









-
Alfalfa (Medicago sativa L.) is a perennial legume that is widely recognized for its rich nutritional profile and exceptional quality as a fodder crop. It has a well-developed root system, vigorous tillering, and abundant foliage, along with key advantages such as high yield, excellent quality, strong adaptability, and broad distribution. It has become a widely utilized forage species with the largest planted area in China[1]. However, the development and yield of alfalfa are limited by extreme environmental stressors, and soil salinization is particularly detrimental[2]. Therefore, deciphering how plants cope with salt stress is a crucial step in breeding salt-tolerant crops, which is essential for satisfying the steadily rising global food demand.
Glutathione S-transferases (GSTs), an ancient family of proteins, play crucial roles in various catalytic and regulatory functions. These enzymes are instrumental in the S-conjugation process, which involves the attachment of the thiol group of glutathione (GSH, composed of glutamic acid, cysteine, and glycine) to the electrophilic groups of hydrophobic and potentially harmful substrates[3]. Their principal role involves detoxifiying noxious compounds of exogenous origin, such as herbicides and foreign pathogens, or endogenous stress-induced metabolites. GSTs can be induced by a variety of stresses and play a direct role in reducing oxidative damage or in detoxification[4,5]. Furthermore, GSTs can be activated by various phytohormones such as abscisic acid (ABA), auxin, ethylene, salicylic acid, cytokinins, and methyl jasmonate, which are essential for plant growth and development[6,7]. GSTs are key regulators of signaling pathways and are vital for numerous developmental processes, including apoptosis and growth modulation[8]. In photosynthetic eukaryotes, the GST family is categorized into 14 distinct classes, including tau (GSTU), phi (GSTF), zeta (GSTZ), theta (GSTT), and tetrachlorohydroquinone dehydrogenase (TCHQD), which feature GSTs that possess serine at their active sites. Iota (GSTIs), DHARs, hemerythrins (GSTHs), GHRs, lambda (GSTLs), mPGES-2s, and metaxins are categorized as Cys-GSTs, while the catalytic residues in the EF1Bγ and Ure2p classes remain undefined[9,10]. Specifically, GSTUs, GSTFs, GSTLs, and TCHQDs are related to plants[11].
In plants, the function of GSTs is closely related to stress response. For instance, the BES/BZR family transcription factor TaBZR2 enhances the drought resistance of wheat (Triticum aestivum) through the activation of TaGST1 and mediates the crossover between the brassinosteroid (BR) and drought signaling pathways[12]. In poplar (Populus alba × Populus glandulosa), salt stress markedly upregulates the expression of PtrGSTF8, and the overexpression of PtrGSTF8 enhances salt tolerance in tobacco by reducing reactive oxygen species (ROS) levels[13]. GSTU is crucial for scavenging hydrogen peroxide (H2O2) and superoxide anions (O2·−) to maintain ROS homeostasis under various stresses[14]. Arabidopsis plants with increased expression of AtGSTU19 demonstrate enhanced resistance to salt, drought, and methyl viologen stress[15]. Ectopic expression of rice OsGSTU4 in Arabidopsis can enhance the tolerance to salt and oxidative stress at different developmental stages[16]. MsCML10 interacts with MsGSTU8 and MsFBA6, activating them to maintain ROS homeostasis at low temperatures, promote sugar accumulation, and ultimately enhance the cold tolerance of alfalfa[17]. Overexpression of tomato (Lycopersicon esculentum) LeGSTU2 enhances the resistance to NaCl and mannitol-induced salt and osmotic stress in transgenic Arabidopsis[18]. In poplar, PtrGSTU23/40 overexpression regulates the activity of GST enzymes to modulate ROS scavenging capacity, ultimately enhancing salt tolerance[19]. The jasmonic acid signaling pathway mediated by CsJAZ1-CsMYC2.2 regulates tea plant (Camellia sinensis [L.] O. Kuntze) sensitivity to Colletotrichum camelliae by modulating the accumulation of H2O2 through CsGSTU45[20]. Overexpression of Glycine soja GsGSTU13 enhances salt tolerance in alfalfa[21]. In Medicago truncatula, an expression profile analysis of GST family genes showes that salt stress significantly induces or inhibites the expression of GST genes, indicating that GST family genes are closely related to salt stress reactions[22]. However, the extent to which the GST family genes in salt stress tolerance in alfalfa remains unclear.
Salt-responsive genes in alfalfa have been explored using transcriptomics[23]. MsGSTU17 is upregulated in the highly salt-tolerant cultivar 'Magnum Salt' following salt treatment[23]. However, the salt tolerance function of MsGSTU17 in alfalfa has not been confirmed. In the present study, we identified MsGSTU17 as a gene that responds to salt stress and encodes a protein belonging to the GST family. Furthermore, we studied the salt tolerance of transgenic Arabidopsis plants overexpressing MsGSTU17 and its role in regulating salt tolerance by modulating salt-response genes and the antioxidant defense system.
-
Medicago sativa (Zhongmu No.1) and Nicotiana benthamiana were grown in a growth medium consisting of fertile soil and perlite in a 2:1 volumetric ratio. They were cultivated under controlled light conditions, with an irradiance of 200 μmol·m−2·s−1 and a 16 h/8 h (light/dark) cycle in a greenhouse maintained at 23 °C. Seeds from the Columbia ecotype (Col-0) of Arabidopsis thaliana were disinfected with immersion in a 10% (v/v) solution of sodium hypochlorite for 16 min to eliminate surface contaminants, followed by three rinses with sterile water. Sterilized seeds were sown on a medium containing 1/2 Murashige and Skoog (MS) nutrients and stored at 4 °C for 3 d to promote germination. Seedlings (7 days old) were replanted into a soil matrix that included a blend of nutrient-rich soil and perlite at a volume ratio of 2:1, and accommodated in a growth chamber at 23 °C with an irradiance of 200 μmol·m−2·s−1 and a 16 h/8 h (light/dark) cycle.
Sequence analysis
-
The sequences of the MtGST gene family, used as baits to search for the target sequence of alfalfa, were obtained from Hasan et al.[22]. Supplementary Table S1 provides information on MsGST proteins. The phylogenetic tree was generated using MEGA X software, employing the maximum-likelihood (ML) method with parameters set to JTT + G + F and 1,000 bootstrap replicates. The amino acid sequences of MsGSTU17 homologs, MtGSTU17, and MtGSTU18 from Medicago truncatula and AtGSTU25 from Arabidopsis, were aligned using DNAMAN software. The sequences of MtGSTU17, MtGSTU18, and AtGSTU25 were obtained from Phytozome (
https://phytozome.jgi.doe.gov/ ). The 2,000 bp genomic DNA sequence upstream of the transcription start site of MsGSTU17 was retrieved from the Medicago sativa genome[24]. The cis-acting elements in the promoter region of MsGSTU17 were identified through the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ). The transmembrane domains and signal peptide of the MsGSTU17 protein were predicted by analyzing its protein sequence using the TMHMM Server v. 2.0 (https://services.healthtech.dtu.dk/services/TMHMM-2.0/ ) and SignalP-5.0 (http://www.cbs.dtu.dk/services/SignalP/ ), respectively.Subcellular localization of MsGSTU17 protein
-
The coding region of MsGSTU17, excluding the termination codon, was amplified and inserted into the pCAMBIA-1305 vector to create a fusion protein with a green fluorescent protein (GFP) driven by the CaMV 35S promoter. Using Agrobacterium-mediated transformation, the recombinant construct, along with an empty vector as a control, was co-transformed into the abaxial side of four-week-old Nicotiana benthamiana leaves, together with the 35S::AtAKT1-mCherry vector. After 48−72 h of transformation, fluorescence was observed using a confocal laser scanning microscope (Carl Zeiss SAS, Jena, Germany).
Quantitative real-time PCR analysis (qRT-PCR)
-
Total RNA was extracted using the RNAprep Pure Plant Kit (Tiangen Inc., Beijing, China), and cDNA was synthesized using HiScript III RT SuperMix for qPCR (Vazyme, Nanjing, China). The qRT-PCR was conducted using the Thermal Cycler Dice Real-Time System from Takara (Otsu, Japan) with diluted cDNA as the template. The alfalfa β-actin gene (JQ028730.1) served as the internal control for normalizing template quantities and Arabidopsis actin gene was similarly utilized for the normalization of template amounts[25]. The normalized (2−ΔΔCᴛ) method was utilized to measure the relative gene expression levels. Supplementary Table S2 provides a comprehensive list of the primers used.
Generation of transgenic plants
-
We inserted the coding DNA sequence (CDS) of MsGSTU17 into the pGAMBIA3301 vector driven by the CaMV 35S promoter. Arabidopsis thaliana (Col-0) plants at four weeks of age were genetically transformed using the Agrobacterium tumefaciens strain EHA105 harboring the recombinant plasmid used for gene expression by employing a modified flower dipping method. We subjected the progeny to Basta selection and collected homozygous T3 transgenic seeds for further analysis.
Salt tolerance assays
-
Sterile seeds of both wild type (WT) and transgenic Arabidopsis were transferred onto 1/2-strength MS medium. Seeds were subjected to 3 d of cold stratification at 4 °C, followed by a 4-d incubation at 22 °C for further growth. The germinated seeds were transferred to fresh medium with varying concentrations of NaCl (0, 100, and 125 mM), and the seedlings were cultivated vertically for 12 d in a growth chamber to measure root elongation and fresh weight. To determine salt tolerance, plants grown on 1/2 strength MS medium were transferred to soil. They were then allowed to grow for 16 additional days in a growth chamber at 22 °C. Irrigation of the plants commenced with 3-d treatments with 200 and 250 mM NaCl solutions, followed by a subsequent 6-d treatment with 300 mM NaCl solution. Electrolyte leakage was assessed as previously described[26]. The survival rate was calculated by counting the surviving plants after a 12-d treatment with 300 mM NaCl[27].
Measurements of enzyme activities
-
To extract superoxide dismutase (SOD) and catalase (CAT), 0.2 g of fresh leaf material was flash-frozen in liquid nitrogen, then ground into a fine powder in 3 mL of ice-cold 50 mM phosphate buffer (pH 7.8) using a mortar and pestle. For ascorbate peroxidase (APX) extraction, leaves were homogenized in 3 mL of 50 mM phosphate buffer (pH 7.0), cooled on ice, and enriched with 1 mM EDTA and 1 mM ascorbic acid (AsA). Both homogenates were centrifuged at 12,000 × g for 20 min at 4 °C and the supernatants were collected for subsequent enzyme activity and protein concentration assays following the method described by Guo et al.[28]. To determine the GST and glutathione reductase (GR) activities and GSH content, 0.2 g of the sample was homogenized in sodium phosphate buffer and centrifuged, then the absorbance of the supernatant was measured using a spectrophotometer at specific wavelengths using commercial kits as per the manufacturer's instructions (Grace Biotechnology, Suzhou, China)[15].
3,3'-diaminobenzidine (DAB) staining and nitrioblue tetrazolium (NBT) staining
-
To detect H2O2, leaf samples were incubated for 1 h in DAB solution at a concentration of 1 mg·mL−1. To identify O2·−, the leaflets were treated with a 5 mg·mL−1 NBT solution for 12 h in darkness[29]. To facilitate imaging and deplete chlorophyll, the leaflets were immersed in a 95% (v/v) ethanol solution.
Statistical analysis
-
All data were analyzed using a completely randomized design model with the SPSS software (SPSS Inc., Chicago, IL, USA). To determine the significance of mean differences among treatments or plant lines, ANOVA was implemented, followed by Duncan's test to ascertain significance at the 0.05 level.
-
To explore the role of MsGSTU17, a 675 bp open reading frame (ORF) encoding a 224-amino acid peptide (GenBank accession No. PQ816732) was isolated from the salt-tolerant alfalfa cultivar 'Zhongmu No.1' . It was highly homologous to MtGSTU17 based on the phylogenetic analysis of Medicago truncatula (Fig. 1). Multiple sequence alignments of MsGSTU17 with MtGSTU17 (Medtr3g467420), MtGSTU18 (Medtr3g467430), and AtGSTU25 (At1g17180) revealed high sequence identities of 94%, 89%, and 65%, respectively (Fig. 1). The Pfam database revealed that MsGSTU17 contains two conserved domains, GST-N and GST-C, which are characteristic of the GST family (Supplementary Fig. S1a). Structural modeling of MsGSTU17 predicted seven distinct protein folds (Supplementary Fig. S1b). The 2,000 bp promoter segment of MsGSTU17 was scrutinized for cis-regulatory elements using the PlantCARE platform. The existence of four ABA-responsive, one MeJA-responsive, and five low-temperature-responsive elements suggests that MsGSTU17 is involved in stress response mechanisms (Supplementary Fig. S1c).
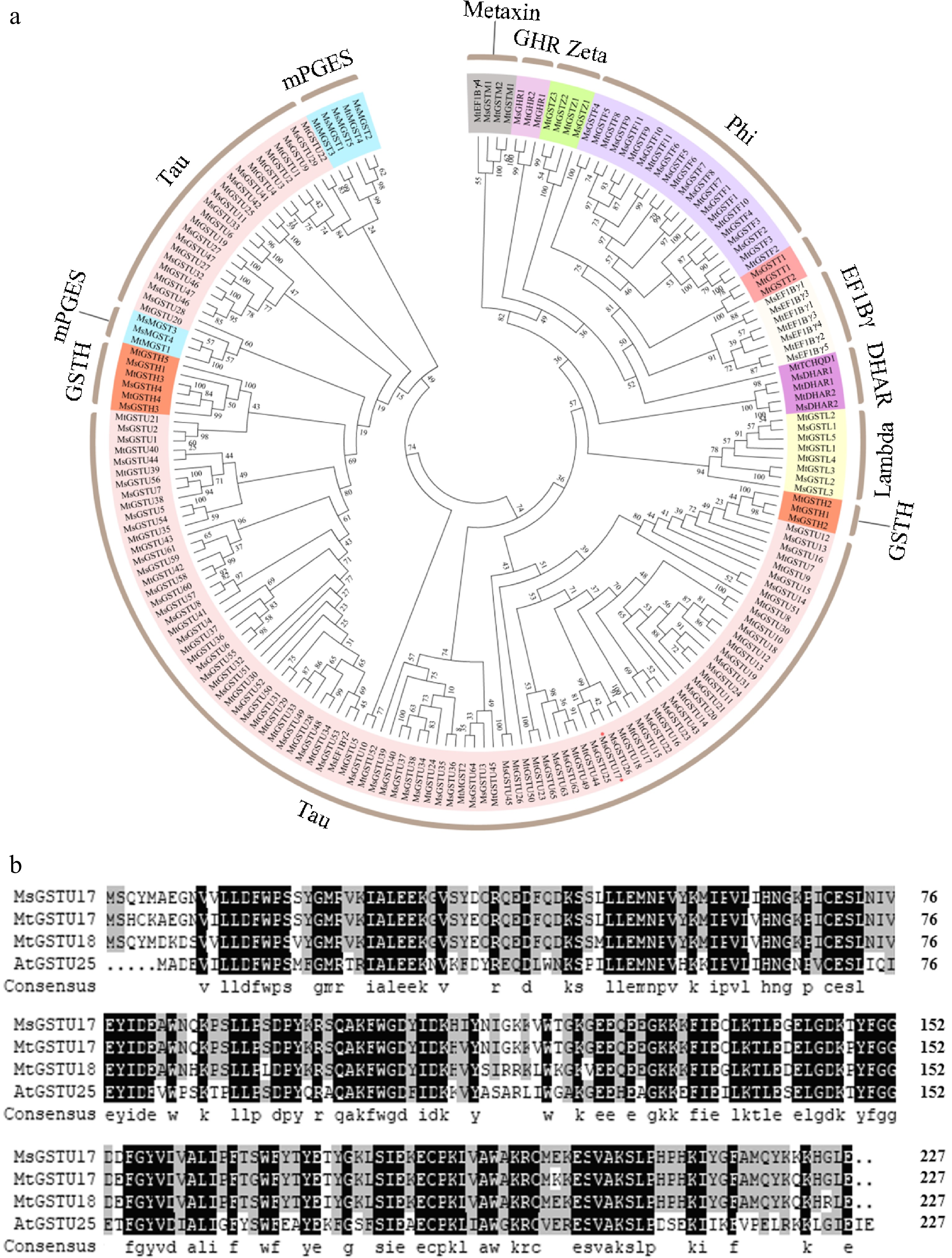
Figure 1.
Phylogenetic evaluations and alignments of multiple GST sequences from various plant species were performed. (a) Phylogenetic tree of GSTs constructed based on Medicago truncatula and alfalfa. (b) Multiple sequence alignment of MsGSTU17, MtGSTU17, MtGSTU18, and AtGSTU25 was performed. Those amino acids which were the same are distinguished by a black background, while those with a similarity of ≥ 75% are represented in gray.
Subcellular localization of MsGSTU17 and its expression pattern
-
Analysis of MsGSTU17 revealed the absence of transit peptides, signal peptides, and transmembrane domains (Supplementary Fig. S2), and evidence strongly suggested that MsGSTU17 is localized to the cytoplasm. To validate this hypothesis, the cellular localization of MsGSTU17 was assessed by co-expressing MsGSTU17-GFP and AtAKT1-mCherry in tobacco leaves. The data revealed that the MsGSTU17 fluorescence signal did not coincide with the AtAKT1 fluorescence signal (Fig. 2), confirming that MsGSTU17 was localized in the cytoplasm.
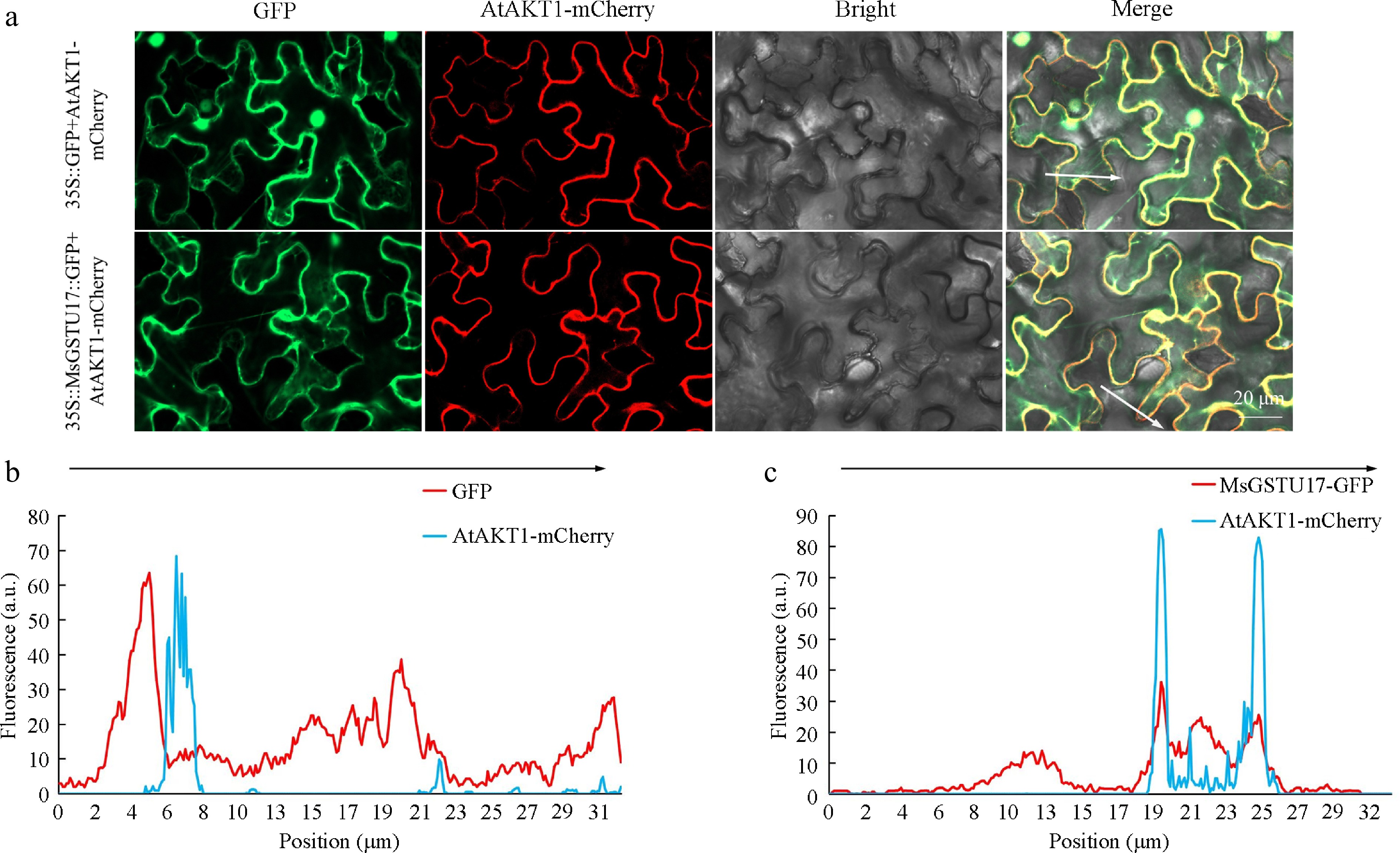
Figure 2.
Subcellular localization analysis of MsGSTU17. (a) The study evaluated the colocalization of MsGSTU17-GFP and AtAKT1-mCherry in the context of tobacco leaf tissue. Presented are representative laser-scanning photographs illustrating the fluorescence of GFP, the fluorescence of mCherry, the bright field, and an overlay combining all three. The fluorescence intensities (in arbitrary units) that crossed the tobacco leaf cell (indicated by the white line) along the arrow in (b) 35S::GFP, and (c) MsGSTU17-GFP were analyzed using ImageJ software. Scale bars = 20 μm.
The tissue-specific expression pattern of MsGSTU17 was examined using qRT-PCR. The findings revealed that MsGSTU17 was expressed in all tissues tested, including the leaves, stems, flowers, roots, nodules, and seeds of alfalfa. Notably, its expression was highest in the leaves, whereas lower levels were observed in the roots and stems (Fig. 3a). After salt treatment, MsGSTU17 was expressed in both the roots and leaves. Transcription levels of MsGSTU17 increased in both the roots and leaves after 2 h of salt stress and decreased after 24 h of salt stress (Fig. 3).
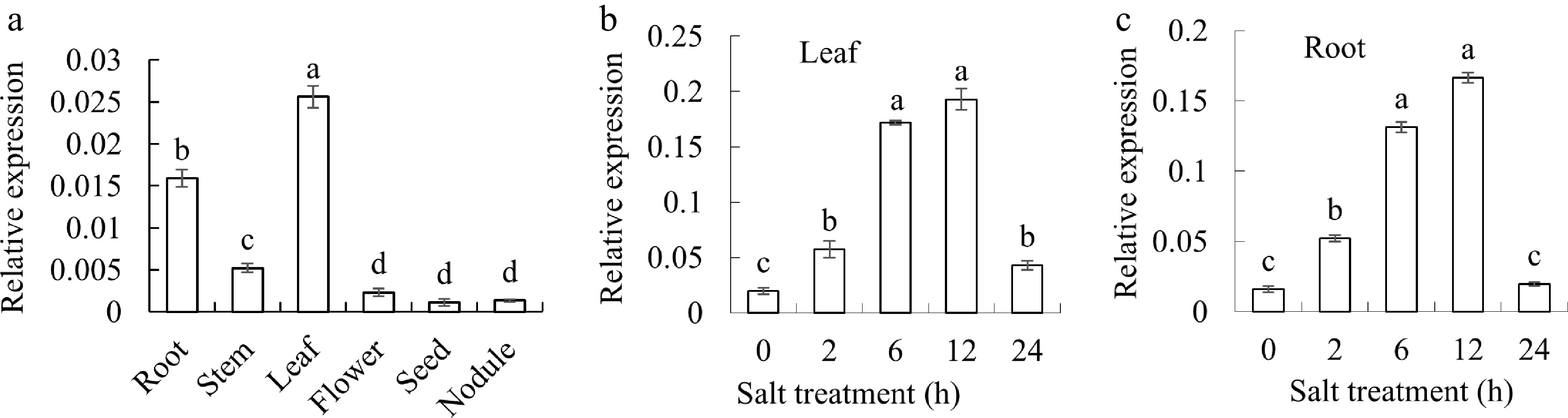
Figure 3.
Analyzing the expression profile of MsGSTU17 gene. (a) Investigating the expression patterns of MsGSTU17 across diverse tissues of Arabidopsis, encompassing roots, stems, leaves, flowers, seeds, and nodules. Under conditions of salt stress, a quantitative assessment of MsGSTU17 expression was conducted in both (b) leaves, and (c) roots. The mean values with their standard deviations are depicted in the bar charts (n = 3), at the significance level of p < 0.05, bars on the chart that bear the same letter indicate no statistically differences.
Overexpression of MsGSTU17 augmented the salinity endurance of transgenic Arabidopsis
-
Transgenic Arabidopsis plants overexpressing MsGSTU17 were successfully created, and the presence of MsGSTU17 transcripts was confirmed in two homozygous lines (OE6 and OE13). In contrast, no transcripts were detected in the wild-type (WT) plants (Fig. 4a). The transgenic lines had significantly longer roots than the WT plants under salt stress. Despite the general growth inhibition caused by salt stress, transgenic plants sustained a higher fresh weight than WT plants (Fig. 4). To investigate the involvement of MsGSTU17 in salt tolerance, plants that had been cultivated in soil for three weeks underwent irrigation with 300 mM NaCl solution for 12 d. Under salt stress, the survival rate of the transgenic lines ranged from 66.67%–70.83%, which was significantly higher than that of the wild type (33.33 %), and their ion leakage rates were also notably lower than that of the WT plants (Fig. 5). These results suggest that MsGSTU17 overexpression significantly improved the salt tolerance of transgenic plants.
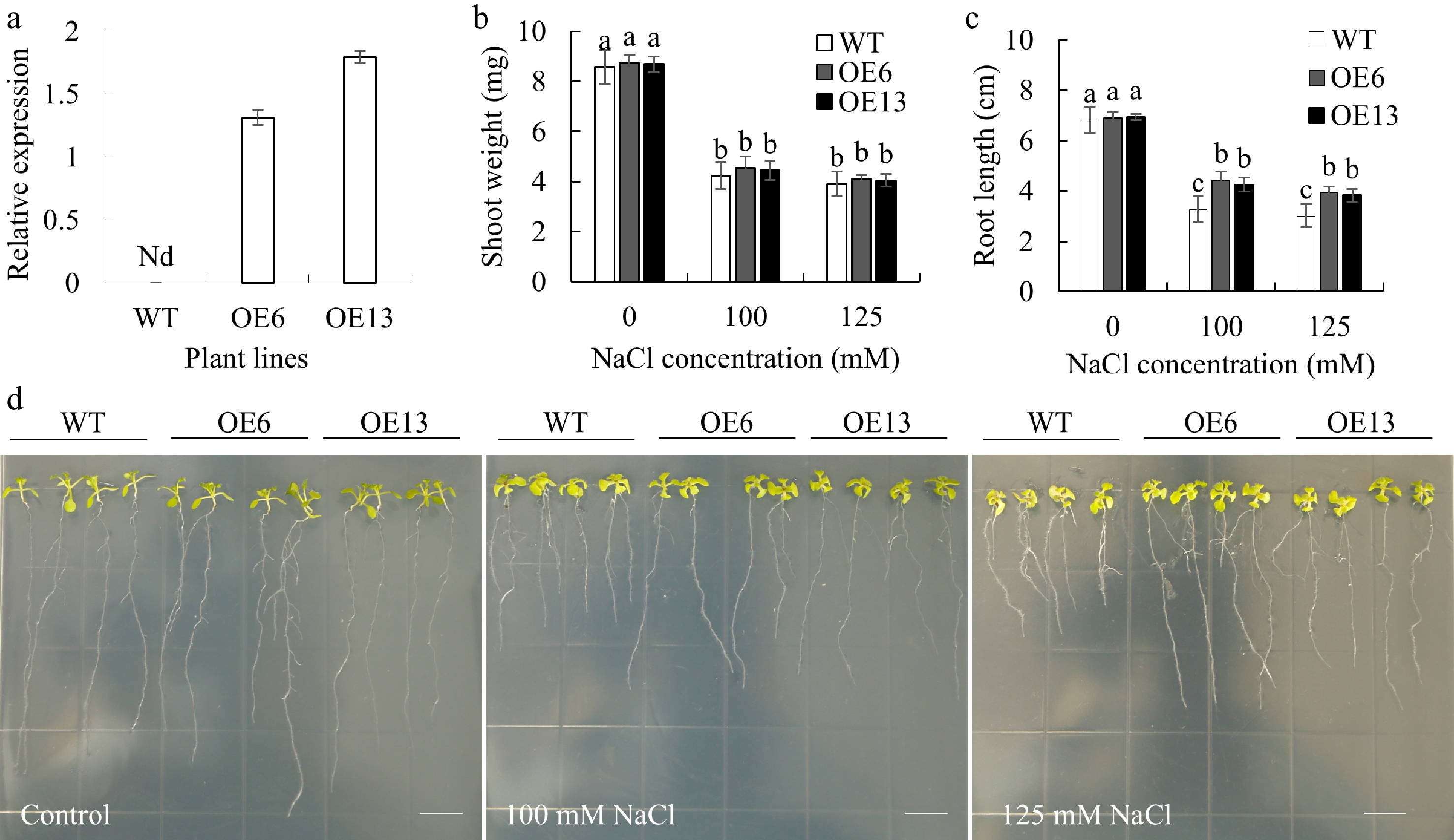
Figure 4.
The root length and shoot fresh weight of transgenic Arabidopsis plants overexpressing MsGSTU17 after salt stress treatment. WT (Col-0) and MsGSTU17 overexpressing seedlings (OE6 and OE13) were included in the treatment. (a) qRT-PCR analysis of MsGSTU17 transcription levels in plants. The (b) fresh shoot weight, and (c) root length of each group of seedlings after 12 d of growth in 1/2 MS medium containing different NaCl concentrations. The same letters above the bar indicate no significant difference in the corresponding items (p < 0.05). The sample size of seedlings in each experiment is greater than 30. (d) Phenotype of each group of seedlings grown on 1/2 MS medium with different NaCl concentrations for 12 d. Bar = 10 mm.
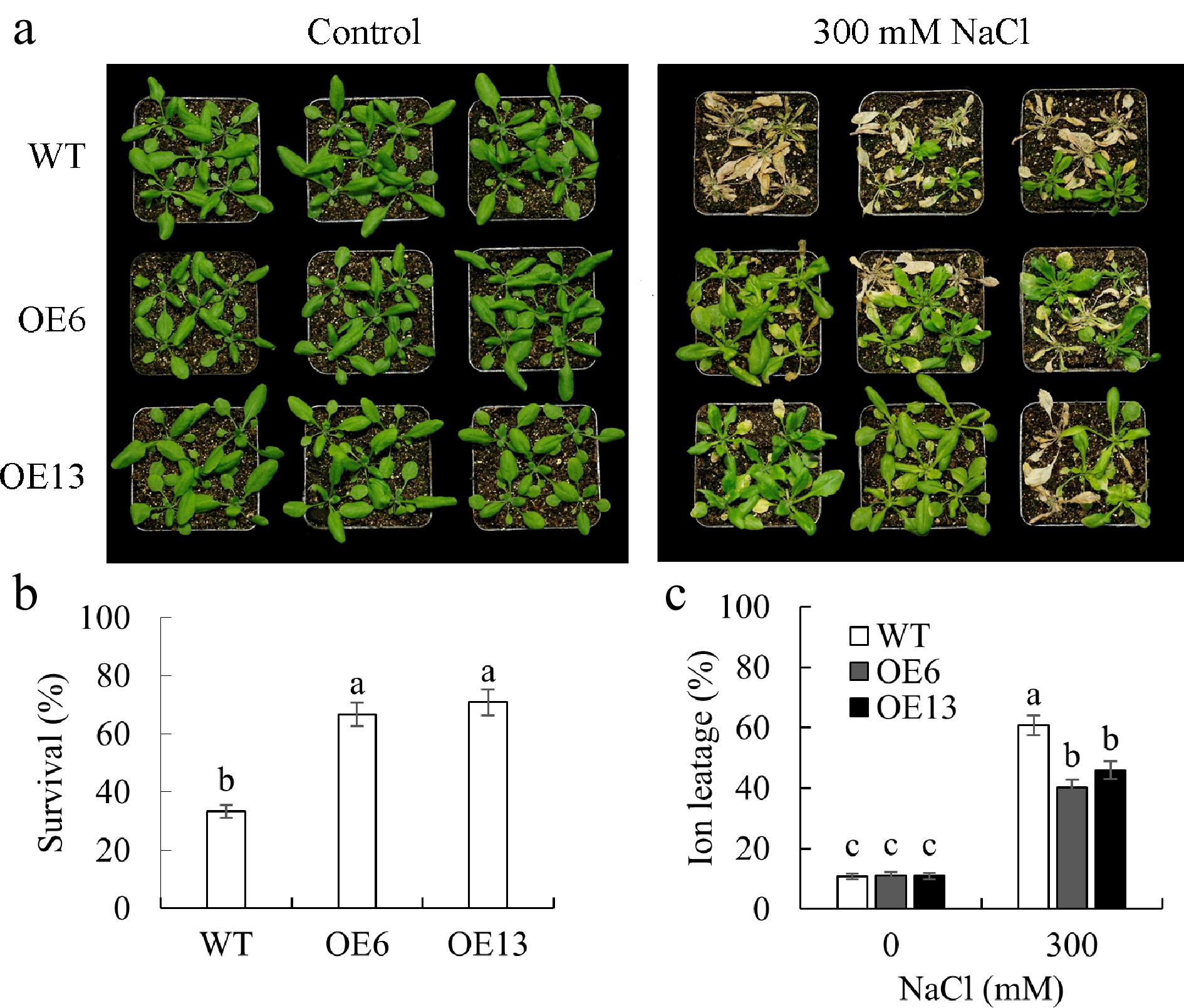
Figure 5.
The survival performance of transgenic Arabidopsis plants overexpressing MsGSTU17 in a salt stress environment was investigated. WT (Col-0) and MsGSTU17 overexpressing seedlings (three weeks old) were treated with 200, 250, and 300 mM NaCl for 3, 3, and 6 d, respectively. (a) The photos depicting the growth status of seedlings. (b) The survival rate. (c) The degree of ion leakage in the leaves is represented as the mean ± standard deviation (n = 3 replicates, four plants/replicate). The same letters above the bar chart signified that there was no significant difference in the corresponding items (p < 0.05).
ROS accumulation and antioxidant enzyme activities
-
The NBT and DAB staining results showed that under control conditions, there were no differences in O2·− and H2O2 levels between transgenic and WT plants. The relative increase in O2·− and H2O2 levels in transgenic plants was lower than that in WT plants under salt stress. The activities of SOD, CAT, and APX were similar between the WT and transgenic plants under control conditions. However, transgenic plants exhibited a more substantial increase in enzyme activity than WT plants under salt stress (Fig. 6).
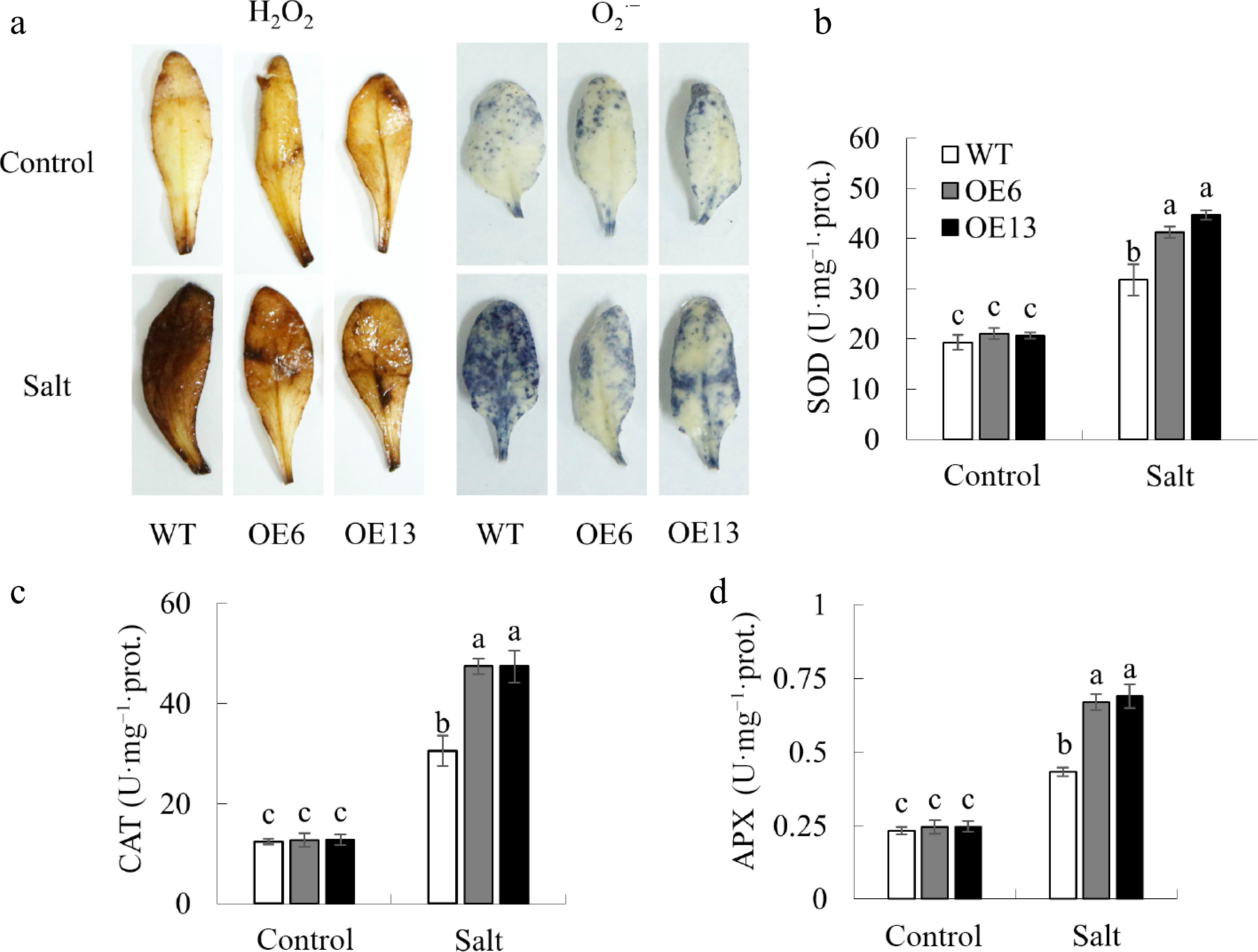
Figure 6.
The impact of MsGSTU17 overexpression on the activity of antioxidant enzymes and accumulation of ROS. (a) Reactive oxygen species, including O2·− and H2O2, were detected in leaves. The enzymatic activities of (b) SOD, (c) CAT, and (d) APX were quantified. The data are presented as mean ± SD based on three independent replicates. The same letter above the bars indicates that there is no significant difference between the groups (p < 0.05).
Changes in contents of GSH and related enzymes under salt stress
-
Compared to the WT plants, transgenic Arabidopsis exhibited markedly higher levels of GST activity and GSH content under control conditions, without notable variation in GR activity. After salt treatment, GST and GR activities increased in all genotypes, whereas the GSH content decreased. A comparative analysis of WT and transgenic plants under salt stress revealed that transgenic Arabidopsis thaliana exhibited significantly elevated GST and GR activities, as well as increased GSH levels, compared to those in WT specimens (Fig. 7).
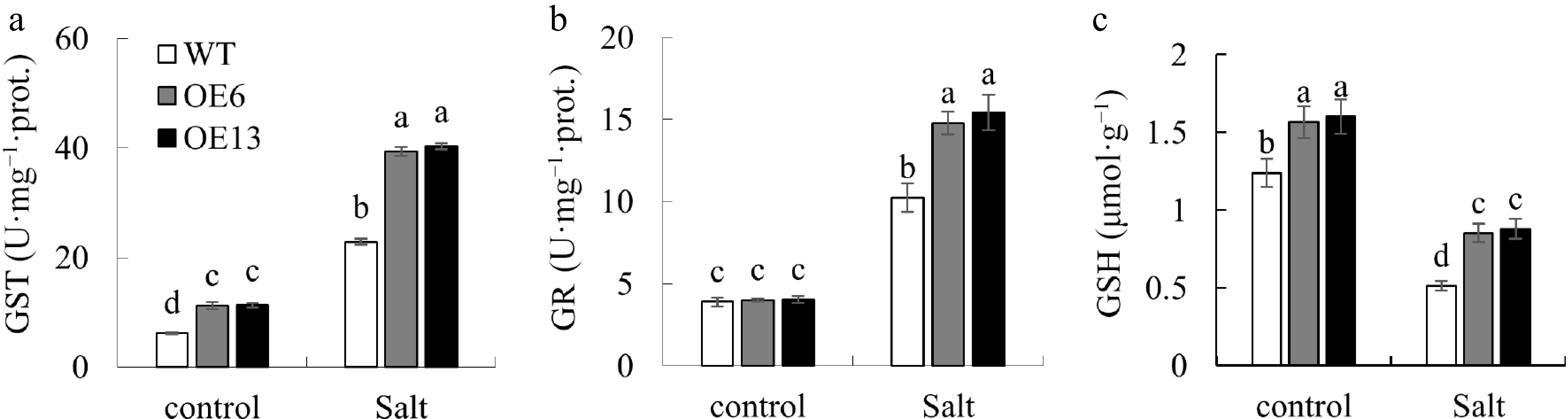
Figure 7.
Changes of GSH content and related enzymes. Variations in (a) GST, and (b) GR activities, along with (c) GSH levels, were observed in 15-day-old wild-type and transgenic Arabidopsis plants following a 20-d exposure to 0 or 200 mM NaCl. Distinctive letters above the bars signify statistically significant disparities among the treatments at the p < 0.05 level.
Transcriptional analysis of gene expression patterns under salt stress conditions
-
To investigate how MsGSTU17 responds to salt stress, we analyzed the expression levels of the salt stress-related genes SOS1, SOS2, SOS3, RD29A, KIN2, and HKT1 in 3-week-old plants treated with 200 mM NaCl using qRT-PCR. Under normal conditions, the transgenic lines showed no significant difference in the expression of these genes, except for SOS2, when compared to wild-type plants. However, under salt stress, the expression of all these genes was markedly higher in the transgenic lines than in the wild-type plants (Fig. 8).
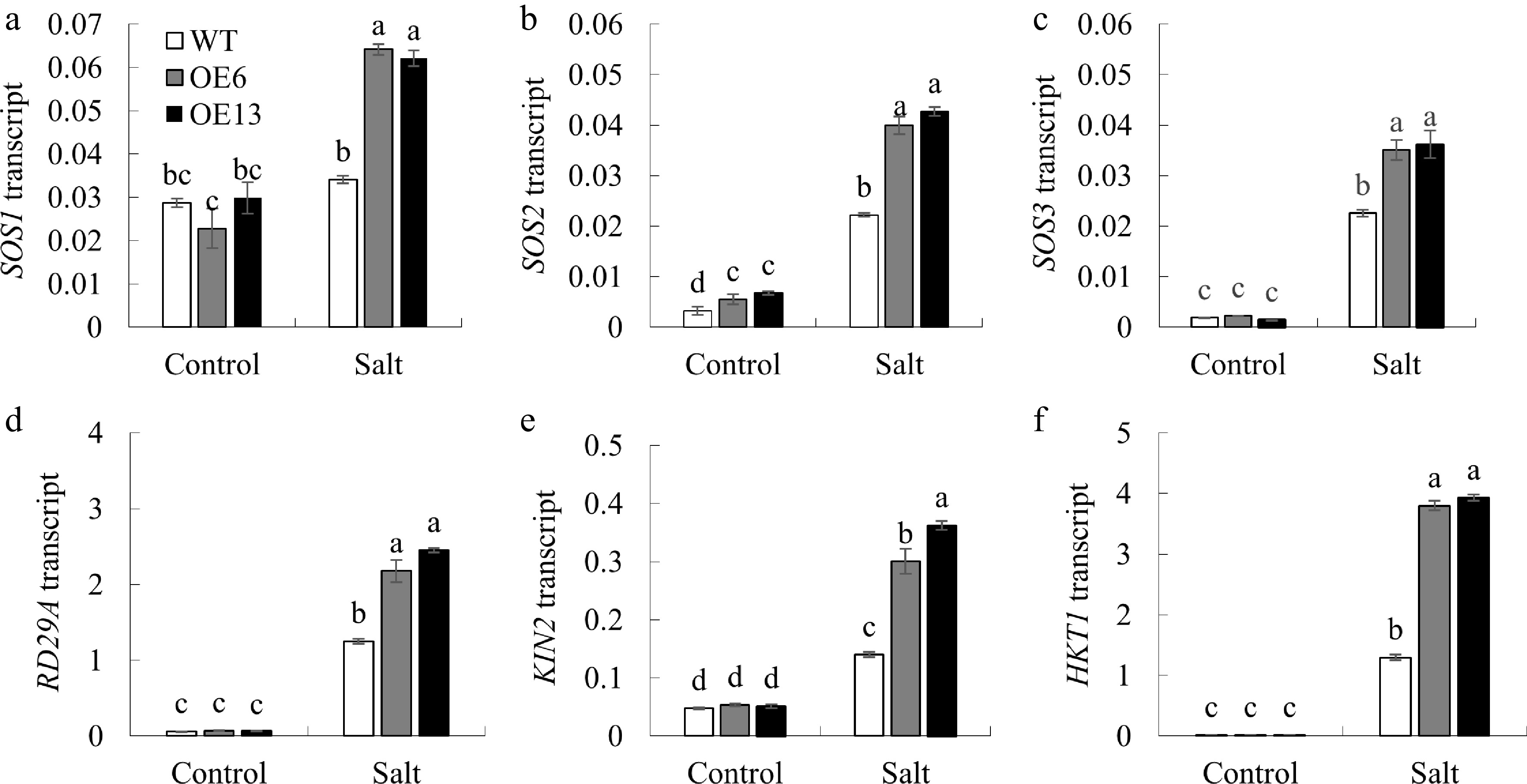
Figure 8.
The impact of salt stress on the expression of genes involved in ion homeostasis and stress response in Arabidopsis. After treating 7-day-old Arabidopsis seedlings with 125 mM NaCl for 6 h, the expression of (a) SOS1, (b) SOS2, (c) SOS3, (d) RD29A, (e) KIN2, and (f) HKT1 was assessed. Values are presented as mean ± SD (n = 3 replicates). Data are shown as the mean ± standard deviation (n = 3 replicates). Groups with the same letter above the bars are not significantly different at the p < 0.05 level.
-
Salinity stress is a major abiotic factor affecting plant growth and development. It primarily disrupts the dynamic balance of ions, osmosis, and ROS, severely reducing plant yield and quality[30,31]. The ascorbate-glutathione (AsA-GSH) cycle exists in various organisms. It plays a vital role in the antioxidant defense systems of plants and in their responses to diverse abiotic stresses. Within the AsA-GSH cycle, GSTs, proteins found extensively across diverse organisms, are crucially involved in detoxification mechanisms[32]. The tau class is one of the most important and widespread GST classes in plants[33]. Previous studies have shown that the tau class in Medicago trancatula consists of 52 members. A previous study reports that the expression levels of MtGSTU5, MtGSTU8, MtGSTU17, MtGSTU46, and MtGSTU47 increase significantly in response to various abiotic and biotic stress conditions. Among the MtGST family, MtGSTU17 stands out for its exceptional responsiveness to stress, particularly its strong affinity for traditional substrates, such as reduced GSH and 1-chloro-2,4-dinitrobenzene[23]. In the present study, we characterized a salt-inducible glutathione S-transferase gene, designated MsGSTU17, which showed a high degree of sequence similarity to the previously characterized MtGSTU17. The orthologous gene of MsGSTU17 in Arabidopsis thaliana, designated AtGSTU25 (At1g17180), exhibited 65% sequence similarity and was hypothesized to participate in the assimilation of sulfur and possess a spectrum of detoxification functions against environmental contaminants[34]. Analysis of the 2,000 bp MsGSTU17 promoter revealed the presence of abscisic acid-responsive elements (ABREs), MeJA-responsive elements, low-temperature response elements, and MYB-responsive elements. Among these, ABREs are primarily involved in regulating the ABA response or drought expression[35]. Based on the obtained data, we speculate that MsGSTU17 facilitates the conjugation of GSH and ROS in the cytoplasm, contributing to plant defense against abiotic stressors.
Understanding the subcellular localization of proteins is crucial for the functional analysis of genes. OsGSTU4 is detected in both the cytoplasm and nucleus, where it plays a role in enhancing the expression of genes related to stress responses and cellular detoxification[16]. PeGSTU58 is also observed in these two compartments, contributing to improved salt-alkaline tolerance and increased methionine content[36]. MdGSTU12 from Malus domestica is localized in the cytoplasm, functioning as a cytoplasmic transporter and promoting anthocyanin accumulation[37]. MsGSTU8 from Medicago sativa is found in the cytoplasm, where it is involved in salt stress responses[38]. Like MdGSTU12 and MsGSTU8, MsGSTU17 was cytoplasmic in location. However, the influence of different subcellular localizations on the functional roles of GSTs remains an area that requires further exploration.
Overexpression of the MsGSTU8 gene from alfalfa in transgenic tobacco enhances saline-alkali stress tolerance by reducing ROS accumulation and boosting antioxidant enzyme activity[38]. Similar to MsGSTU8, MsGSTU17 overexpression also significantly enhanced plant growth under salt stresses. This finding is also consistent with previous studies that demonstrated the key role of GSTUs in plant responses to environmental stimuli[5]. Under salt stress, the transcription of MsGSTU17 was upregulated in both leaf and root tissues. Compared with the WT, overexpression of MsGSTU17 resulted in increased shoot weight, root length, and survival rate after salt treatment, whereas ion leakage was reduced, indicating that MsGSTU17 positively regulates salt tolerance. Similarly, the soybean gene GmGSTU23 improves plant tolerance to salt stress by facilitating the removal of ROS and GSH, as evidenced by increased GST and GR activity and GSH content in transgenic lines exposed to salt stress[39]. Overexpression of GmGSTU4 in tobacco enhances salt stress tolerance by regulating metabolic shifts towards protective metabolites and detoxification pathways, including increased proline and trehalose, and activation of the oxidative pentose phosphate pathway[40].
The salt overly sensitive (SOS) signaling pathway plays a crucial role in regulating the balance of Na+ and K+ ions in plants, which is essential for their tolerance to salt stress[41]. SOS1 is a Na+/H+ exchanger, SOS3 acts as a Ca2+-binding protein, and SOS2, a serine/threonine protein kinase, plays a crucial role in the SOS pathway[42]. RD29A, KIN2, and HKT1 serve as stress- and ABA-responsive marker genes that contribute to salt tolerance in plants[43]. Among the absence of stress, the transcript levels of SOS1, SOS3, RD29A, KIN2, and HKT1 in the transgenic lines were comparable to those in WT plants, transcript level of SOS2 significantly increased. Under salt stress, the stress-responsive genes SOS1, SOS2, SOS3, RD29A, KIN2, and HKT1 were transcriptionally upregulated in both wild-type (WT) and transgenic plants. However, the expression levels in the transgenic plants were higher than those in the WT plants, which corresponded to the enhanced salt tolerance observed in the transgenic lines.
Exposure to abiotic stresses, such as salinity, leads to a substantial increase in ROS levels within plants, potentially inducing oxidative damage. The antioxidant defense system maintains a steady state between ROS production and scavenging, thereby preventing excessive ROS accumulation during stress[44]. When exposed to salt stress, significant accumulation of O2·− and H2O2 was observed in WT plants compared to that in the transgenic strains. This phenomenon was consistent with the increased enzyme activities of SOD, CAT, and APX detected in transgenic plants, indicating that their antioxidant responses were more effective. SOD is responsible for the detoxification of O2·−, whereas CAT and APX are involved in scavenging H2O2[45]. These findings imply that the enhanced salt tolerance in transgenic plants is linked to elevated antioxidant enzyme activities. Compared to WT plants, the transgenic lines exhibited significant increases in GST and GR activities, as well as a significant increase in the prototype GSH content. These data suggested that MsGSTU17 directly boosts GST activity and engages in ROS scavenging via GSH, thereby improving plant tolerance to salt stress. Nevertheless, there is limited knowledge regarding the interactions between MsGSTU17 and other transcription factors, necessitating further studies. Consequently, future investigations should employ techniques such as yeast two-hybrid screening and RNA sequencing to explore the role of MsGSTU17, to enhance our understanding of the mechanisms underlying salt tolerance in plants.
-
In this study, a novel MsGSTU17 gene was found to positively regulate salt tolerance. The elevated expression of MsGSTU17 improved salt tolerance in Arabidopsis and this overexpression induced alterations in the transcription levels of genes responsive to salt stress. The process mitigated the accumulation of reactive ROS by augmenting enzymatic antioxidant defenses, specifically by upregulating SOD, CAT, APX, GST, and GR catalytic activities, and by elevating intracellular GSH concentrations. Genes homologous to MsGSTU17 in crops may serve as potential targets for improving salt tolerance.
This work was funded by National Natural Science Foundation of China (32171687 and 32211530421), Personnel Startup Project of the Scientific Research and Development Foundation of Zhejiang A&F University (2023LFR039 and 2021FR041).
-
The authors confirm contribution to the paper as follows: study conception and design: Yu S, Zhang L; data collection: Yu S, Zhuang X; analysis and interpretation of results: Yu S, Chen S, Xie Y, Zhang L; draft manuscript preparation: Yu S, Zhuang X, Chen S, Xie Y, Zhang L. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available on request from the corresponding author.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Information on MsGST proteins.
- Supplementary Table S2 Primer sequences used in RT-qPCR.
- Supplementary Fig. S1 Bioinformatics analysis of MsGSTU17.
- Supplementary Fig. S2 The transmembrane domains and signal peptide analysis of MsGSTU17.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu S, Zhuang X, Chen S, Xie Y, Zhang L. 2025. A tau class glutathione S-transferase in alfalfa, MsGSTU17, regulates salt tolerance in transgenic Arabidopsis thaliana. Grass Research 5: e005 doi: 10.48130/grares-0025-0005 

A tau class glutathione S-transferase in alfalfa, MsGSTU17, regulates salt tolerance in transgenic Arabidopsis thaliana
- Received: 03 December 2024
- Revised: 06 January 2025
- Accepted: 21 January 2025
- Published online: 21 February 2025
Abstract: High soil salinity negatively affects crop yield, intrinsic quality, and economic profitability. The tau class of glutathione S-transferases (GSTs) is pivotal in plant defense mechanisms against salt stress. In this study, we characterized a salt-responsive tau-class GST gene, designated as MsGSTU17, in Medicago sativa. The MsGSTU17gene was found to be located in the cytoplasm and had high expression levels in both leaves and roots. Compared to the wild-type plants, overexpression of MsGSTU17 in Arabidopsis under salt stress led to increased root length, shoot weight, and survival rate, along with a reduced ion leakage rate. At the same time, the transgenic Arabidopsis lines showed significantly increased expression of genes related to salt stress, including SOS1, SOS2, SOS3, KIN2, RD29A, and HKT1, compared to the wild-type. Additionally, these transgenic plants exhibited an enhanced antioxidant defense system characterized by elevated activities of catalase (CAT), superoxide dismutase (SOD), ascorbate peroxidase (APX), GST, and glutathione reductase (GR), and higher levels of glutathione (GSH). In summary, our results suggest that overexpressing MsGSTU17 in transgenic plants can mitigate salt-caused damage by scavenging reactive oxygen species and increasing GSH levels via upregulation of GST activity.
-
Key words:
- Antioxidant system /
- Glutathione S-transferases /
- Medicago sativa L. /
- Salt tolerance