










-
Cold temperatures are a critical environmental constraint that affects the productivity and geographical distribution of plants. This condition can lead to reduced water availability, decrease plant membrane fluidity in plants and disrupt the balance between photosynthesis and respiration, thereby adversely affecting plant growth and survival[1]. Cold stress is typically defined as temperatures ranging from 0 to 15 °C[2]. Due to early spring's unstable temperatures in China, plants often experience sudden drops to chilly conditions[3], which can significantly impact their growth, productivity, and quality[1].
Plants have evolved various adaptation strategies to cope with cold stress by maintaining cellular homeostasis and mitigating stress-induced damage. These adjustments predominantly involve alterations in physiological processes[4], metabolism[5], and transcription[6]. The fluidity of the cell membrane is crucial for maintaining its functional activity, and it is highly sensitive to temperature fluctuations[7]. Studies have indicated that under cold conditions, damage to the cell membrane is predominantly due to lipid peroxidation and unsaturation of fatty acids[8]. Furthermore, increased levels of a lipid peroxidation product malondialdehyde (MDA) correlate with reduced cold resistance in plants[9]. Additionally, soluble sugars, which interact with membrane lipids, play a crucial role in stabilizing cell membranes[10]. During cold stress, the leakage of electrons can impair ATP synthesis, leading to metabolic disruptions and hindering the energy supply to the plant[11]. This impairment further compromises plant health and viability. Moreover, intricate regulatory networks involving metabolism and physiology are essential for enhancing plant cold tolerance[12]. Central to this mechanism is the activation of the antioxidant defense system, which involves the accumulation of nitrogenous compounds with low molecular weights, such as betaine and proline (Pro)[13], and soluble sugars[14]. Additionally, antioxidant enzymes, such as peroxidase (POD), dismutase (SOD), and superoxide catalase (CAT), are crucial for mitigating the buildup of reactive oxygen species (ROS) within plant cells[15]. Soluble sugars and nitrogenous compounds with low molecular weights, which are both cryoprotective agents, not only aid ROS removal but also help maintain cell membrane integrity, protein stability[13,14], and ion homeostasis[16]. Moreover, various critical metabolites, including phenylpropanes, carbohydrates, flavonoids, anthocyanins, and steroids, are instrumental in the plant's cold response by functioning as osmotic regulators and antioxidants[15]. Plant hormones also significantly affect plant cold response, with changes in the levels of jasmonic acid (JA), abscisic acid (ABA), ethylene (ET), and gibberellins (GA), each contributing to the regulation of cold acclimation[17] and enhancing cold tolerance.
In recent years, the rapid advancements in genome sequencing[18], transcriptomics[19], and metabolomics[20] have significantly enhanced our knowledge of the complex strategies employed by plants to withstand cold stress. RNA sequencing (RNA-seq) studies have documented variations in the transcriptomes of various plants, such as maize[21], Arabidopsis[22], and grape[18]. The MYB, WRKY, AP2/ERF, bZIP, NAC, and ZFP transcription factor (TF) families are specifically responsive to cold signals in these plants[23,24]. Moreover, certain proteins like the heat shock protein 70, cold-regulated 413 plasma membrane protein, dehydration-responsive proteins, and ethylene-responsive transcription factors, are crucial for adaption to cold conditions[25]. Beyond transcriptomics, metabolomics plays a vital role in analyzing plant metabolic responses under cold stress. This approach has detected various metabolites, including amino acids[26], carbohydrates[20], and abscisic acid (ABA)[27], which are significant in responding to cold stress. Metabolomics offers a detailed view of the regulatory mechanisms underlying energy transmission and cellar signal transduction through the identification of end products of gene expression and modification of proteins[20].
The integration of transcriptome and metabolomic data is crucial for unraveling the intricate regulatory networks that govern plant responses to cold stress. For instance, combined analyses in wheat have highlighted the importance of jasmonyl isoleucine and ABA in cold adaptation[28]. This integrated approach has also been applied extensively to investigate plant responses in apples and other species to cold stress[29]. Such research provides valuable insights, particularly into the cold responses in woody species like fruit trees. However, there is still a gap in our understanding of these responses in other woody plants, such as tea trees, where research remains limited.
Environmental stress conditions significantly impact plant growth, development, and yield. Research over the past three decades has highlighted silicon's 'quasi-essential ameliorative' role in combating various environmental stresses in plants[30]. This beneficial effect involves regulating morphology and growth, promoting osmolyte accumulation, improving photosynthesis, enhancing antioxidant enzyme activities, facilitating nutrient uptake, and modulating other metabolic pathways under diverse abiotic stresses like temperature fluctuations, drought, heavy metal toxicity, UV radiation, salinity, and nutrient deficiency[31]. Commonly used forms of silicon include ortho-silicic acid[32] and silicate salts such as sodium silicate, potassium silicate, and calcium silicate[33]. Under cold conditions, silicon has demonstrated advantageous effects across multiple plant species. For instance, it has been observed that maize can recover from cold exposure through the application of silicon, which alleviates oxidative stress and helps balance the hormonal levels[34]. Additionally, silicon has been shown to enhance the accumulation of biomass in maize by increasing the levels of proteins, protective pigments, glutathione, and ascorbic acid, and by improving photochemical reactions[35]. In barley, silicon treatment has led to increased levels of soluble carbohydrates and proteins, as well as enhanced antioxidative enzyme activities in leaf apoplasm when exposed to cold stress[36].
Rice recovery from cold stress is facilitated by the up-regulation of cellular metabolisms and over-expression of the Lsi1 gene[37]. The resilience of wheat under cold conditions depends on enhanced antioxidant defense activity, increased membrane permeability, and improved water retention in leaf tissues[38]. Notably, the plants mentioned in these reports are primarily members of the Gramineae family. In the realm of horticultural crops, recent studies have shown that bamboo[39] and aloe vera[40] recover from cold stress by increasing SOD, POD, and CAT activities, reducing MDA levels and membrane damage, and boosting concentrations of amino acids and soluble sugars. Despite these findings, extensive transcriptomic and metabolomic studies are still needed to fully elucidate the mechanisms through which silicon alleviates cold stress. It is essential to gain a deeper understanding of regulatory elements, transcription factors, and silicon-related transporters, especially in how they interact with hormonal signaling, antioxidant responses, cold stress mitigation, and other key physiological pathways. Silicon amelioration has evolved into a prominent foliar application strategy for enhancing plant productivity and stress resilience; however, further investigations are necessary to uncover its mechanisms and regulate key metabolic and defense pathways.
In this study, the tea plant cultivar 'Fuding Dabaicha' from the genus Camellia in the Theaceae family, known for its sensitivity to cold stress[41], was treated with potassium silicate to examine its phenotypic, physiological, and biochemical responses under short-, medium-, and long-term cold treatments. Transcriptome sequencing and metabolomics technology were employed to identify key candidate genes and essential regulatory pathways, providing a theoretical basis for silicon-mediated amelioration of cold tolerance in tea plants.
-
The study involved one-year-old seedlings of the Camellia sinensis 'Fuding Dabaicha'[42,43], sourced from Nanjing Yarun Tea Industry Co., Ltd., China, located at 36°28' N, 116°82' E. These seedlings were potted in 14 cm2 by 18 cm deep plastic containers using a soil mixture composed of peat, sand, and perlite in a 2:1:1 ratio. The growth environment was controlled within a cultivation chamber at Shandong Agricultural University, China, maintaining 80% relative humidity, a 16 h/8 h light/dark photoperiod, and a 25 °C/18 °C day/night temperature cycle.
Seedlings of a uniform height (approximately 15 cm) were subjected to a 4 °C cold treatment for durations of 1, 3, and 5 d. The treatments included LTWL (seedlings treated with 1.80 mmol/L sodium sulfate at 4 °C for 1, 3, and 5 d) and LTSiL (seedlings treated with 1.80 mmol/L sodium silicate at 4 °C for the same durations). Ten seedlings were selected for each treatment, with three biological replicates maintained. At the end of each treatment duration (1, 3, and 5 d), the leaves were collected — excluding the winged leaves, petioles, and midribs — promptly flash-frozen in liquid nitrogen and preserved at −80 °C until use. Each leaf sample was bifurcated for different analyses; one portion was utilized to assess physiological indices like lipid damage, antioxidant enzyme activity, and osmotic regulatory substance content, while the remaining samples were designated for metabolomic analysis and transcriptomic sequencing.
Physiological measurements
-
To assess lipid damage, the degree of lipid peroxidation was determined using the thiobarbituric acid reactive substances (TBARS) assay by quantifying malondialdehyde (MDA) levels. Absorbance readings were taken at 450, 532, and 600 nm, and the formula: C = 6.45 × (A532 − A600) − 0.56 × A450 was used to calculate MDA concentration (μmol/L)[44]. Additionally, 0.5 g of each leaf from each treatment was mixed with 20 mL of deionized water in a 50 mL centrifuge tube and agitated at room temperature for 10 h at a speed of 150 r/min. A conductivity meter (DDS-307, Rex, China) was used to measure initial electrical conductivity (EC1). Samples were then boiled at 100 °C for 30 min, followed by another agitation for 10 h at room temperature, after which the final electrical conductivity (EC2) was recorded. The equation REL (%) = (EC1/EC2) × 100 was used to calculate the relative electrolyte leakage (REL)[45]. All measurements were replicated three times.
CAT, SOD, and POD activities were measured using specific assay kits (Nanjing Jiancheng, China), following methods referenced in previous studies[46]. These measurements were performed in triplicate. The anthrone method was used to determine soluble sugar content by measuring absorbance at 620 nm to investigate osmotic regulatory substances[47], while proline content was determined by its reaction with acidic ninhydrin to form a red product, measured at 520 nm[48]. These measurements were also performed in triplicate. Ultraviolet spectrophotometry (MAPADA, UV-1200) was used to determine the activities of SOD, POD, MDA, and CAT, as well as the contents of soluble sugars and proline.
Liquid chromatography-mass spectrometry (LC-QQQ-MS, Agilent, USA) was used for the determination of jasmonic acid (JA) contents. Frozen leaves of approximately 100 mg were ground to a fine powder in liquid nitrogen and mixed with a standard (Methyl jasmonate, 98%, Sigma). The mixture was centrifuged at 4 °C, after which the clear supernatant was carefully decanted into a 2 mL test tube. The remaining residue underwent a second extraction using 0.4 mL of 70% methanol (v/v). The combined supernatants were then dried using a vacuum concentrator (Eppendorf, Germany). Subsequent analysis was conducted with a flow rate of 0.4 mL/min and a column temperature of 40 °C[49].
To determine free amino acid contents, an amino acid analyzer was employed[50]. The extraction procedure involved using 20 mg of the sample, which was treated using 1 mL of 4% sulfosalicylic acid. The mixture was then subjected to ultrasonic extraction for 30 min, followed by centrifugation for another 30 min. The sediment was then re-extracted using the same method. Up to 2 mL of the resulting supernatant was collected, and 1 mL of this supernatant was then filtered for subsequent analysis. For detection, 20 μL of both standards and the sample were introduced into a high-speed amino acid analyzer (L-8900, Hitachi, Japan). Analytical conditions were set as follows: a 0.35 mL/min mobile phase flow rate, a 0.3 mL/min derivatization reagent flow rate, a post-column reaction temperature of 130 °C, and a column temperature of 38 °C. The autosampler temperature was maintained at 4 °C. The peak area of each compound was compared against the corresponding amino acid standard. These standards included L (+)-Glutamic acid, glycine, L-Histidine, L-Threonine, L-Phenylalanine, L-Valine, L-Tyrosine, and L-Tryptophan, with purities ranging from 98.5% to 99.5% (Sigma).
Catechin content was quantified using high-performance liquid chromatography (HPLC)[51]. A uniformly ground sample (0.2 g) was treated with 5 mL of 70% methanol aqueous solution that had been preheated to 70 °C and immersed in a water bath set at 70 °C for 10 min. The solution was centrifuged once cooled to ambient temperature, and the supernatant was decanted into a 10 mL volumetric flask. The remaining residue underwent re-extraction following the same procedure using another 5 mL of the aforementioned 70% methanol aqueous solution. These extracts were then combined, diluted to a total volume of 10 mL, and filtered to obtain the mother liquid. Subsequently, 2 mL of this mother liquid was taken and added to a 10 mL volumetric flask, then mixed with a stabilization solution consisting of 25 mL of 10 mg/mL EDTA-2Na solution, 25 mL of 10 mg/mL ascorbic acid solution, 50 mL of acetonitrile, and water to fill up to 500 mL. The solution was then brought up to volume, thoroughly mixed, and filtered back into the mother liquor. For HPLC analysis, 10 μL each of the standard working solution and the assay solution were injected into an HPLC system (U3000, Thermo, USA). Chromatographic conditions used were: detection wavelength: 280 nm, column: Cosmosil 5C18-AR-II (250 mm × 4.6 mm, 5 μm), injection volume: 20 μL, column temperature: 30 °C, mobile phase: phase A: pure water, phase B: 50 mmol/L phosphoric acid, phase C: acetonitrile, and flow rate: 0.8 mL/min. The standards used included EGCG (98% purity, Sigma), EGC ((−)-EGC, 98%, Sigma), ECG ((−)-Epicatechin gallate, 95%, Sigma), C ((−)-Catechin, 97%, Sigma), EC ((−)-Epicatechin, 95%, Sigma), and gallic acid (99%, Sigma).
RNA isolation and transcriptomic analysis
-
Total RNA was extracted from a composite sample of frozen leaves weighing 500 mg, derived from ten individual seedlings (one from each pot), utilizing the Universal Plant Total RNA Extraction Kit (Spin-column) by Bioteke Corporation, Beijing, China. This process was replicated three times for each treatment to ensure biological reproducibility. The RNA sequencing (RNA-Seq) analysis was performed utilizing the Illumina HiSeq platform (Illumina Inc., San Diego, CA, USA)[52] at Qingdao oeBiotech Co., Ltd. The high-quality clean reads obtained from sequencing were aligned directly to the Camellia sinensis genome, which was accessed from the 'Shuchazao' genome website (
www.ncbi.nlm.nih.gov/datasets/genome/GCF_004153795.1/ ) using the HISAT2 software. The criteria for DEG identification were set as follows: a P-value of less than 0.05, an absolute log2 (Fold Change) of at least 1, and a false discovery rate (FDR) below 0.05.Gene functions were comprehensively annotated based on databases such as the Kyoto Encyclopedia of Genes and Genomes (KEGG), the Protein Family Database (Pfam), Gene Ontology (GO), the National Centre for Biotechnology Information (NCBI) non-redundant protein sequences (NR), Swiss-Prot, TrEMBL, eukaryotic Orthologous Groups (KOG). This extensive annotation helped in understanding the biological implications of the observed gene expression changes.
Gene and accession numbers
-
The sequence data related to this study are available in the tea plant genome database:
https://doi.org/10.1073/pnas.1719622115 . The specific accession numbers assigned to the genes studied are as follows:CSS0014932: NAC; CSS0021931: ERF; CSS0012308: MYB; CSS0029008: WRKY; CSS0020832: C3H; CSS0050119: HSF; CSS0006857: AOC; CSS0012232: AOS1; CSS0028417: COI1; CSS0049044: MMK1; CSS0007714: CHS1; CSS0034616: CYP81Q32; CSS0035512: PLDZETA1; CSS0038386: DGK5; CSS0045289: LAC17.
Quantitative real-time PCR
-
Fifteen DEGs were randomly selected for qRT-PCR confirmation, with three biological replicates, respectively. The sequences of the Forward and Reverse primers designed using Primir version 5.0 (Premier Biosoft International, CA, USA) are listed in Supplementary Table S1. qRT-PCR was carried out according to Zhou et al.[53], and GAPDH was selected as a reference gene[54] listed in Supplementary Table S1.
Metabolome analysis
-
Widely targeted metabolomics analysis was conducted at Qingdao oeBiotech Co., Ltd. (www.qdoebiotech.cn), following the protocols detailed in Huang et al.[55]. Metabolites with an absolute log2(fold change) value greater than 1 and a variable importance in projection (VIP) score greater than 1 are classified as differentially abundant metabolites (DAMs), according to the orthogonal projections to latent structures-discriminant analysis (OPLS-DA). The oeBiotech metabolite database and KEGG database[56] (
www.kegg.jp/kegg/pathway.html ) were used for DAM function annotation. Pairwise Pearson correlation coefficients (PCCs) between DEGs and DAMs were calculated using the 'corrplot' function in the R package. Interactive networks were constructed using correlations with a p-value less than 0.05 and a PCC exceeding |0.8|, and visualized using Cytoscape 3.8.2[57]. Commonly affected pathways were identified by mapping the DAMs and DEGs to the KEGG database.Statistical analyses
-
One-way analysis of variance (ANOVA) was used to assess statistical significance using SPSS Statistics 26.0 (IBM Corp.), with p < 0.05. For a graphical representation of the data, Adobe Photoshop 2020, Prism 8.0.2, and Microsoft Excel 2010 were used for generating charts. The oeBiotech cloud platform (
https://cloud.obBiotech.cn ) was used for correlation analysis. These tools collectively facilitated comprehensive data analysis and visualization, aiding in the interpretation of complex statistical results. -
Tea plants treated with sodium sulfate and subjected to 4 °C exhibited symptoms of chlorosis and wilting within three days. Many of these plants were close to death after the cold treatment was extended to five days (Fig. 1a & b). However, a significant increase in chilling tolerance was observed when tea plants were treated daily with sodium silicate after initially being exposed to 4 °C (Fig. 1). This study demonstrates that silicon can significantly enhance the ability of tea plants to acclimate to cold stress.

Figure 1.
Effects of silicon-amelioration on tea plant in 1, 3, and 5 d under 4 °C. (a) Effects on whole plant growth. (b) Effects on leaf development. LTSiL: the leaves with silicon-amelioration under 4 °C; LTWL: the leaves without silicon-amelioration under 4 °C.
Under cold stress conditions, plant cells initiate various physiological adaptations to sustain their normal functions by synthesizing diverse biomolecules. In the present study, tea plant seedlings were treated with either sodium sulfate or sodium silicate and subjected to a temperature of 4 °C. Observations were recorded at intervals of 1, 3, and 5 d. Changes in REL, MDA, SOD, CAT, POD, Pro, soluble sugars, and JA content were measured across different time points. All physiological indices showed either increasing or decreasing trends as the exposure duration extended (Fig. 2a−h).
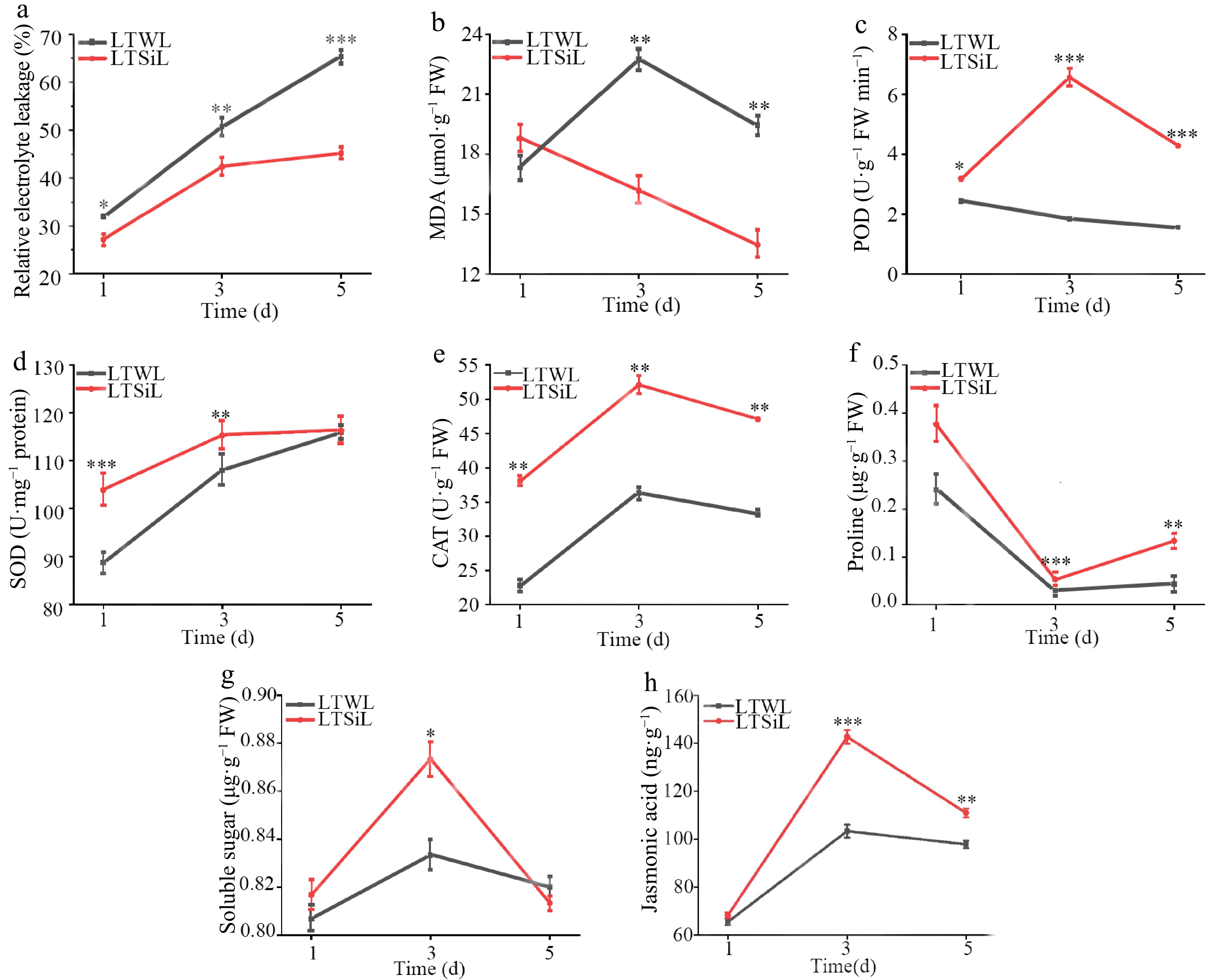
Figure 2.
Physiological changes in the leaves of tea plants with / without silicon-amelioration under 4 °C. (a) Relative electrolyte leakage (REL) values. (b) Malondialdehyde (MDA) content. (c) Peroxidase (POD) activity. (d) Superoxide dismutase (SOD) activity. (e) Catalase (CAT) activity. (f) Proline content. (g) Soluble sugar content. (h) JA content. LTSiL: the leaves with silicon-amelioration under 4 °C; LTWL: the leaves without silicon-amelioration under 4 °C. X-axis of (a)−(g) represents cold stress time. Y-axis of (a)−(g) represents substance content. The error bars represent means ± SD from three replicates.
While the SOD levels showed no noticeable differences between the two treatments during cold stress, significant differences were observed in the levels or activities of soluble sugars, REL, MDA, CAT, POD, and Pro in the seedlings treated with sodium silicate, especially after 3 d of exposure. Given these findings, leaves that were exposed to 4 °C for 3 d were chosen for detailed transcriptome sequencing and metabolomic analysis. Additionally, the changes in the contents of Val, Thr, Tyr, Trp, Phe, and Gly in the seedlings over time under cold stress were analyzed, noting a general decline in these components throughout the cold stress (Fig. 3a−f). In Fig. 4, changes in contents of Epigallocatechin Gallate (EGCG), Epicatechin Gallate (EGC), and Catechin (C) in tested seedlings at different times under cold stress were measured, all of which presented a downward trend, however, the contents of EGCG and EGC with silicon amelioration were more than those without silicon-amelioration.
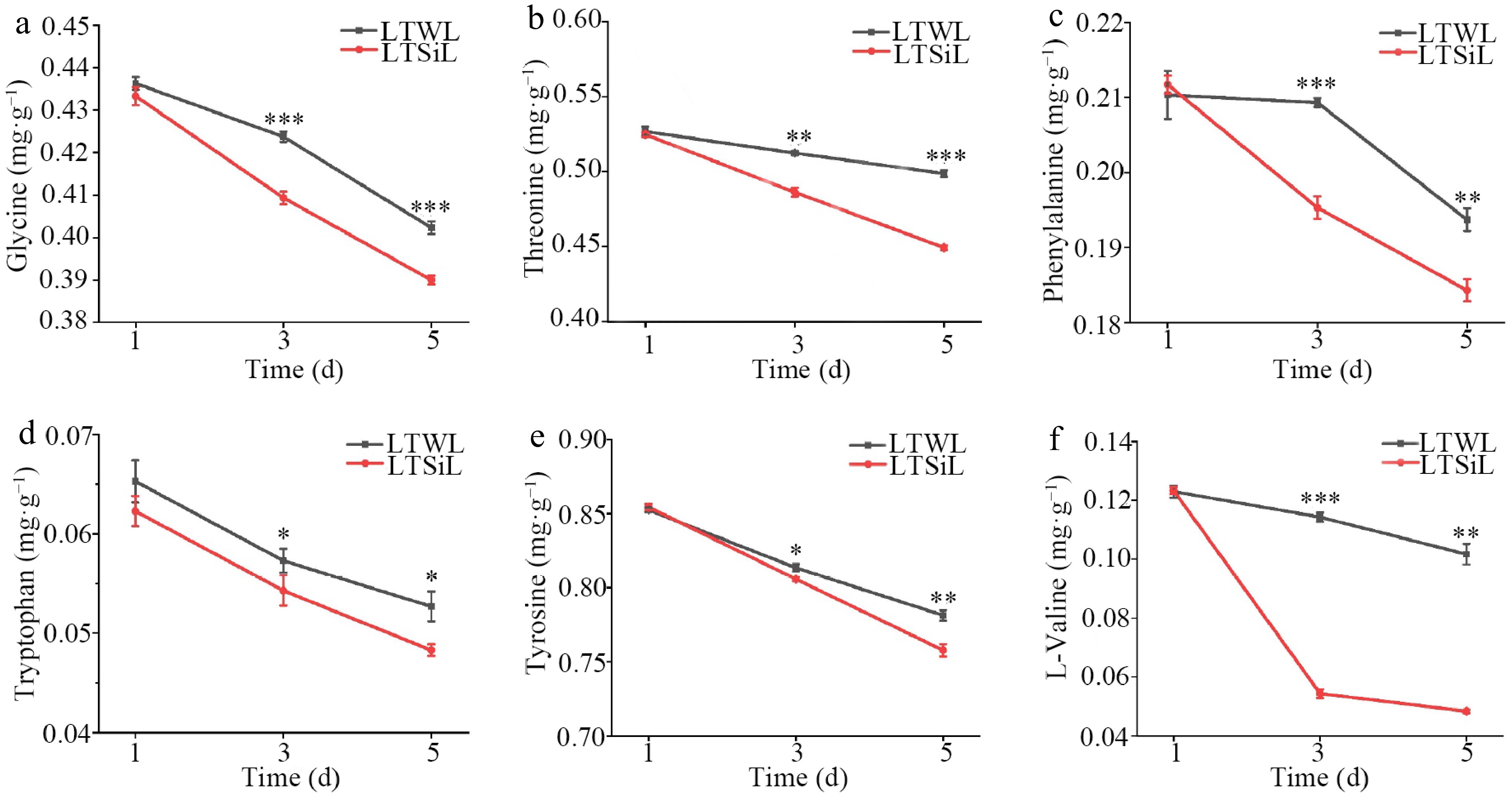
Figure 3.
Amino acids associated with physiological changes in the leaves of tea plants with / without silicon-amelioration under 4 °C. (a) Glycine (Gly) content. (b) Threonine (Thr) content. (c) Phenylalanine (Phe) content. (d) Tryptophan (Trp) content. (e) Tyrosine (Tyr) content. (f) Valine (Val) content. LTSiL: the leaves with silicon-amelioration under 4 °C; LTWL: the leaves without silicon-amelioration under 4 °C. X-axis of (a)−(h) represents cold stress time. Y-axis of (a)-(g) represents substance content. The error bars represent means ± SD from three replicates.
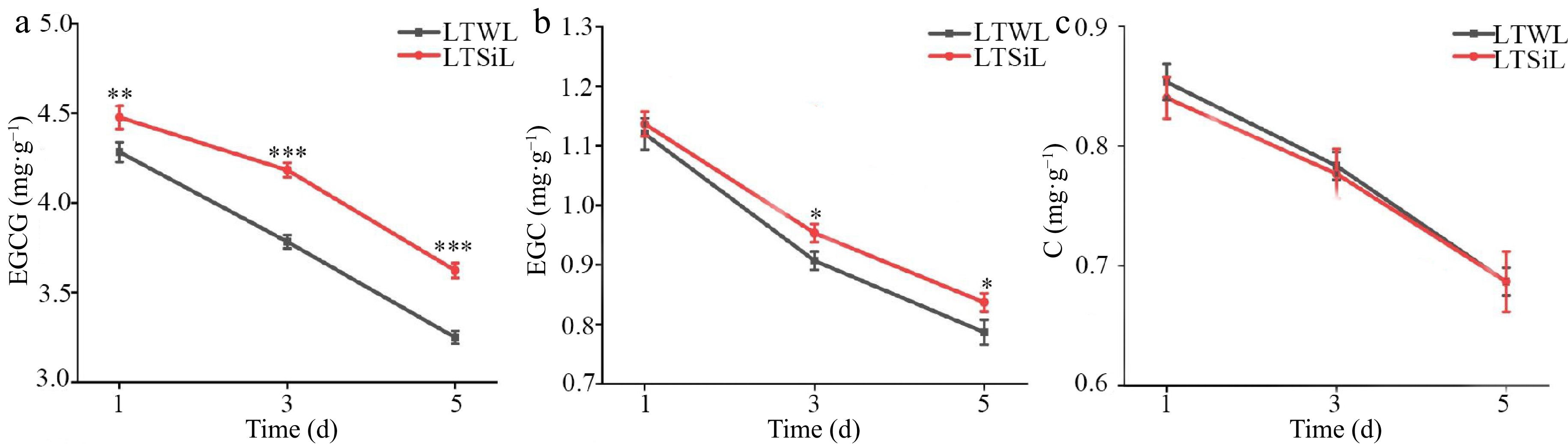
Figure 4.
Catechin associated with physiological changes in the leaves of tea plants with / without silicon-amelioration under 4 °C. (a) Epigallocatechin Gallate (EGCG) content. (b) Epicatechin Gallate (EGC) content. (c) Catechin (C) content. LTSiL: the leaves with silicon-amelioration under 4 °C; LTWL: the leaves without silicon-amelioration under 4 °C. X-axis of (a)−(f) represents cold stress time. Y-axis of (a)-(g) represents substance content. The error bars represent means ± SD from three replicates.
Enrichment analysis of DEGs in tea plants with Silicon-Amelioration (LTSiL) vs without (LTWL) under cold stress
-
Out of 871 DEGs identified between LTSiL and LTWL, 360 (41.31%) were up-regulated and 412 (58.69%) were down-regulated (Supplementary Fig. S1a). These DEGs underwent GO analysis for functional annotation and enrichment analysis, classified into three main GO categories: cellular component (CC), biological process (BP), and molecular function (MF). Specifically, three BP terms 'lignin catabolic process,' 'plant-type secondary cell wall biogenesis,' and 'phenylpropanoid metabolic process' were notably enriched in LTSiL compared to LTWL (Supplementary Fig. S1b). Additionally, enrichment in three CC terms including 'extracellular region,' 'apoplast', and 'cell wall' was observed (Supplementary Fig. S1b). Furthermore, five MF terms including 'hydroquinone: oxygen oxidoreductase activity', 'oxidoreductase activity', 'oxidizing metal ions,' 'copper ion binding', 'heme binding', and 'peroxidase activity' were also highlighted in the LTSiL group (Supplementary Fig. S1b).
KEGG pathway analyses revealed distinct metabolic responses in tea plants under cold stress. In the LTSiL group (silicon-ameliorated), pathways such as the 'mutual conversion between pentose and glucuronic acid', 'fructose and mannose metabolism', 'α-linolenic acid metabolism', and 'phenylpropane biosynthesis' were notably enriched, reflecting enhanced responses in carbohydrate metabolism, lipid metabolism, and biosynthesis of secondary metabolites in 'Fuding Dabaicha' (Supplementary Fig. S1c). Conversely, in the down-regulated DEGs of LTSiL compared to LTWL, pathways including 'Tyrosine metabolism', 'alanine, aspartate and glutamate metabolism', 'biosynthesis of various plant secondary metabolites', and 'photosynthesis' were prevalent, indicating a regulatory effect of these pathways on amino acid metabolism, secondary metabolite biosynthesis, and energy metabolism in response to cold stress (Supplementary Fig. S1d). These findings suggest that the enhanced cold resistance in LTSiL may be attributed to these metabolic adjustments.
Analysis of cold-responsive transcription factors in LTSiL vs LTWL tea plants under cold stress
-
Transcription factors (TFs) are crucial for regulating plant physiological development and responses to biotic and abiotic stresses in challenging environments. In the context of cold stress, a comparison between LTSiL (silicon-ameliorated) and LTWL (control) revealed significant differences in TF expression: 171 TFs were differentially expressed, with 72 up-regulated and 99 down-regulated in LTSiL. Figure 5b shows the top 13 TF families that were most responsive to cold stress in LTSiL compared to LTWL. Notably, six primary TF families—NAC, ERF, MYB, bZIP, MIKC_MADS, and WRKY—comprised 66.08% of the total TFs identified, highlighting their significant roles in mediating the cold stress response in tea plants.
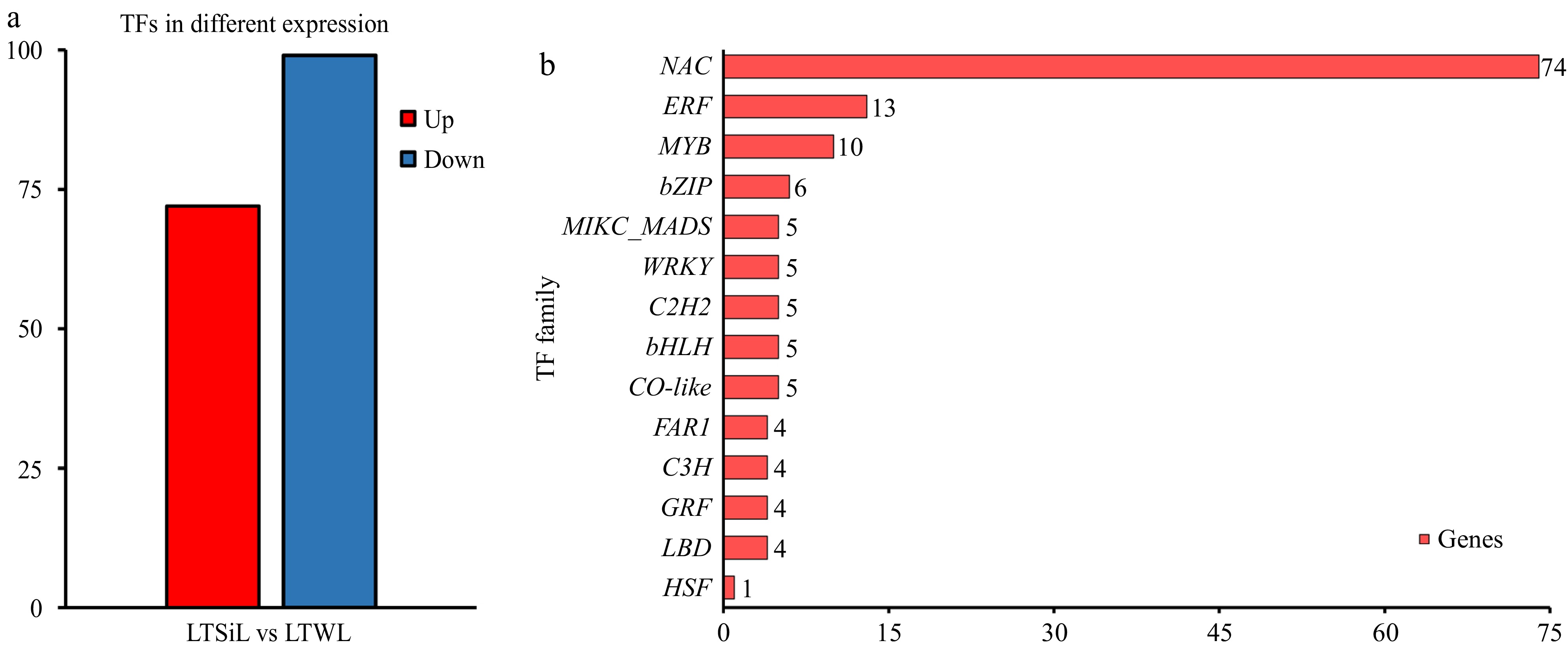
Figure 5.
Overview of the transcription factors differentially expressed in tea plants under 4 °C treatment. (a) The number of TFs identified in the pairwise comparison of LTSiL vs LTWL in tea plants. (b) The top 15 TF families for LTSiL vs LTWL that responded strongly to cold treatment. X-axis represents gene number and Y-axis represents the top 15 transcription factor families.
qRT-PCR analysis of DEGs in LTSiL vs LTWL tea plants under cold stress
-
To validate the transcriptome data, qRT-PCR analysis was conducted on 15 randomly selected candidate DEGs that are involved in various biological pathways. These pathways include 'lipid metabolism', 'isoflavone biosynthesis', 'phenylpropanoid biosynthesis', 'plant hormone signal transduction', 'amino acid synthesis', and 'flavonoid biosynthesis'. Additionally, the selection included genes encoding membrane proteins and TFs, representing both up-regulated and down-regulated DEGs between LTSiL and LTWL leaves. The expression profiles from the qRT-PCR analysis were consistent with those obtained from RNA-seq (Fig. 6a). Furthermore, the qRT-PCR results displayed a strong correlation with the RNA-seq data, achieving a Pearson correlation coefficient (r) of 0.9491, which substantiates the reliability of the RNA-seq findings (Fig. 6b).
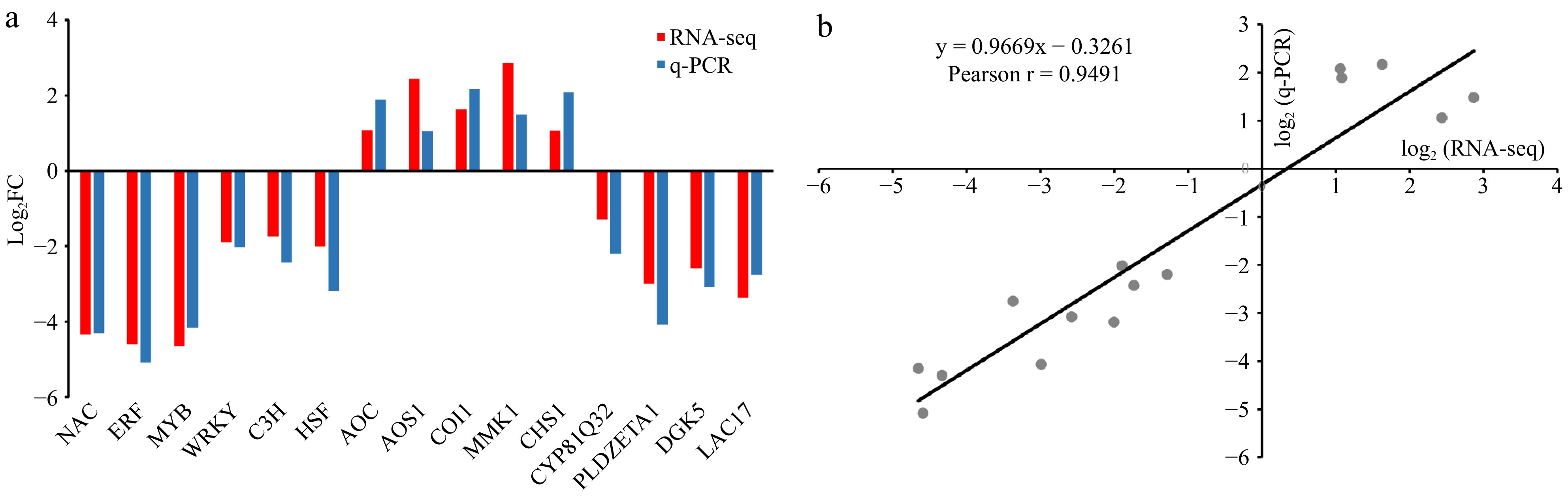
Figure 6.
Expression patterns of 15 DEGs of tea plants under cold stress by qRT-PCR. (a) Expression patterns in LTSiL vs LTWL. All relative expression values were shown with log2 (fold change) on the Y axis, X-axis are genes. (b) Correlation between RNA-seq data and qRT-PCR detection. Each RNA-seq expression data was plotted against qRT-PCR data and subjected to linear regression.
Metabolomic analysis
-
Metabolomic studies were conducted to examine the metabolites involved in the cold-stress response of 'Fuding Dabaicha' under two different treatments. A total of 815 DAMs were identified between LTSiL and LTWL. Of these, 352 metabolites were found to be up-regulated, whereas 463 were down-regulated in LTSiL compared to LTWL (Fig. 7a). To further explore the metabolic differences underlying cold resistance, the top 50 DAMs induced by low temperature were clustered and analyzed, revealing that the levels of polyketides and flavonoids were significantly higher in LTSiL than in LTWL. Conversely, the levels of organooxygen compounds and their derivatives, phenylalanine, amino acids, and their derivatives, and organic acids were markedly lower in LTSiL (Fig. 7b). These changes in metabolite content might contribute to the enhanced cold resistance observed in LTSiL. Overall, these DAMs are likely to play a crucial role in mediating the response to cold stress in plants treated with silicon.
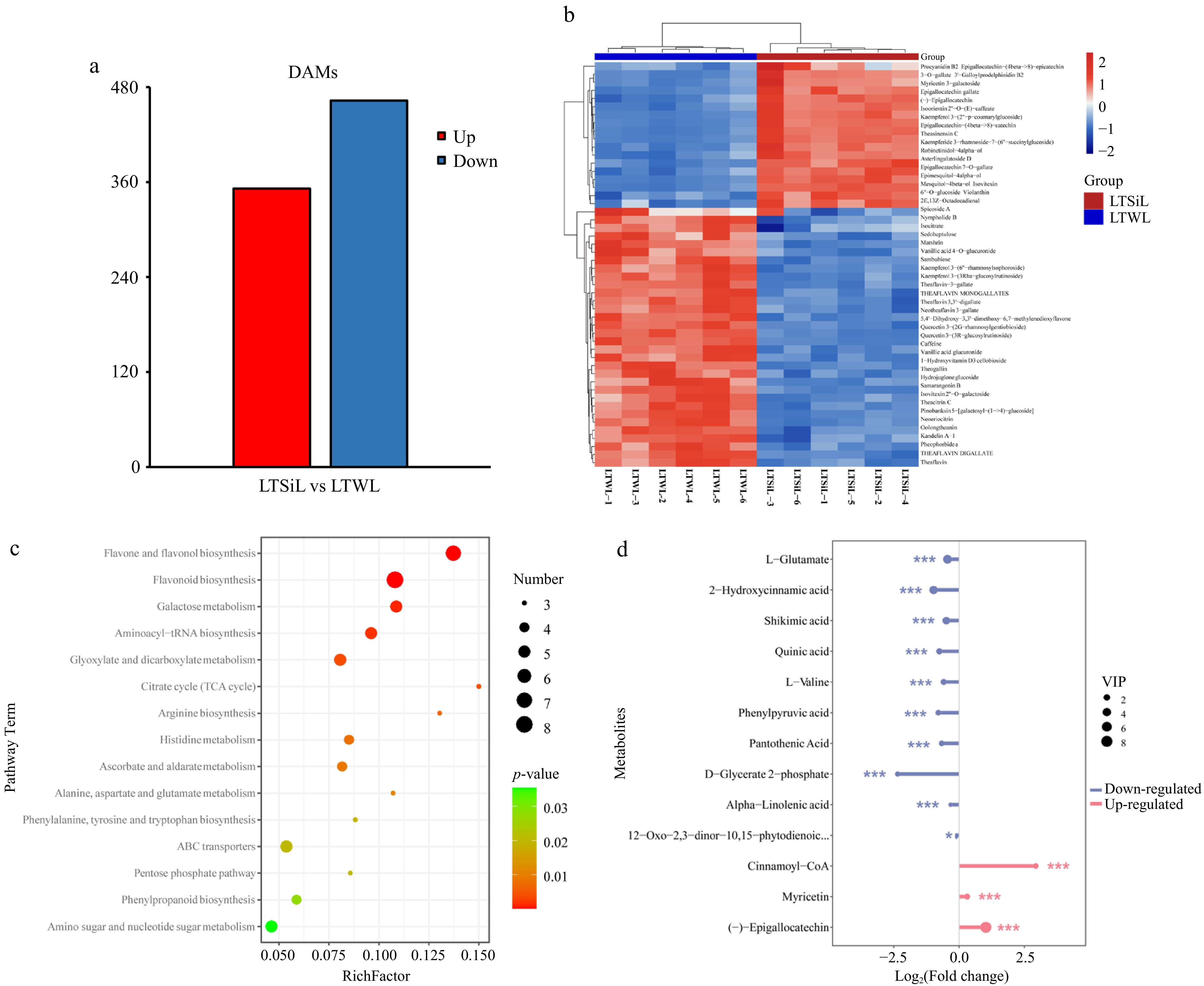
Figure 7.
Overview of metabolite analysis of tea plants subjected to 4 °C treatment. (a) The number of DAMs identified in the pairwise comparison of LTSiL vs LTWL in tea plants. (b) Hierarchical clustering analysis of the DAMs in LTSiL and LTWL. The abscissa indicates sample names and hierarchical clustering results; the ordinate indicates DAMs and hierarchical clustering results. 1, 2 and 3 refer to the three replicates. R Red color indicates high expression; blue color indicates low expression. (c) KEGG analysis of LTSiL and LTWL. X-axis represents Rich factor. The greater the Rich factor is, the greater the degree of enrichment is. The larger the point is, the greater the number of differential genes enriched in the pathway is. The redder the color of the dot is, the more significant the enrichment is. (d) Analysis of key metabolites between LTSiL and LTWL under cold treatment. X-axis represents types of differential metabolites. Y-axis represents differential metabolites fold change value.
In-depth KEGG pathway analysis was utilized to explore the metabolic pathways enriched in the DMAs between LTSiL and LTWL under cold stress. Key pathways identified include 'flavonoid biosynthesis', 'flavone and flavonol biosynthesis', 'galactose metabolism', 'aminoacyl tRNA biosynthesis', 'glyoxylate and dicarboxylate metabolism', 'arginine biosynthesis, and 'histidine metabolism', with 'flavonoid biosynthesis' and 'flavone and flavonol biosynthesis' showing the highest enrichment (Fig. 7c, Supplementary Fig. S2). These pathways suggest a potential link to cold tolerance, underscoring the need for further studies to elucidate the mechanisms by which silicon amelioration influences cold stress response.
Further analysis focused on key metabolites with significant changes, defined by a log2 (fold_change) greater than 1 and a p-value less than 0.05. As shown in Fig. 7d, flavonoid biosynthesis pathways were notably up-regulated in LTSiL compared to LTWL, while pathways involving terpenoid and alkaloid metabolites were significantly down-regulated. Specific metabolites, such as Cinnamoyl-CoA and (−)-Epigallocatechin, were up-regulated more than once, whereas others including D-Glycerate 2-phosphate, 2-Hydroxycinnamic acid, Phenylpyruvic acid, Pantothenic acid, Quinic acid, Shkimic acid, L-Valine, L-Glutamate, 12-oxophytodienoic acid (OPDA), and α-linolenic acid (α-LeA) were down-regulated. Notably, Cinnamoyl-CoA exhibited an eight-fold increase, and D-Glycerate 2-phosphate a five-fold decrease in LTSiL.
Comprehensive analysis of the metabolomic and transcriptomic data
-
The integrated analysis of transcriptomic and metabolomic data revealed significant overlaps in the KEGG pathways enriched for both DEGs and DAMs. Thirty-three KEGG pathways that were common to both DEGs and DAMs in LTSiL vs LTWL tea plants were identified (Fig. 8a). Furthermore, 17 KEGG pathways were significantly enriched (p < 0.05) for either DEGs or DAMs in both treatment groups (Fig. 8b). Notably, there was one pathway that was commonly and significantly enriched in both DEGs and DAMs across the two treatments, underscoring its potential critical role in the response mechanisms to cold stress in these plants.
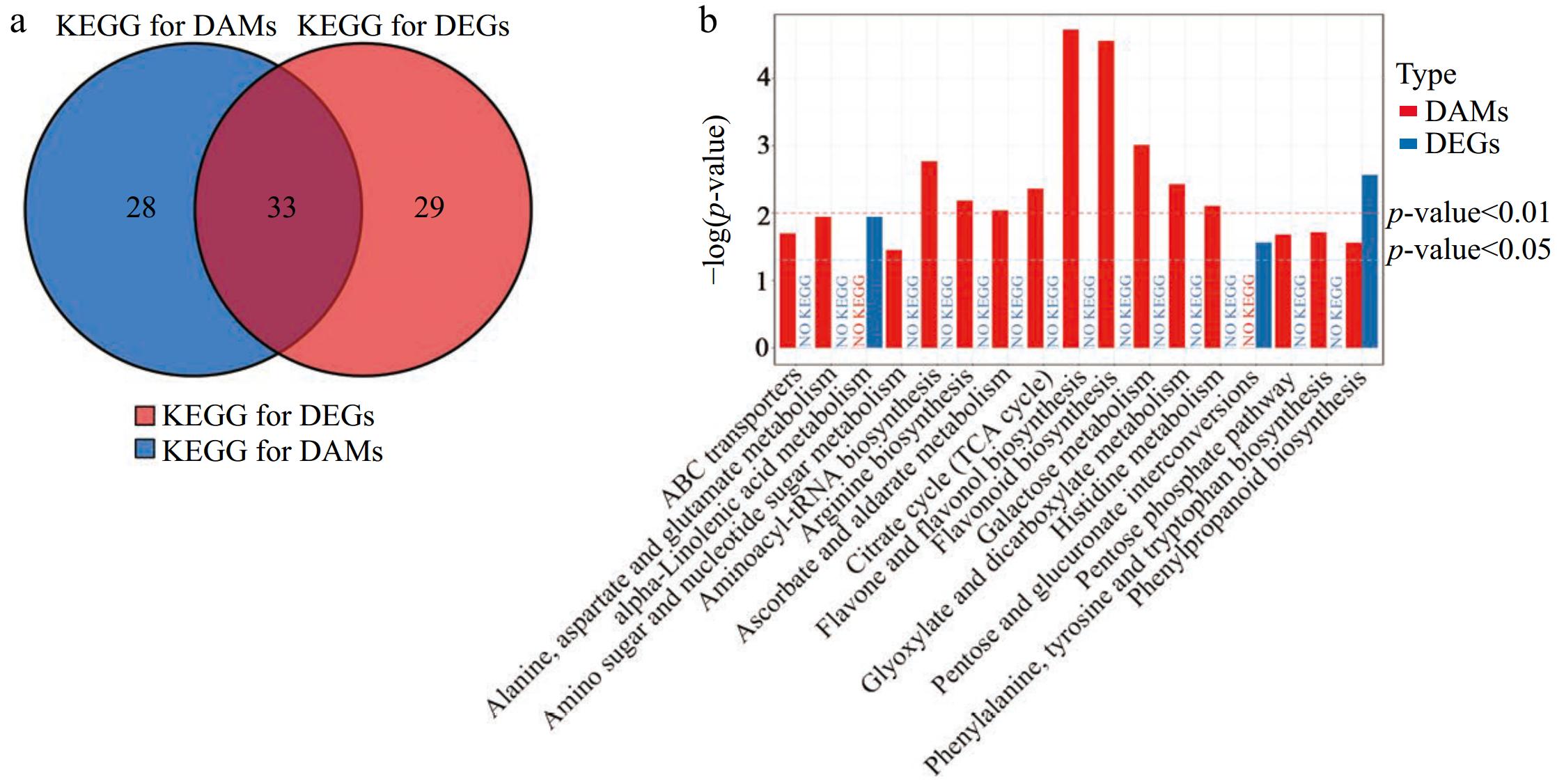
Figure 8.
Conjoint analysis of transcriptomic and metabolomics data. (a) Venn analysis of KEGG pathways for DEGs and DAMs. (b) Statistics of all KEGG pathways for DAMs and/or DEG with a p < 0.05 (b) in LTSiL vs LTWL.
-
In this study, the regulatory mechanisms of silicon-mediated amelioration were investigated in 'Fuding Dabaicha' under cold stress by conducting extensive biochemical, transcriptomic, and metabolomic analyses. Plants employ various physiological and metabolic strategies to protect against abiotic stresses. Significant physiological alterations were observed in 'Fuding Dabaicha' when exposed to cold stress without silicon amelioration. Furthermore, notable changes in metabolite levels and gene expression in 'Fuding Dabaicha', both with and without silicon treatment were detected through metabolomics and transcriptomics after 3 d of cold stress.
Physiological responses of tea plants with silicon amelioration under cold stress
-
Abiotic stress results in the accumulation of ROS, which destabilizes the plasma membrane in plant cells[58]. Both the REL and MDA levels serve as indicators of membrane permeability and can be utilized to assess plant cold tolerance. Increased levels of REL and MDA indicate greater damage to plants from cold exposure[9,59]. In the present study, the measurements of REL and MDA in LTSiL were found to be lower than those in LTWT during cold treatment (Fig. 2a & b). In addition, the activity of several antioxidant enzymes is enhanced to mitigate ROS-induced damage in plant cells[60], particularly through POD, a critical enzyme for scavenging ROS[61,62]. In this study, activities of SOD, POD, and CAT were observed to be higher in LTSiL compared to LTWL (Fig. 2c−e). Specifically, POD activities in LTSiL showed a continuous increase during cold exposure periods of 1, 3, and 5 d. By contrast, POD activities in LTWT were decreased after 3 d of cold exposure, slightly increased after 5 d, but remained substantially lower than LTSiL after day 1 (Fig. 2c).
Proline, commonly found in plants and acting primarily as an osmotic regulator, accumulates significantly under environmental stresses and assists in stabilizing subcellular structures and managing cellular redox potential during abiotic stress[63,64]. During the first day of cold stress, proline accumulation was gradual in LTWL but swift in LTSiL. Subsequently, proline levels decreased to relatively low levels in both LTWL and LTSiL, with a notably sharper decline observed in LTSiL compared to LTWL. However, by day 5 of cold stress, proline levels in LTSiL had nearly returned to pre-stress levels (Fig. 2f). These findings align with previous research indicating that the rapid breakdown of proline post-stress provides a substantial supply of reductants to aid recovery from stress-induced damage[65,66]. LTSiL is more resistant to low temperature. In addition, soluble sugars, crucial osmoregulatory molecules in plants, accumulate in the cytosol to prevent dehydration of the protoplasm and enhance cold tolerance[67]. In contrast to the relatively stable levels of soluble sugars in the LTWL treatment, LTSiL tea plants exhibited a peak in sugar accumulation at 3 d followed by a rapid decline (Fig. 2g). This pattern indicates an early osmotic response of soluble sugars to mitigate cold-induced damage in tea plant leaves. This decline in sugar content has been linked to reduced photosynthetic activity during cold exposure[4]. Consequently, the varying degrees of cold resistance observed between LTSiL and LTWT tea plants may result from differences in antioxidant enzyme activity and levels of osmoprotectants, which are influenced by silicon amelioration in tea plants.
Plant signal transduction pathways associated with silicon amelioration under cold stress
-
Phytohormones play a vital role in plant growth, development, metabolism, and particularly in adapting to and responding to cold stress[68,69]. Research has demonstrated that cold stress impacts plant growth and development[68,69]. For instance, exposure to 4 °C increases jasmonic acid (JA) levels in rice, and significantly elevates the transcription of genes such as OsAOS1/2, OsAOC, OsLOX2, OsOPR1/7, OsJAZ1, OsbHLH148, and OsCOI1a[70], suggesting that JA signaling plays a crucial role in rice's response to cold stress, where JA, as a signaling molecule, facilitates the production of proteins that counteract cold stress and enhances cold tolerance. In the present research, pathways related to plant signal transduction were notably enhanced in both LTSiL and LTWL tea plants, with numerous genes involved in jasmonic acid biosynthesis being modulated under cold conditions (Fig. 6a). Similarly, the findings indicate that in LTSiL compared to LTWL, the expression of AOC, AOS, and COI1, which are positive regulators of JA metabolism, was significantly elevated, potentially helped stabilize JA levels (Fig. 2h). These observations indicate that JA biosynthesis and signaling may regulate plant responses and adaptations to varying cold stress conditions when augmented by silicon treatment.
Flavonoid metabolites associated with silicon amelioration under cold stress
-
Flavonoid metabolites, significant secondary metabolites in plants, tend to accumulate substantially under environmental stress[57]. Flavonoids are known for their roles as ROS scavengers and as signaling molecules that activate defense-related pathways and regulatory mechanisms[71]. Studies have shown that the flavonoid content in Arabidopsis is positively correlated with cold resistance[72,73]. In this study, flavonoid metabolites were significantly upregulated in LTSiL compared to LTWL tea plants (Figs 4a, b & 7c). Previous research on tobacco under cold stress revealed a substantial accumulation of flavonoids, which was influenced by phenylpropanoid biosynthesis, highlighting the crucial role of flavonoids in cold stress regulation[29]. Additionally, the combined transcriptomic and metabolomic analyses demonstrated significant increases in both phenylpropanoid and flavonoid biosynthesis in LTSiL tea plants (Figs 7c, 8b). These findings suggest that silicon (LTSiL) can modulate the expression of flavonoid metabolites, thereby enhancing the cold tolerance of tea plants.
Amino acids metabolism associated with silicon amelioration under cold stress
-
Studies have shown that the number of free amino acids in plants increases under abiotic stress conditions[74,75]. However, plants may actively synthesize specific amino acids with beneficial roles during stress responses, making it crucial to identify these amino acids and understand their functions to fully comprehend the mechanisms behind enhanced stress resistance. To manage osmotic stress, plants reduce their intracellular osmotic potential by accumulating compatible solutes like proline[75]. Proline synthesis is vital for stress tolerance, as it also functions as a ROS scavenger[13]. The findings of this study (Fig. 2f) aligns with previous research, which demonstrated that proline synthesis is rapidly induced during cold stress[76,77]. Transcriptomic and metabolomic analyses revealed that, unlike some highly abundant amino acids such as proline, typically low-abundance amino acids also accumulate during abiotic stress due to increased protein turnover (Fig. 7d, Supplementary Fig. S2a). For example, experimentally validated conclusions indicate that branched-chain amino acids, such as valine, are degraded through partially overlapping pathways[45,78]. Additionally, Threonine (Thr) deamination, consistently down-regulated during abiotic stress (Figs 3b, 7c & Supplementary Fig. S2b), is recognized as the primary pathway for producing 2-oxobutyrate, a precursor for isoleucine synthesis[79,80]. Moreover, plant stress response involves the activation of intricate signaling networks, leading to significant metabolic changes across the plant[81,82]. Furthermore, amino acid catabolism plays a crucial role in maintaining metabolic homeostasis within the stress-activated pathways[78]. For example, under stress conditions, plants activate alternative pathways to supply substrates for mitochondrial respiration. Proteins and branched-chain amino acids, such as valine (Val), undergo degradation (Figs 3f, 7d & Supplementary Fig. S2a), and the alternative ETF/ETFQO respiratory pathway is engaged, aiding in the mitigation of stress-induced damage[83−85]. Plants also respond to stress by degrading aromatic amino acids like Gly, Tyr, Trp, and Phe (Figs 3a, c−e, 7c & Supplementary Fig. S2c), which serve as substrates for the synthesis of hormones and secondary metabolites[86].
-
The present analysis found that the cold resistance of 'Fuding Dabaicha' in silicon amelioration treatment (LTSiL) was significantly higher than that in the control treatment (LTWL) through the physiological indexes. Compared with LTWL, the expression level of some genes involved in TFs, plant hormones, and JA signaling pathways were significantly up-regulated in LTSiL through the transcriptomics analysis, besides that flavonoid metabolites were also significantly enriched and amino acids metabolites were significantly varied through the metabolomics analysis of LTSiL under cold stress.
This research was supported by the Key Research and Development Project from the Department of Science & Technology of Tai'an City, China (2022NYLZ08), and the National Natural Science Foundation of China (32302437). We thank A&L Scientific Editing (
www.alpublish.com ) for their expertise in content and language editing during the preparation of this manuscript.-
The authors confirm their contributions to the paper as follows: methodology, data curation: Han X; experimentation, visualization: Yang S, Xie X, Zhao X; initial draft preparation: Xie X; manuscript review: Han X; provision of resources: Shang T; funding acquisition, project supervision: Han X, Xie X. All authors have reviewed the results and approved the final version of the manuscript.
-
Due to administrative requirements, the datasets generated and/or analyzed during this study are not publicly available but are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xueying Xie, Shuqing Yang
- Supplementary Table S1 Primers used for qRT-PCR.
- Supplementary Fig. S1 Overview of the transcriptome analysis of tea plants subjected to 4 ◦C treatment.
- Supplementary Fig. S2 Pathways related to amino acid metabolism under low temperature stress.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xie X, Yang S, Zhao X, Shang T, Han X. 2025. Mechanisms of silicon-mediated amelioration for Camellia sinensis using physiology, transcriptomics, and metabolomics under cold stress. Beverage Plant Research 5: e003 doi: 10.48130/bpr-0024-0034 

Mechanisms of silicon-mediated amelioration for Camellia sinensis using physiology, transcriptomics, and metabolomics under cold stress
- Received: 20 June 2024
- Revised: 27 August 2024
- Accepted: 23 September 2024
- Published online: 12 February 2025
Abstract: Cold stress significantly impacts the growth, quality, and yield of tea plant leaves. Additionally, silicon (Si) is a widely abundant element on earth and is recognized for its beneficial effect on plant growth and development. Here, a comprehensive transcriptomic, biochemical, and metabolomic investigations on tea plant leaves was conducted, examining the impact of Si-mediated improvement during short- (1 d), medium- (3 d), and long-term (5 d) exposure to cold stress. Biochemical and physiological analyses demonstrated that Si-mediated amelioration enhanced the activity of protective enzyme systems during all three cold stress treatments. There was a marked increase in soluble sugar content and a significant decrease in chlorophyll B content during the medium-term cold stress. Integrated transcriptomic and metabolomic analyses revealed that the differentially expressed genes (DEGs) primarily targeted pathways in phenylpropane biosynthesis, α-linolenic acid metabolism, pentose and glucuronic acid interconversion, and nitrogen metabolism. Similarly, differentially abundant metabolites (DAMs) linked to cold stress response in tea plants predominantly involved flavone and flavonol biosynthesis, flavonoid biosynthesis, aminoacyl-tRNA biosynthesis, galactose metabolism, and phenylpropanoid biosynthesis. The integrated analysis of the metabolites associated with these pathways and expression profiles further confirmed the involvement of the identified DEGs in responding to cold stress and the beneficial effect of Si in tea plants. This research expands our knowledge of the regulatory networks underlying cold stress responses in tea plants.
-
Key words:
- Camellia sinensis /
- Cold stress /
- Silicon /
- Plant physiology /
- Transcriptomics /
- Metabolomics