









-
β-caryophyllene is a sesquiterpene commonly found in plants with a variety of pharmacological and biological functions. β-caryophyllene has anti-inflammatory and antifungal properties[1,2]. Simultaneously, β-caryophyllene is capable of mediating signaling between different organisms and plays an important role in plant-induced defense responses. Flowers can attract insect pollination by releasing β-caryophyllene in Arabidopsis[3]. β-caryophyllene released from maize roots attracts natural enemies of pests[4]. Two terpene synthesis pathways exist in plants. The mevalonic acid (MVA) pathway located in the cytoplasm and the methyl erythritol-4-phosphate (MEP) pathway located in the plastids. Among them, β-caryophyllene is generated via the MVA pathway and finally catalyzed by the terpene synthase (TPS)[5].
The JASMONATE ZIM DOMAIN (JAZ) proteins originally known as ZIM (zinc finger proteins expressed in inflorescence meristem), work as inhibitors of the JA signaling pathway[6]. JAZ subfamily generally contains two highly conserved domains: the ZIM domain (also called TIFY), and the Jas domain (also called CCT_2)[7], which are essential for JAZ to interact with other proteins[8,9]. Under normal growth conditions, the JA-Ile level in plants is low. Then JAZ proteins bind to MYC2 and other transcription factors to inhibit the activity of transcription factors (TFs), thereby suppressing JA signaling[10]. However, under stress conditions, JA signaling molecules are synthesized in plants to form JA-Ile or other amino acid derivatives, prompting the COI1 protein of the SCFCOI1 complex to interact with JAZ proteins[8]. Subsequently, the JAZ proteins are ubiquitinated and degraded via the 26S proteasome pathway and TFs like MYC2 are released to activate the expression of downstream JA response genes[11]. Research found that when JA response occurred, JAZ genes would be significantly induced, which in turn inhibited the activity of MYC2, forming a feedback regulation mechanism, so that the plants would not produce too strong JA response, and avoiding excessive energy consumption within the plants[12,13]. Studies have shown that JAZ is involved in the regulation of plant terpenoid metabolism. In Artemisia annua, the AaJAZ8 protein inhibited the artemisinin biosynthesis by repressing the expression of artemisinin synthesis gene[14]. In Salvia miltiorrhiza, SmJAZ3 could induce the accumulation of salicylic acid and tanshinone and promote plant growth and development by interacting with SmWD40-170[15]. In Curcuma wenyujin, CwJAZ4/9 proteins inhibited the terpene pathway and jasmonic acid response by forming the CwJAZ4/9 - CwMYC2 regulatory cascade, thereby reducing terpene accumulation[16]. Although JAZs play an important regulatory role in plant growth and development, there is currently little known about the regulatory factors upstream of JAZ.
Non-heading Chinese cabbage (NHCC) (Brassica campestris (syn. Brassica rapa) ssp. chinensis) is a common leafy vegetable that originated in China and has gradually been cultivated worldwide. 'XQC', one cultivar of NHCC, is named for its unique volatile flavor. β-caryophyllene is one of its volatile components in NHCC. In our previous studies, BcTPS21 was considered a key gene in NHCC involved in β-caryophyllene biosynthesis. However, there are few knowledge on the regulation of β-caryophyllene synthesis in NHCC.
In this study, the JAZ gene family in NHCC have been characterized. The interaction between BcJAZ2 and BcMYC2 has been confirmed and the role of BcJAZ2 in regulating β-caryophyllene synthesis in NHCC was verified through heterologous expression and gene silencing. Additionally, we found that BcbHLH137, BcHBI1.1, and BcHBI1.2 could bind the BcJAZ2 promoter and activate its expression. These results provide a basis for further exploration of β-caryophyllene biosynthesis.
-
The JAZ gene sequences of NHCC were retrieved from the NHCC database[17] (
http://tbir.njau.edu.cn/NhCCDbHubs/ ) and TBGR database[18] (www.tbgr.org.cn ) and were screened using HMM search and two rounds of BLASTP in TBtools v 2.096[19]. Hidden Markov models (HMMs) of TIFY domain (PF06200) and Jas domain (PF09425) were downloaded from the Pfam database (http://pfam-legacy.xfam.org/ ). Finally, the screened candidate genes were validated through the NCBI Batch CD-Search (www.ncbi.nlm.nih.gov/ ) and only selected genes with TIFY and Jas domains. Their physicochemical properties were calculated by the ExPASy (https://web.expasy.org/compute_pi/ ) and TBtools, respectively. The prediction of subcellular localizations of BcJAZs was conducted using WOLF PSORT website (https://wolfpsort.hgc.jp/ ).The conserved motifs of the BcJAZs were predicted using the MEME online website[20] (
https://meme-suite.org/meme/ ) and visualized by TBtools. The Jalview software was performed for multiple sequence alignment of BcJAZ proteins[21]. Based on the NHCC genome GFF file, the chromosomal mapping of BcJAZ genes was visualized using TBtools software. Utilizing MCScanX in TBtools, we identified intraspecific gene duplication events and interspecific collinearity relationships of BcJAZ family members. Additionally, the cis-elements of BcJAZ genes upstream 2,000 bp promoter sequences were predicted via the PlantCARE database[22].Phylogenetic analysis of BcJAZ gene family
-
The gene sequences of Arabidopsis and rice that have been reported were accessed via the TAIR database (
www.arabidopsis.org ) and Rice Genome Database (http://rice.plantbiology.msu.edu ). We utilized the MEGA 11.0 software to align the JAZ protein sequences from Arabidopsis, NHCC, and rice, and constructed a phylogenetic tree using the neighbor-joining (NJ) method with 1,000 bootstrap replicates to study the evolution of the BcJAZ gene family in NHCC[23].Plant materials and treatments conditions
-
The 'XQC' variety of NHCC, Arabidopsis thaliana and Nicotiana benthamiana mentioned in this experiment were acquired form the Nanjing Agricultural University (Nanjing, China). All plant materials were cultivated in light 16 h/22 °C and dark 8 h/18 °C. Various treatments were carried out with three biological replicates when the seedlings were about one-month-old. The low-temperature treatment conditions were 4 °C for both daytime and nighttime and the hormone treatments were spraying the leaves with 100 μM methyl jasmonate (MeJA) and 100 μM gibberellic acid (GA). Samples without MeJA and GA treatments (taken at 0 h) as control. Plant leaves were taken at 0, 1, 3, 6, 12, 24, and 48 h after treatments. The BcJAZ2 transgenic lines were created by dipping flowers using Arabidopsis thaliana Columbia-0 (Col-0) wild type.
qRT-PCR analysis
-
Employing the RNA simple Total RNA Kit (Tiangen, Beijing, China), we extracted total RNA of NHCC and Arabidopsis and using HiScript III RT SuperMix for qPCR (+gDNA wiper) (Vazyme, China) synthesized cDNA. qPCR was carried out with ChamQ SYBR qPCR Master Mix (Vazyme, China) and the relative expression of genes was calculated using 2−ΔΔCᴛ method. The reference genes were BcGAPDH and AtACTIN for NHCC and Arabidopsis[24]. All primers used in this study are supplied in Supplementary Table S1.
Plasmid construction
-
For subcellular localization, BcJAZ2 was inserted into the PRI101 vector. The 35S:BcJAZ2-GFP overexpression vector was transformed into Agrobacterium tumefaciens strain GV3101 (pSoup) (TOLOBIO, China).
For Y1H and Y2H assays, the BcJAZ2 promoter segments pAbAi-proBcJAZ2-P1 (−1,250 to 0 bp) were integrated into the pAbAi vector. To verify trans-activation activity of transcription factors, the bait proteins BcbHLH137, BcHBI1.1, BcHBI1.2 and BcJAZ2 were recombined into pGBKT7(BD) vectors, while the prey protein BcMYC2 to be validated were integrated into the pGADT7 (AD) vector.
For BiFC assays, the open reading frames (ORF) of BcJAZ2 and BcMYC2 were cloned and constructed into BiFC-YC and BiFC-YN vectors, respectively. The created recombinant vectors (YC-BcJAZ2 and YN-BcMYC2) were introduced into Agrobacterium tumefaciens strain GV3101 (pSoup) (TOLOBIO, China).
For LUC assays, the CDS without stop codon of BcHBI1.1, BcHBI1.2, and BcbHLH137 were integrated into the PRI101 vector to create effector vectors. BcJAZ2 truncated promoter fragment proBcJAZ2-P1 (−1,250 to 0 bp) was constructed into pGreenII-0800-LUC to create a reporter vector. The recombinant plasmids were introduced into Agrobacterium tumefaciens strain GV3101 (pSoup) (TOLOBIO, China). All primers used in this study are supplied in Supplementary Table S1.
Subcellular localization
-
Agrobacterium tumefaciens strain carrying the GFP vector was mixed with nuclear marker in a 1:1 ratio, and injected into tobacco leaves. The tobacco was cultivated in darkness for 24 h followed by normal cultivation for 24−48 h. Through the confocal laser scanning microscopy (Zeiss, LSM 780, Germany), we detected the position of GFP signals.
Bimolecular fluorescence complementation assays
-
Agrobacterium tumefaciens strains containing the BiFC-YC and BiFC-YN plasmids were blended in the same ratio and injected into tobacco leaves. Tobacco treatment conditions were the same as for subcellular localization. Using the confocal laser scanning microscope (Zeiss, LSM 780, Germany), distinct fluorescence signals were discovered after 48−72 h.
Virus-induced BcJAZ2 gene silencing in NHCC
-
Virus-induced gene silencing (VIGS) is a rapid method used to reveal gene function[25]. Based on the CDS sequence and conserved domains of BcJAZ2, a 40 bp specific fragment was designed, which was then reverse-complemented and ligated to the previous 40 bp fragment to form an 80 bp palindromic hairpin structure. The DNA sequence is shown in Supplementary Table S1. Subsequently, the sequences were constructed into PTY-empty vector, and then PTY-empty and PTY-BcJAZ2 plasmids were transformed into the NHCC variety 'XQC' by the method of gene gun bombardment (Bio-Rad, PDS1000/He), respectively. The phenotypes of the silenced plants were observed after two weeks. Extracted RNA and detected the expression level of BcJAZ2 by qRT-PCR.
Yeast one-hybrid library screening and yeast two-hybrid analysis
-
Using the lithium acetate procedure (Takara, Japan), the linearized bait vector pAbAi-proBcJAZ2-P1 (−1,250 to 0 bp) was introduced into yeast Y1H Gold strain, which was grown on SD/-Ura deficient medium containing different doses of Aureobasidin A (AbA) for 2−3 d to find the lowest AbA concentration that repressed self-activation (Supplementary Fig. S1). The One Step Yeast Direct PCR kit (TOLOBIO, China) was used to identify the positive recombinant clones. A cDNA library of 10 μL was added to the 600 μL Y1H Gold cells containing pAbAi bait and was grown on 200 ng/mL AbA SD/-Leu solid medium at 28 °C for 5−7 d. The colonies grown on solid medium were detected by PCR and the sequencing results were compared using the NCBI website (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ). The prey vectors pGADT7-BcbHLH137, pGADT7-BcHBI1.1, pGADT7-BcHBI1.2 and empty pGADT7 were then transferred into yeast strains containing the bait vector, respectively, and cultured for 3−5 d at 28 °C in SD/-Leu deficient medium containing AbA.For Y2H assays, the bait protein BcJAZ2-BD and prey protein BcMYC2-AD were co-transformed into yeast Y2H Gold strain by the lithium acetate method (Takara, Japan). Similarly, the bait proteins BcbHLH137-BD, BcHBI1.1-AD, and BcHBI1.2-AD were co-transformed into yeast Y2H Gold strain with AD empty vector, respectively. The co-transformed yeast cells were first cultured on the SD/-Leu-Trp medium and then moved to the SD/-Leu-Trp-Ade-His medium to grow for 3−5 d at 28 °C.
Dual-luciferase reporter assay
-
Agrobacterium tumefaciens strains containing effector and LUC reporter were blended in the same ratio and injected into tobacco leaves, separately. Tobacco treatment conditions were the same as for subcellular localization. After 48−72 h, the luciferase activity was detected using Dual Luciferase Reporter Assay Kit (Vazyme, China).
GC-MS analysis
-
Five-week-old Arabidopsis inflorescences (0.2~0.5 g) and leaves of BcJAZ2 silenced plants were collected for β-caryophyllene determination. To repress the enzyme reaction, the plant samples were ground into powder and 2 mL NaCl saturated solution was added together and poured into headspace vials (Agilent, CA, USA). A gas chromatograph (TRACE 1310, Thermo Scientific) and a triple quadrupole mass spectrometer were employed for β-caryophyllene substance detection. The method of testing was consistent as described by Wang et al.[26].
Statistical analysis
-
The experiments in this study were set up with three biological and technical replicates and the data in this study were analyzed by Student's t-tests (*p ≤0.05, **p ≤ 0. 01, ***p ≤ 0. 001, ****p ≤ 0.0001). Error lines represent standard deviations. Statistical significance is indicated by different asterisks.
-
We used bioinformatics techniques to identify 25 JAZ family members in high quality NHCC genomes which were distributed on nine chromosomes (Fig. 1a). The gene sequence lengths of BcJAZs ranged from 393 bp (BcJAZ19) to 1,062 bp (BcJAZ2), and the encoded proteins varied from 130 aa (BcJAZ19) to 353 aa (BcJAZ2). Their theoretical isoelectric points and molecular masses varied from 5.10 (BcJAZ6) to 9.99 (BcJAZ7) and 14.97 to 37.58 KDa, respectively. Subcellular localization predictions indicated most BcJAZs showed nuclear distribution (Supplementary Table S2).
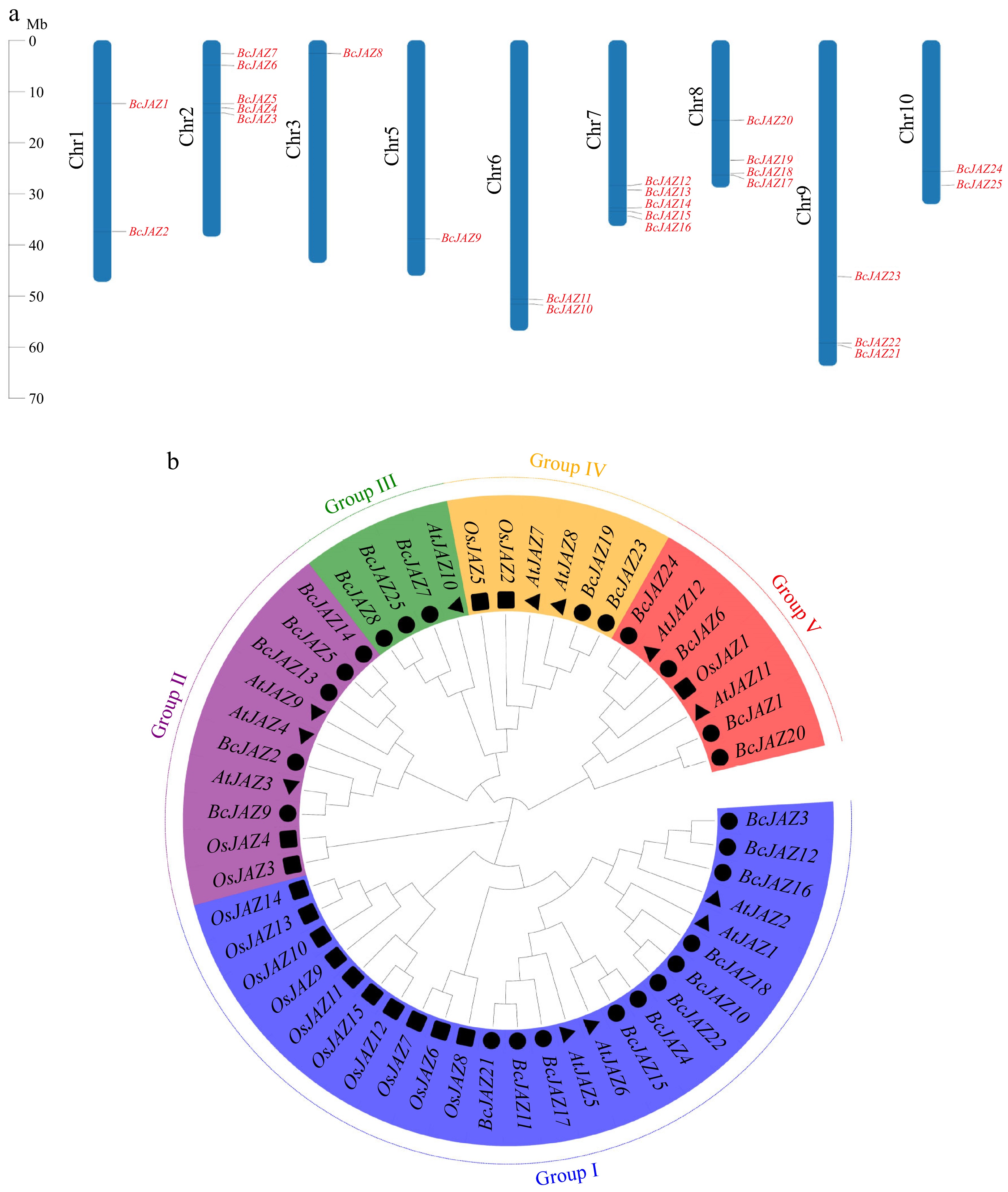
Figure 1.
Chromosomal mapping and phylogenetic tree of BcJAZ gene family. (a) Chromosomal mapping of BcJAZ family members. (b) Phylogenetic tree of NHCC, Arabidopsis, and rice. Subfamilies I, II, III, IV, and V were indicated by different colors. The black circles represent the 25 BcJAZs in NHCC. The black triangles represent the 12 AtJAZs in Arabidopsis. The black squares represent the 15 OsJAZs in rice.
Aiming to explore the phylogenetic relationship of JAZ among NHCC, Arabidopsis, and rice, we established a phylogenetic tree containing 25 BcJAZs, 12 AtJAZs, and 15 OsJAZs proteins (Fig. 1b). According to the phylogenetic tree, we found that all JAZs were mainly clustered in five groups according to the evolutionary distance. Group I contained 11 BcJAZ members which contained most of the BcJAZ genes of NHCC. Interestingly, most of the OsJAZ genes (10) were also distributed in group I. Group II was comprised of five BcJAZ genes, BcJAZ2, BcJAZ5, BcJAZ9, BcJAZ13, and BcJAZ14. Group III was comprised of three BcJAZ genes, BcJAZ7, BcJAZ8, and BcJAZ25. Group IV was comprised of two BcJAZ genes, BcJAZ19, and BcJAZ23. Group V was comprised of four BcJAZ genes, BcJAZ1, BcJAZ6, BcJAZ20, and BcJAZ24.
Multiple sequence alignment, conserved domain, gene structure of the BcJAZ genes
-
Multiple sequence alignment results showed that BcJAZ proteins had two very obvious conserved domains and were identified: motif 1 corresponded to the Jas domain, while motif 2 corresponded to the TIFY domain (Fig. 2a, b). As shown in Fig. 2c, the majority of the BcJAZ genes sharing similar motifs grouped in closer evolutionary branch. The gene structure and predicted motifs of BcJAZs were visualized using TBtools. The findings indicated that the BcJAZ gene family showed variations in CDS quantities (Fig. 2d).

Figure 2.
Multiple sequence alignment, motifs and gene structure of BcJAZ genes. (a), (b) Multiple sequence alignment and conserved domains of BcJAZ. (c) Conserved motifs of BcJAZs. (d) The gene structure of BcJAZs; CDS are represented by yellow boxes, introns by grey lines, and UTRs by green boxes.
Collinearity analysis of the BcJAZ genes
-
Intraspecific collinearity analysis of the BcJAZ genes revealed 31 gene pairs underwent segmental duplication events (Fig. 3a, Supplementary Table S3), and no tandem duplication clusters were detected, indicating the main gene duplication event for JAZ genes in NHCC was segmental duplication. Interspecies collinearity analysis showed that there are 36 collinear gene pairs between NHCC and Arabidopsis, and two collinear gene pairs between NHCC and rice (Fig. 3b). The collinearity analysis indicated that the BcJAZs of NHCC had closer relationship with the AtJAZs of Arabidopsis, which was consistent with the present cognition of plant evolutionary history.
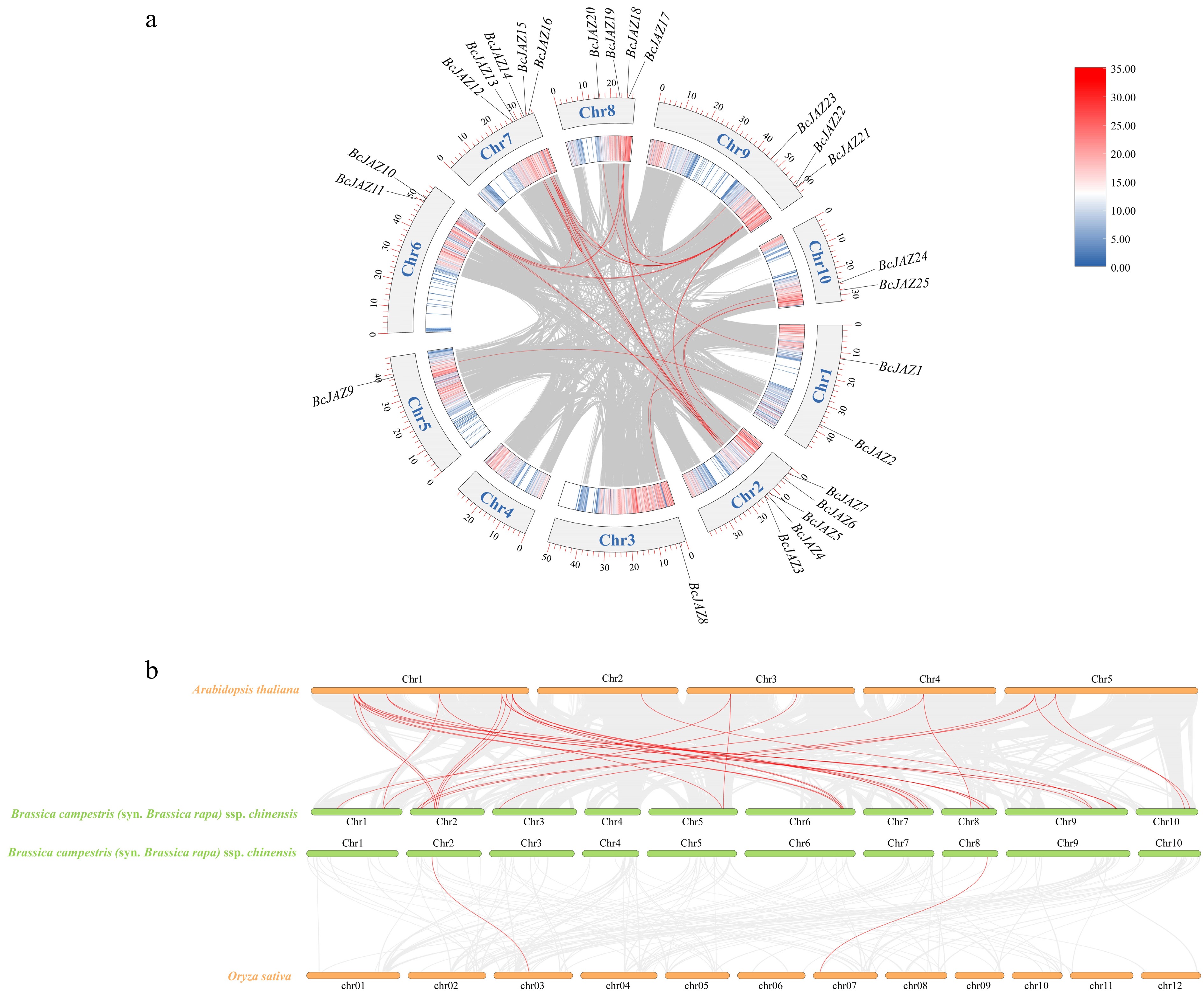
Figure 3.
Collinearity analysis of the BcJAZs. (a) Intraspecies collinearity analysis of BcJAZs. (b) Interspecific collinearity analysis of BcJAZs in NHCC, Arabidopsis thaliana, and Oryza sativa.
Cis-elements of BcJAZ gene family
-
Gene function can be predicted by analyzing the promoter cis-elements. Based on the predictions of the Plantcare website, we have cis-elements categorized into five groups: core/binding, development, abiotic/biotic elements, light, and hormones (Fig. 4). All 25 BcJAZs promoters contained numerous core elements and binding sites, including CAAT-box and TATA-box. Most BcJAZs contained certain abiotic-responsive and light-responsive elements, including the drought-responsive element like DRE core, the low temperature-responsive element like LTR, as well as the light-responsive elements BOX4, GT1-motif, and G-box. Some hormone response elements, including SA (TCA-element), IAA (TGA-element), GA (GARE-motif, P-box), MeJA (CGTCA-motif, TGACG-motif), and ABA (ABRE-element), were also detected in most of the BcJAZs promoters. These results indicated that BcJAZs likely serve in development, abiotic stress, and multiple phytohormone signaling pathways.
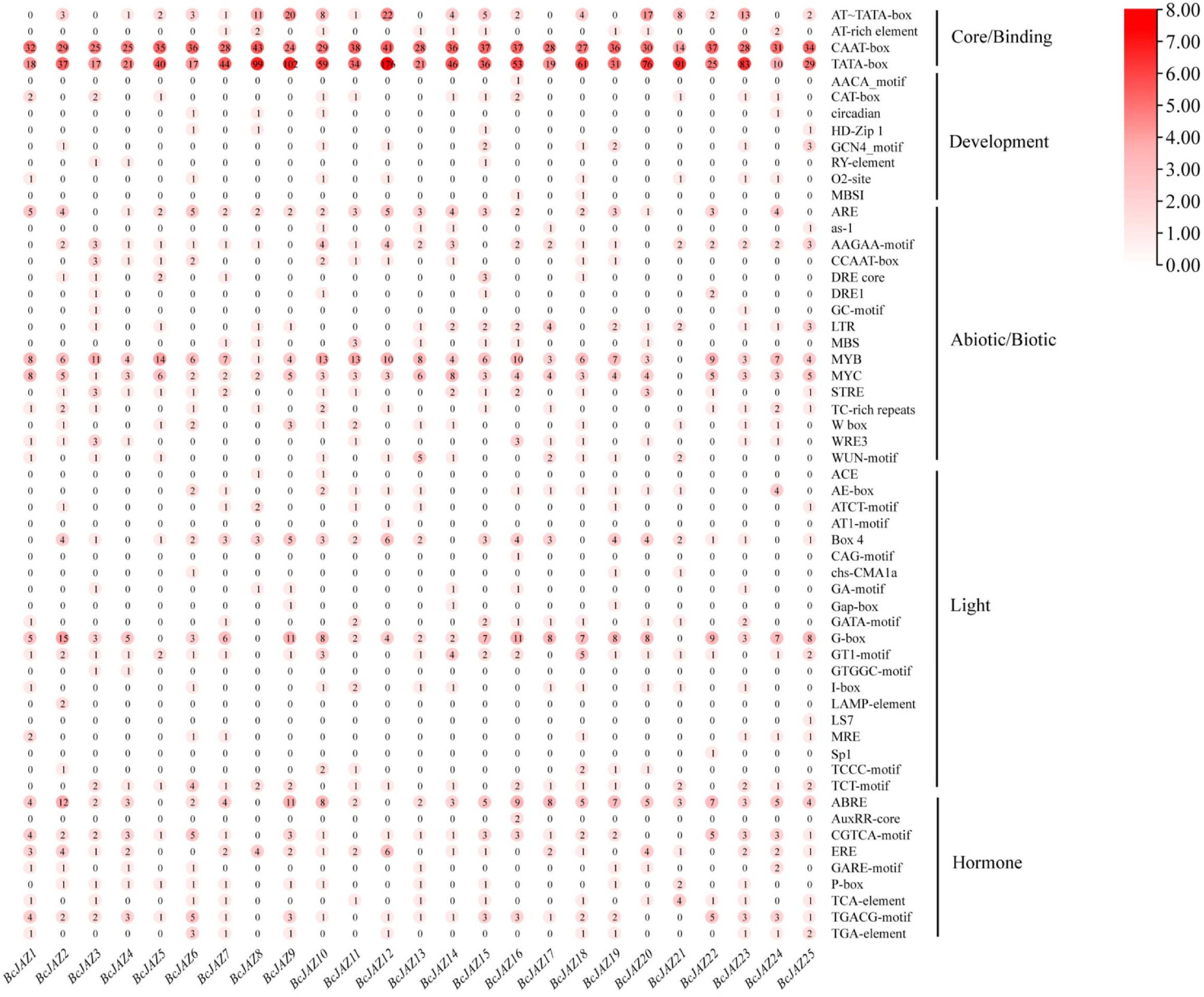
Figure 4.
Cis-elements analysis of BcJAZs promoters in NHCC. The numbers within the red circles indicated the quantities of cis-elements.
Expression patterns of BcJAZs in response to abiotic stress and hormone treatments
-
RNA-Seq analysis showed that under low- and high-temperature stress, ten BcJAZ genes (BcJAZ2, BcJAZ3, BcJAZ4, BcJAZ5, BcJAZ9, BcJAZ12, BcJAZ14, BcJAZ16, BcJAZ21, and BcJAZ24) were strongly induced (Fig. 5a). Previous research has demonstrated that BcJAZs participated in hormone responses, and their expression patterns under different treatments may reflect genes biology function. qRT-PCR data indicated that all genes responded to MeJA treatment (Fig. 5c), among which BcJAZ2, BcJAZ4, BcJAZ5, BcJAZ9, BcJAZ14, BcJAZ16, and BcJAZ24 showed 'up-down' expression trends after treatment. Under GA treatment (Fig. 5d), the expression levels of BcJAZ2, BcJAZ3, BcJAZ5, BcJAZ14, BcJAZ16, and BcJAZ21 showed 'up-down' expression trends after treatment. Although BcJAZ9 and BcJAZ12 showed similar expression patterns, the expression of BcJAZ9 reached its peak at 1 h after GA treatment, while the expression level of BcJAZ12 was highest at 3 h. Conversely, the expression levels of BcJAZ16 and BcJAZ24 continuously decreased until 3 and 12 h after treatment, respectively.
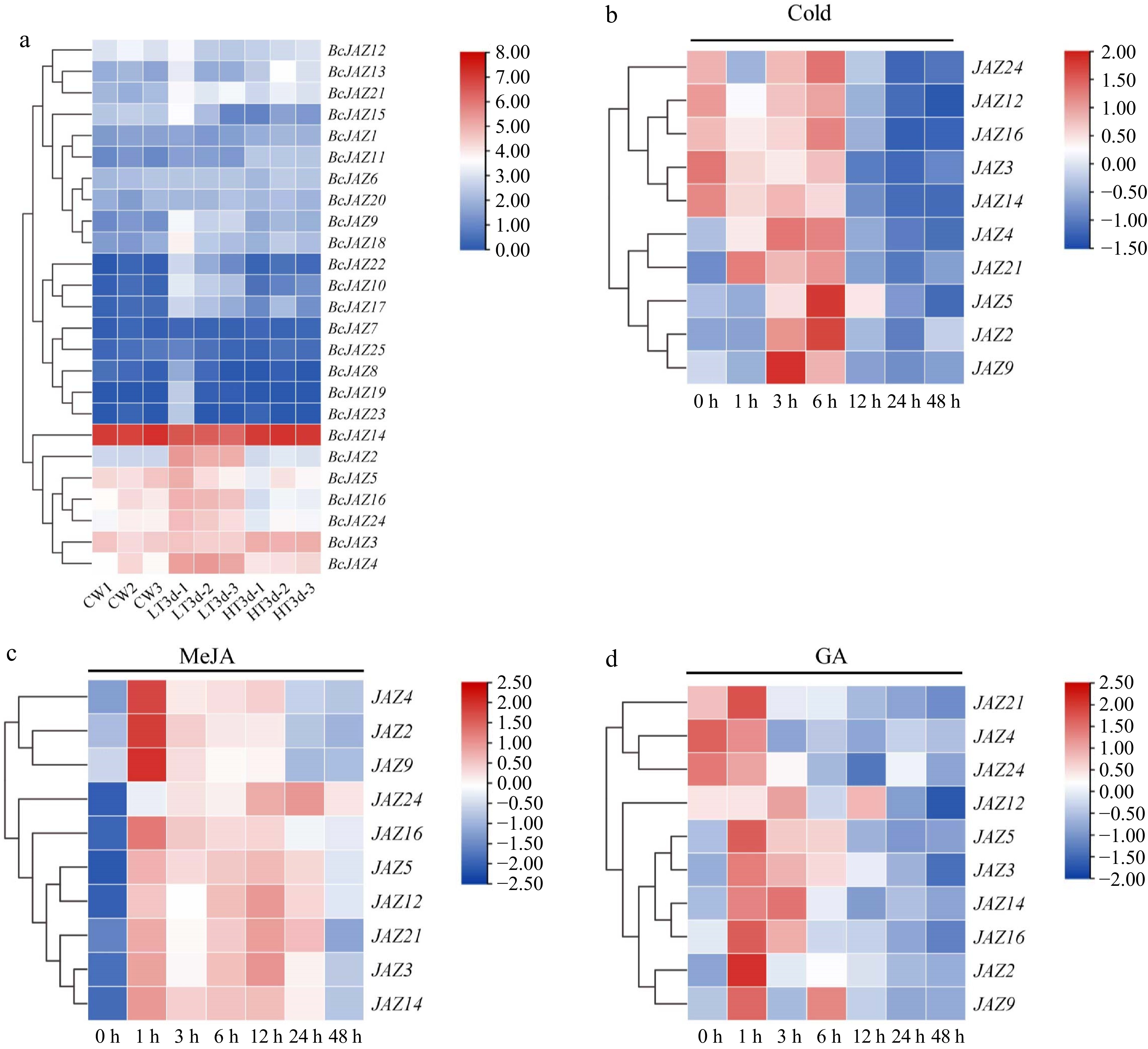
Figure 5.
Expression patterns of BcJAZ genes in response to abiotic stresses and hormone treatments. (a) RNA-Seq data of the BcJAZ gene family at different temperatures (CW represented normal temperature, LT3d represented low temperature 8/4 °C and HT3d represented high temperature 38/25 °C). The bar graph shows the Log2 (TPM + 1) values, indicating expression levels from low to high. (b) The expression patterns of BcJAZs under 4 °C, (c) MeJA, and (d) GA treatment. After being normalized, the expression levels were visualized using TBtools.
In particular, the expression levels of BcJAZ2 changed most significantly under long-term low-temperature treatment (Fig. 5a). Meanwhile, we analyzed the expression levels of JAZs under short-term (48 h) low-temperature treatment and the expression level of BcJAZ2 was significantly up-regulated after 6 h of treatment (Fig. 5b). Previous studies showed that low temperatures inhibited β-caryophyllene synthesis in NHCC (unpublished data), and BcJAZ2 was significantly induced by both short- and long-term low temperature treatments. Therefore, we hypothesized that BcJAZ2 may be a key negative regulator of β-caryophyllene synthesis.
BcJAZ2 interacts with BcMYC2
-
The results of website prediction and subcellular localization were consistent and both revealed that BcJAZ2 was located in the nucleus (Fig. 6a). According to predictions of the STRING 12.0 database, BcJAZ2 appeared to be functionally connected with proteins including MYC2/MYC3/MYC4, COI1, Ninja family proteins, and JAZ family proteins (Fig. 6b). It has demonstrated that MYC2 in Arabidopsis can directly bind to the promoter of the β-caryophyllene synthase gene TPS21 and activate its expression[27]. We verified the relationship between BcMYC2 and BcJAZ2 of NCHH using Y2H (Fig. 6c). Each combination could grow normally on SD/-Trp-Leu mediums, whereas only yeast strains transformed with BcJAZ2-BD and BcMYC2-AD plasmids and a positive control could grow normally on SD/-Trp-Leu-His-Ade medium, suggesting that BcJAZ2 could interact with BcMYC2 in yeast. To further validate the interactions between BcJAZ2 and BcMYC2, it was verified by BiFC experiments. The results showed that when BcJAZ2-cYFP and BcMYC2-nYFP were co-injected, yellow fluorescence could be detected, indicating that BcJAZ2 and BcMYC2 could interact in plants (Fig. 6d). These results suggested that BcJAZ2 may repress the expression of the β-caryophyllene synthase gene through BcMYC2.
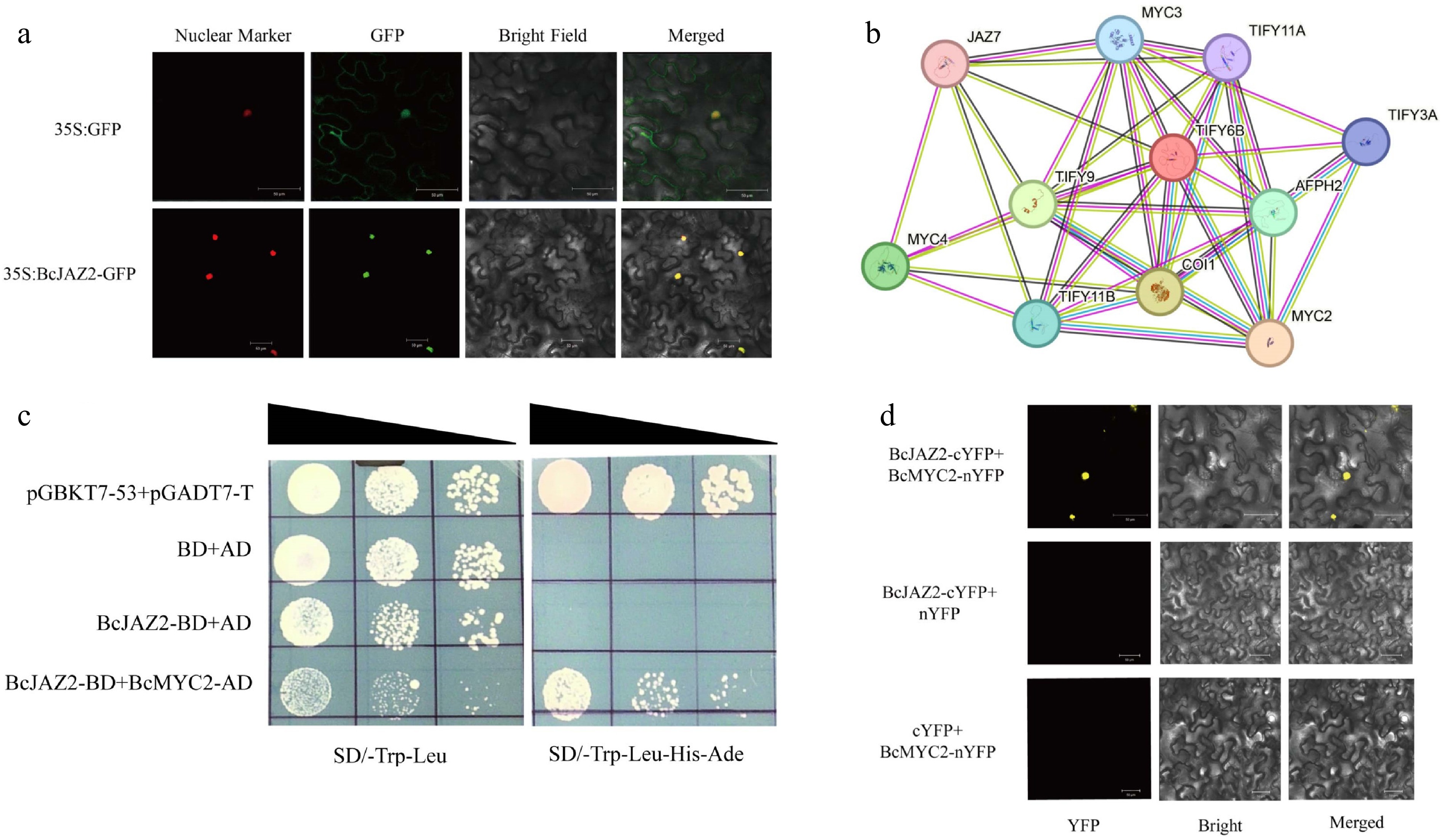
Figure 6.
BcJAZ2 interacts with BcMYC2. (a) Subcellular localization of BcJAZ2. The scale bar was 50 μm. (b) The protein-protein interaction (PPI) network of BcJAZ2 (TIFY6B) from the STRING 12.0 database. (c) Yeast two-hybrid assays. Positive control was yeast co-transformed with pGBKT7-53 and pGADT7-T and yeast co-transformed with pGBKT7 and pGADT7 was negative control. (d) Bimolecular fluorescence complementation assay. The scale bar was 50 μm.
Overexpression of BcJAZ2 negatively regulates β-caryophyllene biosynthesis in Arabidopsis
-
To further explore the function of BcJAZ2, we constructed and identified BcJAZ2 transgenic lines in the background of the wild-type (WT) model plant Arabidopsis thaliana (Fig. 7a, b). The formation of almost all floral volatile sesquiterpenes in Arabidopsis is attributed to the sesquiterpene synthase genes TPS21 and TPS11, with TPS21 being accountable for producing of β-caryophyllene[27]. In inflorescences of BcJAZ2 transgenic lines, the expression of AtTPS21 and AtTPS11 was significantly repressed compared with WT (Fig. 7c). To determine whether BcJAZ2 influences the JA signaling, we analyzed the expression levels of JA signaling genes by qRT-PCR. As expected, AtCOI1 and AtMYC2 expression levels were suppressed (Fig. 7c). The effects of transcriptional regulation of BcJAZ2 on β-caryophyllene synthesis in overexpression lines were also further monitored by GC-MS (Fig. 7d). Compared with WT, the β-caryophyllene content of BcJAZ2 transgenic lines Arabidopsis decreased approximately 45%. Together, these data suggested that BcJAZ2 negatively regulated the synthesis of β-caryophyllene, by inhibiting JA signaling and further suppressing the expression of the sesquiterpene synthase.
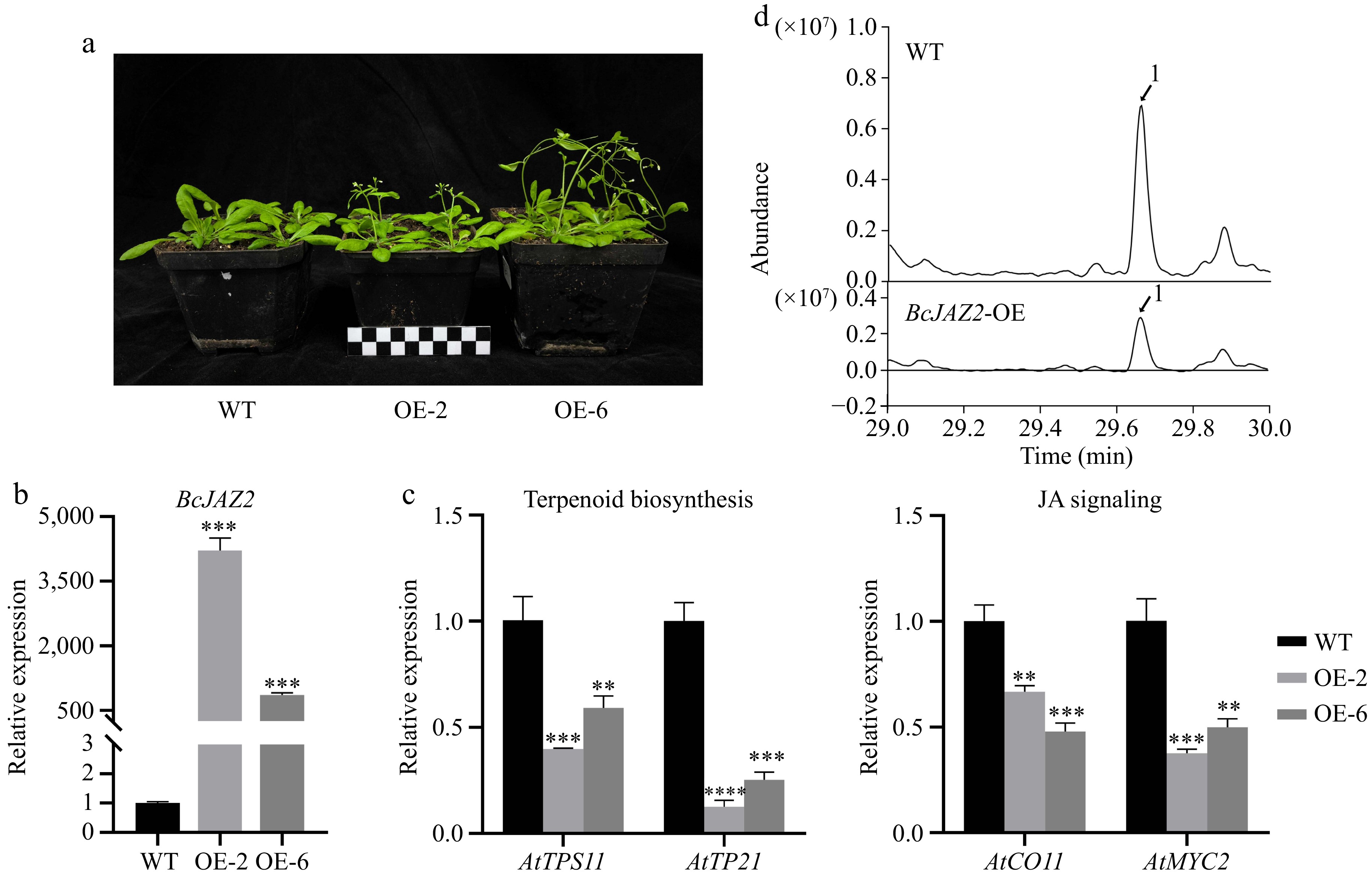
Figure 7.
Overexpression of BcJAZ2 inhibits β-caryophyllene biosynthesis in Arabidopsis. (a) The phenotypes of BcJAZ2 transgenic Arabidopsis. The black and white boxes are scale bar and one box represents 1 cm. (b) Relative expression of BcJAZ2 in Arabidopsis transgenic lines inflorescences. (c) Analysis of sesquiterpene synthase and JA signaling genes expression level in inflorescences of BcJAZ2 transgenic Arabidopsis. Student's t-test: **p ≤ 0. 01, ***p ≤ 0.001, ****p ≤ 0.0001. Three experiments were repeated and values were means ± SD. (d) GC-MS chromatogram analysis of β-caryophyllene emitted by Arabidopsis inflorescences in both wild-type (WT) and transgenic lines. Peaks with number represented β-caryophyllene.
Silencing BcJAZ2 by VIGS promotes sesquiterpene biosynthesis in NHCC
-
To elucidate the biological function of BcJAZ2 in NHCC, we obtain PTY-BcJAZ2 silenced plants using the virus-induced gene silencing approach (Fig. 8a). The expression of BcJAZ2 was significantly reduced in the silenced plants (PTY-BcJAZ2 #11, #13) compared to the control (PTY), confirming that BcJAZ2 was successfully silenced (Fig. 8b). qRT-PCR data suggested that the expression levels of β-caryophyllene synthase gene BcTPS21 and key genes for JA signaling in silenced plants were significantly promoted (Fig. 8c). Additionally, further monitoring of the impact of BcJAZ2 silencing on the β-caryophyllene biosynthesis was conducted by GC-MS (Fig. 8d). Consistent with the gene expression patterns, an approximately 3-fold increase in β-caryophyllene content was measured in PTY-BcJAZ2 plants compared to the PTY control.
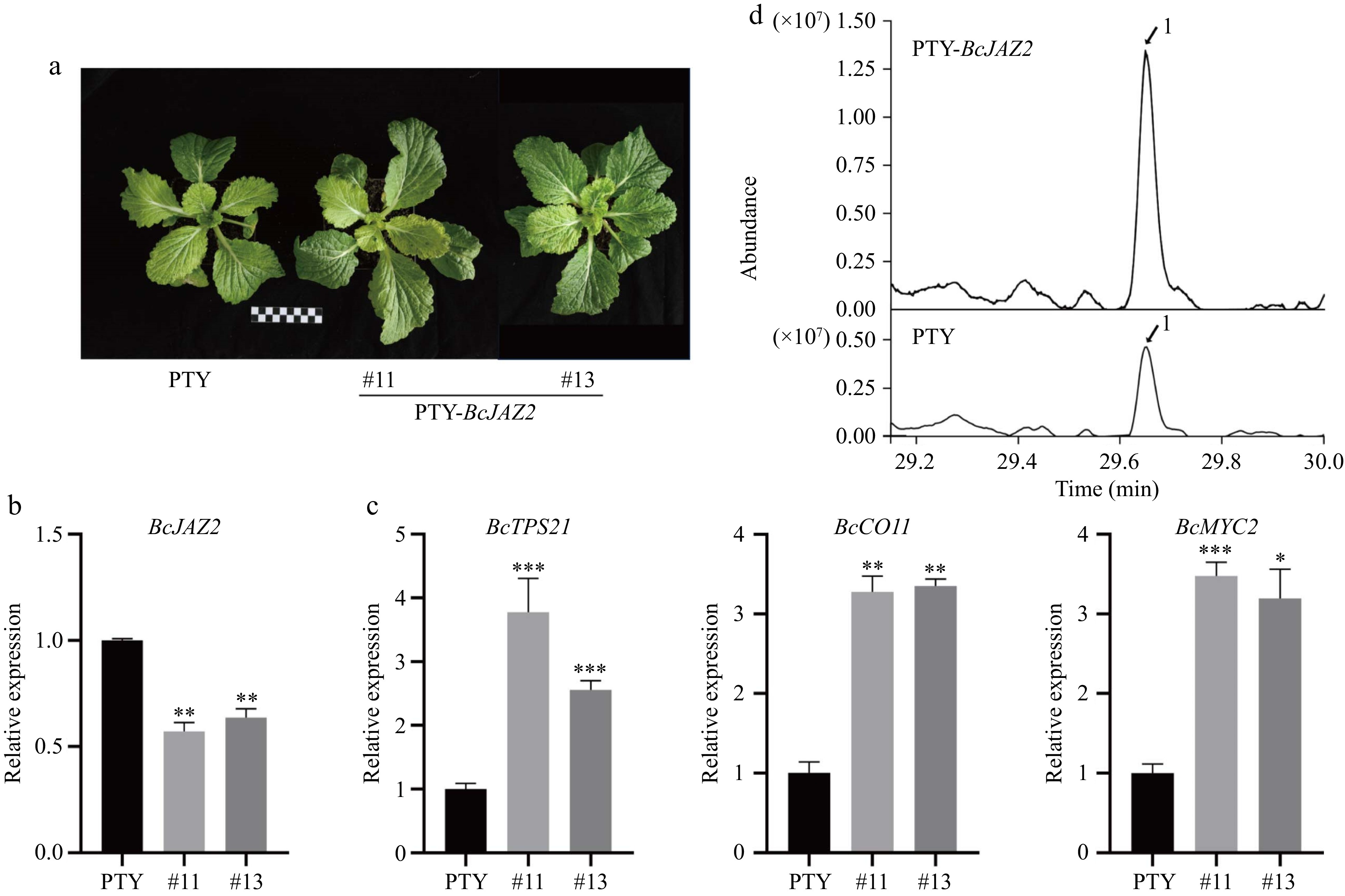
Figure 8.
Silencing BcJAZ2 by VIGS promotes β-caryophyllene biosynthesis in NHCC. (a) The phenotypes of BcJAZ2 in silenced plants. The black and white boxes are scale bar and one box represents 1 cm. (b) Relative expression of BcJAZ2 in silenced and control plants. (c) Analysis of sesquiterpene synthase and JA signaling genes expression level of BcJAZ2 in silenced and control plants. Student's t-test: *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001. Three experiments were repeated and values were means ± SD. (d) GC-MS chromatogram analysis of β-caryophyllene in silenced and control plants. Peaks with number represented sesquiterpenes β-caryophyllene.
BcbHLH137, BcHBI1.1, and BcHBI1.2 are upstream regulators of BcJAZ2
-
To search for relevant transcription factors that may regulate BcJAZ2 expression, we used the yeast one-hybrid screen library technique. Cis-elements analysis of the BcJAZ2 promoter of upstream 1,250 bp sequence showed that there were 13 kinds of cis-elements in this region (Supplementary Table S4). Finally, three candidate genes (BcbHLH137, BcHB1.1, BcHBI1.2) were screened. Subcellular localization and trans-activation activity assays demonstrated that these three genes are located in the nucleus and all have trans-activation activity capacity, consistent with the characterization of transcription factors (Fig. 9a, b). The Y1H and LUC results showed that BcbHLH137, BcHBI1.1, and BcHBI1.2 all positively regulated BcJAZ2 (Fig. 9c−e). These results provided candidate genes for upstream regulation of BcJAZ2 expression, which may provide new clues for exploring more functions of BcJAZ2.
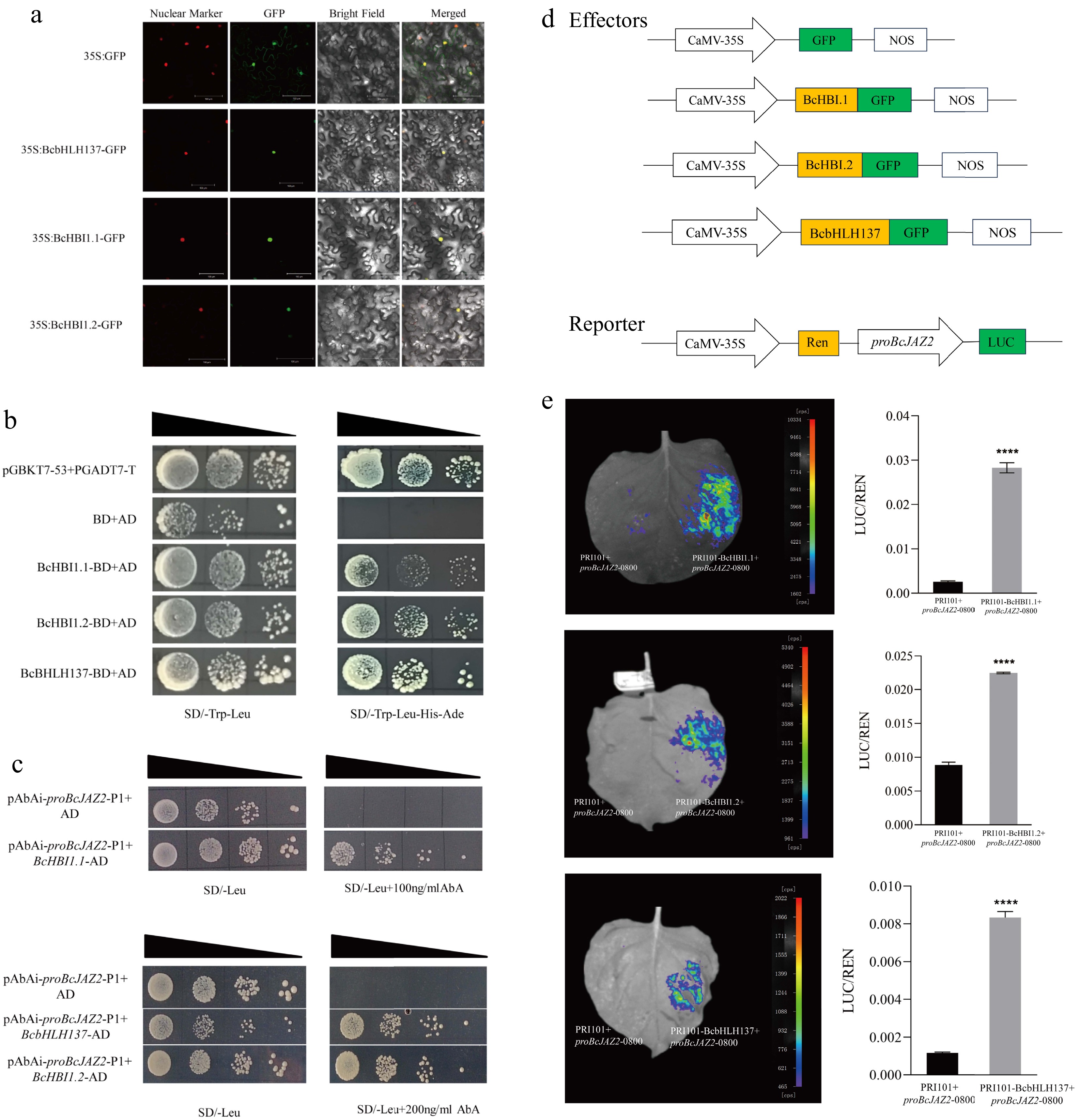
Figure 9.
Screening of upstream regulatory genes of BcJAZ2. (a) Subcellular localization of BcbHLH137, BcHBI1.1, and BcHBI1.2. The scale bar was 100 μm. (b) Trans-activation activity of BcHBI1.1, BcHBI1.2, and BcbHLH137 using yeast two-hybrid assay. (c) Yeast one-hybrid assay. (d) Schematic diagram of the effectors and reporter vectors. (e) Dual-luciferase assay showed that the screened transcription factors promoted BcJAZ2 expression. The data were from three experiments replicates and values were means ± SD. Student's t-test: ****p ≤ 0.0001.
-
Plants possess complex biological regulatory networks that control their growth cycles and the synthesis and accumulation of various substances essential for their survival[28]. JAZ is a key negative regulator of the JA signaling pathway and participates in many biological processes as well as complex synergistic and antagonistic interactions between JA and other hormones[29−32]. Although JAZ gene family has been identified in many species including Arabidopsis[7], rice[33], maize[34], B. oleracea[35], and B. napus[36], there are few reports on the JAZ genes in NHCC. Twenty five BcJAZ genes were identified in this study, named BcJAZ1-25 (Supplementary Table S2). Phylogenetic relationships analysis showed that BcJAZ gene family members are categorized into five families (I, II, III, IV, and V) (Fig. 1b). It was shown that 56 and 31 JAZ genes were identified in the reference genomes of B. napus and B. oleracea[37]. Compared with 12 JAZ genes in Arabidopsis, this is consistent with a whole-genome triplication (WGT) event that have been demonstrated during the evolution of Brassica species. The presence of TIFY and Jas domains in all BcJAZ proteins examined suggested that these domains were highly conserved and may have significant structural characteristics (Fig. 2). Gene duplications confer new functions for the evolution of species, and help them adapt to the environment. Consistent with the findings in B. napus and B. rapa[37,38], there were no instances of tandem duplications in the BcJAZ genes within the NHCC genome (Supplementary Table S3), Intraspecies and interspecies collinearity analyses indicated that segmental duplication was the major duplication event for BcJAZ genes, meanwhile, chromosomal and gene amplification events may have occurred during the evolution of NHCC (Fig. 3a, b).
The dynamic expression of genes under different treatments may reflect the gene functions. By analyzing the promoter cis-elements of 25 BcJAZ genes, some types of development-related response elements, stress-responsive elements, light-responsive elements, and hormone response elements were detected in most BcJAZ promoters (Fig. 4). qRT-PCR data demonstrated that these BcJAZ genes exhibited distinct expression profiles in response to low temperature (4 °C), MeJA, and GA treatments (Fig. 5b−d), which was in accordance with the findings from other Brassica crops[37]. The response to different hormone treatments suggested that JAZ proteins were involved in the crosstalk of different hormone signals to maintain normal plant growth. Notably, RNA-seq data revealed that the expression level of BcJAZ2 was most significantly up-regulated after three days of low temperature (Fig. 5a), while BcJAZ2 was also induced under short-term low-temperature treatment. However, previous studies showed that low temperatures inhibited β-caryophyllene synthesis in NHCC (unpublished data). Therefore, we hypothesized that BcJAZ2 may be a key negative regulator of β-caryophyllene synthesis. Similarly, high-temperature treatment significantly induced the expression of the JA signaling pathway repressor CsJAZ6 in Camellia sinensis, which partially mediates secondary metabolite biosynthesis by affecting the formation of the MBW complex[39].
In our study, we further explored the molecular mechanism by which BcJAZ2 was involved in the regulation of β-caryophyllene. Subcellular localization confirmed that BcJAZ2 was a nuclear-localized protein (Fig. 6a). As a core component of the JA signaling cascade, MYC2 is responsible for the regulation of secondary metabolism across diverse species[16,27,40]. AtMYC2 in Arabidopsis can directly bind to the promoters of the sesquiterpene synthase genes AtTPS21 and AtTPS11 and activate their expression, leading to the release of sesquiterpenes, especially β-caryophyllene[27]. Here, we showed that BcJAZ2 interacted with BcMYC2 through Y2H and BiFC assays (Fig. 6c, d). Furthermore, the transcription of sesquiterpene synthase and JA signaling genes were significantly repressed and the content of sesquiterpene β-caryophyllene was decreased in BcJAZ2 overexpression Arabidopsis compared with the wild-type (WT) Arabidopsis (Fig. 7). As expected, the results in BcJAZ2 silenced plants were opposite (Fig. 8). Hence, we propose that BcJAZ2 serves the role of a transcriptional repressor within the JA signaling, forming a BcJAZ2-BcMYC2 cascade to regulate the biosynthesis of sesquiterpene in NHCC.
Transcription factors can bind specific DNA sequences and regulate gene expression through activation or repression. Three transcription factors (BcbHLH137, BcHB1.1, BcHBI1.2) were screened and identified using the yeast one-hybrid (Fig. 9c). LUC results showed that these three TFs all have positive regulation of BcJAZ2 expression (Fig. 9e). Interestingly, these three TFs all belong to the bHLH family, which regulate cell proliferation and elongation, and their transgenic Arabidopsis exhibits an early flowering phenotype[41,42]. Recent studies showed that JAZ3 transgenic Arabidopsis displayed shorter hypocotyls and larger cotyledons under specific light or temperature environments, compared with the wild-type Arabidopsis[43]. In this investigation, we also found that BcJAZ2 transgenic Arabidopsis have big cotyledons and early flowering phenotypes. Plants can utilize phytohormones to integrate and transmit various signals in order to adjust their growth and development to adapt to the environment, with various phytohormones involved in this regulatory process by interfering with each other. Among them, some key regulators in the hormone signaling pathway can act on multiple hormone signaling processes. Previous studies revealed that the bHLH transcription factors specifically bind to the E-box element[44] (5'-CANNTG-3'). From this, we suspected that bHLH137 and HBI1 might participate in the transcriptional regulation of plant development and terpene under diverse environments and phytohormone signals, possibly through binding to the BcJAZ2 promoter E-box element. However, the precise regulatory mechanisms underlying these processes require further investigation.
-
In conclusion, we performed a genome-wide analysis of the JAZ gene family in NHCC. Expression pattern analyses indicated that BcJAZs gene were responsive to low temperature, MeJA, and GA. In addition, we confirmed that BcJAZ2 negatively regulates β-caryophyllene synthesis by interacting with BcMYC2. Three transcription factors (BcbHLH137, BcHBI1.1, BcHBI1.2) were screened and positively regulated BcJAZ2 expression. However, more evidence is needed to determine whether these transcription factors are also involved in the regulation of β-caryophyllene synthesis in NHCC.
This research was funded by the Jiangsu Seed Industry Revitalization Project (JBGS(2020)15), National Key R&D Program of China (2023YFD2300700) and the Earmarked Fund for China Agriculture Research System (CARS-23-A-16). The authors appreciate those contributors who provide related genomes available in public databases.
-
The authors confirm contribution to the paper as follows: experiments Design, data analysis, and manuscript writing: Han T, Wang H; experimental guidance: Liang Z, Bai A; manuscript revision: Han T, Wang H, Xu H, Li Y; experimental resources offering: Wang J, Liu T, Hou X, Li Y; experiments supporting: Li Y. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Tiantian Han, Haibin Wang
- Supplementary Table S1 Primers used in this article.
- Supplementary Table S2 Physicochemical properties of BcJAZ proteins in NHCC.
- Supplementary Table S3 Paralogous relationships among BcJAZ genes family members in non-heading Chinese cabbage.
- Supplementary Table S4 BcJAZ2 promoter cis-elements prediction.
- Supplementary Fig. S1 The minimum AbA concentration to inhibit yeast self-activation.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Han T, Wang H, Liang Z, Bai A, Xu H, et al. 2025. Genome-wide identification of the JAZ gene family in non-heading Chinese cabbage and the functional verification of BcJAZ2 in the biosynthesis of β-caryophyllene. Vegetable Research 5: e007 doi: 10.48130/vegres-0025-0001 

Genome-wide identification of the JAZ gene family in non-heading Chinese cabbage and the functional verification of BcJAZ2 in the biosynthesis of β-caryophyllene
- Received: 28 September 2024
- Revised: 05 December 2024
- Accepted: 23 December 2024
- Published online: 04 March 2025
Abstract: JASMONATE ZIM DOMAIN (JAZ) proteins function as negative regulators of the JA signaling pathway and participate in plant development, stress responses, and secondary metabolism. β-caryophyllene is a volatile sesquiterpene compound that contributes to the formation of plant aromas and possesses antibacterial, anti-inflammatory, and antifungal biological activities. In our previous experiments, we found that the BcJAZ2 was closely related to β-caryophyllene synthesis under low-temperature treatment in non-heading Chinese cabbage (NHCC). To further explore the function of BcJAZ2, we characterized JAZ gene family in NHCC. In this study, 25 BcJAZ genes were discovered in NHCC, and comprehensively analyzed the evolutionary relationships and structural characterizations of BcJAZs. BcMYC2, a positive regulator of terpenoid synthesis, interacted with BcJAZ2 confirmed by yeast two-hybrid and bimolecular fluorescence complementation assays. Overexpression of BcJAZ2 in Arabidopsis and silencing of BcJAZ2 in NHCC showed that BcJAZ2 acted as a negative regulator of β-caryophyllene biosynthesis. In addition, three transcription factors BcbHLH137, BcHBI1.1, and BcHBI1.2 were confirmed to be positive regulators of BcJAZ2 by yeast one-hybrid and LUC assays. The above results enrich our understanding of the regulation of β-caryophyllene synthesis and provide the foundation for in-depth exploration of regulatory mechanisms of BcJAZs.