proteins. Prediction of subcellular localization revealed that most BvSUTRs are located in the plasma membrane, chloroplast, and vacuole (Supplementary Table S1, Fig. 4a). Based on the physicochemical properties of proteins, predicted family members may play different functions.
Evolutionary analysis of SUTRs
-
Phylogenetic analysis separated BvSUTRs into five groups based on their evolutionary relationships (Fig. 1). According to the phylogenetic tree, 11 BvSUTRs in group I, 12 BvSUTRs in group II, eight BvSUTRs in group III, 14 BvSUTRs in group IV, and 16 BvSUTRs in group V were located. Most BvSUTRs from groups I and II had sugar/inositol transporter domains, and BvSUTRs from group III possessed sucrose/H+ symporter domains. In addition, subgroup IVa had a nucleotide-sugar transporter domain, and BvSUTRs from subgroup IVb had a sugar phosphate transporter domain. All the BvSUTRs with sweet sugar transporter domains were located in group V. The number of exons/introns is among the factors that are affected by evolutionary events and can also affect the speed of the process of gene expression[69,70]. The bvSUTR genes from group II had the most exons, and the members of group I had the least number of exons (Fig. 2). The pI ranged little among the different groups of BvSUTRs, and the members of groups II and III exhibited greater diversity and had different pI ranges (Fig. 4b). Chromosome mapping analysis revealed that a total of 61 BvSUTR genes were distributed on nine chromosomes of Beta vulgaris, seven of which were located on unknown chromosomes (Fig. 3). Most BvSUTRs (13 genes) were observed on chromosome 4. Two duplication events were recognized between BvSUTR38−BvSUTR40 and BvSUTR57−BvSUTR61 (Fig. 3).
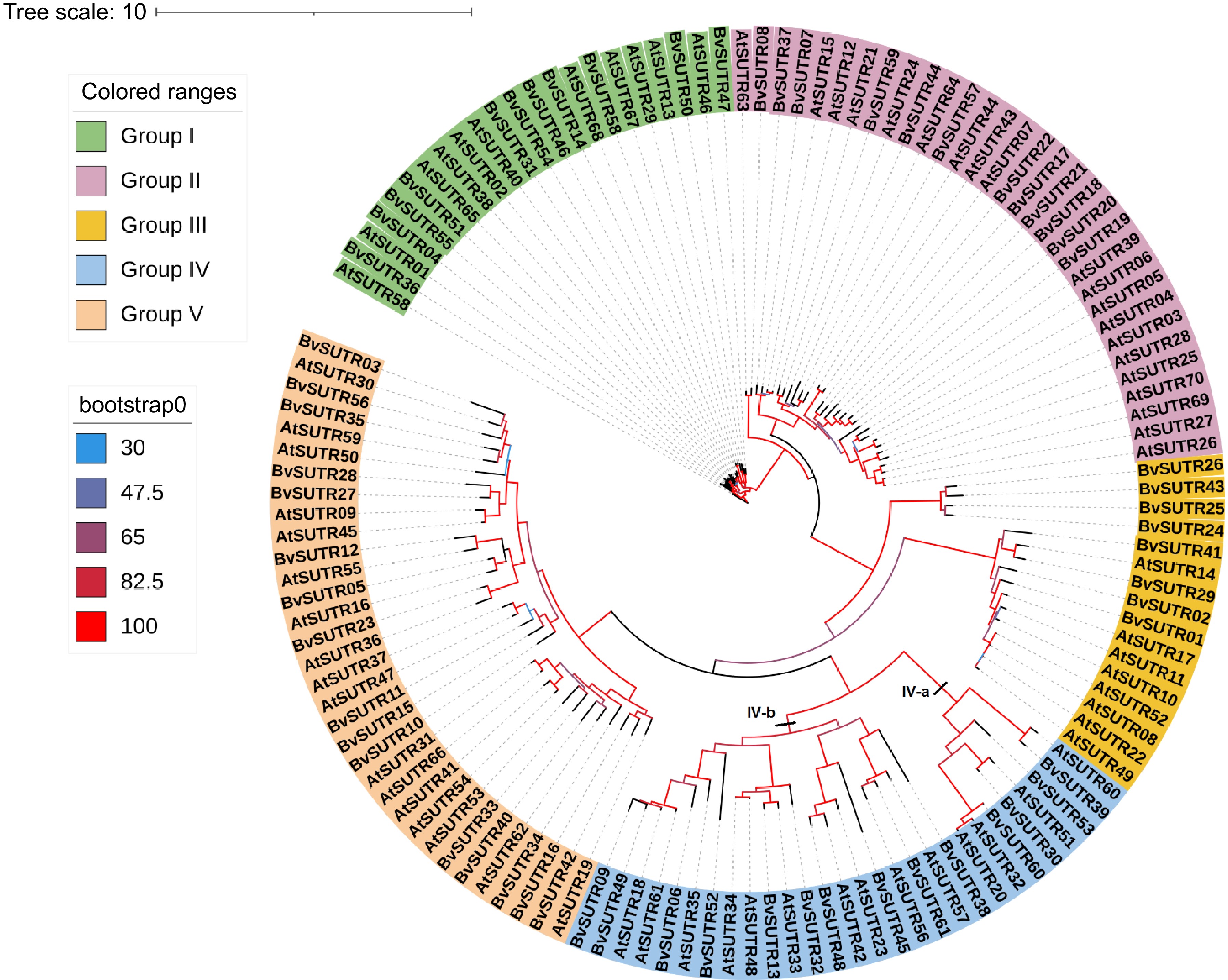
Figure 1.
Phylogeny tree of the SUTR family in Beta vulgaris (starting with Bv) and the dicot model plant Arabidopsis thaliana (starting with At).
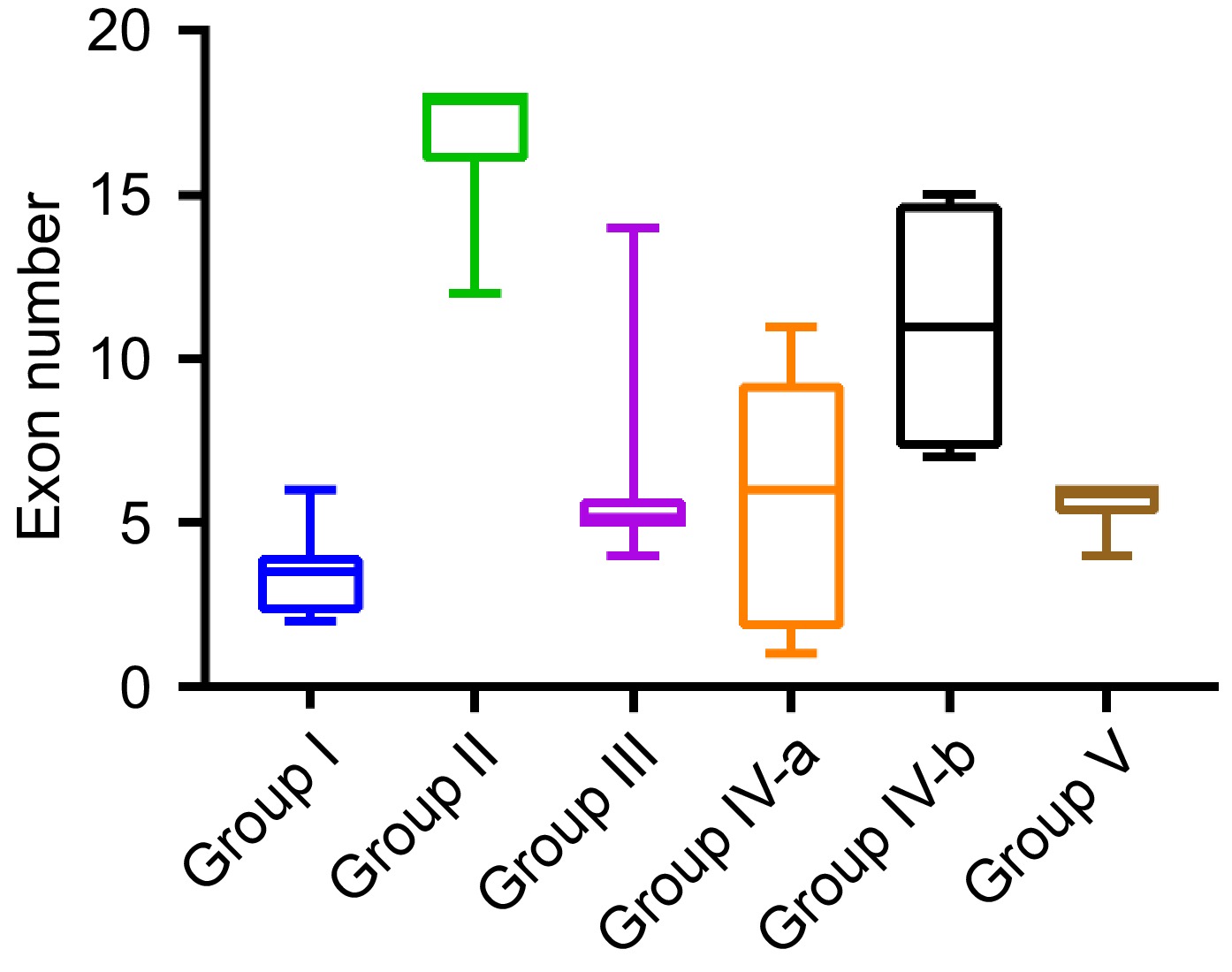
Figure 2.
Range of exon numbers in BvSUTR genes based on phylogenetic analysis.
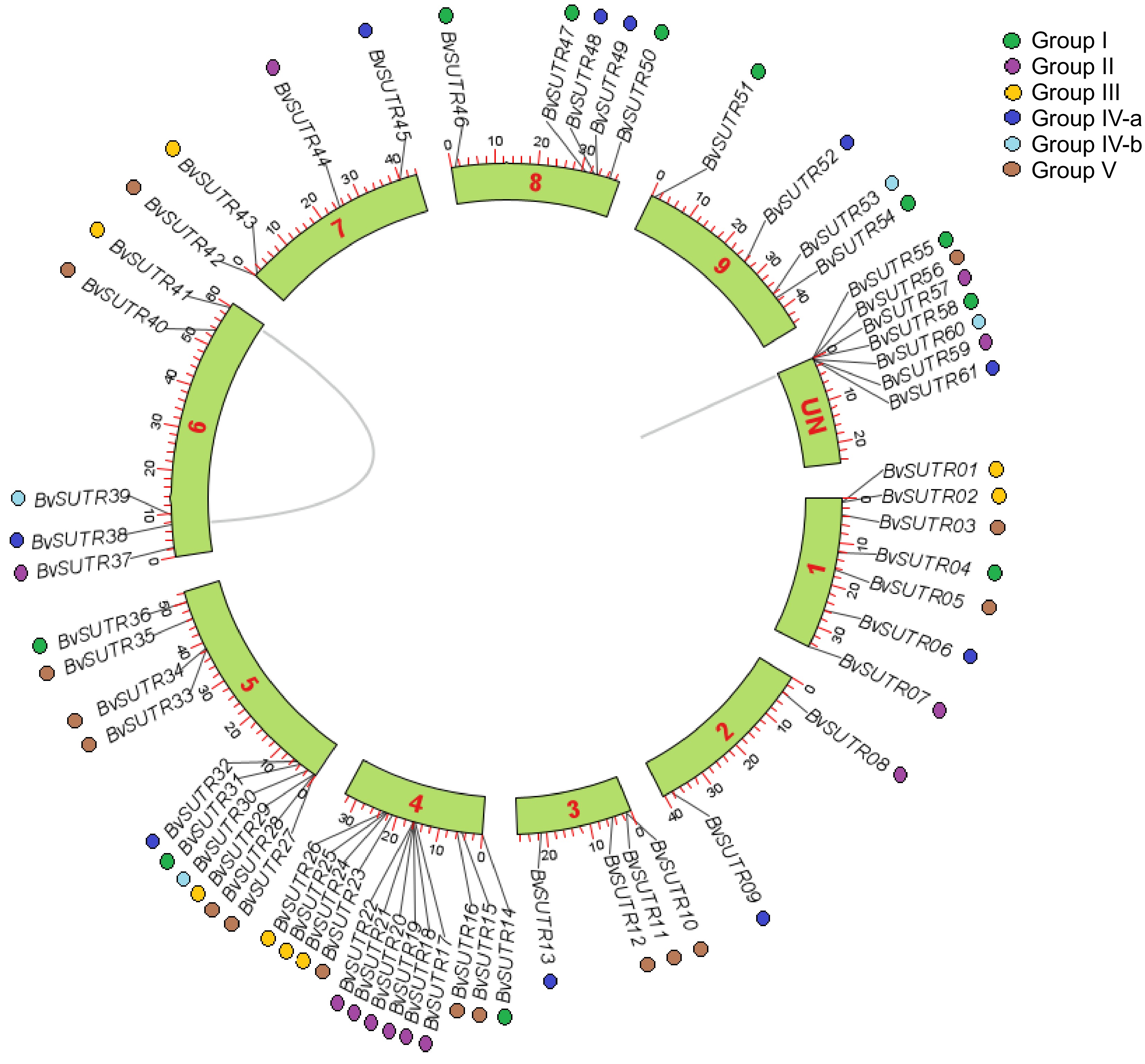
Figure 3.
Distribution of BvSUTR genes in the genome of Beta vulgaris.
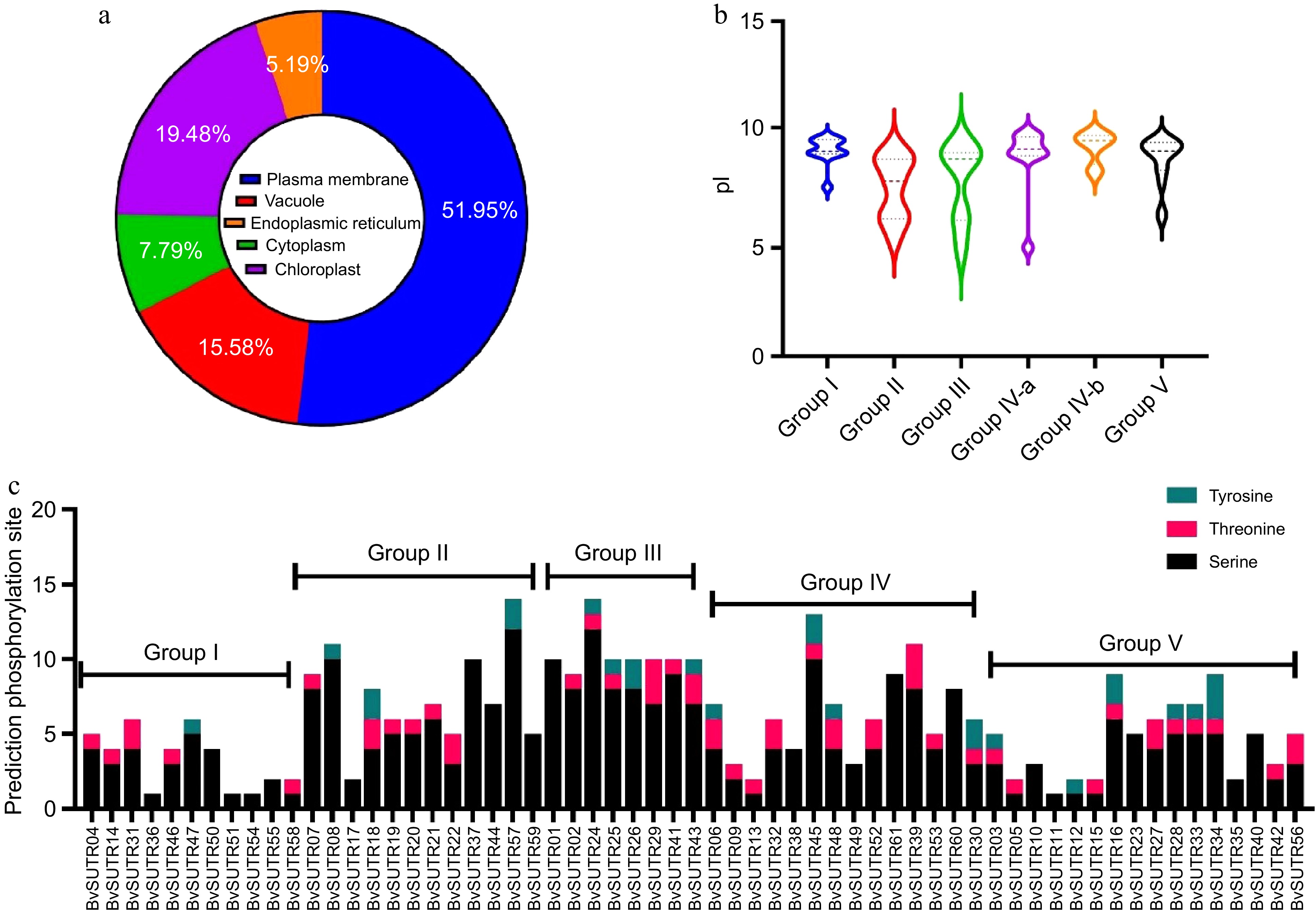
Figure 4.
Prediction of the subcellular localization of (a) BvSUTRs, (b) the range of isoelectric points (pIs), five groups of BvSUTRs, (c) and the prediction of phosphorylation sites in BvSUTRs.
Phosphorylation site in BvSUTRs
-
Posttranslational modifications were investigated based on the prediction of phosphorylation regions in BvSUTR proteins (Fig. 4c). The range of phosphorylations varied from 1 (BvSUTR36, 51 and 54 from group I and BvSUTR11 from group V) to 25 (BvSUTR24 from group III and BvSUTR57 from group II). In addition, members of group III, which included the sucrose/H+ symporter, exhibited increased potential for phosphorylation, while members of group I (which included the sugar/inositol transporter domain) and group V (which included the SWEET sugar transporter domain) were predicted to have fewer phosphorylation sites.
Prediction of the transmembrane structure of SUTRs
-
Membrane proteins such as transporters are involved in many essential processes, including regulation, signaling and transport. Determining the structure of this group of proteins and how they are located in the membrane can provide a better understanding of their function and enable comparisons between them. In the present study, diverse transmembrane structures of BvSUTRs were predicted between subfamilies. All group V members have 5 to 7 transmembrane helices, most group IV members have 8 to 10 transmembrane helices, and members of groups I, II, and III have 11 to 12 transmembrane helices (Fig. 5). The N-terminal ends of all BvSUTRs are located inside the cell, and the N-terminus is longer than the C-terminus. The positions of terminal ends are important for sugar and signal recognition as well as for regulating the rate of sugar transport.
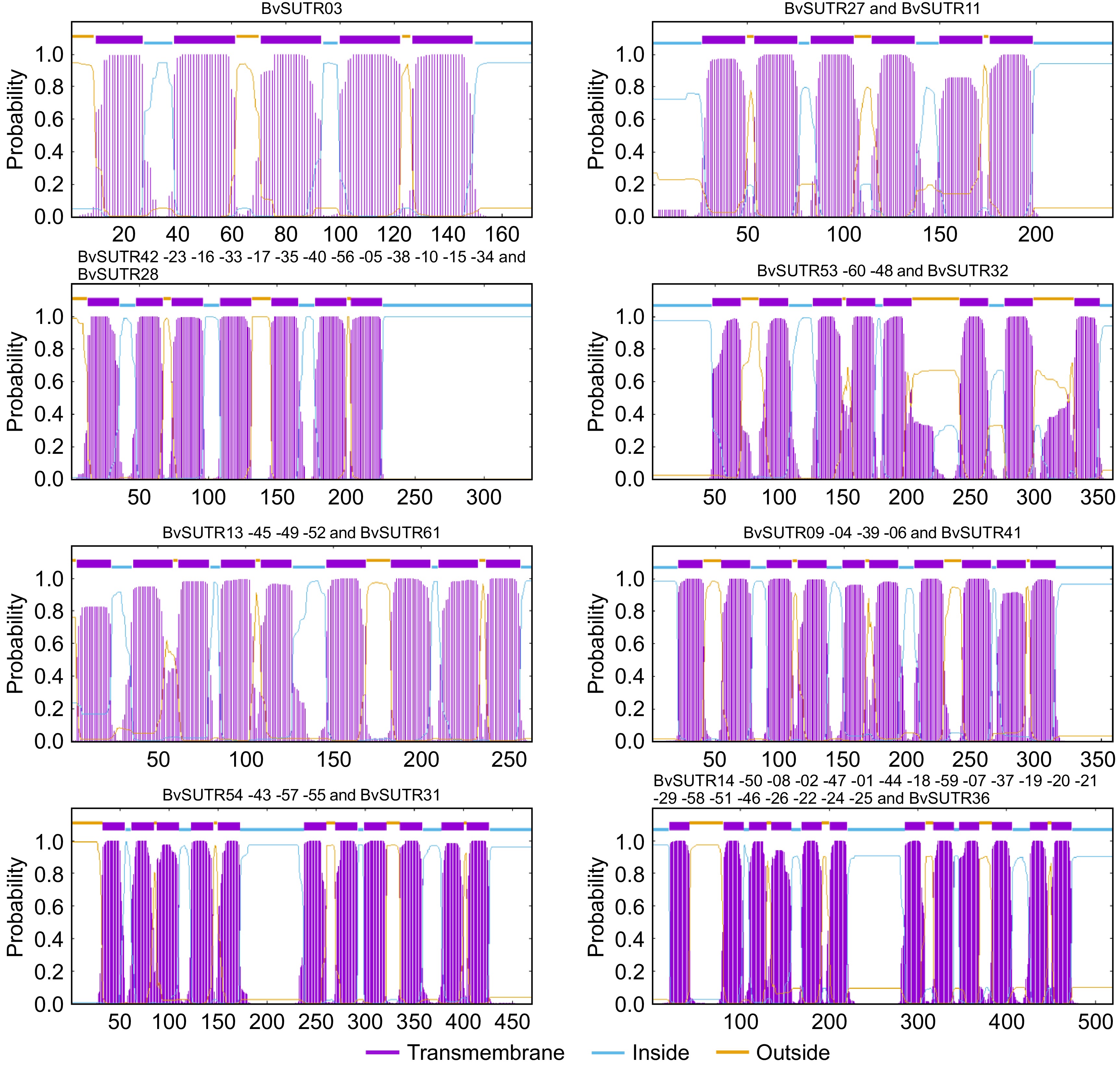
Figure 5.
Transmembrane helical structure of BvSUTRs.
Three-dimensional structure of BvSUTRs
-
Three-dimensional structures and binding sites were modeled for all BvSUTR proteins, and seven conserved structures based on grouping from phylogenetic analysis are illustrated in Fig. 6. The members of groups III and IV had similar structures, and two types of structures were observed among the members of group V, including SWEET sugar transporters. Determining the active and interacting regions of proteins is possible by studying the pocket regions. In the present study, the percentage of each amino acid present in the ligand-binding regions was calculated for the members of each group of the BvSUTR family, and the results indicated that BvSUTR proteins are different in terms of their ligand-binding regions (Fig. 7). Among the members of group I, leucine, phenylalanine, and glycine were more common in the ligand-binding site, and glycine, serine, and isoleucine were more common in the ligand-binding regions of the BvSUTRs from group II. In group III, leucine, valine, and glycine were frequently distributed in ligand-binding sites. Asparagine was more common in the ligand-binding sites of the members of group IVa than in those of group IVb, while leucine and serine were more common. In addition, serine amino acids were strongly repeated in the ligand-binding sites of proteins from group V. Overall, serine and leucine are more common in the pocket sites of BvSUTR family members.
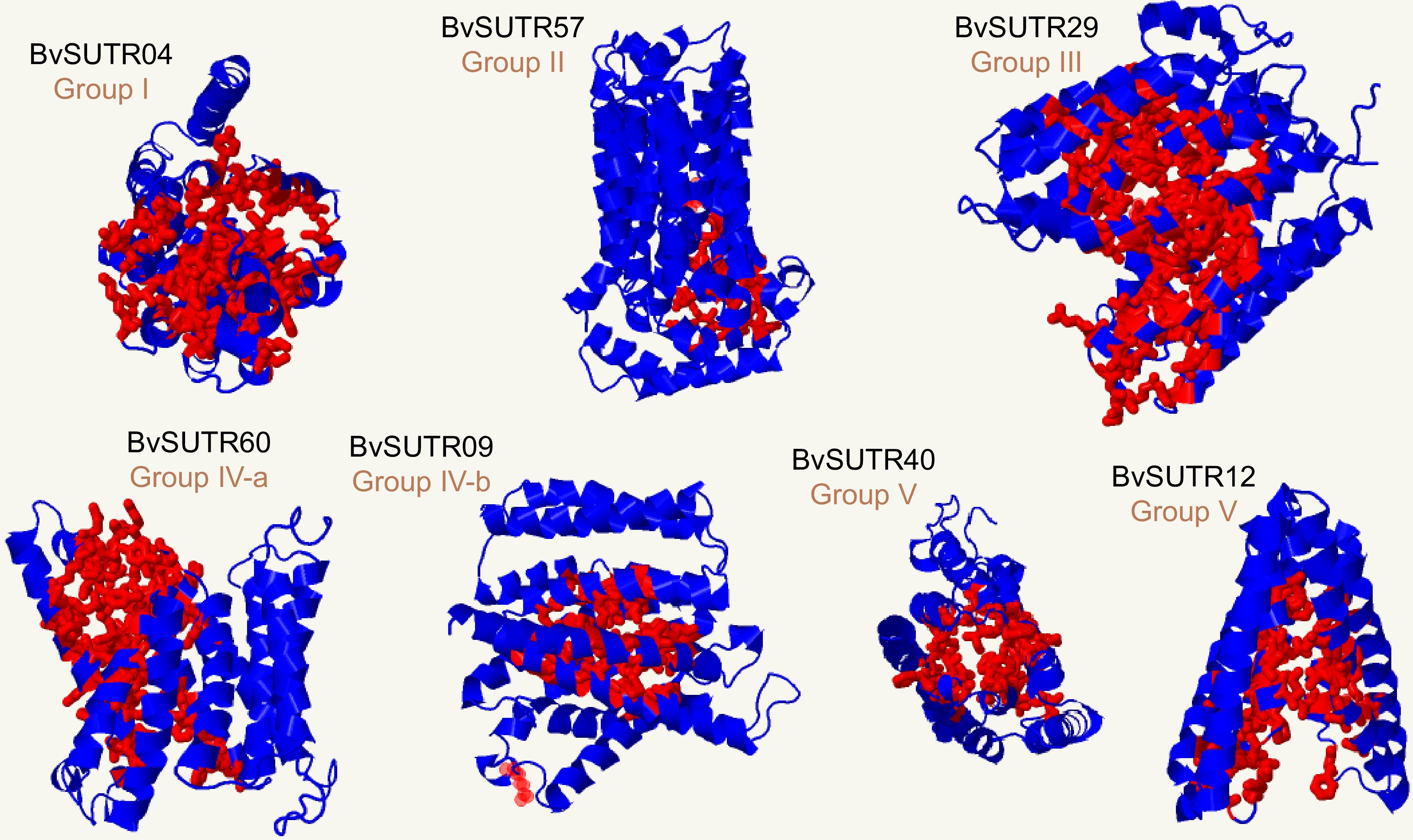
Figure 6.
3D structure and binding sites of BvSUTRs. The red indicates the binding site in the 3D structure of the candidate BvSUTR proteins.
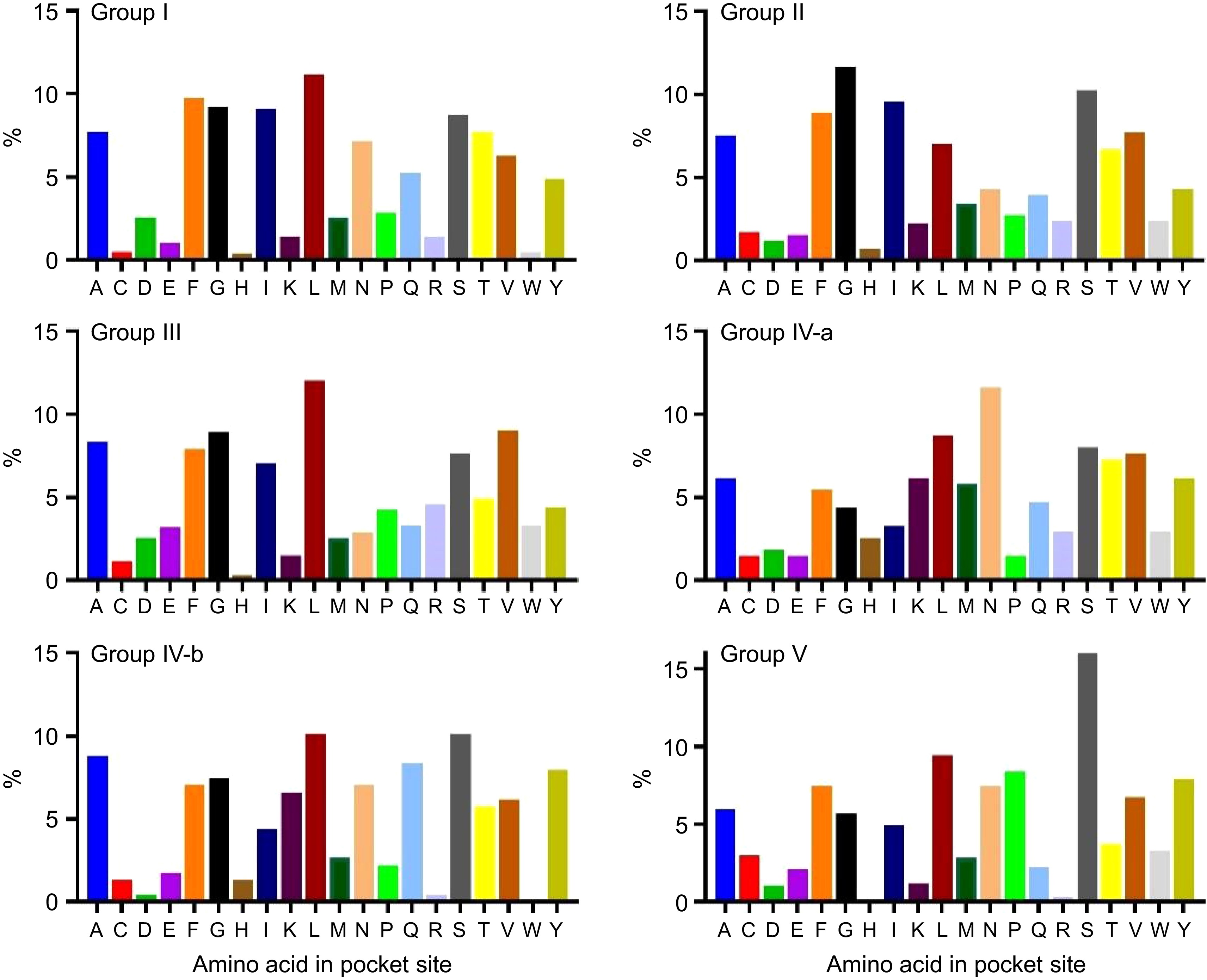
Figure 7.
Percentage of each amino acid in the pocket site of the BvSUTR groups based on phylogenetic analysis. Alanine (A); Cysteine (C); Aspartic acid (D); Glutamic acid (E); Phenylalanine (F); Glycine (G); Histidine (H); Isoleucine (I); Lysine (K); Leucine (L); Methionine (M); Asparagine (N); Proline (P); Glutamine (Q); Arginine (R); Serine (S); Threonine (T); Valine (V); Tryptophan (W); Tyrosine (Y).
Promoter analysis of SUTRs
-
The putative cis-regulatory elements present in the promoter region of the BvSUTR genes were classified into four groups: plant growth and light-responsive elements (REs), hormone REs, stress REs, and binding REs (Fig. 8a & Supplementary Table S2). Putative cis-regulatory elements, such as the CAT-box (related to meristem expression), Box 4 and G-box (light responsiveness), MYB, ABRE (abscisic acid responsiveness), and GARE-motif (MeJA responsiveness), were frequently recognized in the upstream region of the BvSUTR genes (Supplementary Table S2). Cis-regulatory elements related to growth and light (approximately 71.20%) and stress (approximately 17.23%) were more common in the promoter region of the BvSUTR genes, while cis-regulatory elements (approximately 8.91%) had roles in hormone responsiveness. The genes encoded abscisic acid (ABA), auxin, salicylic acid (SA), gibberellins (GA), and methyl jasmonate (MeJA) in the regulatory regions of the BvSUTRs, suggesting that the BvSUTR may play a key role in the response to stress conditions. The frequency of ABA and MeJA responsiveness was within the promoter regions of the BvSUTR genes (Fig. 8b).
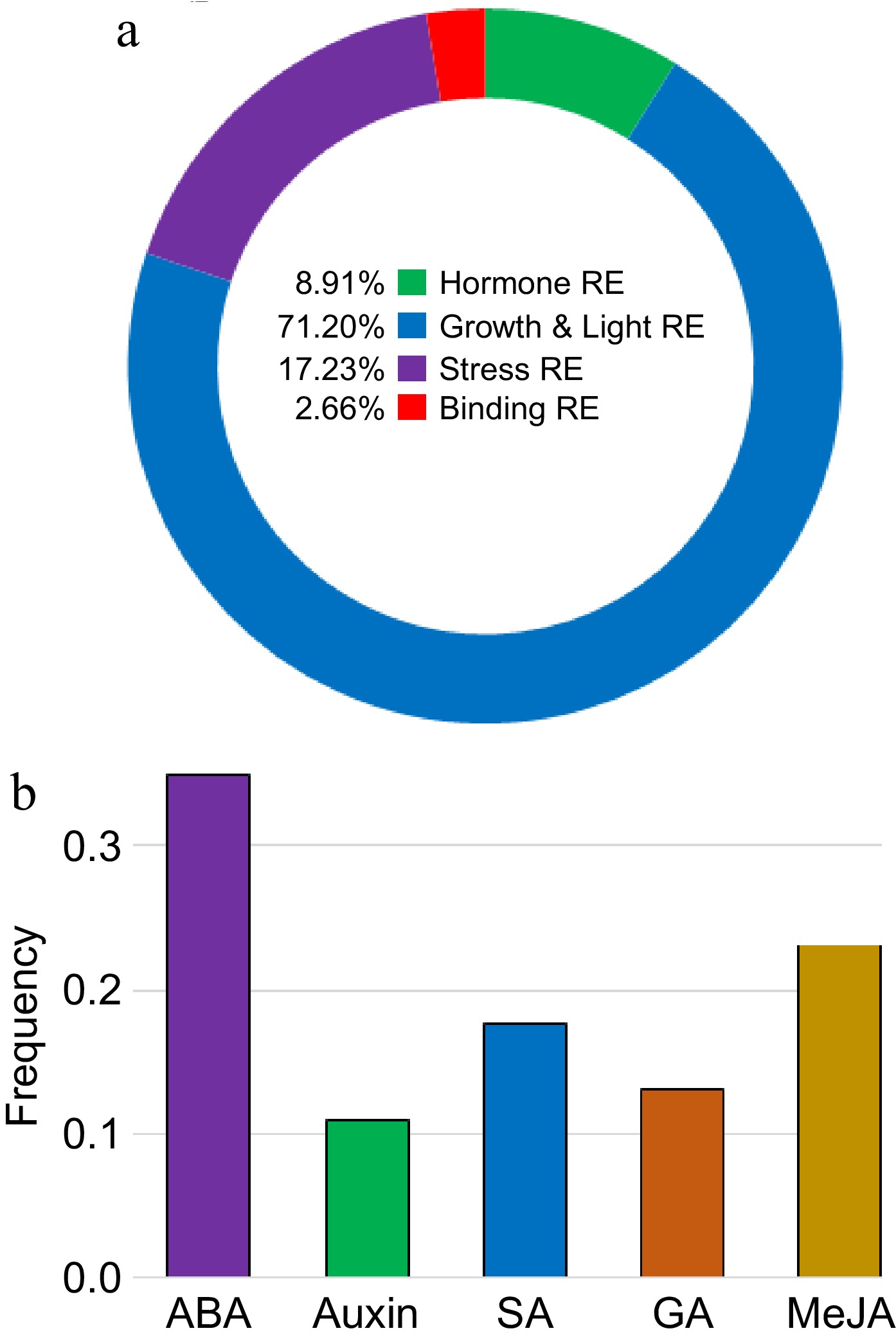
Figure 8.
Promoter analysis of BvSUTRs. Classification of cis-regulatory elements present in the promoter site of (a) BvSUTR genes and (b) involved in the response to hormones. The full details are presented in Supplementary Table S2.
Expression profile of SUTR genes under abiotic and biotic stimuli via RNA-seq data
-
RNA-seq experiments were also employed to further study the expression of the identified SUTRs in various tissues and under abiotic stresses. The expression levels of SUTRs were studied in shoot and root tissues under cold, salt, drought, wounding, and heat conditions (Fig. 9a). The expression profile of SUTRs demonstrated a high expression level of the BvSUTR40 gene under salt stress conditions; in contrast, the BvSUTR40 gene showed low expression in shoots at wounding and heat conditions. In addition, the BvSUTR03 and BvSUTR56 genes were upregulated in shoots under salt stress conditions, while the BvSUTR50 gene exhibited the lowest level of expression in shoots under salt conditions. The genes with high expression under salt conditions were categorized in group V for coping with salt stress, suggesting the potential of these genes for coping with unfavorable conditions. Furthermore, the BvSUTR23, BvSUTR05, BvSUTR40, and BvSUTR07 genes were upregulated in shoots under cold stress conditions (Fig. 9a). The expression pattern of SUTRs demonstrated a remarkable expression level in the BvSUTR58 gene in roots under salt stress conditions, and the BvSUTR58 gene was upregulated under cold and drought conditions. Additionally, the BvSUTR01 and BvSUTR35 genes were upregulated in roots under salt stress conditions, whereas the BvSUTR22 gene exhibited low expression in roots under salt stress conditions (Fig. 9a). The BvSUTR35 gene exhibited a moderate level of expression in root tissue under cold, salt, drought, and wounding stress conditions, while downregulated expression was observed in shoot tissue under abiotic stress conditions (Fig. 9a).
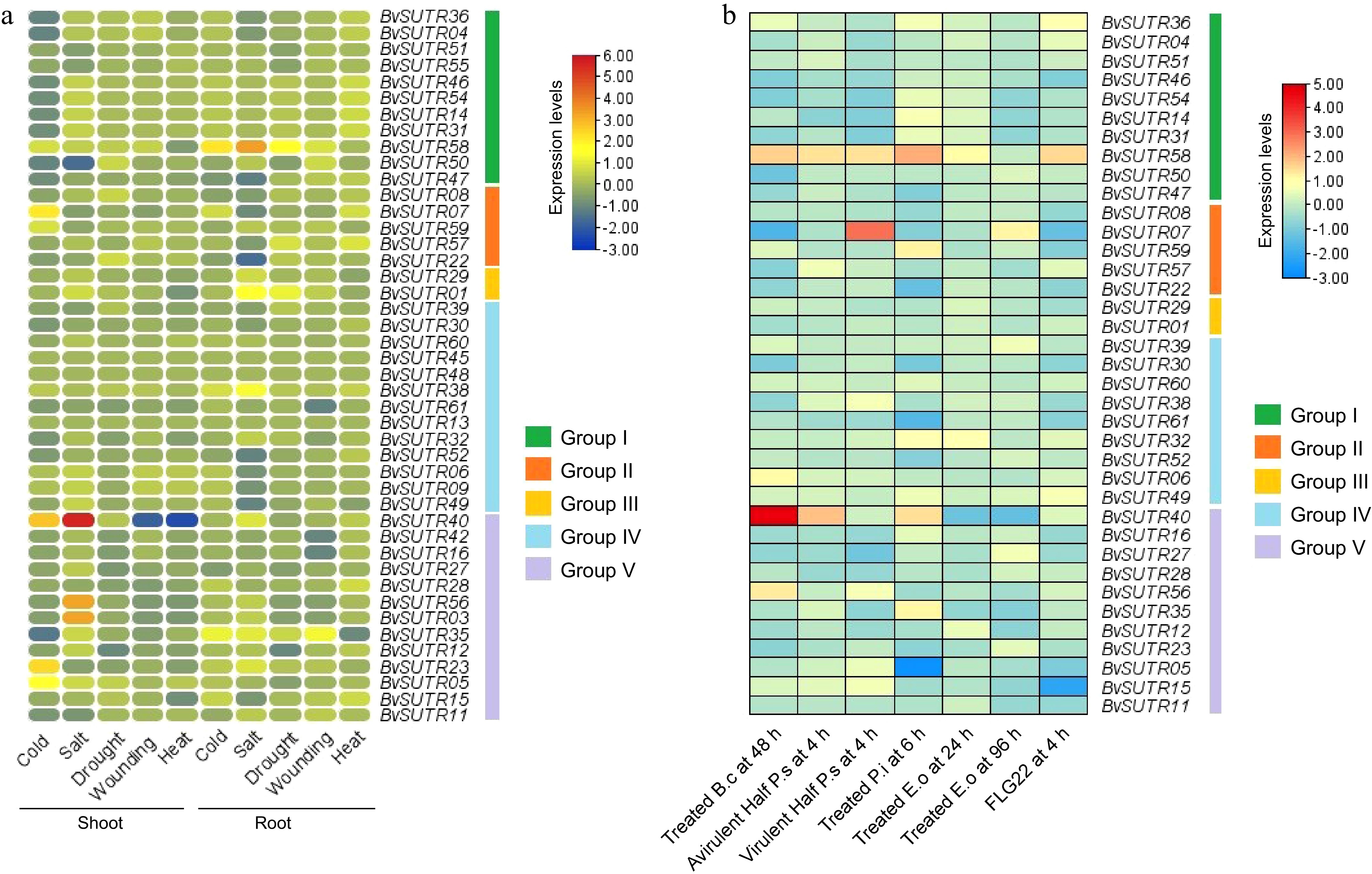
Figure 9.
Expression profile of SUTRs in response to abiotic stresses in (a) shoot and root tissues, and (b) expression profile of SUTRs in response to biotic stresses. B.c: Botrytis cinerea; P.s: Pseudomonas syringae; P.i: Phytophthora infestans; E.o: Erysiphe orontii; FLG22: flagellin 22.
The expression patterns of SUTR genes were investigated in Botrytis cinerea, Pseudomonas syringae, Phytophthora infestans, Erysiphe orontii, the avirulent half of the P.S. at 4 h, the virulent half of the P.S. at 4 h, and flagellin 22 using RNA-seq data sets from Beta vulgaris. According to the expression pattern, the BvSUTR40 gene was highly expressed under 48 h of Botrytis cinerea stress, and it was upregulated in Phytophthora infestans and the avirulent half P.S. after 4 h of stress. In addition, the strains belonged to group V according to the results of the phylogenetic analysis. Furthermore, the BvSUTR07 gene was highly expressed in the virulent half of the P.S. at 4 h. The BvSUTR58 gene was upregulated under all the studied conditions except for treatment with Erysiphe orontii at 96 h (Fig. 9b). The BvSUTR05 and BvSUTR15 genes exhibited low expression under Phytophthora infestans (at 6 h) and flagellin 22 (at 4 h) stress conditions (Fig. 9b). The expression levels of SUTR genes in Beta vulgaris revealed differential expression patterns under biotic stress conditions, and the highly expressed genes belonged to groups I, II, and V (Fig. 9b).
Interaction network of SUTRs
-
The interaction network of BvSUTRs was constructed based on available data from the dicot model Arabidopsis thaliana (Fig. 10). The most strong interactions were observed for the BvSUTR41, BvSUTR34, BvSUTR58, BvSUTR50, BvSUTR29, BvSUTR02, and SWEET11 genes, for which carbohydrate transmembrane transporter activity and disaccharide transmembrane transporter activity were predicted for BvSUTR34 and SWEET11. BvSUTR08 was coexpressed with MEX1 and DPE1, and 4-alpha-glucanotransferase activity and beta-butylalanine-alpha-glucanotransferase activity were predicted to be involved (Fig. 10). There were no interactions or coexpressed genes for BvSUTR61, BvSUTR25, BvSUTR09, BvSUTR32, BvSUTR39, BvSUTR52, or BvSUTR53 (Fig. 10). However, some of the coexpressed genes were not annotated for predicted molecular functions, such as F17A22.21, A0A1P8AXP5, A0A19LLD6, T5K18.220, MWD22.7, M3E9.90-2, MUF8.1, MBK23.26, MSN9.8 TRM28, and F4D1. The gene ontology (GO) enrichment analysis of network elements revealed the following molecular function terms: carbohydrate transmembrane transporter activity, secondary active transmembrane transporter activity, pyrimidine nucleotide sugar transporter activity, sucrose transmembrane transporter activity, disaccharide transmembrane transporter activity, salicylic acid transmembrane transporter activity, symporter activity, antiporter activity, 4-alpha-glucanotransferase activity, hexose transmembrane transporter activity, beta maltose 4-alpha-glucanotransferase activity, and ion transmembrane transporter activity. Overall, the results of the interaction network revealed that there are strong interactions between sugar transporters, which probably affect each other's functions.
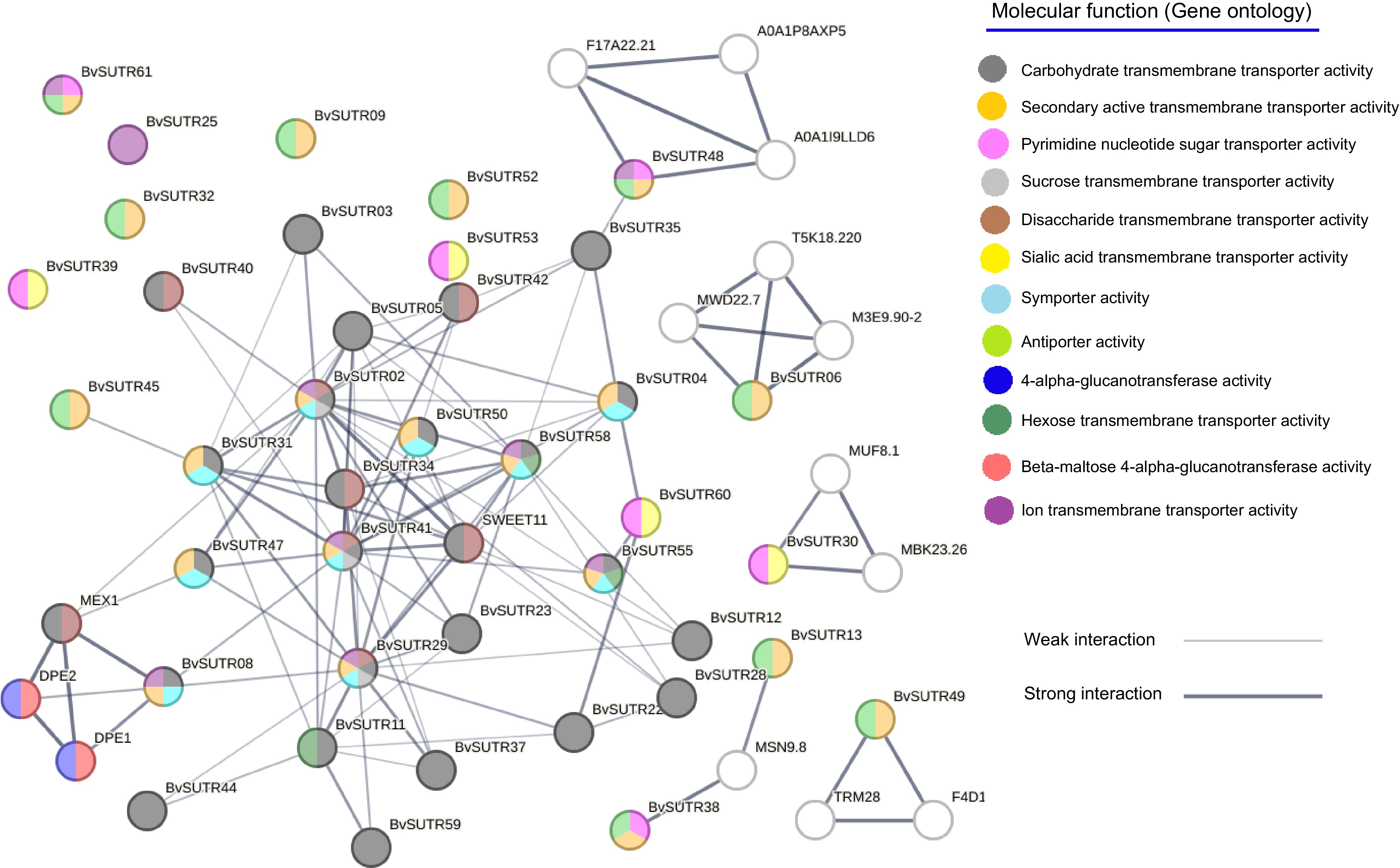
Figure 10.
Interaction network of SUTRs. The background color of each node represents the molecular function.
-
The carbohydrate D-glucose is the main source of energy in living organisms. Unlike animals, as well as most fungi, bacteria, and archaea, plants can synthesize additional sugars, allowing them to become autotrophic organisms. Glucose is stored as polymeric glucan, glycogen in animals, and starch in plants. Despite being a general source of metabolic energy and energy storage, glucose is the main building block for cellulose synthesis and represents the metabolic starting point for carboxylate and amino acid synthesis. Finally, glucose must act as a signaling molecule that conveys the metabolic state of the plant to regulate growth, development, and survival. Therefore, cell-to-cell transport and long-distance transport of sugars throughout the plant body require the precise activity of sugar transporters that facilitate transmembrane transport[2,71,72]. Sugar transporters are divided into subfamilies according to their function, each of which is responsible for transporting sugars to different tissues and organs. To date, there have been very limited studies on these subfamilies. In the present study, a total of 61 SUTR genes (including 16 SWEET genes) were identified from the genome of Beta vulgaris and named BvSUTR1−BvSUTR61. The number of members of this gene family varies among plants; for example, 29 SWEET members in tomato[73], 17 members in Arabidopsis[19], 52 members in soybean[35], 21 members in rice[38], 18 members in pear[74], 68 members in rape[75], 25 members in walnut[76], 27 members in apple[77], 59 members in wheat[78], 16 members in sweet orange[36], 25 members in Medicago truncatula[79], 19 members in Chinese jujube[80], and 25 members in banana[81]. The number of members of a gene family in plants is strongly affected by the polyploidy level and duplication events[82]. Phylogenetic analysis separated BvSUTRs into five groups based on their evolutionary relationships (Fig. 1). Sugar/inositol transporters were in groups I and II, sucrose/H+ symporters were in group III, both nucleotide-sugar transporters and sugar-phosphate transporters were in group IV, and sweet sugar transporters were in group V. The subfamilies were separated from each other based on the phylogenetic tree, and there was great similarity between the BvSUTRs and their orthologs in Arabidopsis. In addition, the number of exons/introns varied between members of each subfamily, revealing that evolutionary pressure has had different effects on each subfamily. Events such as incomplete duplication can increase the number of introns. On the other hand, the number of introns affects the speed of the process of expression and editing of primary mRNAs[83,84]. Additionally, based on the physicochemical characteristics, there was diversity among the members of each subfamily of BvSUTRs, and few duplication events were detected among them.
The BvSUTRs exhibited diverse transmembrane (TM) structures: the SWEET subfamily (group V) members had 5 to 7 transmembrane helices (TMHs), most group IV members (nucleotide-sugar transporters and sugar-phosphate transporters) had 8 to 10 TMHs, and members of groups I, II, and III had 11 to 12 TMHs (Fig. 4). Studies have shown that the expression of SWEET proteins is increased in TMHs under incomplete and tandem duplications, and the primary ancestor of this family had three TMHs[74,85]. Sugar transporters, a class of membrane proteins, are involved in many essential processes, including regulation, signaling and transport[86,87]. Determining the structure of the transmembrane structural domains of proteins and how they are located in the membrane can provide a better understanding of their functional structure. The amino acid sequence affects the formation of the specific shape of the transporter in the membrane, and as a result, the helical state allows dynamic processes during the transport process[88]. A low number of TMHs was detected in the SWEET, which could be related to their function. Unlike monosaccharide transporters (MSTs), SWEET transporters, pH-independent transporters, play a significant role in phloem transport and act as two-way transporters of sugars along a concentration gradient[6,20]. The number of TMHs in BvSUTRs differed from that in other reports, which could be due to variable evolutionary pressures. The three-dimensional structures of the BvSUTR proteins were diverse, indicating that each subfamily has a specific functional structure. However, serine residues were highly repeated in the pocket sites of the BvSUTR proteins. This hypothesis suggests that serine is a key element in the function and interaction of sugar transporters. Considering that serine is an important region in the phosphorylation process, transmitters may be involved in kinase-dependent signaling pathways.
The results of the analysis of the upstream region also confirmed that BvSUTRs are involved in the response to stress and stress-related hormones (ABA and MeJA) and that their functions are dependent on environmental conditions and external stimuli. The expression profile of SUTRs demonstrated that most genes are induced in response to environmental stress and that their expression level decreases. However, the expression levels of some single genes, such as BvSUTR58, increased in response to biotic stresses, and the BvSUTR40 gene was strongly upregulated in response to salinity stress. One of the mechanisms of plant response to environmental stress is the reduction in photosynthesis and vegetative activities, and the movement and transport of sugar can be related to the control of plant growth activities. A strong interaction between sugar transporter family members and several other transporters was estimated, which indicated that the transporters cooperate in a chain system and the performance of each of them is effective in determining the performance of other transporters. The reason for this difference could be that they all participate in maintaining the ionic balance of the cell.
-
In the present study, sugar transporter family members in Beta vulgaris were identified and characterized. Three main subfamilies, monosaccharide transporter, sucrose transporter, and SWEET sugar transporter, were separated and compared. The results revealed that the diversity and expansion of the members of this gene family occurred before the separation of monocots and dicots, and then, some partial duplications occurred within the SUTR genes, which caused an increase in the number of transmembrane domains of these transporters. Based on the protein structure and the study of binding regions, it was predicted that serine amino acids play a key role in the activity of sugar transporters. Of course, there are also high potential areas for phosphorylation. Sugar transporters seem to be influenced by kinase-dependent signaling pathways.
We thank the Shahrood University of Technology for financially supporting and providing the needed facilities.
-
The authors confirm contribution to the paper as follows: study conception and design: Heidari P; data collection: Heidari P, Hosseinzadeh M; analysis and interpretation of results: Heidari P, Ahmadizadeh M, Hosseinzadeh M; draft manuscript preparation: Heidari P, Ahmadizadeh M; manuscript revision: Heidari P, Ahmadizadeh M. All authors reviewed the results and approved the final version of the manuscript.
-
The data generated or analyzed in this study are included in this article. Other materials that support the findings of this study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 BvSUTRs properties.
- Supplementary Table S2 Promoter analysis.
- Supplementary Table S3 GO enrichment of interaction network.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Heidari P, Ahmadizadeh M, Hosseinzadeh M. 2025. Sugar transporter gene family: structure, evolution, and expression analysis in Beta vulgaris. Vegetable Research 5: e002 doi: 10.48130/vegres-0024-0036 

Sugar transporter gene family: structure, evolution, and expression analysis in Beta vulgaris
- Received: 14 May 2024
- Revised: 27 August 2024
- Accepted: 23 September 2024
- Published online: 10 January 2025
Abstract: Sugar transporters are key mediators of the distribution of photosynthesis products in plant organs and play a key role in maintaining the sugar balance in cells. In this study, for the first time, all sugar transporters in Beta vulgaris (BvSUTR) were identified and compared based on their structure and function. A total of 61 BvSUTR genes were identified, and they were classified into four subfamilies: sugar/inositol transporter, sucrose/H+ symporter, nucleotide-sugar transporter, and sweet sugar transporter. Gene structures and physiochemical characteristics varied significantly across the phylogenetic groups, with a lower prevalence of phosphorylation regions detected in individuals belonging to group I and group V, which are part of the sweet sugar transporter subfamily. Additionally, there was a variation in the number of transmembrane helices from 5 to 12, notably fewer in the sweet sugar transporter subfamily. Moreover, the BvSUTR proteins displayed distinct three-dimensional structures and binding sites, where serine and leucine residues were found to be more prevalent in the binding locations. Analysis of the upstream region revealed that cis-regulatory elements related to growth, light, and abscisic acid (ABA) responsiveness are repeated in the promoter region of BvSUTRs. The expression profile revealed that SUTRs are less expressed in response to abiotic stresses. Analysis of the interaction network revealed that there is a strong interaction between sugar transporters and members of this gene family, which further influences the function of their other paralogs. The results of this research can be used in molecular research related to sugar beet breeding programs.
-
Key words:
- Plant transporter gene family /
- Sugar /
- Interaction network /
- Gene structure /
- Genome wide