









-
The heat shock transcription factor (HSF) family plays a pivotal role in plant biology, serving as a key regulator of responses to environmental stress and growth processes[1]. Under abiotic stress conditions such as high-temperature, drought, and salinity, HSFs bind to heat shock elements (HSEs) in the promoters of heat shock genes, triggering the expression of heat shock proteins (HSPs) and other associated genes[2,3]. This activation helps plants in maintaining proper protein folding and cellular functions, thereby enhancing their resilience to harsh environmental conditions[4]. Ultimately, this mechanism supports plant tolerance to adversity and ensures normal growth and development in stressful environments[5].
The HSF family exhibits highly conserved structural features in plants. Its members typically contain DNA-binding domains (DBDs), which recognize and bind to heat shock elements to regulate downstream gene expression[6]. Based on sequence similarity and structural features, the HSF family is classified into several subfamilies (A, B, C, etc.), with each subfamily exhibiting both functional conservation and some degree of specialization[7]. Recent studies have extensively explored the HSF gene family across various plant species, including model organisms such as Arabidopsis thaliana, Oryza sativa, Zea mays, and economically important crops[8,9]. These investigations have significantly advanced our understanding of the structure, function, and regulatory roles of HSFs in plant stress responses and development. For example, different HSF genes in A. thaliana exhibit distinct yet complementary functions in heat stress responses, seed germination, and flowering regulation. In O. sativa, HSF genes contribute to heat tolerance and responses to other abiotic stresses by regulating downstream gene expression, thereby enhancing the resilience, yield, and quality of O. sativa[10]. In addition to their central role in the heat stress response, HSFs are also involved in key developmental processes, including seed germination, seedling growth, and flowering regulation[11]. These factors operate within a coordinated network, in which different HSF members interact to control plant growth and developmental rhythms[12,13].
As a woody perennial species in the Paeoniaceae family, Paeonia suffruticosa is an important ornamental and medicinal plant with substantial economic value and cultural significance[14]. However, it faces numerous environmental challenges during its growth and development. P. suffruticosa is particularly sensitive to abiotic stresses such as high temperature and drought, which can negatively affect its growth, shorten the flowering period, and reduce flower quality—thereby impacting its ornamental and commercial value[15]. Flower development and morphogenesis in P. suffruticosa are regulated by a complex molecular network, in which heat-related transcriptional regulators may play essential roles; however, research on these genes remains limited[16]. Although the HSF family has been partially studied in several plant species, investigations in P. suffruticosa are still lacking. Therefore, a comprehensive analysis of the characteristics, functions, and regulatory mechanisms of the HSF gene family in P. suffruticosa is of great theoretical and practical significance. It can help elucidate the molecular mechanisms of environmental adaptation and contribute to the development of new varieties with improved stress tolerance.
Given the critical role of the HSF gene family in plant stress tolerance and development, and the environmental challenges faced by P. suffruticosa, this study focuses on a genome-wide investigation of the P. suffruticosa HSF gene family. We will comprehensively analyze gene structure, chromosomal localization, phylogenetic relationships, and conserved domain features. Additionally, transcriptome data will be used to explore gene expression patterns and regulatory networks in response to high-temperature stress. This study aims to fill the research gap regarding the P. suffruticosa HSF gene family, offering key insights into its growth, development, and stress adaptation mechanisms. Ultimately, it will provide a theoretical foundation and genetic resources for the genetic improvement and breeding of P. suffruticosa, promoting the development of a high-quality and stress-resistant industry.
-
The genomic data of P. suffruticosa were sourced from the National Gene Bank of China database (
https://ftp.cngb.org/pub/CNSA/data5/CNP0003098 ), while the genomic data for A. thaliana were retrieved from the TAIR database (www.arabidopsis.org )[17]. Genes were initially screened through a BLASTP search, followed by a refined selection of candidate genes using the Hidden Markov Model of HSF proteins (PF0047) from the Pfam database[18,19]. Finally, the BLASTP and HMMER results were combined to take the concatenated set to identify the candidate genes. Conserved structural domains of the candidate genes were identified using the NCBI-CDD (www.ncbi.nlm.nih.gov/cdd ) and SMART (http://smart.embl.de ) databases. Fifteen PosHSF genes were finally identified and renamed according to their order on the chromosome for subsequent analysis. Proteins were analyzed for instability index, aliphatic index, and total hydrotropic mean using the TBtools (https://github.com/CJ-Chen/TBtools ) physicochemical property analysis program. Subcellular localization predictions were made using WoLF PSORT (https://wolfpsort.hgc.jp/ )[20].Phylogenetic tree construction, gene structure, and motif analysis
-
Phylogenetic tree of O. sativa HSF proteins were obtained from the NCBI database (
https://ngdc.cncb.ac.cn/ ). Phylogenetic tree construction was performed in conjunction with A. thaliana and P. suffruticosa data. Phylogenetic trees were constructed using MEGA 6.0 software with 1,000 bootstrap replicates[21]. Conserved structural domains of HSF proteins were analyzed using the NCBI Conserved Domain Database (CDD). Conserved motifs of the PosHSF protein were analyzed using the MEME suite (https://meme-suite.org/meme/ ). Exon and intron structures of the genes were analyzed using the Gene Structure Display Server (GSDS) tool and merged with TBtools. Additionally, amino acid sequences of conserved structural domains were compared and modified using Jalview[22].Promoter and covariate analysis
-
Cis-acting elements (CREs) were identified from the promoter sequences 2,000 bp upstream of each gene using the Plant CARE database and visualized with TB tools[23,24]. The covariance analysis of the genes was performed using the MCScanX tool, and the results obtained were visualized and analyzed using the Quick MCScanX Wrapper program[25].
Expression profiling and data analysis of the PosHSF gene
-
Expression profiling of the PosHSF gene was conducted using RNA-seq data from eight P. suffruticosa tissues: apical, stem, leaf, petal, stamen, pistil, seed, and adventitious root (
https://ftp.cngb.org/pub/CNSA/data5/CNP0003098/ )[26]. In addition to determine the response of PosHSF to high-temperature stress (CK: 0 h, T1: 2 h, T2: 6 h, T3: 12 h, and T4: 24 h), transcriptome raw data were downloaded from NCBI SRA (Accession No. PRJNA1079236)[27]. WGCNA was employed to identify gene modules linked to heat tolerance, while K-means clustering analysis was performed using the HIP-LOT online tool (https://hiplot.com.cn/home/index.html ) to map expression patterns[28].Plant material, RNA extraction, and qRT-PCR
-
Plant materials for this study were collected from the Henan Agricultural University research base. P. suffruticosa leaf RNA was extracted using the E.Z.N.A Plant RNA Kit (OMEGA, USA) from the leaves treated at a high temperature of 40 °C (CK: 0 h, T1: 2 h, T2: 6 h, T3: 12 h, and T4: 24 h) and subsequently reverse transcribed into cDNA using the ReverTra Ace qPCR RT Master Mix (TOYOBO, Japan). The reaction system consisted of 7 μL of ddH analyzed SYBR Green Real-Time Fluorescence Quantitative PCR Premix (TOYOBO, Japan), 1 μL of cDNA, 2 μL of upstream and downstream primers, and 10 μL of SYBR enzyme. By the 2−ΔΔCᴛ method. Primers were designed using the online NCBI Primer-BLAST tool (
www.ncbi.nlm.nih.gov/tools/primer-blast , Supplementary Table S1). One-way ANOVA was performed, and line plots were generated using Microsoft Excel (2019) software. -
Fifteen PosHSF genes were identified in the P. suffruticosa genome using BLAST and HMMER methods. These genes were designated PosHSF01 to PosHSF15 based on their chromosomal locations (Fig. 1). The length of the proteins encoded by these genes ranged from 124 amino acids (PosHSF10) to 689 amino acids (PosHSF06). Based on their predicted amino acid sequences, the molecular weight (MW) of the 15 PosHSF proteins averaged 43,598.53 Da, ranging from 14,042.71 Da (PosHSF10) to 76,198.41 Da (PosHSF06). The predicted isoelectric points (pI) ranged from 4.86 (PosHSF10) to 9.53 (PosHSF13). Instability index analysis showed that 93.3% of the PosHSF proteins (14 out of 15) were predicted to be unstable, with only PosHSF10 having a stability index below 40, suggesting it may be a stable protein. The aliphatic index (A.I.) varied from 60.43 (PosHSF09) and 82.49 (PosHSF02), suggesting slight variations in thermal stability among these proteins. Additionally, all PosHSF proteins were hydrophilic, with negative GRAVY values. Subcellular localization predictions showed that all PosHSF proteins were localized in the nucleus (Supplementary Table S2).
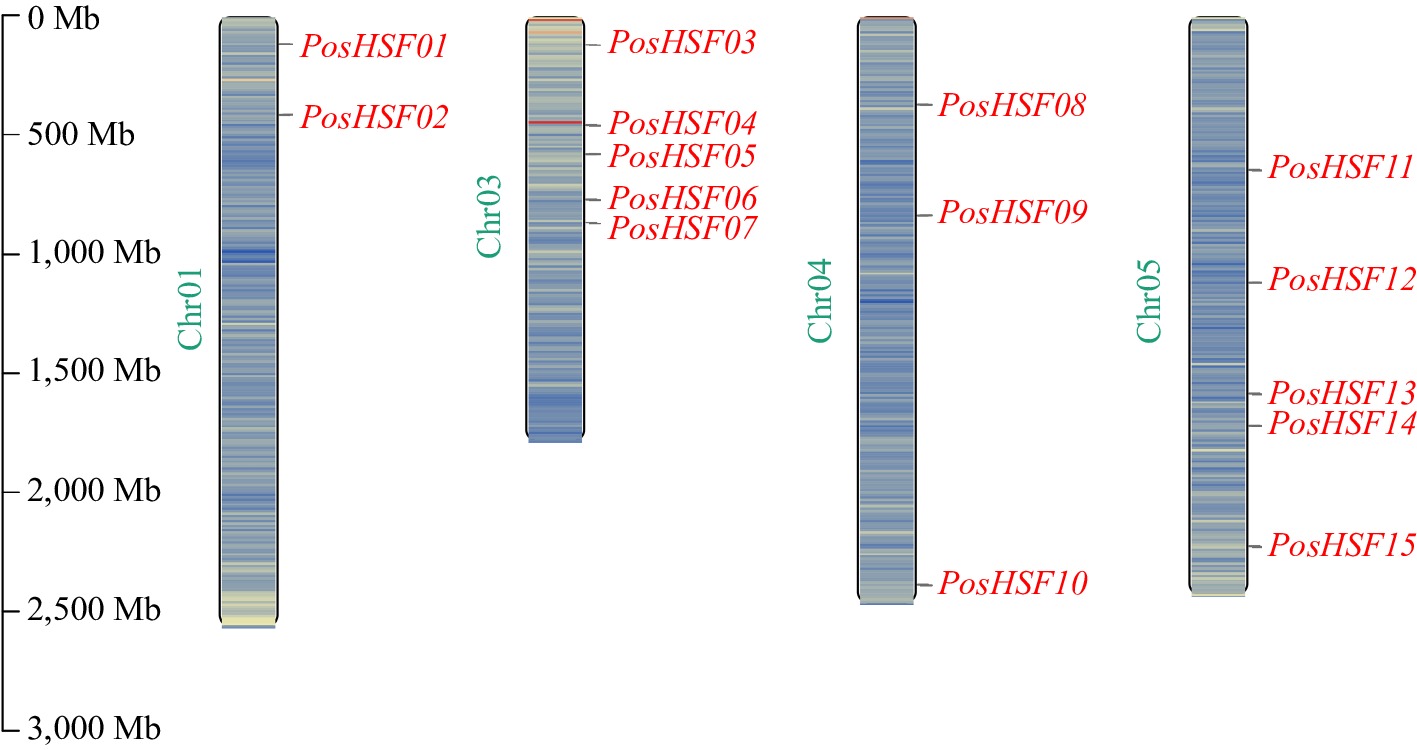
Figure 1.
Chromosomal localization of PosHSF genes in P. suffruticosa.
Phylogenetic analysis and evolutionary classification of PosHSF proteins
-
To investigate the evolutionary relationships of PosHSF proteins, a neighbor-joining (NJ) phylogenetic tree was constructed using 15 PosHSF, 21 AtHSF, and 25 OsHSF proteins (Fig. 2). According to the classification system for the A. thaliana HSF family, the PosHSF proteins were categorized into three main groups: HSF A, HSF B, and HSF C. HSF A includes nine subgroups (A1−A9), with 10 PosHSF proteins identified, although no PosHSF members were found in subgroups A2 and A7. HSF B was divided into four subgroups (B1−B4), with PosHSF proteins primarily distributed in B1 to B3. No PosHSF proteins were assigned to subgroup C2 in HSF C. The phylogenetic tree further revealed that AtHSF proteins were primarily clustered in group A, while OsHSF proteins were mainly found in group B. The distribution of PosHSF proteins resembled that of A. thaliana, highlighting the evolutionary divergence between dicots and monocots.
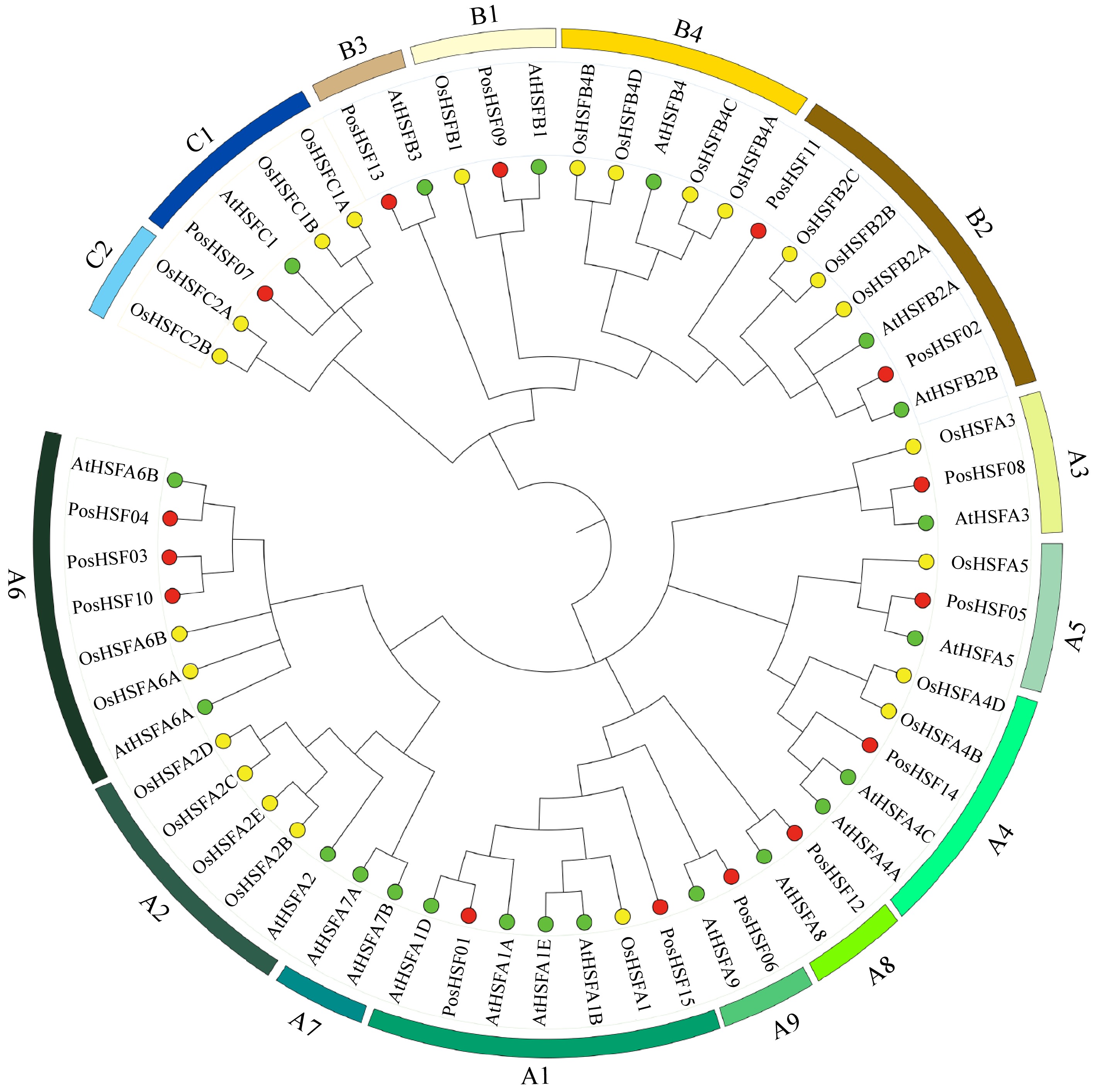
Figure 2.
Phylogenetic relationships of HSF proteins across P. suffruticosa, A. thaliana, and O. sativa.
Conserved structural domains of PosHSF proteins
-
Conserved domains analysis using NCBI-CDD revealed that all 15 PosHSF proteins contained a highly conserved DNA-binding domain (DBD) of approximately 100 amino acids (Fig. 3a, d). To further explore structural diversity, the 10 most conserved motifs (Motifs 1−10) were identified using MEME software (Fig. 3b). The DBD structure comprises motifs 1, 2, and 4, and was present in all PosHSF proteins except PosHSF10, indicating the high conservation of this domain. Gene structure analysis showed that the number of exons in PosHSF genes ranged from 1 to 5, with most genes containing two exons and two introns (Fig. 3c). Additionally, PosHSF proteins with closer evolutionary relationships shared similar exon-intron structures, further supporting their relatedness.
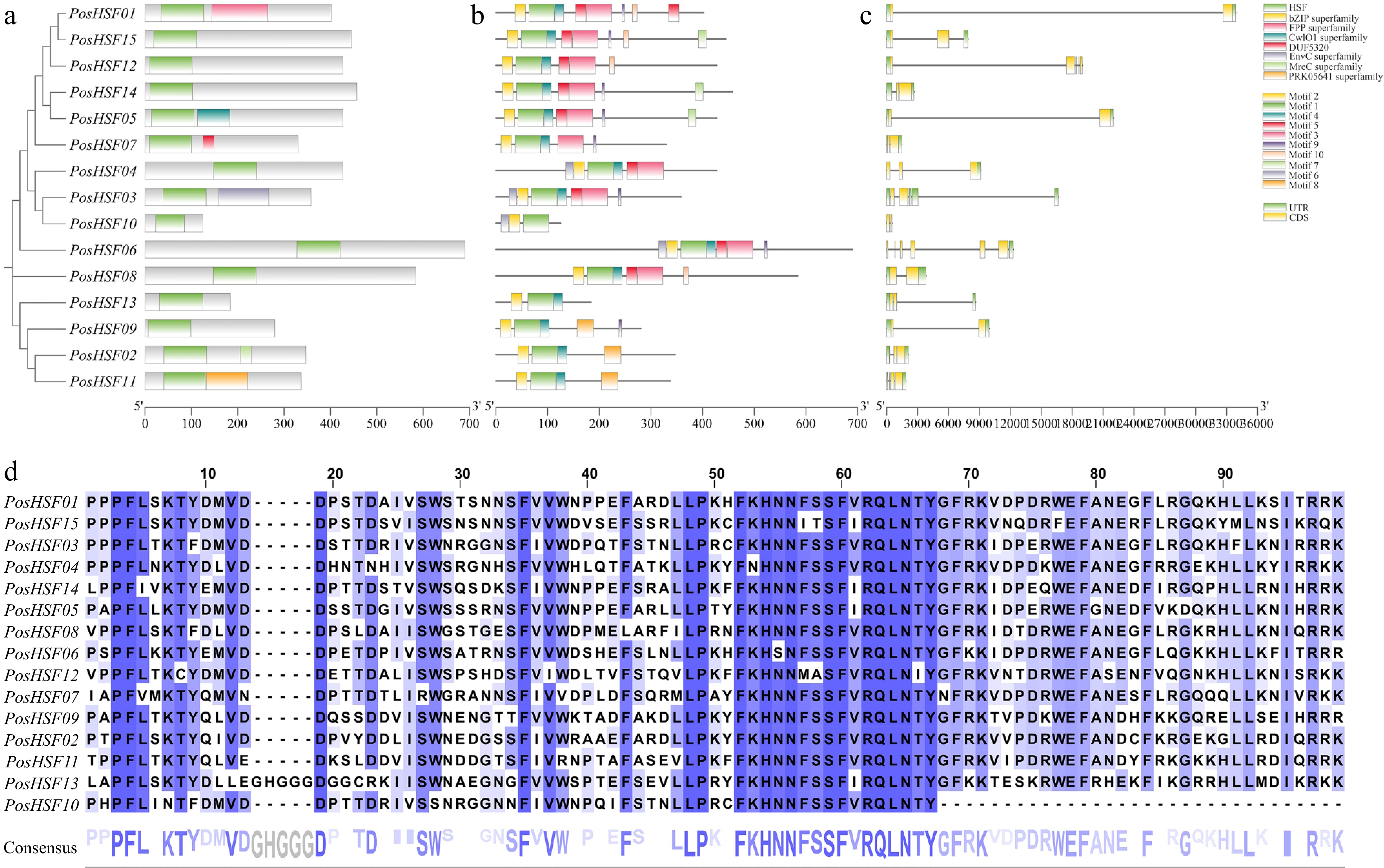
Figure 3.
Characterization of PosHSF genes. (a) Conserved protein structural domains: the green color indicates the most conserved DBD structural domain in the HSF protein. (b) Amino acid motifs in PosHSF: motifs 1−10 in PosHSF are numbered and analyzed. (c) Gene structure of PosHSF, overview of the gene structure in PosHSF genes. (d) Multiple sequence alignment of the DBD structural domain in PosHSF proteins.
Cis-acting element analysis of PosHSF promoters
-
To explore the potential regulatory functions of PosHSF genes, the 2,000 bp upstream promoter regions of the 15 genes were analyzed for cis-acting elements (Fig. 4a). These elements were classified into three categories: responses to abiotic and biotic stresses, phytohormones responses, and plant growth and development (Fig. 4b). Among these growth and development-related elements were the most abundant, with the Box4 element detected in 13 PosHSF genes. Elements related to biotic stress, such as ARE, were present in 14 genes, while the ABRE element, associated with abscisic acid response, was found in 11 genes. These findings suggest that PosHSF genes are likely involved in complex regulatory networks responding to both environmental and hormonal signals.
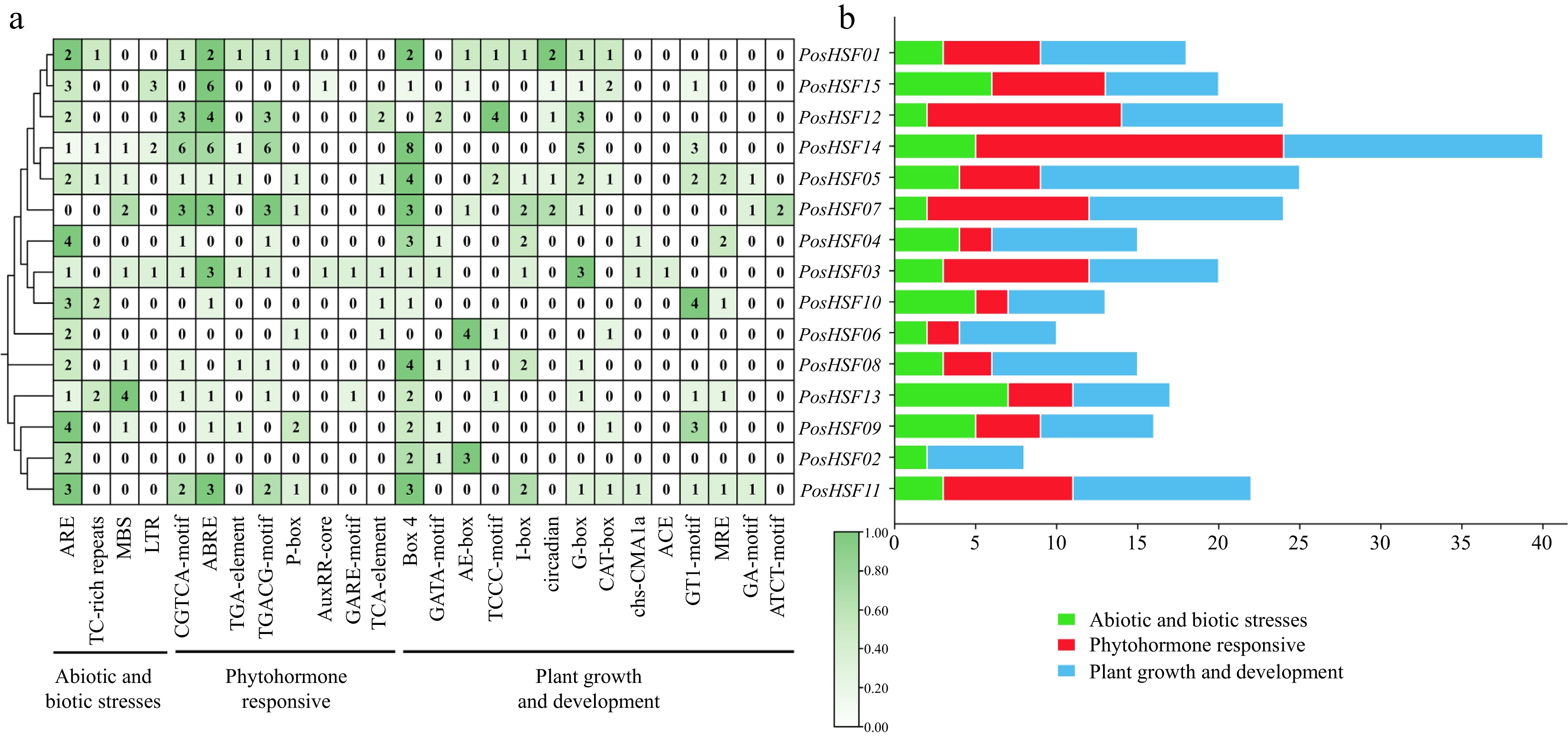
Figure 4.
Analysis of promoter cis-acting elements of PosHSF. (a) Heat map and categorization of PosHSF promoter cis-acting elements. (b) Distribution numbers of the three categories of promoter cis-acting elements in the PosHSF gene.
Covariance analysis of HSF gene homologs
-
To explore the evolutionary conservation of HSF genes, synteny analysis was conducted with the dicot A. thaliana, the monocot Zea mays, and the related species Vitis vinifera and Paeonia ludlowii. The analysis revealed five pairs of homologous genes between P. suffruticosa and A. thaliana and four pairs between P. suffruticosa and Z. mays. Notably, the number of homologous gene pairs between P. suffruticosa and A. thaliana was significantly higher than that between P. suffruticosa and Z. mays, suggesting that P. suffruticosa is more closely related to dicotyledons. In addition, the HSF gene showed a higher degree of variation during the evolutionary process, exceeding the variation of the overall gene (Fig. 5a). Further analysis (Fig. 5b) showed that P. suffruticosa is more closely related to P. ludlowii, with a much higher number of homologous gene pairs than in comparison with V. vinifera. Although there are 12 homologous gene pairs between P. suffruticosa and P. ludlowii, slightly fewer than the 13 pairs between P. suffruticosa and V. vinifera, this result further supports the conclusion that the HSF genes are more variable than the other genes during the evolutionary process.
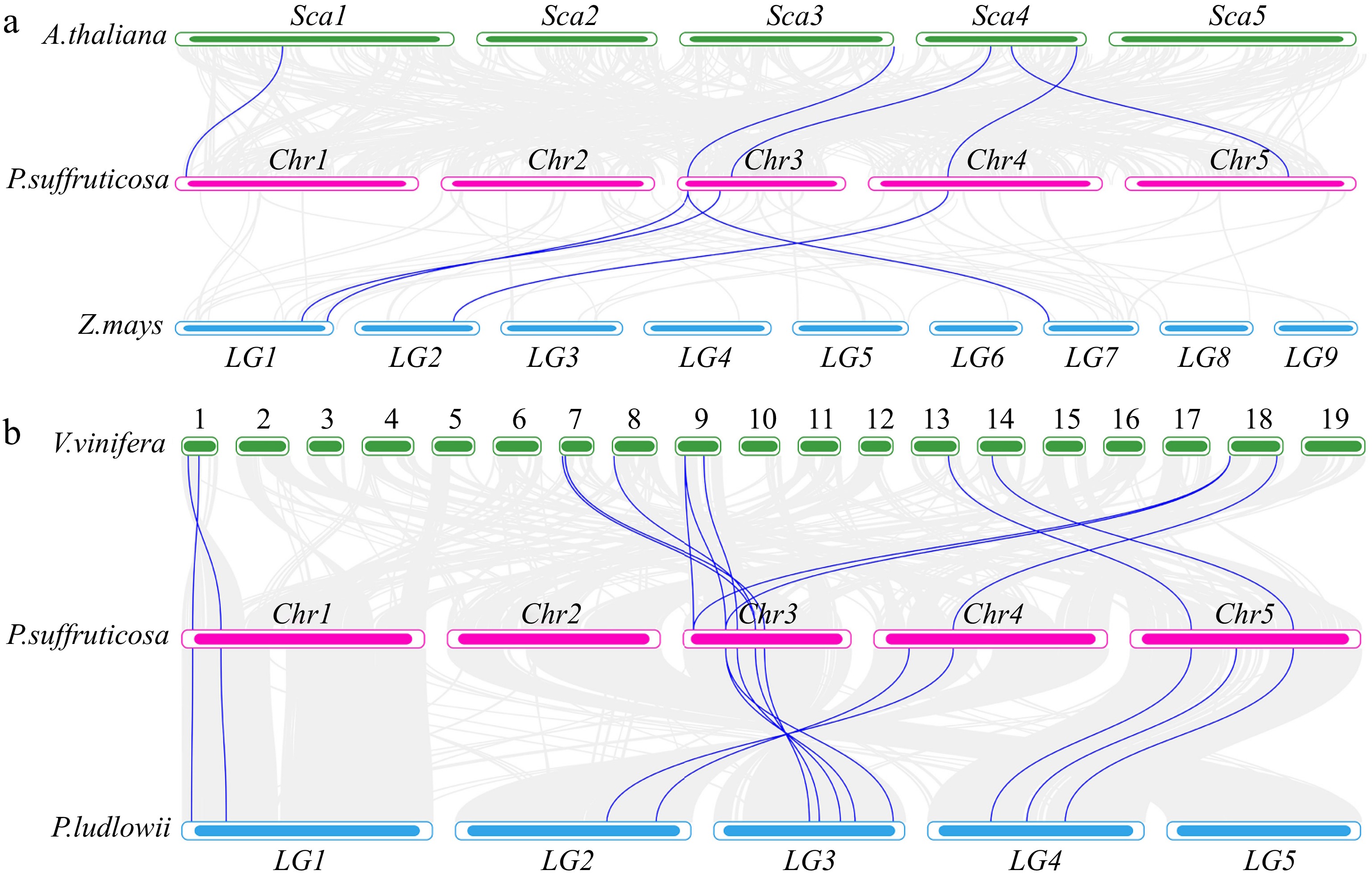
Figure 5.
(a) Synteny analysis of HSF genes in P. suffruticosa, A. thaliana, and Z. mays. (b) Synteny analysis of HSF genes in P. suffruticosa, P. ludlowii, and V. vinifera.
Tissue-specific expression patterns of PosHSF genes
-
To investigate the expression patterns of PosHSF genes across different tissues, a hierarchical clustering heatmap was constructed using NCBI's RNA-seq public data. The expression of 15 PosHSF genes was analyzed in eight tissues: apical, roots, stems, leaves, petals, pistils, seeds, and stamens (Fig. 6). The results showed that PosHSF genes were most prominently expressed in roots; in addition, PosHSF06 and PosHSF11 were more prominently expressed in seeds. Notably, the PosHSF10 gene was not expressed in all tissues, a phenomenon that has been verified identically in other plants such as Medicago sativa. This result strongly verifies that the HSF10 gene is highly conserved during the evolutionary process and provides important theoretical support for subsequent in-depth studies.
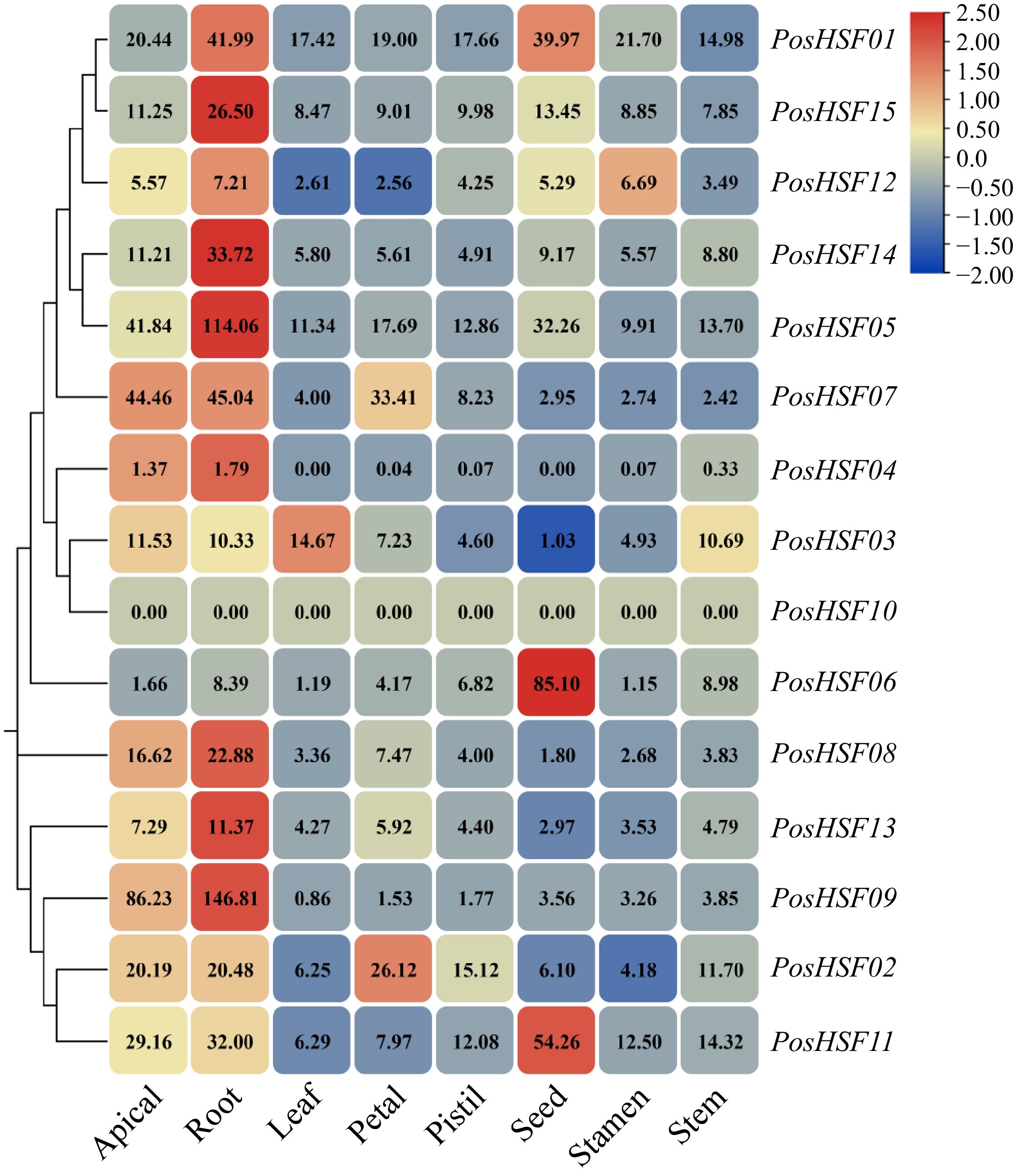
Figure 6.
Expression profiles of tissue-specific PosHSF genes in different tissues.
WGCNA-based identification of heat stress responsive PosHSF genes
-
To investigate the role of PosHSF genes in the heat tolerance of P. suffruticosa, this study analyzed the transcriptome data of P. suffruticosa leaves under high-temperature treatments and screened for key genes using WGCNA (Fig. 7a). WGCNA analysis showed that heat stress-responsive genes in peony were classified into eight co-expression modules. In particular, the genes in the MEblue module showed significant negative regulation, and these genes may play key roles in the response of P. suffruticosa to high-temperature stress. Further K-means clustering analysis categorized the 461 genes in the MEblue module (Fig. 7b), resulting in the classification of these genes into seven classes. To delve deeper into the response of PosHSF genes in the heat tolerance of P. suffruticosa, the study searched these seven modules and found that PosHSF03 (Pos.gene22654.mRNA-1) and PosHSF08 (Pos.gene33889.mRNA-1) were clustered in group 6. The genes in this group showed an expression trend of increasing and then decreasing in response to high-temperature stress. The expression pattern of PosHSF genes is shown in the heatmap (Fig. 7c). The results indicated that PosHSF03 and PosHSF08 were significantly expressed at the early stage of high-temperature treatment (T1), suggesting their key role in the initial response of P. suffruticosa to high-temperature stress. These findings warrant further validation and investigation.
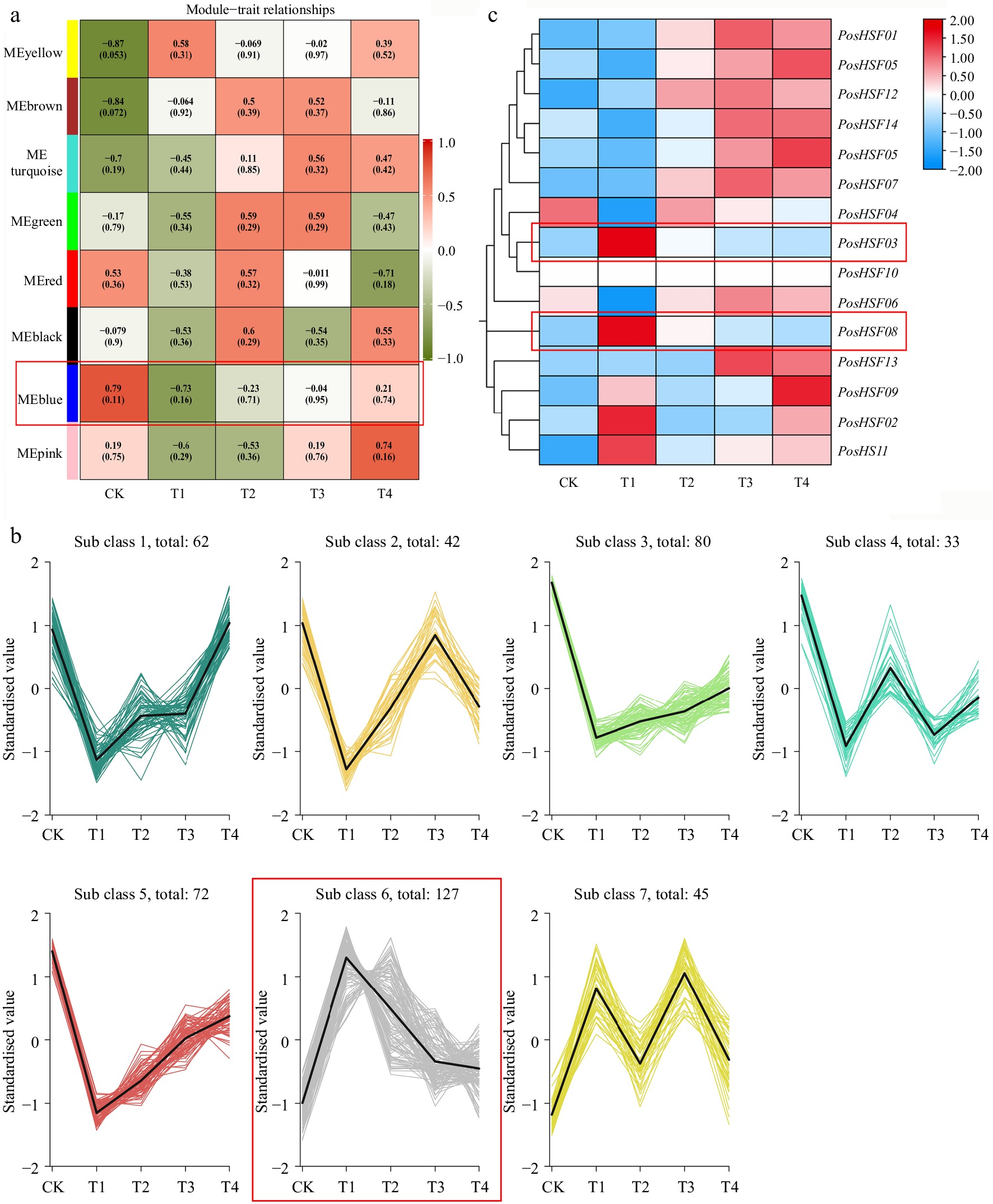
Figure 7.
Analysis of heat tolerance transcriptome data in P. suffruticosa. (a) WGCNA heatmap of module-trait correlations. (b) K-means clustering of gene expression in the blue module. (c) Heatmap of PosHSF expression under high-temperature treatment. Red boxes indicate the modules of focus.
Expression and qRT-PCR analysis of PosHSF genes
-
To further understand the expression of PosHSFs in response to treatments at different high-temperature times, we analyzed the expression of the above 15 PosHSFs members at different high-temperature times (Fig. 8). Comparative analysis of the FPKM values of the genes in the transcriptome data with the expression levels obtained by qRT-PCR showed that the expression patterns were consistent with the transcriptome data.
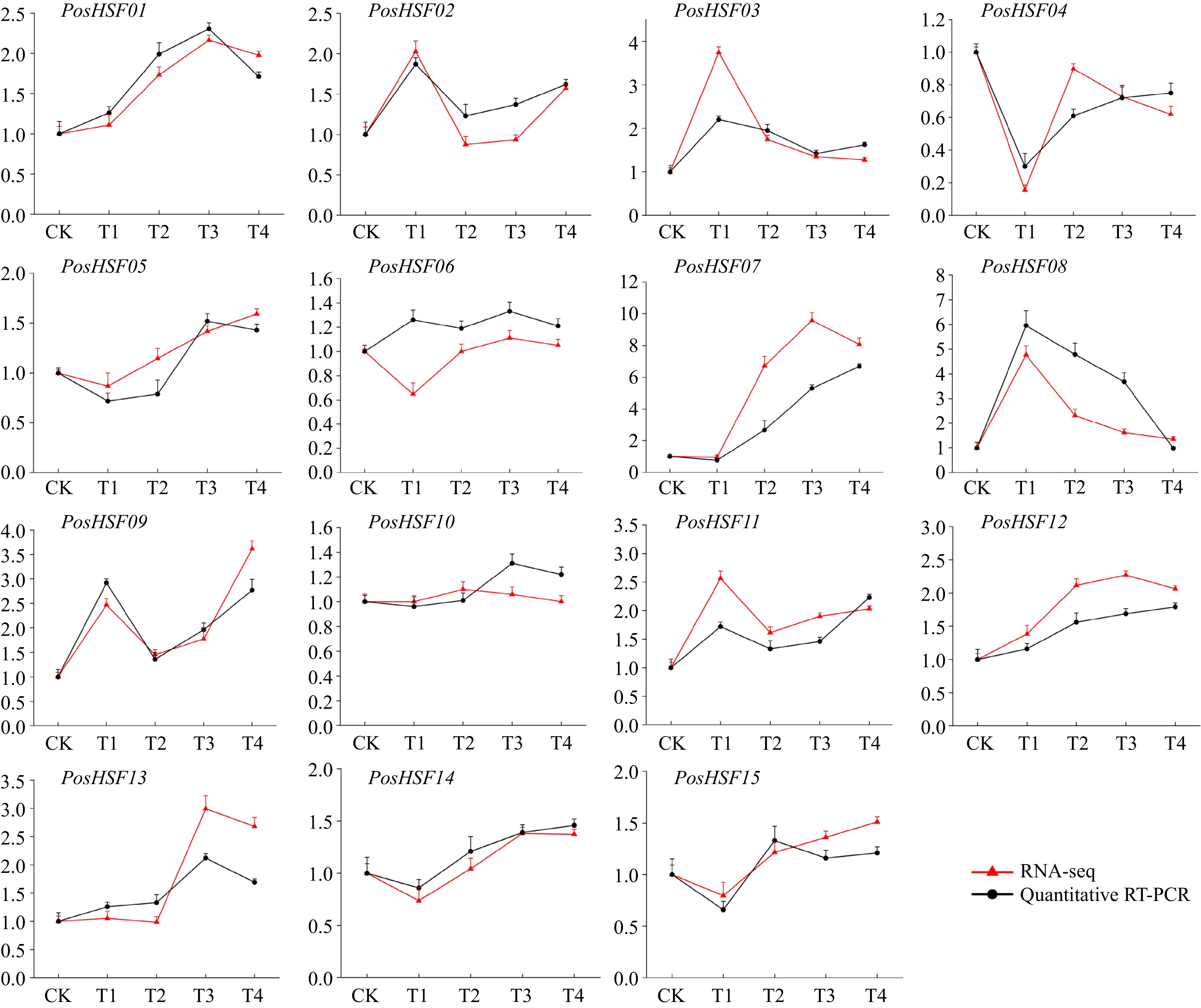
Figure 8.
Expression levels of 15 HSF members of P. suffruticosa in RNA-seq and qRT-PCR. The red line represents the results of RNA-seq, and the black line represents qRT-PCR.
-
HSF (heat shock factors) are crucial transcription factors that regulate plant growth, development, and stress responses[29]. Recent studies on various model plants, crops, and forest trees have highlighted their role in both environmental stress responses and developmental processes, including floral organ formation and stress tolerance[22]. For example, HSFs have been shown to play a crucial regulatory role in response to abiotic stresses such as high temperature and drought in crops such as Triticum aestivum, Poplar, and O. sativa[30−32]. However, studies on HSF genes in P. suffruticosa are still relatively limited; particularly regarding their potential functions in the flowering process and responses to abiotic stresses. Therefore, the systematic identification and functional characterization of the HSF gene family in P. suffruticosa is essential for a deeper understanding of their role in plant growth, development, and environmental adaptation.
Identification and classification of PosHSF genes
-
In this study, 15 PosHSF genes were identified in the P. suffruticosa genome using BLAST and HMMER methods and were named PosHSF01-PosHSF15 according to their chromosomal locations. The number of PosHSF genes distributed across chromosomes indicates that the size of HSF gene families varies significantly among different plant species. This variation is closely related to the genome size and the ecological environments adapted to by each species[33]. For example, O. sativa and A. thaliana contain 22 and 24 HSF genes, respectively, whereas P. suffruticosa contains significantly fewer (15), similar to V. vinifera. Furthermore, the PosHSF genes in P. suffruticosa exhibit phylogenetic variability, similar to that observed in model plants, especially in the distribution patterns between the HSFA and HSFB groups. This suggests that the evolution of the HSF gene family is influenced by species affinity.
Protein characterization of the PosHSF gene
-
Amino acid sequence analysis revealed significant differences in the molecular weights and isoelectric points of PosHSF proteins, which may be related to their functions. Most PosHSF proteins were predicted to be unstable, suggesting their involvement in rapid transcriptional regulation. This finding is consistent with the study by Chauhan et al. on T. aestivum HSF genes, which proposed a role in quick stress responses[34]. Similarly, Cheng et al. reported that plant HSF proteins often exhibit instability under stress conditions, potentially enabling their rapid function in response to environmental challenges such as heat and salinity[35]. The present study supports these observations, implying that the functions of PosHSF proteins in P. suffruticosa are analogous to those in other plant species.
Evolutionary relationships of the PosHSF gene
-
Covariance analysis of P. suffruticosa showed that P. suffruticosa has more homologous gene pairs between mono- and di-cotyledonous model plants and A. thaliana, so that P. suffruticosa is evolutionarily closer to dicotyledonous plants. This finding is consistent with Zhang et al. study of the Glycine max HSF gene families, which suggests that the evolutionary trajectories of HSF genes in different species are closely related to their ecological adaptations[36]. In addition, Yurina pointed out in their study that the HSF gene family of plants has a high variability among similar species, which is the same as the results of the homologous gene pairs in the HSF genes among P. suffruticosa, P. ludlowii, and V. vinifera in the present study, suggesting that there is a high variability of HSF genes in the evolutionary process, which may enable plants to better adapt to different environmental stresses[37].
Transcriptional regulation and stress response of the PosHSF gene
-
Cis-acting element (CRE) analysis of the promoter region revealed that the PosHSF genes play roles in several processes such as plant growth, stress response, and hormone regulation, especially in the enrichment of Box4, ARE, and ABRE elements. The Box4 element is significantly enriched in HSF genes in many plants. Box4 elements are significantly enriched in HSF genes in many plants, especially under environmental stresses such as high temperature and salt stress, where Box4 elements may play an important role. In addition, the study of Yuan et al. revealed the importance of ARE elements in plant stress response, especially under heat stress with a significant regulatory role[38]. The enrichment of these elements suggests that the PosHSF gene may be involved in plant growth and development and stress response through a regulatory network of multiple transcription factors and stress-related genes.
The results of WGCNA analysis in this study showed significant expression of PosHSF03 and PosHSF08 in high-temperature treatments, suggesting that they play key roles in the early stages of P. suffruticosa in response to heat stress. This result coincided with the findings of Kumar et al. and Wang et al. in Z. mays and O. sativa, suggesting that specific HSF genes play important roles in the early stages of heat stress[39,40]. In particular, the expression patterns of PosHSF03 and PosHSF08, which were first increased and then decreased, may be related to the dynamic regulatory mechanism of plant stress response, which also provides new ideas about the role of HSF genes in heat tolerance.
Role of the PosHSF gene in flower morphogenesis
-
A tissue-specific study of P. suffruticosa revealed that the PosHSF gene was significantly expressed in P. suffruticosa petal species. Similar to the study of Chauhan et al. in T. aestivum, this suggests that the HSF gene not only plays a role in stress response but also participates in the regulation of plant organ development[34]. In addition, analysis of P. suffruticosa floral morphogenesis showed that the expression of the PosHSF gene was particularly significant in the pistil, further validating the important role of the HSF gene in floral organ development. Therefore, among the transcription factors related to floral organ development, the HSF gene may play a central regulatory role in the early development of floral organs, which provides new insights into the formation of floral organs in P. suffruticosa.
-
This study presents a comprehensive phylogenetic and functional characterization of the HSF gene family in P. suffruticosa. A total of 15 PosHSF genes were identified, showing conserved structural features and significant roles in high-temperature stress response. Phylogenetic and evolutionary analyses revealed high similarity with HSF genes in A. thaliana, while promoter analysis emphasized their involvement in growth, development, and stress responses. Expression patterns of PosHSF03 and PosHSF08 during early heat stress, revealed by WGCNA, indicates their essential functions in heat tolerance. Overall, this study provides valuable insights into the functional roles of HSF genes in P. suffruticosa and lays a theoretical foundation for breeding programs aimed at improving stress resistance and flower traits in peony.
This work was funded by the Central Plains Scholar Program of Henan Province (Grant No. 244000510002), the Natural Science Foundation of China (Grant No. 32271956), the Science and Technology Program of Shanghai (Grant No. 21DZ1202000), the China Postdoctoral Science Foundation (Grant No. 2024M760753), and the Central Government Led Local Science and Technology Development Fund of Henan Province (Grant No. Z20231811082).
-
The authors confirm contribution to the paper as follows: study design and supervision: Zhang G, He S, Wang Z; writing the manuscript and data analysis: Zhang G, Song Y, Sun Y; assisting in the bioinformatics analysis of gene families: Li D,Wang G, Shi L, Shang W; participating in the experimental procedures: Liu W; providing guidance and revising the manuscript: Zhang G, He S, Wang Z. All authors reviewed the results and approved the final version of the manuscript.
-
The data supporting the results of this study are available in the National Gene Bank of China database, No. CNA0050666, and the high-temperature stress transcriptome data are available in the NCBI database, No. PRJNA1079236.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Guozhe Zhang, Yuke Sun
- Supplementary Table S1 qRT-PCR primer list.
- Supplementary Table S2 Physicochemical property analysis of PosHSF proteins.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang G, Sun Y, Li D, Shi L, Shang W, et al. 2025. Identification, evolution, and functional characterization of the HSF gene family of Paeonia suffruticosa: implications for high-temperature stress response. Ornamental Plant Research 5: e028 doi: 10.48130/opr-0025-0026 

Identification, evolution, and functional characterization of the HSF gene family of Paeonia suffruticosa: implications for high-temperature stress response
- Received: 27 December 2024
- Revised: 09 April 2025
- Accepted: 19 May 2025
- Published online: 03 July 2025
Abstract: The heat shock transcription factor (HSF) family plays a crucial role in regulating plant growth, development, and responses to environmental stresses, particularly those caused by high temperatures and drought. In this study, we systematically characterized the HSF gene family in Paeonia suffruticosa and identified 15 PosHSF genes, which exhibited a high degree of conservation in both gene structure and function. Phylogenetic analysis revealed that the PosHSF genes shared strong similarities with the HSF genes of Arabidopsis thaliana and could be categorized into three groups (HSF A, HSF B, and HSF C). Furthermore, gene evolution and cis-acting element analyses indicated that the promoter region of PosHSF genes was enriched with multiple regulatory elements involved in plant growth, development, and stress responses—particularly in response to high-temperature stress. WGCNA analysis demonstrated that the expression changes of PosHSF03 and PosHSF08 at the early stage of high-temperature treatment suggested their key roles in heat tolerance. This study not only provides new insights into the function of the P. suffruticosa HSF gene family but also offers a theoretical foundation for improving stress resistance and driving varietal innovation in P. suffruticosa, thereby promoting the high-quality development of the P. suffruticosa industry.