









-
The SCF (SKP1-CUL1-F-box protein) E3 ubiquitin-ligase complexes, in combination with F-box proteins TRANSPORT INHIBITOR RESPONSE 1/AUXIN-SIGNALING F-BOX (TIR1/AFB), play a critical role in the bio-process of auxin signal transduction in plants, involving many auxin-mediated responses through transcriptional regulation[1]. The substrate specificity of SCFTIR1/AFB is conferred by the interchangeable F-box protein subunits of the TIR1/AFB family, functioning as auxin coreceptors that interact with the auxin/indole-3-acetic acid (Aux/IAA) transcriptional regulators[2,3]. Auxin acts as a 'molecular glue', strengthening its interaction with auxin receptors TIR1/AFBs, degrading Aux/IAA, releasing auxin-responsive factors (ARFs), and activating the expression of genes involved in auxin-dependent processes[4,5].
Characterized by the structures of F-Box domain and a transport inhibitor response 1 protein domain, TIR1/AFB proteins are identified widely in planta, while phylogenetic studies have found that the TIR1/AFB proteins are conserved across land plant lineages and can be categorized into four clades: TIR1/AFB1, AFB2/AFB3, AFB4/5, and AFB6[6]. Among these TIR1/AFB proteins, AFB6 was identified in Solanaceae species such as tomato (Solanum lycopersicum) and eggplant (S. melongena) but not in Arabidopsis[7,8], while AFB2/3 proteins have not yet been identified or characterized in Solanaceae species.
The TIR1/AFB proteins mediate diverse responses to plant growth and abiotic stress, including embryogenesis, leaf development, fruit development, and salt stress[9−12]. In terms of biotic stress, TIR1/AFB mutants have also been reported with varied responses to disease infestation. The Arabidopsis quadruple mutant tir1 afb1 afb4 afb5 was observed with increased IAA levels along with higher susceptibility to Pseudomonas Syringae PtoDC3000[13]. Compared with Col-0, the afb1 and afb3 mutants of Arabidopsis showed partial resistance to Verticillium dahliae, with defense-related genes such as PR1 and PDF1.2 upregulated[14]. In addition, RNAi-mediated gene knockdown of TaTIR1 increased wheat resistance to Fusarium head blight[15].
Pepper (Capsicum annuum) is one of the most economically important vegetable crops of the Solanaceae family, traditionally cultivated and consumed worldwide both as a vegetable and as a seasoning ingredient owing to its health benefits derived from abundant and diverse bioactive compounds[16]. Despite its economic value, the pepper industry is severely threatened by bacterial wilt caused by Ralstonia solanacearum[17]. Several genes, such as CaWRKY40, have been identified for their roles in pepper's response to R. solanacearum infestation[18−22], but none of the CaTIR1/AFBs have been reported to be involved in response to R. solanacearum in pepper. Research in the pepper genome over the past decade has led to significant insights in this field, while the recent advances in the leaf-injection method for inoculating R. solanacearum and the leaf-to-whole-plant spread bioassay have enabled the accurate timing of inoculation, making it possible to investigate the in-time molecular responses in pepper during R. solanacearum infection[23−25].
In this study, a genome-wide analysis of TIR1/AFB genes in resistant and susceptible pepper varieties was performed to evaluate their expression under R. solanacearum infestation. The CaTIR1/AFBs were subsequently subjected to bioinformatic analysis, qRT-PCR experiment, transient expression in tobacco, and virus-induced gene silencing (VIGS) assays. All the results provided a theoretical basis for studying the characteristics and functions of TIR1/AFBs proteins in pepper when responding to bacterial wilt disease, and could support further research efforts utilizing these key auxin-response genes in pepper breeding.
-
Putative TIR1/AFB genes (PepperBase (
bioinformaticslab.cn )) were retrieved from the pepper genome database[24] based on BLASTP analyses with the criteria of e-value < 0.01, while the previously identified six Arabidopsis TIR1/AFB genes (At1g12820, At3g26810, At3g26980, At4g03190, At4g24390, At5g49980), four tomato TIR1/AFB genes, and five eggplant TIR1/AFB were adopted as reference queries[7,8,26]. Six putative CaTIR1/AFB genes were found (Supplementary Table S1). Further screening was done based on the kinds of Protein Family Analysis and Modeling (PF, PF18791, transport inhibitor response 1 protein domain; PF18511, F-box) (http://pfam.xfam.org/ ) in these six members. Parameters including molecular weight, amino acid length, isoelectric point, and grand average of hydropathicity (GRAVY) were used for further characterization of CaTIR1/AFB proteins through the ExPASy-ProtParam tool[27]. In silico protein subcellular localization was predicted with WoLF PSORT (https://wolfpsort.hgc.jp/ )[28]. The TBtools 'Show Genes on Chromosome' function was used to visualize chromosomal locations of all obtained TIR1/AFB genes.Phylogenetic analysis and classification of CaTIR1/AFB genes
-
Protein sequences of the CaTIR1/AFB genes in Arabidopsis, tomato, eggplant, and pepper were adopted to generate phylogenetic trees using the MEGA X (
www.megasoftware.net ) software[29]. Sequences were aligned using multiple sequence alignment and clustered by the neighbor-joining algorithm, with 1,000 bootstrap replicates.Tandem duplication events of the CaTIR1/AFBs and the synteny analysis
-
The duplicated genes were retrieved and analyzed on the TBtools platform (
https://github.com/CJ-Chen/TBtools ), using BLAST, MCScanX, and Advanced Circos features. The synteny relationships within pepper were established by retrieving collinearity files with MCScanX, followed by generating dual synteny graphics through TBtools[30].Cis-elements in the promoters of the CaTIR1/AFB genes
-
The cis-elements of CaTIR1/CaAFBs were identified from the sequences at 2 kb upstream of the start codon using the web tools in the pepper genome database (PepperBase;
www.bioinformaticslab.cn/PepperBase ), and analyzed using the PlantCARE web-based tool (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )[31], with the findings visualized by TBtools.TIR1/AFB gene structure, conserved domain, and motif analysis
-
The identification of protein-conserved domains was performed on the NCBI conserved domain online server (
www.ncbi.nlm.nih.gov ), while their motifs were analyzed using the MEME Suite (meme-suite.org )[32]. The exon and intron structural prediction of the CaTIR1/AFB gene was conducted with the Gene Structure Display Server (GSDS) (http://gsds.cbi.pku.edu.cn/ )[33], using the genomic sequences and CDS of five CaTIR1/AFB genes.Plant growth, inoculation, and RNA sample collection
-
The inbred pepper lines 'HP-19' (resistant) and 'HP-21' (susceptible) were obtained from the State Key Laboratory for Vegetable Germplasm Enhancement and Utilization of Hebei Vegetable Germplasm Resource Centre, Baoding, China, and R. solanacearum strains were provided by the Fujian Academy of Agricultural Sciences, Fuzhou, China. Pepper seedlings were prepared and inoculated at the six-leaf stage in the plant-growing facilities at Baoding University. The seedlings were grown in an incubator (PGX-350L, Ningbo Saifu Experimental Instrument Co., Ltd, China) at 37 °C with a 16/8 h light/dark photoperiod under 80% humidity. The fourth leaves were inoculated through inocula injection with 0.05 mL R. solanacearum (OD600 = 0.3) suspension in 10 mm/L MgCl2 or the MgCl2 solution as a control, and sampled at 6 h after inoculation for transcriptomic analyses. Leaf samples were collected from at least three plants and instantly frozen in liquid nitrogen, then stored at −80 °C until RNA extraction. Three technical replicates were performed for each morphotype.
RNA-seq library construction, sequencing, and analysis
-
Three grams of each leaf sample were collected for RNA extraction using poly-T oligo-attached magnetic beads, and a NEBNext® Ultra™ RNA Library Prep Kit for Illumina® was used to construct libraries (NEB, Ipswich, MA, USA). Sequencing was performed on an Illumina HiSeq 4000 platform to obtain the 150 bp paired-end reads. Differentially expressed genes (DEGs) were identified from the differential expression analysis using the DESeq R package and the hierarchical cluster analysis. The significant DEGs were filtered based on false discovery rate (FDR) ≤ 0.01 and |log2 fold change| ≥ 1 in each pairwise comparison, and a heat map of expression profiles was constructed using TBtools[34]. Each qRT-PCR analysis included three biological and four technical replicates, respectively, while the 2−ΔΔCᴛ value was determined for the calculation of gene expression[35]. The gene-specific primers for qRT-PCR, subcellular location, and VIGS are listed in Supplementary Tables S1 and S2, annotated and designed based on the pepper genome database (PepperBase;
bioinformaticslab.cn ).Subcellular localization analysis of CaTIR1C-GFP fusion proteins
-
The 35S::CaTIR1C-GFP expression vector was constructed by cloning the full-length coding sequence of CaTIR1C into the pMDC43 plasmid, which was subsequently transferred into Agrobacterium tumefaciens strain GV3101 (Weidi, Shanghai, China) for transient transformation of young tobacco leaves. The subcellular localization of CaTIR1C-GFP was examined using a confocal laser scanning microscope (FV10i, Olympus; Leica Microsystems, Germany).
Virus-induced knockdown of CaTIR1C gene expression analysis
-
In previous studies on pepper resistance to diseases, virus-induced gene silencing (VIGS) showed promising results for the validation of gene functions[36,37]. In this experiment, pTRV1 and pTRV2 VIGS vectors were adopted for gene silencing[38−40], with CaCHlH employed as the gene marker in VIGS assays. About 300 bp synthetic sequence designed by the VIGS tool (
https://vigs.solgenomics.net/ ) corresponding to part of the CaCHlH and CaTIR1C genes was selected and cloned into the pTRV2 vector. The empty vector pTRV2 was used as the control. The constructed TRV-mediated VIGS were then applied to infect one-week-old pepper seedlings, followed by 0.1 mL R. solanacearum (OD600 = 0.3) injected into the fourth leaf in six-leaf stage pepper plants. The experiment included three independent replicates and was repeated twice. -
Using the six Arabidopsis genes, four tomato genes, and five eggplant genes as reference queries[4], six putative TIR1/AFB genes were identified in pepper (Supplementary Table S3). Candidate genes containing at least one F-Box (PF18511) and one TIR1 protein domain (PF18791) were considered as 'true' TIR1/AFBs. Therefore, five genes were kept for further analysis. Predicted genes and related information were presented in Table 1.
Table 1. Predicted pepper TIR1/AFBs genes and related information.
Gene ID Instability index Amino acids GRAVY score Predicted subcellular location Aliphatic index Annotation CaT2T02g02335 37.89 583 −0.058 Nucleus 91.97 CaAFB6 CaT2T03g04037 53.39 581 −0.026 Nucleus 95.51 CaTIR1C CaT2T04g01000 42.25 646 0.051 Nucleus 91.02 CaAFB4/5 CaT2T06g00038 51.41 580 0.015 Nucleus 96.50 CaTIR1B CaT2T06g03723 47.65 658 0.047 Nucleus 98.80 CaTIR1A The average length of these CaTIR1/AFB proteins was 609 amino acids (aa), ranging from 580 aa (CaT2T06g00038) to 658 aa (CaT2T06g03723). These putative TIR1/AFB genes are located on four chromosomes, while two of them were identified on DH06 (Fig. 1).
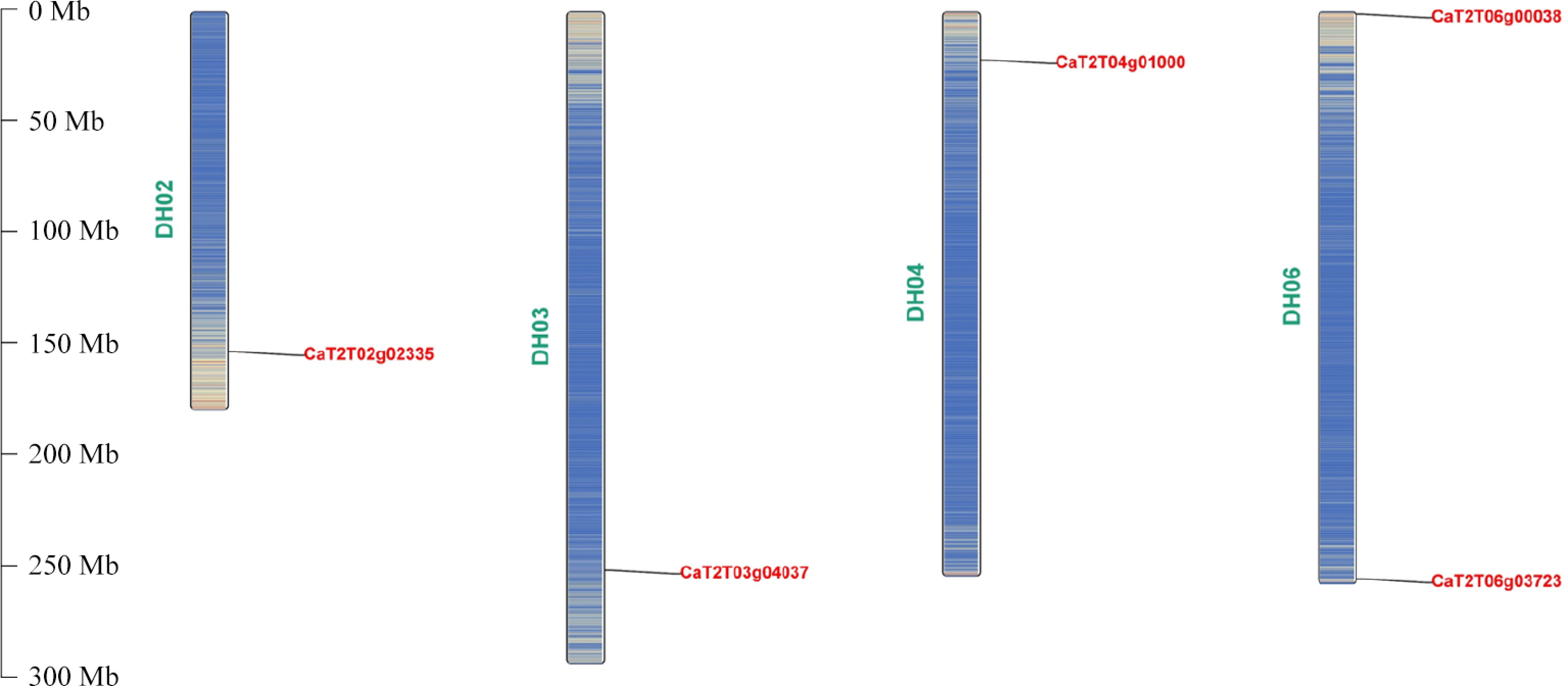
Figure 1.
Chromosomal location of the five CaTIR1/AFB genes.
Comparative phylogenetic analysis and classification of the CaTIR1/AFB gene
-
Phylogenetic studies based on the maximum likelihood algorithm were conducted to investigate the evolutionary relationship of CaTIR1/AFB genes from pepper. The phylogenetic tree was constructed using five TIR1/AFB genes from pepper, five from eggplant, five from Arabidopsis, and four from tomato, while these genes were assigned to four groups: TIR1/AFB1 (Group 1), AFB2/3 (Group 2), AFB4/5 (Group 3), and AFB6 (Group 4) (Fig. 2). The TIR1/AFB1 clade included three CaTIR1/AFB genes from pepper, while AFB4/5 and AFB6 each included one CaTIR1/AFB. Notably, the AFB6 subgroup was present in pepper, eggplant, and tomato but absent in Arabidopsis, suggesting that this subgroup evolved specifically within Solanaceae species. Furthermore, none of the genes from Solanaceae species were identified as closely related to the AFB2/3 subgroup, indicating a possible genetic loss of this subgroup in their genome.
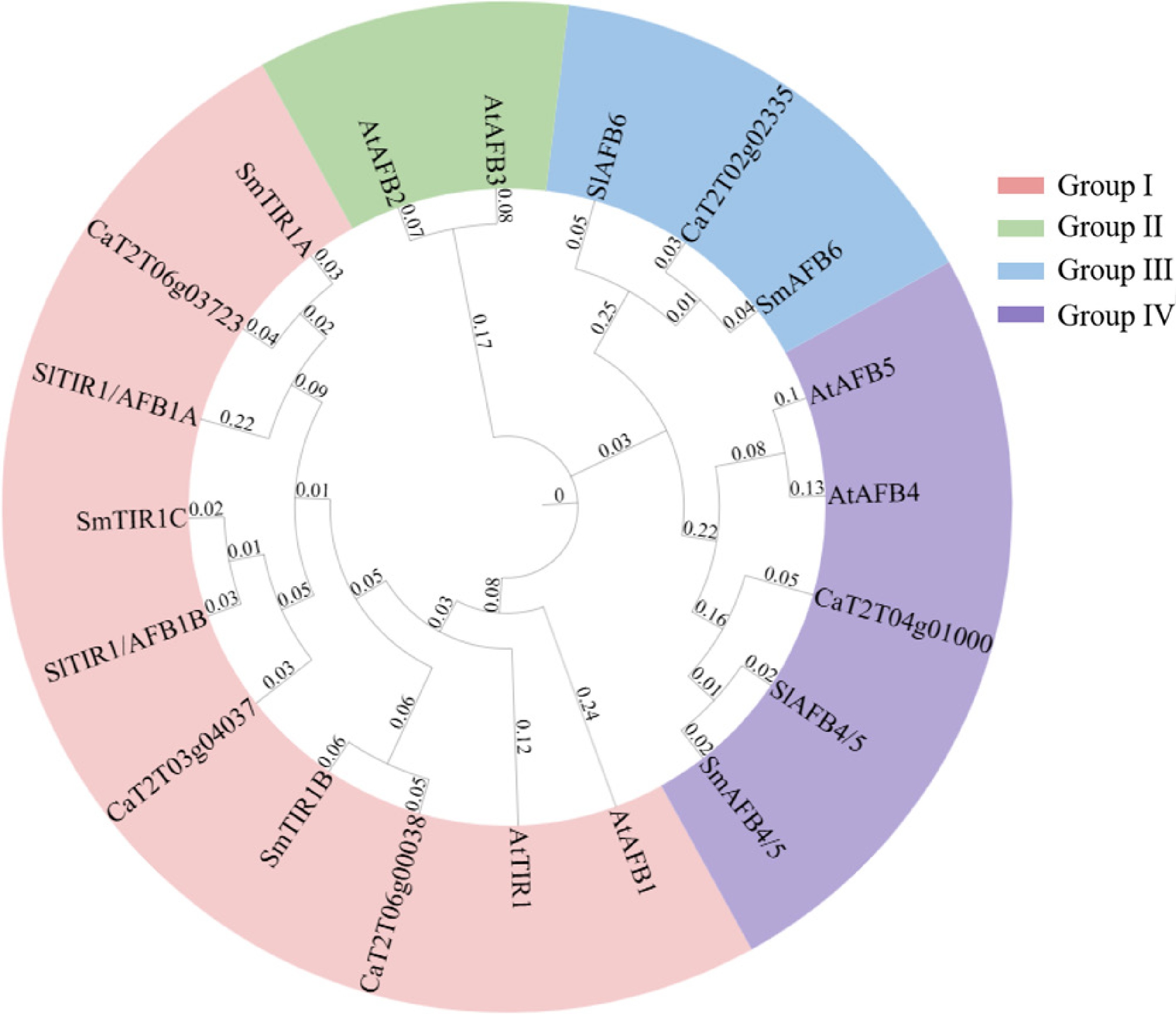
Figure 2.
Phylogenetic analyses of TIR1/AFB proteins identified in pepper, Arabidopsis, tomato, and eggplant. Protein sequences of the five TIR1/AFBs from pepper were used to construct a phylogenetic tree under the neighbor-joining algorism with 1,000 bootstrap replicates. The TIR1/AFBs were clustered into four groups (Groups I, II, III, and IV) with each highlighted in a different color.
TIR1/AFB gene structure and the conserved domain and motif analysis
-
The exon-intron structure analyses of the CaTIR1/AFB genes showed that, although these genes exhibited structural divergence, members within the same phylogenetically identified subgroup shared similar gene structures (Fig. 3a). CaTIR1B and CaTIR1C within the TIR1/AFB subgroup are notably similar, particularly in their CDSs. To further investigate functional characteristics, ten conserved motifs were identified in the CaTIR1/AFBs using the MEME algorithm. While all genes contain motifs of F-box, TIR1, and leucine-rich repeat (LRR), CaAFB6 uniquely featured a motif with an LRR_CC_2 domain (Fig. 3b). Additionally, untranslated regions (UTRs) were identified in CaTIR1C, CaAFB4/5, and CaAFB6, which could potentially differentiate their regulatory or functional roles.
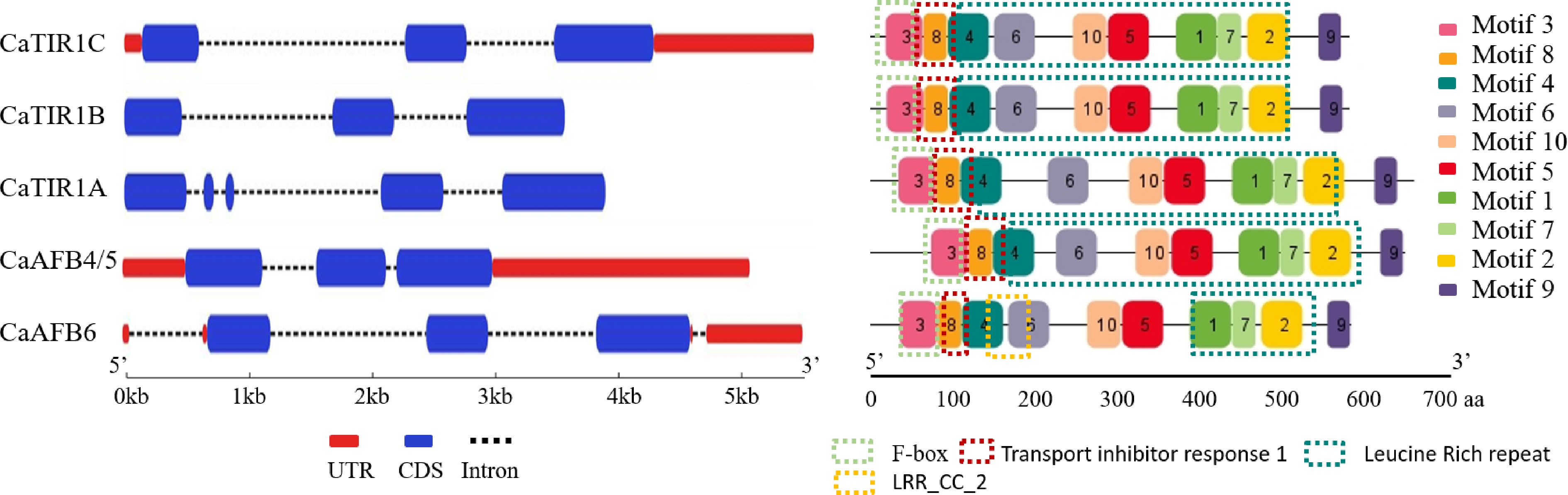
Figure 3.
Gene (left) structure and conserved domain (right) of CaTIR1/AFBs.
Duplication event in the CaTIR1/AFBs and synteny analysis
-
Tandem and segmental gene duplication in CaTIR1/AFBs was subject to BLASTP searches and analyses on TBtools. Out of five CaTIR1/AFB genes, one collinear gene pair, CaTIR1C and CaTIR1B, was identified, which is consistent with the result of the gene structure analysis (Fig. 4), indicating segmental duplication could be important in the expansion of the CaTIR1/AFB family genes.
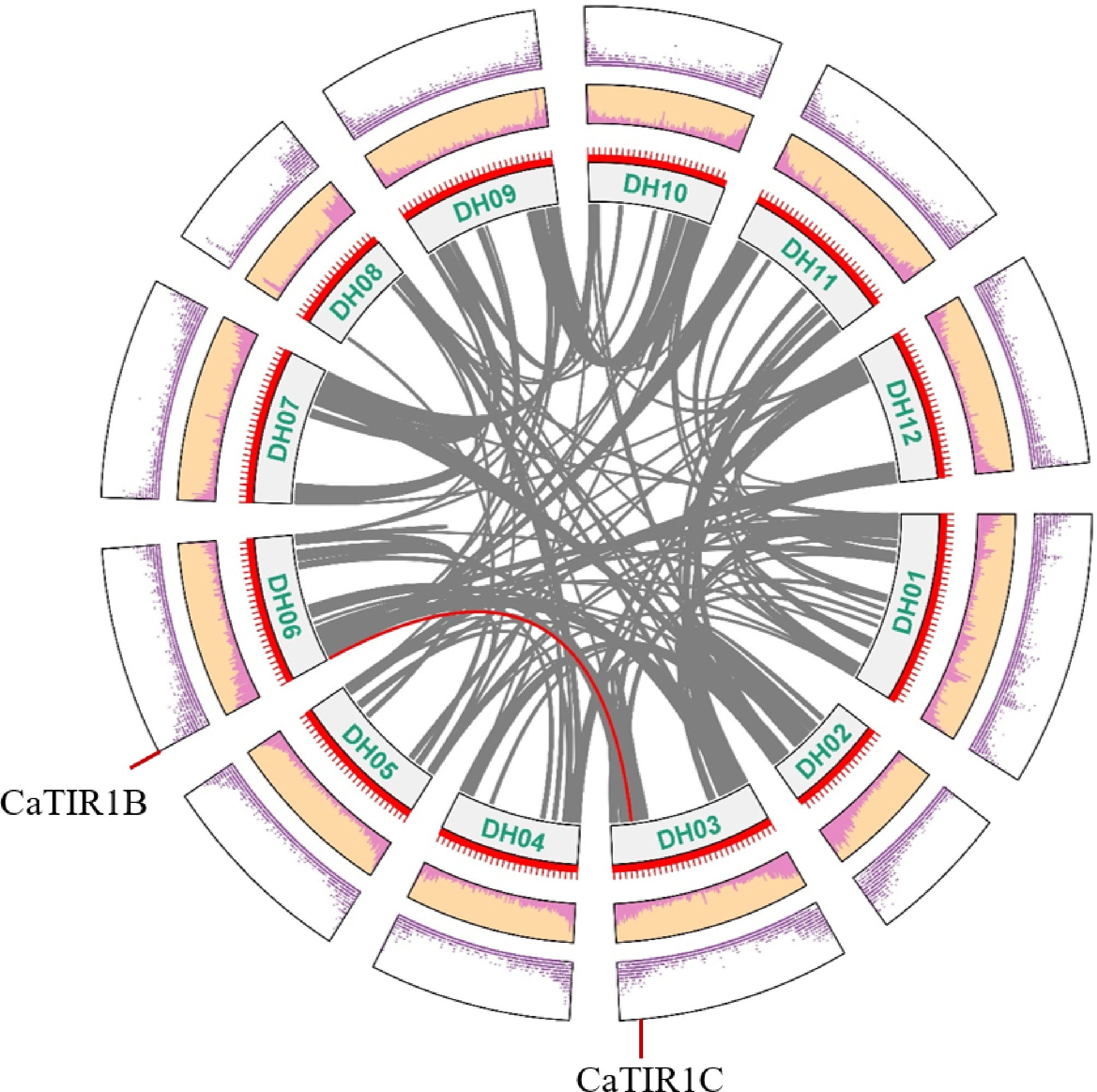
Figure 4.
Circos figure for the chromosome locations with the CaTIR1/AFBs segmental duplication links. Red lines indicate the syntenic cyclin gene pairs between the mentioned genes. Gray lines in the backdrop represent the syntenic cyclin gene pairs of other genes in pepper.
Cis-elements in the promoters of the CaTIR1/AFB genes
-
Sequences from the 2 kb upstream of the translation start sites of CaTIR1/AFBs were analyzed on the PlantCARE web tool, and the cis-elements in the promoters of these genes were identified and compared to explore the potential roles of CaTIR1/AFBs in regulatory mechanisms such as plant hormone signaling, defense, stress, and light responses. Six classes of cis-elements were identified, including responsive elements of light, drought, defense, auxin, abscisic acid (ABA), and gibberellin (GA). Light-responsive elements were identified in the upstream region of all five genes. However, auxin-responsive elements were found only in the upstream of CaAFB4/5 and CaTIR1A, while defense-responsive elements were identified only in the upstream of CaTIR1C and CaAFB6. Similarly, ABA-responsive and drought-responsive elements were only identified in the upstream of CaTIR1C and CaAFB6, respectively (Fig. 5).
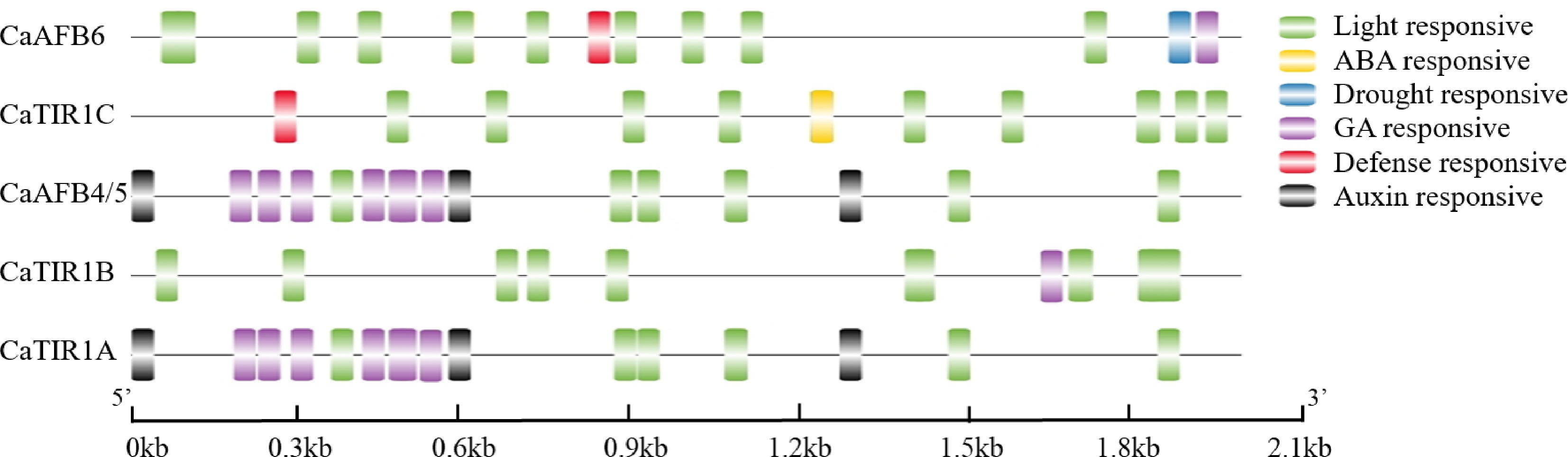
Figure 5.
Identified cis-elements in the promoters at 2 kb upstream of the CaTIR1/AFBs. Hormone, drought, light, and stress-responsive elements were identified and linked to cis-elements of the TIR1/AFBs genes.
Expression profiles of the CaTIR1/AFB genes under R. solanacearum by transcriptome analysis
-
Transcriptomic analyses showed that CaTIR1/AFB genes expressed differently in resistant and susceptible pepper varieties in response to R. solanacearum infection (Supplementary Fig. S1). All CaTIR1/AFB genes were downregulated in the resistant variety, whereas in the susceptible variety, expression levels varied among the genes (Supplementary Fig. S1; Fig. 6). Notably, CaTIR1C was significantly upregulated in the susceptible variety, suggesting that CaTIR1C may act as a positive regulator in response to R. solanacearum. These expression trends were further validated by quantitative real-time PCR (qRT-PCR), which showed general consistency with the RNA-seq data (Fig. 6). Significant expression difference of CaTIR1C between the R. solanacearum and control treatments in susceptible pepper was also confirmed (Fig. 6). The UBI quitin-like gene CaT2T06g03488 was used as the housekeeping gene for normalization.
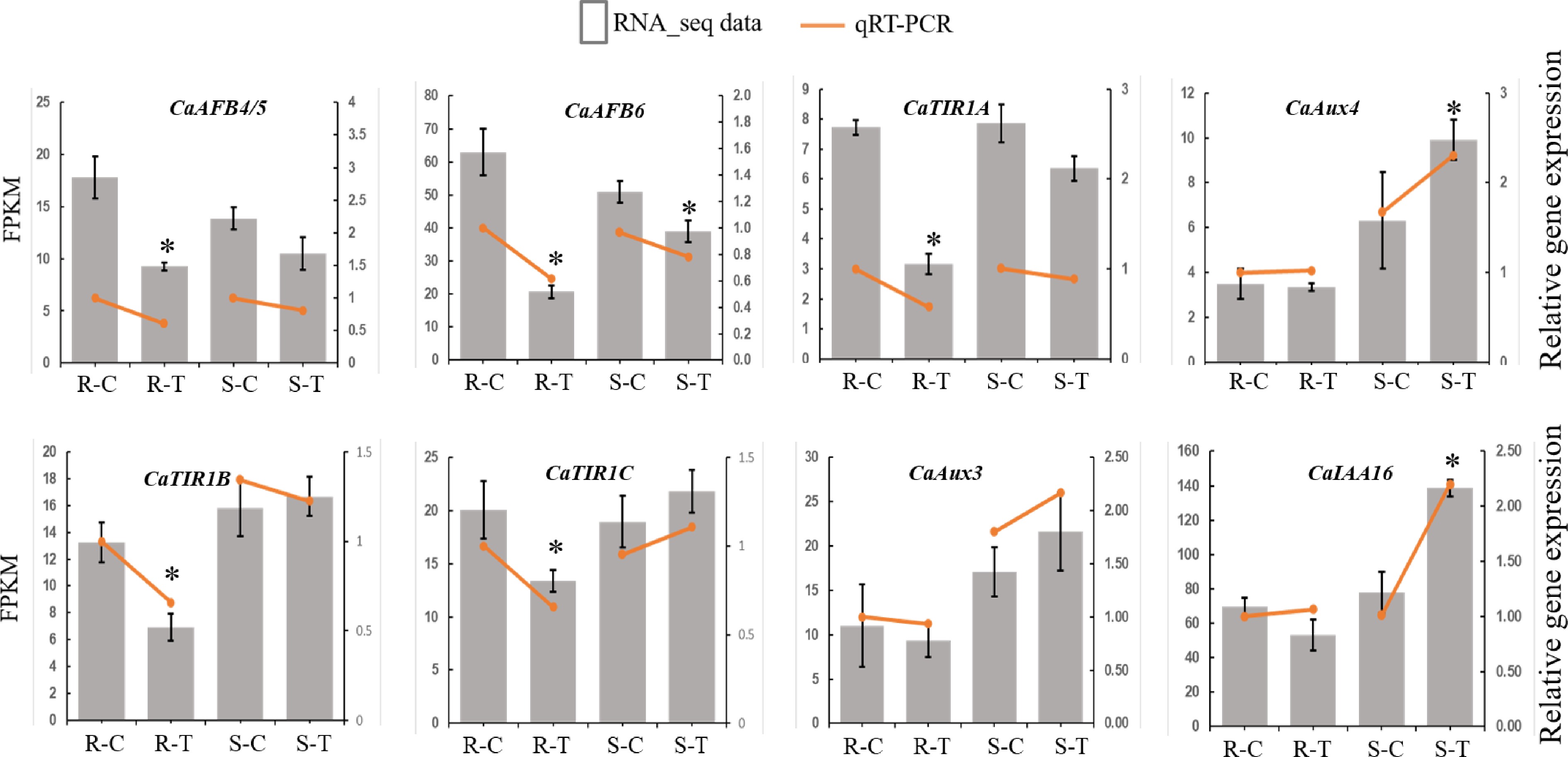
Figure 6.
Expression of CaTIR1/AFB genes under control or R. solanacearum treatment as measured by qRT-PCR (orange symbols) and RNA-seq (bars). For qRT-PCR, gene expression levels were normalized to those of CaUBI, and qRT-PCR expression levels in the resistant variety treated with control (R-C) were set to 1. Results were presented as the means ± SE of three biological replicates. * p < 0.05 (Student's t-test, qRT-PCR data).
Sub-cellular localization of CaTIR1C
-
Previous studies in Arabidopsis have shown that TIR1/AFB family members are localized to the nucleus[8]. To determine the subcellular location of CaTIR1C, the full-length coding region of CaTIR1C was cloned into pMDC43. The plasmids were transformed into Agrobacterium strain GV3101 (Weidi, Shanghai, China). The GV3101 with target plasmids was incubated with Kanamycin at 28 °C overnight in an infiltration buffer consisting of 10 mM MgCl2 (pH 5.7) and 150 mM acetosyringone. The 0.5 mL bacterial fluid (OD600 = 0.5) was injected into a tobacco leaf epidermal cell. After 48–72 h of incubation in darkness, the cells were observed using a confocal laser scanning microscope (Olympus FV10I; Leica Microsystems, Germany). The result revealed that the constructed 35S::CaTIR1C-GFP was selectively enriched in the nuclei of tobacco leaf epidermal cells (Fig. 7), suggesting that CaTIR1C may perform important nuclear functions related to its regulatory role.
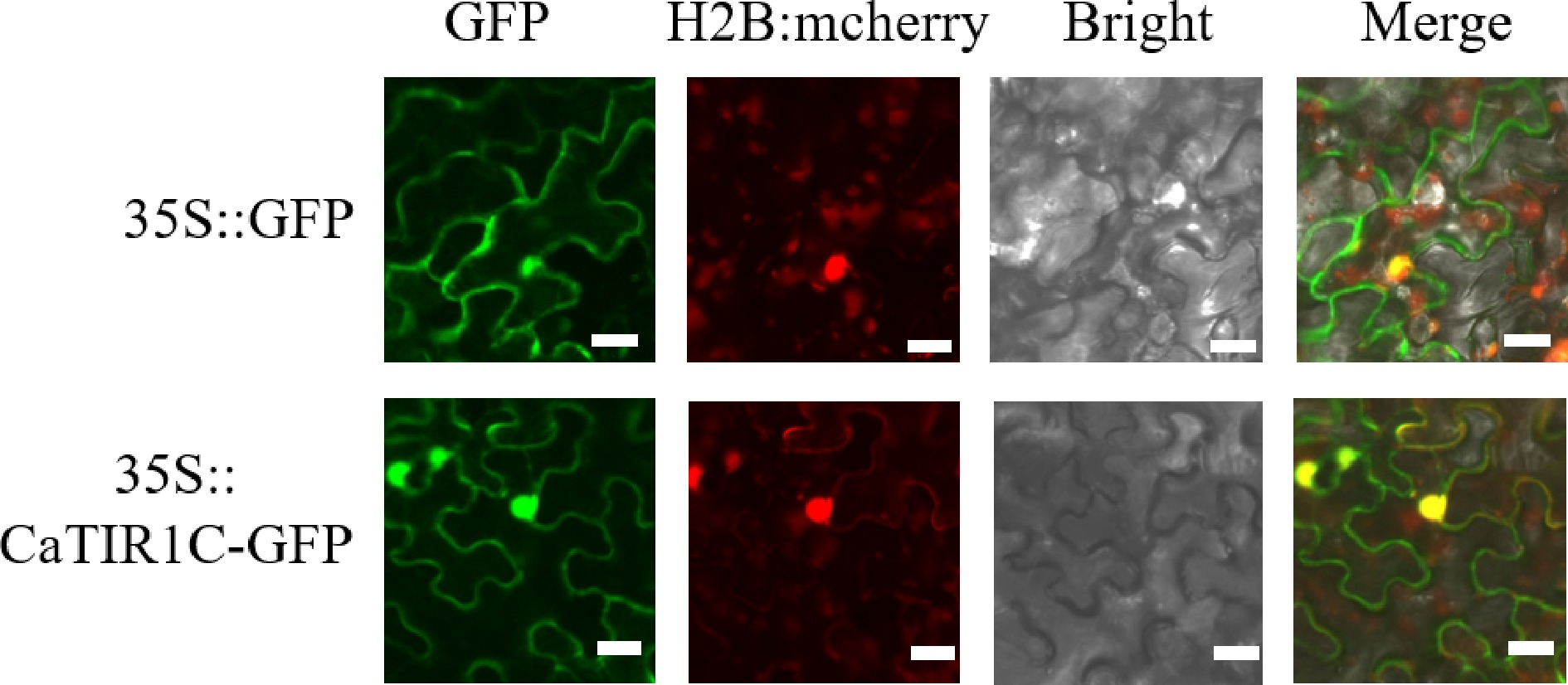
Figure 7.
Subcellular location and expression patterns of CaTIR1C in tobacco epidermal cells. The first column shows the fluorescence of the 35S::GFP, 35S::CaTIR1C-GFP proteins; the second column shows a bright-field image of the 35S::GFP, 35S::CaTIR1C-GFP proteins; the third column shows a bright-field image of the proteins; the fourth column represents an overlay of the fluorescent images. Bar = 20 μm.
Knockdown of CaTIR1C increases resistance to R. solanacearum
-
The VIGS experiment showed that plants transfected with TRV2::CaTIR1C effectively silenced CaTIR1C, with significantly decreased CaTIR1C expression compared to those transfected with the TRV2 empty vector (TRV2::00). Seedlings transfected with TRV2::CaChlH turned yellow-white 25 d after injection (Fig. 8a, b). Six days after R. solanacearum inoculation, control plants exhibited the typical symptom of wilted leaves, whereas CaTIR1C-silenced plants showed resistance with reduced disease symptoms (Fig. 8b; Supplementary Fig. S2). These results showed that CaTIR1C plays a negative role in regulating R. solanacearum resistance.

Figure 8.
Effects of CaTIR1C on pepper susceptibility to R. solanacearum. (a) Expression of CaTIR1C in leaves at 25 d after VIGS infection. Asterisks indicate statistical significance as determined by the two-tailed Student's t-test: **p < 0.01. (b) Young pepper plants at 6 d after the inoculation with 0.1 mL R. solanacearum injection (+ R. solanacearum). Bar = 0.5 cm.
-
With the development of whole genome sequencing, TIR1/AFB families have been identified in more plant species in recent years[6]. As auxin co-receptor proteins, TIR1/AFBs were reported to regulate many aspects of plant growth, development, and responses to salt stress[41−43]. The present study conducted the first genome-wide analysis on TIR1/AFB family in pepper.
In plant genomes, the number of TIR1/AFB genes varies. The Arabidopsis genome contains six TIR1/AFBs, divided into three subgroups: TIR1/AFB1 (TIR1, AFB1), AFB2/3 (AFB2, AFB3), and AFB4/5 (AFB4, AFB5)[8]. In Brassica juncea var. tumida, 18 TIR1/AFB genes have been identified and could be clustered into four subgroups, including TIR1/AFB1, AFB2/3, AFB4/5, and AFB6[44]. In Solanaceae crops, four TIR1/AFB genes have been reported in tomatoes, and five TIR1/AFB genes in eggplant. The present analysis identified five TIR1/AFB genes in the pepper genome (Table 1), which is similar to eggplant but one more than in tomato. These findings suggest that while Solanaceae species share a relatively conserved TIR1/AFB gene count, lineage-specific expansion and diversification.
Extensive functional overlapping and specification have been reported among TIR1/AFB genes[6], with their varied functions due to structural divergence[45]. In this study, differences in gene structure within the CaTIR1/AFB gene family were observed. At least two untranslated regions (UTRs) were detected in CaTIR1C, CaAFB4/5, and CaAFB6, with CaAFB6 containing four UTRs. CaTIR1A contained four coding sequences (CDSs), compared with three CDSs in each of the other four members (Fig. 3), which indicates the unique function of CaTIR1A. While the five CaTIR1/AFB proteins shared similar motif compositions, which are characteristic of resistance proteins known to play crucial roles in plant innate immunity by mediating pathogen recognition and initiating defense responses[46]. A notable LRR_CC_2 domain was observed in CaAFB6, highlighting its potential involvement in specialized signaling functions.
Interestingly, duplication links of the TIR1/AFB genes were found in eggplant (SmTIR1B and SmTIR1C) and pepper (CaTIR1B and CaTIR1C), which are not found in tomato (Fig. 2). This may be connected to the genes' segmental duplication[7]. Duplications of genes involved in pathogenicity or stress responses are widely recognized as a mechanism that enables plants to acquire novel functions or regulatory complexity, facilitating adaptation to diverse biotic challenges[47,48]. This observation suggests that CaTIR1B and CaTIR1C may be functionally related to plant resistance. Notably, differences were detected in the UTR region and defense-responsive cis-elements in the promoters of these two genes, which may lead to the expression differences.
Gene expression patterns are usually associated with their functions[49−51], while genome-wide analyses on these patterns and functions have been utilized to investigate transcript levels. For example, studies have examined the expression profiles of TIR1/AFB under salt stress in Brassica juncea var. tumida and Malus pumila, revealing condition-specific regulatory patterns[44,52]. In this study, expression profiling under the R. solanacearum inoculation treatment demonstrated three distinct expression clusters among the CaTIR1/AFB genes based on their expression patterns.
Notably, CaTIR1B and CaTIR1C are similar in CDS and exon-intron structures; however, CaTIR1C was significantly upregulated in the susceptible pepper variety. This upregulation may be attributed to the presence of unique defense-responsive cis-elements in the promoter region of CaTIR1C. Virus-induced gene silencing (VIGS) of CaTIR1C revealed that its downregulation enhanced resistance to R. solanacearum. These findings suggest that CaTIR1C promotes susceptibility to R. solanacearum, potentially through modulating of auxin-related signaling pathways. Nevertheless, the exact molecular mechanisms by which CaTIR1C contributes to pathogen response require further investigation.
Bacterial wilt poses a major threat to global crop production, particularly affecting Solanaceae crops, including pepper[17]. Research showed that the accumulation of auxin is promoted in Arabidopsis under bacterial wilt infection[53], and genome-wide transcriptomic changes of auxin pathways in the resistant tomato variety were also observed[54]. However, the relationship between bacterial wilt and auxin has not been clearly elucidated. The findings of this study suggest that functional diversification within the CaTIR1/AFB family contributes to variety-specific responses to R. solanacearum infection among resistant and susceptible pepper varieties. Given the physiological diversity of R. solanacearum[55], this gene-level specificity may confer a selective advantage, possibly explaining the conserved nature of CaTIR1/AFB genes within the pepper genome. This is the first study to identify roles for CaTIR1/AFB genes in pepper in the context of R. solanacearum infection. While conclusions are primarily drawn from bioinformatic and transcriptome analyses, further experimental validation is necessary to elucidate the precise roles of TIR1/AFB genes within the broader regulatory network of plant growth and immune responses.
In conclusion, five TIR1/AFB genes were identified in the pepper genome. Bioinformatic and transcriptome analysis of gene structure and conserved motifs revealed both functional redundancy and diversification in CaTIR1/AFBs under R. solanacearum. Notably, CaAFB6 was identified in pepper, together with AFB6 in tomato and eggplant, suggesting lineage-specific expansion in Solanaceae. Among the identified genes, CaTIR1C uniquely harbored defense-responsive cis-elements and displayed distinct expression patterns between resistant and susceptible pepper varieties upon R. solanacearum infection. Further functional evidence from expression profiling, subcellular localization, and VIGS assays highlighted CaTIR1C as a key regulator in pepper's response to R. solanacearum. These findings provide valuable insights into the roles of TIR1/AFB genes in pepper, establishing a foundation for future functional studies with approaches such as gene overexpression, RNA interference, and genome editing.
This work was supported by the Doctoral Fund of Baoding University (Grant No. 2023B03), and the Found of Innovation Capability Enhancement Special Project of Baoding Science and Technology Bureau (Grant No. 2494N005).
-
The authors confirm contributions to the paper as follows: methodology: Liu P, Jia Z, Xia M, Hua J; formal analysis, investigation: Liu P, Jia Z, Karamat U, Zhang C, Teng Z, Zhang B; preparation, validation: Karamat U, Zhang C, Teng Z, Zhang B; writing, original draft: Du W, Yu Z, Miao X; manuscript editing: Liu P, Jia Z, Du W, Yu Z, Miao X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Wenchao Du, Zhiyu Yu
- Supplementary Table S1 Primers for qRT-PCR analysis.
- Supplementary Table S2 Primers for VIGS and subcellular location analysis.
- Supplementary Table S3 TIR1/AFB genes identified in Capsicum annuum.
- Supplementary Fig. S1 Transcriptomic expression analysis of the CaTIR1/AFB genes of resistant variety (R) and susceptible variety (S) under control (C) or R. Solanacearum treatment (T).
- Supplementary Fig. S2 Representative seedlings six days after 0.1 mL R. solanacearum injection (+R. solanacearum).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Du W, Yu Z, Miao X, Teng Z, Zhang C, et al. 2025. Unveiling the role of the TIR1/AFB gene family in Capsicum annuum: genome-wide identification, characterization, and transcriptomic analysis under Ralstonia solanacearum infection. Vegetable Research 5: e034 doi: 10.48130/vegres-0025-0028 

Unveiling the role of the TIR1/AFB gene family in Capsicum annuum: genome-wide identification, characterization, and transcriptomic analysis under Ralstonia solanacearum infection
- Received: 06 May 2025
- Revised: 10 July 2025
- Accepted: 21 July 2025
- Published online: 16 September 2025
Abstract: TIR1/AFB proteins play key roles in plants responding to Solanum crop disease. In this study, five putative TIR1/AFB proteins were identified in pepper (Capsicum annuum). Chromosomal mapping revealed that CaTIR1/AFB genes are located on chromosomes DH02, DH03, DH04, and DH06. Phylogenetic tree analysis based on TIR1/AFB protein sequences from Arabidopsis, tomato, and eggplant revealed that the CaTIR1/AFB genes in pepper belong to three of the four identified subgroups. One collinear gene pair (CaTIR1C and CaTIR1B) was identified in pepper, suggesting the TIR1/AFB gene family in pepper experienced segmental duplications in evolution. Seventy-eight cis-elements related to the hormone, defense, drought, and light response were identified. RNA-sequencing data and expression profiling showed that TIR1/AFB genes were expressed differently under R. solanacearum treatment; downregulated expression of all CaTIR1/AFBs was observed in the resistant variety, while CaTIR1C was upregulated in the susceptible variety. Subcellular localization analysis showed that CaTIR1C is a nuclear-localized protein. Further virus-induced gene silencing (VIGS) assays revealed that knocking down CaTIR1C enhanced pepper resistance to R. solanacearum, indicating that CaTIR1C is a negative regulator in response to R. solanacearum. These findings contribute to the molecular breeding of bacterial wilt-resistant pepper and hold the potential to improve disease control efficiency by further exploring the function of CaTIR1C.
-
Key words:
- Capsicum annuum /
- Genome-wide analysis /
- TIR1/AFB gene family /
- VIGS /
- CaTIR1C /
- Ralstonia solanacearum