










-
As the most dominant plants on the planet, flowering plants play a critical role in providing food and habitat for terrestrial animals[1]. They have evolved rapid diversification in flowering habits to adapt to environmental changes[2]. Understanding flowering habits offers potential strategies to mitigate heat damage in agricultural production under a warming climate. In the past half-century, heat stress has intensified in terms of frequency, intensity, and duration, and the trend is projected to worsen globally by the end of this century[3]. This changing environment poses a significant threat to agricultural production, especially when heat stress coincides with crop flowering[4]. Regrettably, the flowering time of many staple crops, such as maize (Zea mays L.), rice (Oryza sativa L.), sorghum (Sorghum bicolor (L.) Moench), and soybean (Glycine max (L.) Merr.), often coincides with periods of high-temperature stress in practical agricultural settings[5−7].
Both stress avoidance and tolerance are crop adaptations to abiotic stimuli[8]. Avoiding or reducing the overlap between heat stress and flowering time is considered an effective measure to mitigate the impact of heat stress on crop reproduction[9,10]. Nevertheless, completely avoiding the overlap by adjusting flowering time through altering phenology and sowing dates is highly challenging, especially in multi-cropping regions. The diurnal flowering habit, which determines the time of day when a floret opens each day, becomes crucial for facilitating crop reproduction and avoiding heat stress during hot seasons[6,11]. In the plant kingdom, there are nearly 295,000 known species of flowering plants worldwide, with over 100 of these being significant cultivated species[12]. They are categorized into different groups (e.g., morning-, afternoon-, and night-flowering) based on their diurnal floret opening time (DFOT) within the 24-h cycle. This timing is closely linked to both internal circadian rhythms and external environmental conditions[2,13]. DFOT is also dependent on the structure and function of floret organs, such as lodicules in the grass plants and petals in the eudicot plants[14]. Key genes like OsMYB8 and EARLY MORNING FLOWERING 1 identified in rice can alter DFOT by regulating the cell wall structure of lodicules[11,15]. This modulation facilitates cross-fertilization between indica and japonica varieties. Generally, indica rice opens its florets 1−3 h earlier each day compared to japonica. This suggests that the stable DFOT of plants can be intentionally altered, providing a strategy to enhance crop heat stress resistance. In this study, we analyzed the global-scale warming trend since 1850, compared the diurnal variation of temperature during the hottest months across global croplands over the past 20 years, collected DFOT data from 102 plant species (primarily cultivated crops), and summarized the molecular mechanism underlying DFOT regulation in cereal crops. We classified these plant species into three flowering-time groups based on DFOT: morning-flowering (5:00–10:00), midday and afternoon-flowering (11:00–17:00), and night-flowering (18:00–3:00). We then compared their optimum flowering temperatures. To fully elucidate the relationships between DFOT and heat escape, we selected four cereal crops involved in heat stress experiments: maize (morning-flowering), rice (indica, morning-flowering; japonica, midday and afternoon-flowering), sorghum, and pearl millet (night-flowering). This study concludes with evidence that the early-morning flowering (EMF)/pollination trait enhances heat escape in crops. These findings have significant implications for breeding climate-resilient crop varieties.
-
Global annual land temperatures from 1850 to 2023, derived from NOAA (2024), were analyzed to calculate temperature anomalies relative to the 1901–2000 average, highlighting the global temperature trend over time. Hourly temperatures every day during the hottest months of the year in croplands of Asia, Europe, Africa, North America, and South America over the past 20 years were downloaded from NASA POWER (2024)[16]. We selected 10 sites each in Asia, Europe, Africa, North America, and South America. These sites are evenly distributed across croplands. High-temperature increases over time are reflected as the rise in daily maximum temperature (Tmax) and the extension of midday heat episode duration (Dmax). Dmax was defined as the total hours in a day when the hourly temperature exceeded 29 °C[17]. For each year from 2004 to 2023, the average values of Tmax and Dmax during the hot season were calculated.
Diurnal floret opening time (DFOT) in flowering plant species
-
Data on the DFOT of plant species were collected using the Web of Science (WoS,
http://apps.webofknowledge.com/ ) and China National Knowledge Infrastructure (CNKI,www.cnki.net ) to search peer-reviewed research articles published prior to October 2024. We adopted keyword combinations of 'flowering time', or 'flowering rhythm', or 'floret opening time', or 'diurnal floret opening time', or 'flower opening time'. After searching through thousands of articles, we identified 115 studies (published between 1905 and 2024) that specifically reported the DFOT of 102 plant species (Supplementary Table S1). Among these, the optimum flowering temperatures for floret opening of 47 plant species were identified.DFOT and high temperature in cereal crops
-
To detect the responses of plant species with different DFOTs to high temperature, we focused on temperature-controlled experiments conducted in both field and greenhouse worldwide. Flower structure may affect DFOT responses to high temperature; hence, we only focused on cereal plants, including maize, rice, sorghum, and pearl millet, which represent plant species with different DFOTs. Maize tassels and indica rice typically open their florets in the morning, japonica rice open florets during midday and afternoon, while sorghum and pearl millet open florets at night or in the early morning hours[18−21]. In this section, we also searched journal articles using WoS and CNKI. The keyword combinations used were 'high temperature', or 'heat', or 'yield', or 'seed number', or 'kernel number', or 'seed set'. Additionally, we included species-specific keywords: 'maize', or 'corn', or 'Zea mays' for maize, 'rice', or 'Oryza sativa' for rice, 'sorghum', or 'Sorghum bicolor' for sorghum, and 'pearl millet', or 'Pennisetum glaucum' for pearl millet. The studies should meet the following three criteria to reduce publication bias. First, factorial experiments had several temperature treatments, including a control group and different high temperature treatments, either in a greenhouse or in the field. In these experiments, crop management regarding weeds, pests, diseases, and fertilizers was well performed. Second, the temperature treatments were performed during or near the flowering stage. Temperature treatments lasting only a few hours were excluded from our database. Third, studies that did not measure the seed set ratio were excluded to enhance the precision of comparisons between different plant species. In total, 31 journal articles met the criteria and were included (Supplementary Table S2).
Early-morning flowering (EMF) and heat escape
-
Varieties with EMF trait in wheat and rice have been reported[22,23]. In wheat, control and EMF varieties were grown under control temperature of 24/16 °C (day/night) in a chamber until the start of flowering. When the first anther was visible, half plants of control and EMF varieties were exposed to a high temperature of 34/16 °C for 10 d, with the other half maintained under control temperature[23]. In rice, 6-year field experiments that included control and EMF varieties were performed[22]. In this study, sowing dates were strategically selected to ensure high-temperature exposure during the flowering stage. In maize, we did not find studies that used EMF varieties as plant materials. Instead, a temperature-controlled experiment was conducted with five hand-pollination timings (from 7:00–8:00 to 11:00–12:00 at one-hour intervals) and two temperature levels (30/20 °C and 40/30 °C, maximum/minimum) during the pollen-shedding period[24]. In both temperature treatments, the day-time temperature steadily increased at 7:00 and peaked at 13:00, and then decreased back to night-time temperature at 19:00. Hand-pollination was only performed once for one pollination timing treatment. Sufficient pollen grains were collected under the same temperature conditions to ensure consistent pollination of the silks of the ear. To prevent contamination, all ears were enclosed in paper bags both before and after hand-pollination. Maize plants were subjected to high-temperature stress (40/30 °C) for one day and were subsequently transferred to a moderate temperature regime (30/20 °C) for one week. Following this period, all plants were returned to natural field conditions for further growth and development. These studies in wheat, rice, and maize all measured the seed set ratio.
Molecular basis of DFOT
-
The sketch illustrating jasmonic acid (JA) changes in rice florets during a flowering day is adapted from He et al.[25]. Depictions of DFOT in rice are illustrated, including DFOT1 and PME40/42 from Wang et al.[11], OPR7, HAN1, and JAZ7/9 from Wang et al.[26], MYB and JAR1 from Gou et al.[15], MYC2 from Zhu et al.[27], GLN2 from Xu et al.[28], AOC from Liu et al.[29], as well as miR167 and its targets ARF6/12/17/25 from Zhao et al.[30]. The data of opened spikelets in maize inbreds lines of Chang7-2 and Qi319, which are heat tolerant and sensitive, respectively, are derived from Gu[31]. The amino acid sequences of DFOT-related proteins in rice are used to search the homologs in maize using BLAST (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ), and the candidates with the highest identified percentage were screened. The expression levels of maize DFOT genes are derived from Stelpflug et al.[32] and normalized by Z-score. The original data of protein abundance of maize lodicules in Chang7-2 and Qi319 under control (32/22 °C) and heat stress (40/30 °C) are from Liu et al.[21]. The ratio of protein abundance of individual inbred lines between the control and heat is calculated. The amino acids about OsDFOT1 (Os01g0611000/LOC_Os01g42520) were used for BLAST across plants. The phylogenetic tree was constructed using MEGA via the neighbor-joining statistical method with 500 bootstrap replications.Statistics
-
Temperature changes in degrees and duration were expressed as:
$ \Delta \mathrm{T}\mathrm{m}=\mathrm{T}\mathrm{m}\mathrm{a}\mathrm{x}\;\left(\mathrm{y}\mathrm{e}\mathrm{a}\mathrm{r}\_b\right)-\mathrm{T}\mathrm{m}\mathrm{a}\mathrm{x}\mathrm{ }\;(\mathrm{y}\mathrm{e}\mathrm{a}\mathrm{r}\_a) $ Where ΔTm is the annual increase in the daily maximum temperature (Tmax) during the hottest season between year_a and year_b.
$ \mathrm{Rate}_{increase}=\dfrac{\Delta Tm}{\Delta T} $ Where Rateincrease is the annual rate of increase in Tmax during the period from year_a to year_b.
$ \Delta D=Duration\;\left(year\_a\right)-Duration\;(year\_b) $ Where ΔD is the extension of daily high temperature duration in the hottest season between year_b and year_a.
$ \mathrm{Rate}_{duration}=\dfrac{\Delta D}{\Delta T} $ Where Rateduration is the annual rate of daily high temperature duration in the hottest season between year_a to year_b.
Based on results from temperature-controlled experiments in maize, rice, sorghum, and pearl millet worldwide, the relationships between their seed set and temperatures were fitted, respectively, using linear and non-linear [quadratic and generalized additive models (GAM)] regressions. The Akaike Information Criterion (AIC) was used to decide the best model and associated thresholds[33,34]. The best model has the lowest AIC value. The threshold might only manifest when non-linear regressions provide a better fit to the data. We found variables were better fit by either quadratic or GAM models than by linear models, which confirmed the existence of temperature thresholds. There were three linear regressions, including segmented, step, and stegmented models that can determine the thresholds. Based on AIC values and data distribution, we fitted the regression manually using a segmented model. Bootstrap analysis, based on 200 samplings per database, was conducted on linear regressions to determine whether thresholds significantly affected slopes[35]. We then calculated changes in slope before and after the identified thresholds for each crop. All codes used for analysis and figure creation are on Figshare at doi:
10.6084/m9.figshare.26954419.v1 .One-way ANOVA analysis was performed to compare the optimum temperature for flowering among different plant species groups using SPSS 22 (SPSS Inc., Chicago, IL, USA). Student's t-test was used for comparisons of DFOT between heat resistant inbred line Chang7-2 and heat sensitive inbred line Qi319. A p-value less than 0.05 was considered statistically significant.
-
The Earth is experiencing unprecedented warming, and the year 2023 marked the warmest in the 174-year observational record since recordkeeping began in 1880 (Fig. 1a). In July, August, and September of 2023, global temperatures exceeded the long-term average by more than 1.0 °C for the first time in NOAA's recorded history[36]. During these months, many crops, such as maize and rice, flower and set seed in their major global producing regions[37,38], often coinciding with heatwaves. In this study, our focus was on the hottest months of the year across various global croplands, examining hourly temperature fluctuations within a single day of those months over the past 20 years (Fig. 1b, c). We selected 10 sites each in Asia, Europe, Africa, North America, and South America. These sites are evenly distributed across croplands (Fig. 1b). In this study, 29 °C was regarded as critical temperature level examining duration of midday heat episodes[17]. During the hot season, the daily maximum temperature (Tmax) rapidly increased by 1.25 °C from 2004 to 2023 with a mean rate of 0.062 °C/year (Fig. 1c). While globally, the duration of midday heat episodes extended by 1.08 h in the past 20 years with an average rate of 0.05 h/year. The average Tmax during the hottest month in the croplands of Africa ranked first (Tmax = 34.0 °C in 2023), followed by Asia (Tmax = 32.6 °C; Fig. 1b). North America had the third highest Tmax, but had the highest growth rate in Tmax (0.154 °C/year). North America also had the fastest extending rate in midday heat episodes over the past 20 years (0.13 h/year), closely followed by Africa (0.094 h/year), more than twice faster than that in South America, Asia, and Europe. Croplands of Europe had the lowest Tmax and the slowest growth rate and extending rate in Tmax during the hot season.
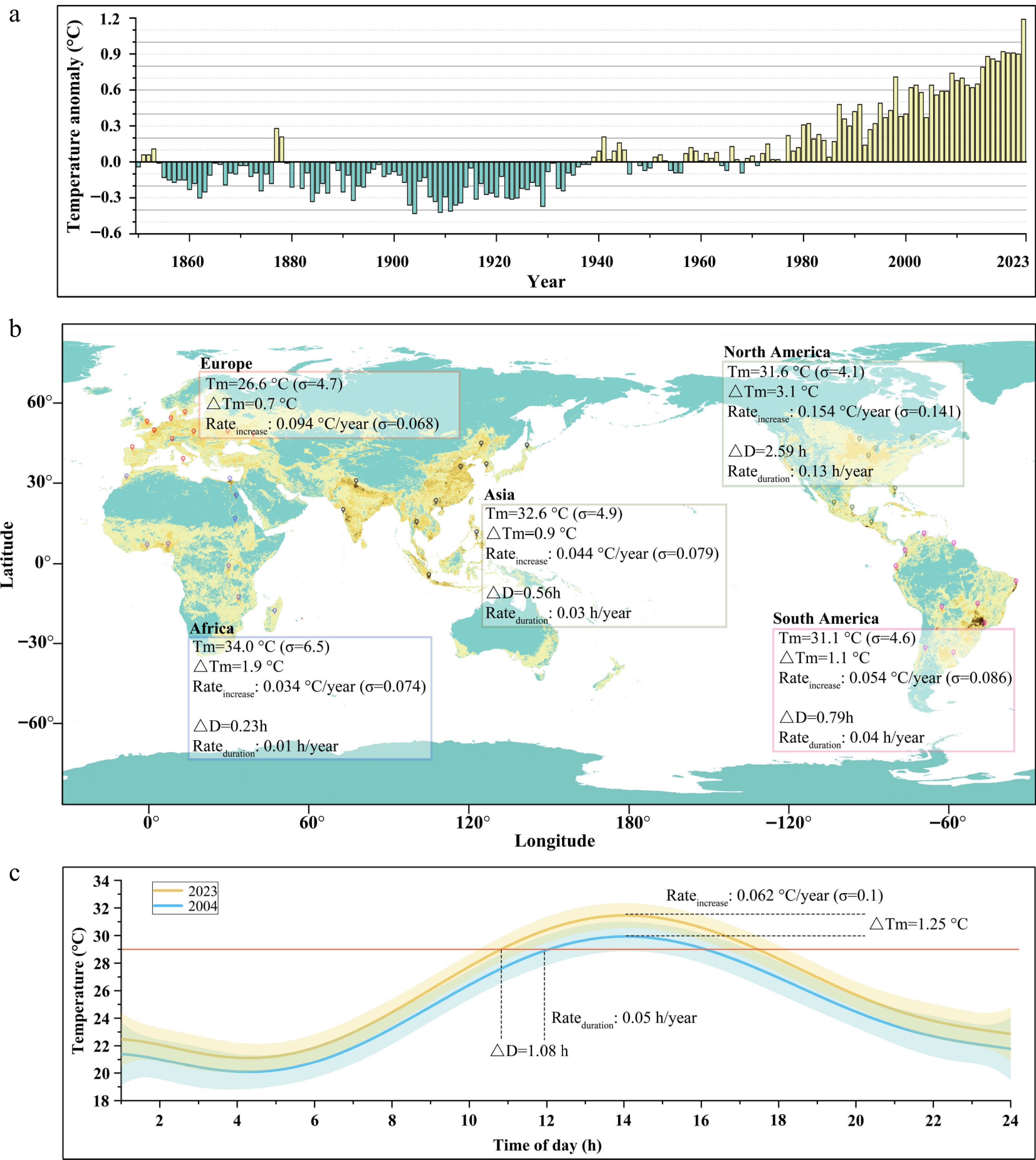
Figure 1.
Global temporal and spatial patterns of changes in temperature. (a) Anomalies of global land annual temperature with respect to the 1901−2000 average, data from NOAA (2024) (
www.ncei.noaa.gov/access/monitoring/monthly-report/global/202313 ). (b) Temperature changes in croplands of Asia, Europe, Africa, North America, and South America from 2004 to 2023. There are 10 sites selected in each continent that are located at the major croplands. Tm refers to the average daily maximum temperature (Tmax) in the hottest months across 10 sites in each region. σ represents the standard deviation of the average maximum temperature of 10 sites. ΔTm indicates the increase in the average Tmax from 2004 to 2023, and Rateincrease represents the rate of annual increase in the average Tmax. ΔD indicates the average extension of midday heat episodes in the hottest months from 2004 to 2023. Due to many crops considered in this study, 29 °C was regarded as critical temperature level examining duration of midday heat episodes[17]. Rateduration indicates the annual extension rate of midday heat episode in the hottest months. Since the average Tmax did not reach 29 °C in Europe, there was neither ΔD nor Rateduration in this continent. Background color in the map represents crop production[39]. (c) Changes of global average Tmax in the hottest months from 2023 to 2004. Data in (b) and (c) are derived from NASA POWER (2024) (https://power.larc.nasa.gov/data-access-viewer/ ). The shading in (c) shows the 95% confidence interval.DFOT in flowering plant species
-
The high species richness of flowering plants and their rapid diversification are often referred to as 'Darwin's second abominable mystery'. In these flowering plant species, DFOT exhibits diverse characteristics to adapt to complex growing environments (Fig. 2). We collected detailed information about the floret opening times of 102 flowering plant species from published articles, spanning from early morning (5:00) to the next early morning (3:00) (Fig. 2a; Supplementary Table S1). Based on DFOT, we classified these plant species into three groups: early-morning flowering, midday and afternoon flowering, and night flowering groups. Each group includes both dicots and monocots. To examine the responses of these groups to temperature during floret opening, we collected their optimum temperatures for floret opening. There was no significant difference in the mean optimum temperature for floret opening between the groups, but there was considerable variation within each group, likely attributable to their differences in the floret structure (Fig. 2b). To reduce/avoid confounding effects of the floret structure, we further compared optimum temperature for floret opening in three groups that are only comprised of cereal plants (Fig. 2c). The mean optimum temperatures of each group are 26.1, 22.3, and 29.3 °C, respectively, with significant differences. We selected five representative cereal crops for comparison. Maize and indica rice are classified as morning-flowering crops, japonica rice as midday and afternoon flowering crop, and sorghum and pearl millet as night-flowering crops. According to previous reports and data calculation[40−47], the respective optimum flowering temperatures of these five crops are approximately 30.5, 28.3, 24.3, 31, and 33.75 °C, respectively (Fig. 2d).
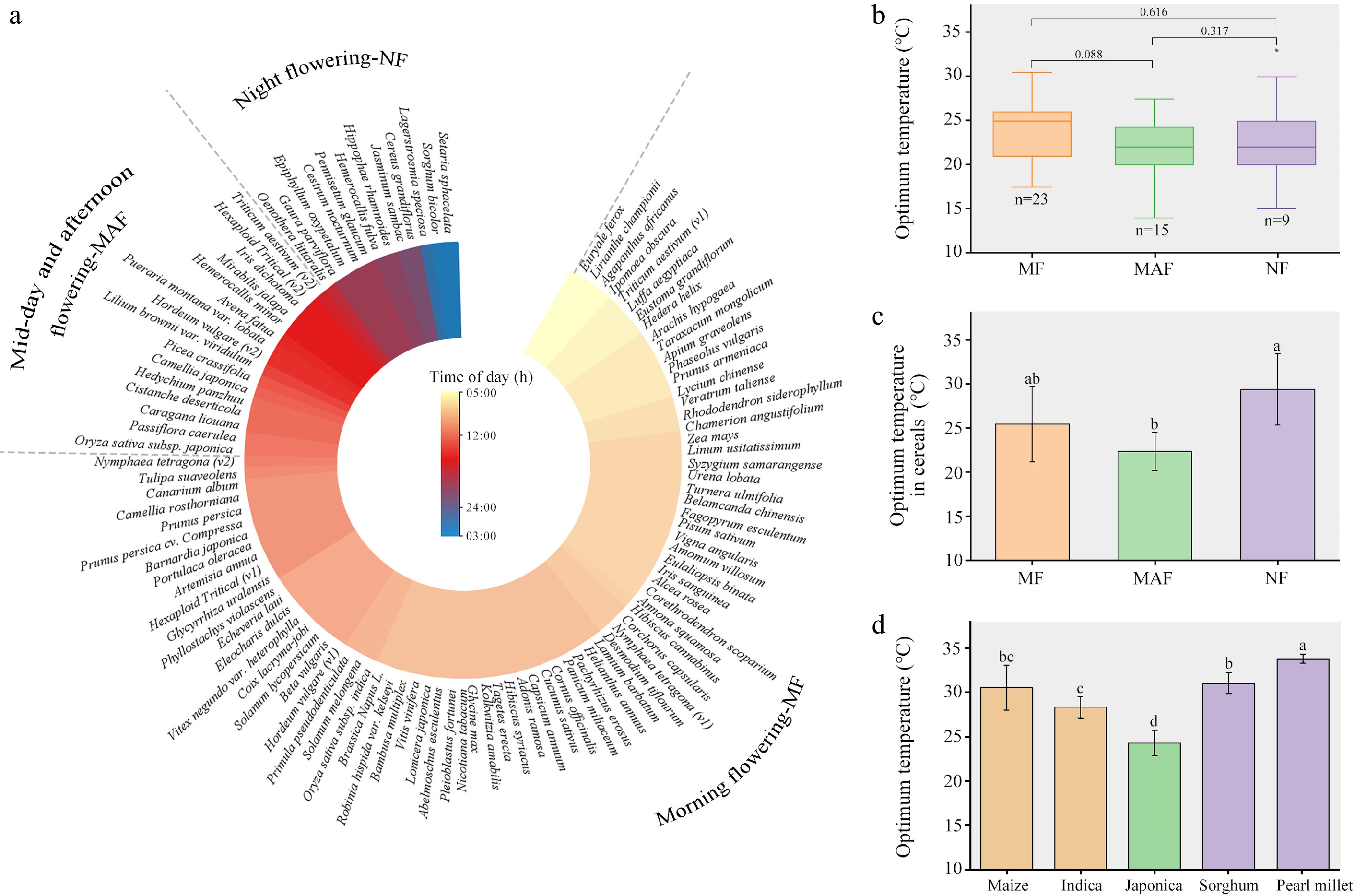
Figure 2.
Diurnal flowering time of plant species and the optimum temperature for flowering. (a) Diurnal floret opening time (DFOT) of 102 plant species. Based on DFOT, these plants are divided into morning flowering (MF, 5:00–11:00), midday and afternoon flowering (MAF, 11:00–17:00), and night flowering (NF, 18:00–3:00). (b) Optimum temperature for floret opening of three groups. The black line in each box represents the mean value. (c) Optimum temperature for floret opening in three groups that only contain cereal plants. (d) Respective optimum temperature for floret opening of maize, indica, japonica, sorghum, and pearl millet. The black line in each box in (b) and (c) represents the mean value. The error bar indicates standard error of the mean. The comparisons in optimum temperature between plant species groups were performed using one-way ANOVA analysis. Details about the diurnal flowering time and optimum temperatures for flowering of plant species are shown in Supplementary Table S1.
DFOT and high temperatures in cereal crops
-
Given their similarity in the floret structure and the frequency of exposure to heat stress events during flowering, we focused on cereal crops, including maize, rice, sorghum, and pearl millet. Maize tassels and indica rice proceed with floret opening in the morning, while japonica rice opens florets in the midday. Sorghum and pearl millet open their florets either at night or in the early morning. We collected published results concerning the effects of high-temperature stress on seed set in these four crops globally and identified their respective temperature thresholds (Fig. 3a1–a3; Supplementary Table S2).
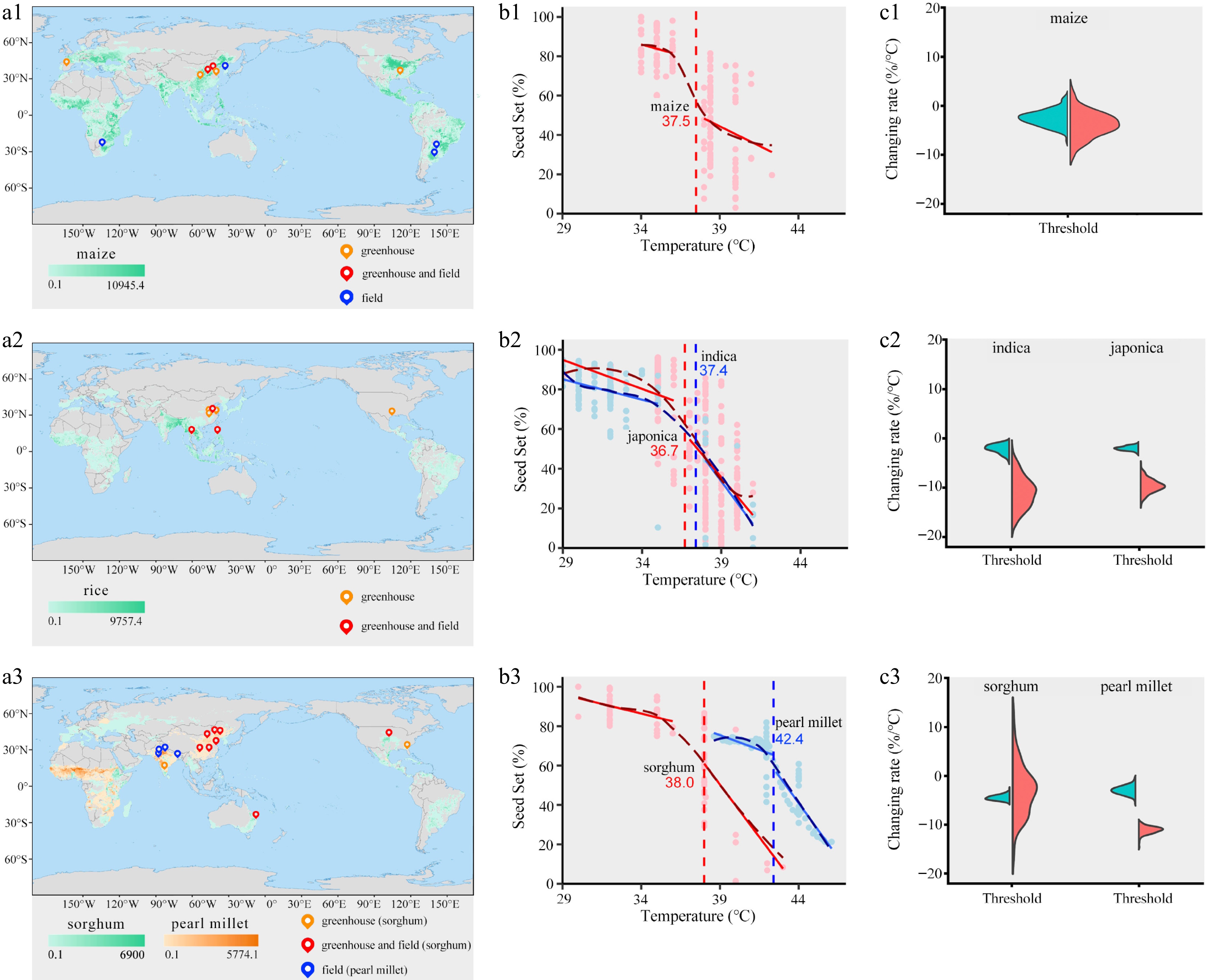
Figure 3.
Fitted high-temperature thresholds for cops. (a1)–(a3) Distribution of temperature-controlled experiments across global croplands in maize, rice, sorghum, and pearl millet, respectively. Background color in the map represents harvest area[39]. The colors of location points in the map represent experimental types (greenhouse, field, or both). (b1)–(b3) Curved dashed lines and solid lines represent the smoothed trend fitted by a generalized additive model and the linear fits, respectively. Inset numbers and the vertical dashed lines both describe the high temperature threshold. (c1)–(c3) Violin diagrams show bootstrapped values of the slope of the two regressions existing each side of each high temperature threshold (blue, before the threshold; red, after the threshold). Experimental data that are collected from peer-reviewed articles are shown in Supplementary Table S2. Details about model comparisons are shown in Supplementary Table S3.
We fitted linear and non-linear (quadratic and generalized additive models [GAM]) regressions to the relationships between seed set and temperatures, and used the Akaike Information Criterion (AIC) to decide the best model and associated thresholds. The most likely model has the lowest AIC value (Supplementary Table S3). Based on AIC values and data distribution, we fitted the regression manually using a segmented model (Fig. 3b1–b3). Bootstrap analysis was conducted on linear regressions to determine whether thresholds significantly affected slopes. The temperature thresholds for seed set in maize, indica, and japonica were 37.5, 37.4, and 36.7 °C, respectively (Fig. 3b1, b2). Sorghum and pearl millet exhibited higher high-temperature thresholds (38 and 42.4 °C, respectively; Fig. 3b3). When temperatures exceeded the thresholds, the decline in seed set ratio was slower in sorghum compared to maize and rice. Pearl millet showed a similar pattern to rice, possibly because it has a relatively higher threshold, resulting in a rapid decrease in seed set once the temperature exceeded the threshold (Fig. 3c1–c3). Particularly, as different rice subspecies, indica and japonica had more similarities in the floret structure, but the former showed a higher temperature threshold (37.4 °C vs 36.7 °C).
Early-morning flowering (EMF) and heat escape
-
Several experiments have demonstrated that rice and wheat varieties with EMF trait exhibit enhanced heat escape during the flowering stage (Fig. 4a, b; Supplementary Table S4). We re-analyzed the data reported by Aiqing et al.[23] and Ishimaru et al.[22] to explore EMF and heat escape in wheat and rice. Consequently, on average, across various studies, the mean seed set ratio of EMF varieties in wheat and rice was comparable to that of their respective controls under normal conditions. However, the seed set ratio was much higher in EMF wheat and rice than the control varieties, respectively, under high-temperature condition. The difference in seed set between EMF and control became larger with increased temperatures (Fig. 4b). In maize, results of varieties with the EMF trait have not been released, but we re-examined a previous study[24] and found that advancing hand-pollination time to early morning significantly increased the seed set ratio under heat stress (Supplementary Table S5). When pollination time was delayed after 9:00, seed set ratio significantly decreased from 80% to less than 60% under 40/30 °C (maximum daily temperature/minimum temperature); however, under control conditions (32/22 °C), the seed set ratio remained above 80% even under the treatment of delayed pollination time (Fig. 4c). In this temperature-controlled experiment, abundant fresh pollen grains were manually applied to maize silks. Therefore, pollen viability and fertilization are identified as the two critical factors determining seed set. These pieces of evidence indicate that the EMF trait is an effective strategy to escape heat stress (Fig. 4d). Additionally, given the global decline of pollinators[48,49], plant EMF trait could enhance heat adaptation during flowering in insect-pollinated species, because pollinators favor cooler early morning hours on hot days.
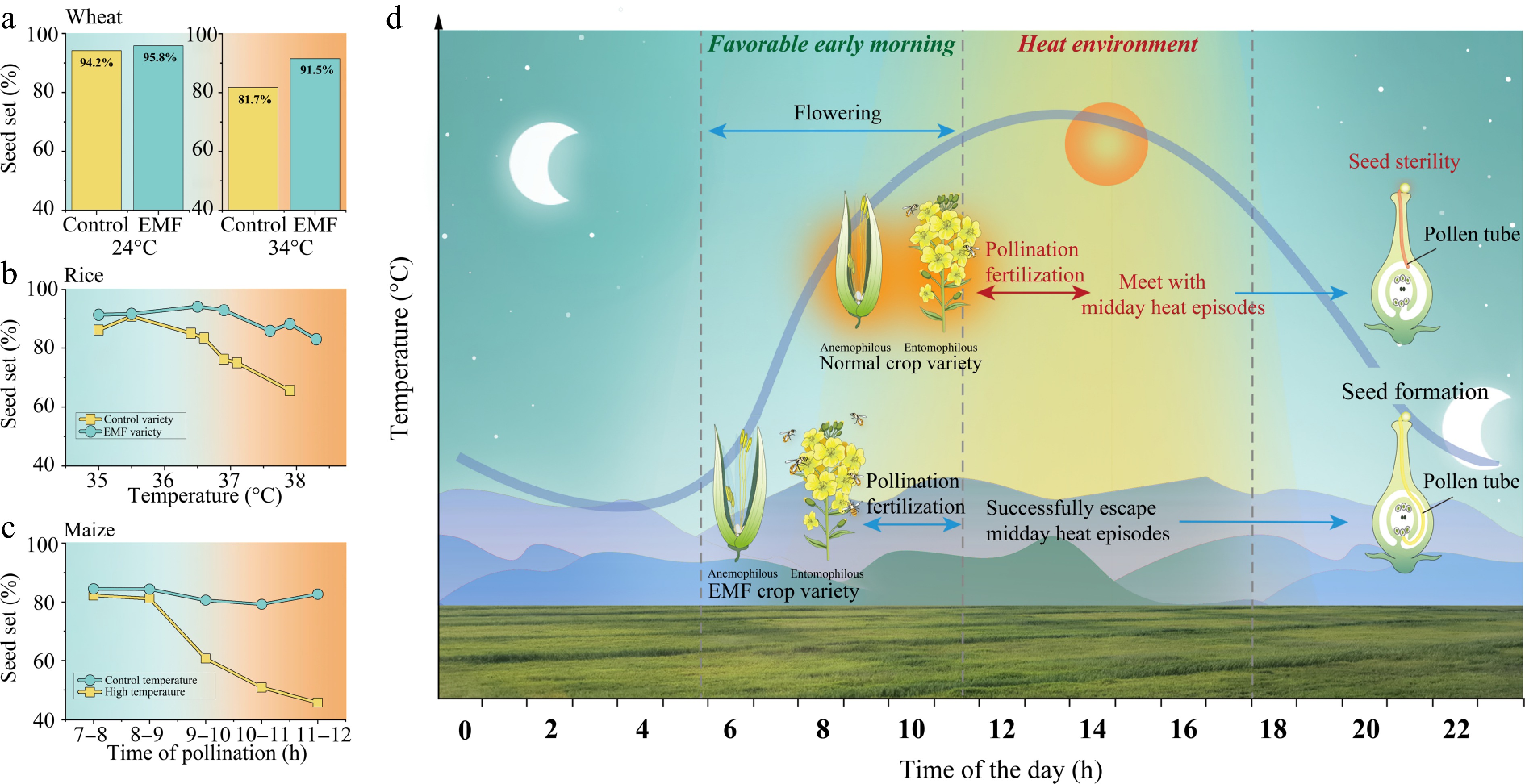
Figure 4.
Heat escape of early-morning flowering (EMF) crops. (a) Seed set ratio of EMF and control varieties in wheat under 24 and 34 °C during flowering. (b) Seed set ratio of EMF and control varieties in rice under different temperature levels. (c) Seed set ratio as effect of pollination time that ranges from 7:00–8:00 to 11:00–12:00 in one-hour interval under control (30/20 °C; daily maximum temperature/daily minimum temperature) and high temperature (40/30 °C). (d) Illustration of heat resistance of EMF crops to high temperature. Experimental data that are collected from previous reports[18–20] and detailed data are shown in Supplementary Tables S4 and S5.
Molecular basis of DFOT
-
The cereal spikelet typically consists of a pair of glumes that enclose florets, and lodicules are floral organs specific to grasses[14]. Taking rice as an example, the two lodicules are located at the base of florets, and are composed of large thin-wall cells and vascular bundles (Fig. 5a1–a3). In rice spikelets, levels of JAs increased as flowering approaches, peaking at flowering and subsequently decreasing (Fig. 5b1). OsAOC, OsOPR7, OsMYC2, and OsMYB8-OsJAR1 positively regulated JA biosynthesis, whereas knockout of OsHAN1 could reduce JA levels in the spikelets (Fig. 5b2). DFOT1 negatively regulated DFOT in rice; however, loss-of-function of the genes in pectin methylesterase (PME) family, such as PME40 and PME42, led to an earlier floret opening (Fig. 5b2). DFOT1 interacted with GLN2 to modulate the cell wall components in lodicules (Fig. 5b2). Activated by JA signals, OsMYC2 interacted with OsJAZ9, a transcriptional repressor of the JA pathway, forming an OsMYC2-JA feedback loop that regulates DFOT through cell wall loosening (Fig. 5b2). Enhanced JA signals resulting from the knockout of OsJAZ7 and OsJAZ9 increased water potential in lodicules, attributed to higher levels of potassium ions and soluble sugars (Fig. 5b2), facilitating water absorption of lodicule expansion. Additionally, overexpression of OsmiR167d suppressed auxin response factors (ARFs) of ARF6/12/17/25 and narrowed lodicule width (Fig. 5b2).
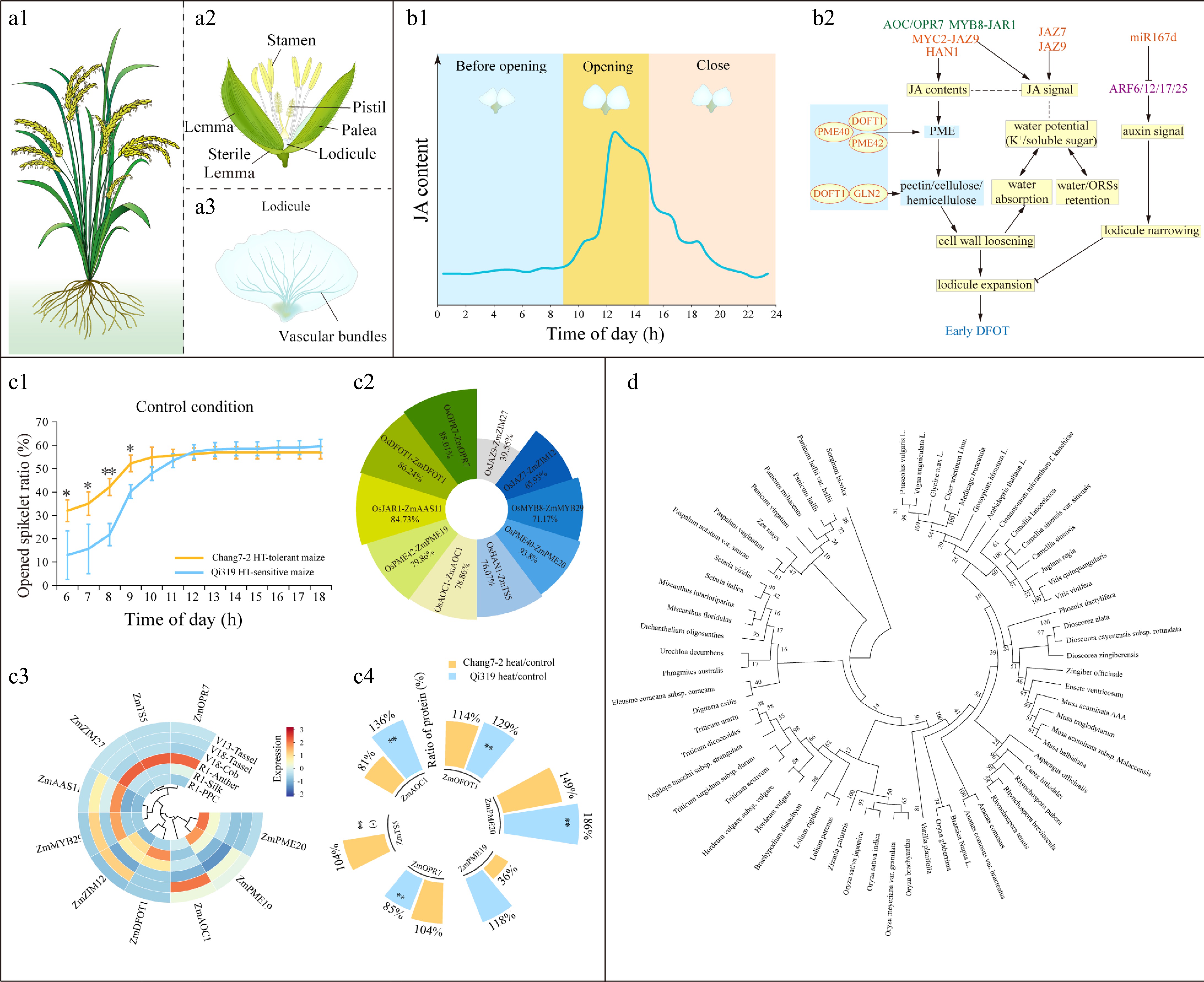
Figure 5.
Molecular mechanisms of diurnal floret opening time (DFOT) in cereal crops. (a1)–(a3) Illustration of flowering plant, spikelet, and lodicules of rice. (b1), (b2) Jasmonic acid (JA) contents in a flowering day and mechanism of JAs for early DFOT in rice[25]. The blue and yellow colors indicate a suppression (decrease) and enhance (increase), respectively, in protein interaction, enzyme activity, and metabolite accumulation. Triangle arrow and t-arrow indicate positive and negative effects. Solid and dashed lines indicate the confirmed and putative interactions. Genes marked with orange and purple mean positive and negative roles in early DFOT. ORSs, osmotic regulation substances. (c) DFOT in maize in response to high-temperature stress. (c1) Ratio of opened spikelet in heat-tolerant inbred lines Chang7-2 and heat-sensitive inbred line Qi319 under high-temperature stress (40/30 °C) during flowering. (c2) Similarities of homologs involved in DFOT between rice and maize. (c3) Gene expression levels of potential DFOT genes in maize. The transcript levels are obtained from Gu[31], and normalized by Z-score for analysis. PPC, pre-pollination cob. (c4) Ratio of protein abundance in Chang7-2 and Qi319 under heat stress compared with relative controls. Raw data are obtained from Liu et al.[21]. The ratio of protein abundance in the comparison heat versus control is calculated (* p < 0.05 and ** p < 0.01). (-) means '0' of the ratio. The ratio greater than 100% means increased in the comparison; otherwise, it is decreased. (d) A phylogenetic tree of OsDFOT1 and its homologs in other plant species. OsDFOT1 mediates diurnal floret opening time in rice[11], and the amino acids of OsDFOT1 were used for BLAST (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ). The phylogenetic tree was constructed according to the amino acid sequences of OsDFOT1 and its homologs. Details about BLAST results and homolog accessions are shown in Supplementary Table S6.Unlike rice, evidence uncovering DFOT in other crops is rather rare. The ratio of opened spikelets was higher in Chang7-2 (heat resistant) compared to Qi319[31] (heat sensitive) (Fig. 5c1). The homologs of rice genes involved in DFOT were investigated in maize, revealing high similarities between the two crops (Fig. 5c2). The transcript levels of DFOT genes exhibited varying changes between maize tassels and other tissues at the silking stage (R1). The levels of ZmDFOT1 were relatively stable and lower in tassels during vegetative stages compared to R1. ZmAOC1, ZmZIM2, ZmMYB29, ZmAAS11, and ZmZIM12 in tassels increased with successive development (Fig. 5c3). To further verify the roles of DFOT genes related to heat escape in maize, their protein abundances in lodicules of Chang7-2 and Qi319 were examined (Fig. 5c4). Among the ten genes involved in maize DFOT six were significantly responsive to heat. ZmDFOT1 and ZmPME20 were induced by heat stress in both inbred lines. ZmPME19 showed decreased abundance under heat stress in Chang7-2 compared to the control condition; however, there were no significant changes in Qi319 between the control and heat groups. In addition, high-temperature stress declined protein accumulation of ZmOPR7 in the heat-sensitive inbred line Qi319. DFOT plays a pivotal role in mediating flowering rhythm via lodicule expansion in rice; the homologs of OsDFOT against other 65 plant species, including monocots and dicots, were examined, with similarity of 61%–95% (Supplementary Table S6). The phylogenetic tree indicated that OsDFOT from japonica rice is closely related to that from indica rice, but distantly related to those from sorghum and maize (Fig. 5d).
-
The Earth is experiencing unprecedented warming[50,51]. In a single day during the hot seasons, the duration of heat stress extended at an average rate of 0.05 h/year globally over the past decades (Fig. 1). Crops such as maize, rice, and sorghum undergo flowering during late spring, summer, or early autumn, inevitably exposing them to heat stress. The timing of these crucial reproductive events often coincides with extended heat episodes, threatening crop production through impaired pollen shedding, disrupted pollination, failed fertilization, and compromised grain development. Existing evidence confirms that flower fertility in many crop species is not adequately prepared to withstand heat stress driven by climate conditions[4, 6]. Advances in crop heat resistance through metabolic modifications have not kept pace with climate warming, resulting in a lag in the development and widespread use of heat-resistant crop varieties[52]. Hence, we are shifting our focus towards exploring heat escape strategies in agricultural production[22, 53]. Our results suggest that adjusting the diurnal floret opening times (DFOT) to ensure the proper timing of flower opening might help crops adapt to various types of heat stress, such as heatwaves, heat episodes, and prolonged periods of high temperatures.
Although a direct causal link between flowering time and optimum temperature remains unconfirmed, structural adaptations in floret organs, such as lodicules or petals, critically modulate flowering regulation in plants[14]. Subspecies-specific optimal temperatures for floret opening are observed (indica rice: 28.3 °C, japonica rice: 24.3 °C), correlating with distinct lodicule morphologies. Japonica exhibits larger lodicule-lemma gaps and more vascular bundles than indica[54]. In the grass plants, rice and maize share similar floral organ structures; enhancing lodicule expansion in rice makes DFOT earlier, and increasing lodicule size promotes maize flowering and improves heat resistance[21, 27]. These findings demonstrate that floral organ structure would govern flowering timing and underpin heat escape in crops.
Early-morning flowering (EMF) trait is considered an effective escape strategy in coping with heat stress in rice[22, 55], wheat[23], and maize[24]. In our study, sorghum, maize, and rice have similarities in spikelet and floret structure, but sorghum has a higher high-temperature threshold for seed setting during flowering, which may be attributed to its night-flowering habit (Fig. 2). Furthermore, as different rice subspecies, indica and japonica have more similarities in floret structure, but the former has a higher temperature threshold probably due to its earlier flowering time in the morning. These results indicate that enhancing heat escape is more feasible than improving heat tolerance when aiming to increase crop heat adaptation during flowering. DFOT is partly affected by the growing environments of plants, but the peak floret opening time remains relatively stable across different environments for a given variety of different species[22,23, 53], a clear clue, based on which we can select of or breed for heat-escape varieties. Several field experiments verified that rice and wheat varieties with EMF trait owned enhanced heat escape during the flowering stage (Fig. 4). Inspiringly, recent studies have identified several genes associated with the EMF trait in rice (Fig. 5), demonstrating promising potential for enhancing heat escape. Based on these EMF genes, several homologs have been identified in plant species, which implies that it is feasible to breed EMF varieties. Inspired by the EMF trait, drone-assisted pollination in the early morning into crop management practices under heat stress during the flowering stage has been integrated, and it significantly increases crop grain yield. Furthermore, the EMF trait may enhance the heat escape of entomophilous plants during flowering, as pollinators tend to be more active in the cooler early morning hours on hot days. Shifting flowering time shows promise for heat stress avoidance, while field implementation should account for early morning humidity, which may trigger pollen clumping and compromises fertilization.
-
Plant kingdoms have long demonstrated climate-smart strategies to adapt to heat stress, exemplified by traits like night flowering in sorghum and pearl millet. To date, the molecular mechanism underlying diurnal floret opening in these crops remains largely unexplored, and the key genes responsible for the night-flowering trait in sorghum and pearl millet have yet to be identified. Moreover, the causal links between flowering time and critical temperature thresholds, as well as the molecular mechanisms through which the circadian clock confers heat adaptation in crops by regulating flowering time, require further investigation. Collectively, there are innovative strategies that aim at enhancing crop heat escape through precise improvements in variety breeding and crop management, which are particularly crucial in a warming climate.
This study was supported by the National Key Research & Development Program of China (2023YFD2303304), the National Science Foundation of China (32272214), the 2115 Talent Development Program of China Agricultural University, the Pinduoduo-China Agricultural University Research Fund (PC2023B02006), and the S&T Program of Hebei (225A9908D).
-
The authors confirm their contributions to the paper as follows: study conception and design: Song R, Wang X, Mao F, Huang S; graphs and draft manuscript preparation: Song R, Wang M, Yao Q, Wang X, Huang S; materials visualization: Wang X, Mao F, Han Y; manucript revision: Wang P, Liu P, Gao Y, Wang X, Mao F, Huang S. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are openly available in figshare at https://doi.org/10.6084/m9.figshare.26954419.v1.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Ruoyu Song, Minxing Wang, Qian Yao
- Supplementary Table S1 Diurnal floret opening time and optimum temperatures for floret opening of different plant species.
- Supplementary Table S2 Seed set ratio of maize, indica rice, japonica rice, sorghum, and pearl millet under different temperature levels during flowering.
- Supplementary Table S3 Comparisons of models used to identify high temperature thresholds for seed set in different crops.
- Supplementary Table S4 Seed set ratio of control and early-morning flowering (EMF) varieties under different temperature levels during flowering stage in wheat and rice.
- Supplementary Table S5 Seed set ratio of maize as effect of pollination time and temperature treatments on the 4th day after anthesis.
- Supplementary Table S6 The similarity of rice diurnal floret opening time against different plant species.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Song R, Wang M, Yao Q, Han Y, Liu P, et al. 2025. Shifting flower timing to beat the heat of warming climate in crops. Seed Biology 4: e013 doi: 10.48130/seedbio-0025-0013 

Shifting flower timing to beat the heat of warming climate in crops
- Received: 16 May 2025
- Revised: 29 July 2025
- Accepted: 05 August 2025
- Published online: 28 August 2025
Abstract: Climate-induced heat stress is intensifying across a spectrum of timescales, spanning from annual and seasonal cycles to monthly and even daily fluctuations. The development of heat-resistant crops is imperative to safeguard food security in the face of a warming climate. This study investigates the warming trend in global croplands, explores a heat escape strategy and its underlying mechanism, and evaluates its efficacy in mitigating heat stress. We analyzed the global-scale warming trend since 1850, compared the diurnal variation of temperature during the hottest months across global croplands over the past 20 years, collected diurnal floret opening time (DFOT) data from 102 plant species, and summarized the molecular mechanism underlying DFOT regulation in cereal crops. During the hottest season globally, the daily maximum temperature increased at an average rate of 0.062 °C per year, while the duration of mid-day heat episodes in global croplands extended by an average of 0.05 h per year. In the plant kingdom, species like sorghum and pearl millet escape midday heat during hot seasons by shifting their floret opening time to early morning, thereby effectively coping with prolonged heat episodes. Multiple pieces of evidence confirm that the early-morning flowering trait significantly enhances heat resistance in rice, wheat, and maize. The molecular mechanism of the early-morning flowering trait is relatively clear in rice, based on which, we identified homologous genes in more than 60 plant species, which provides insights into the potential for widely enhancing crop heat escape in a warming climate.
-
Key words:
- Heat stress /
- Plant kingdom /
- Diurnal floret opening time /
- Early-morning flowering /
- Grain yield