









-
Kale (Brassica oleracea var. acephala) is a horticultural variety of wild cabbage from the Brassicaceae family. Its curly leaves are highly nutritious, making it one of the healthiest vegetables in the world[1]. Kale is rich in many vitamins, including C, A, K, B1, B2, B3, and minerals such as iron, phosphorus, and calcium that help lower blood pressure and protect cardiovascular system[2]. Additionally, kale contains β-carotene, a potent antioxidant, as well as indole-3-methanol and sulforaphane, which exhibit both anti-cancer activity and antioxidant functions, and may also enhance the flavor profile of kale[3]. The significant health benefits of kale have led to an increase in consumer interest and increased demand for its cultivation and quality.
Selenium is an essential trace element for humans, playing crucial roles in antioxidant defense and immune function[4]. Although selenium cannot be synthesized endogenously in humans, it is absorbed and metabolized by plants, where it undergoes biotransformation into more bioavailable forms with higher biological activity[5,6]. There are inorganic and organic forms of selenium in nature. Previous studies on selenium biofortification have primarily focused on inorganic forms[7,8]; however, their assimilation efficiency is limited, and they pose potential ecotoxicity[9]. Organic selenium mainly includes seleno-amino acids, selenium-proteins, etc., which serve as vital sources of exogenous selenium supplementation due to their high bioavailability and low toxicity[10]. L-Selenomethionine (SeMet), molecular formula C5H11NO2Se, is an amino acid formed by selenium replacing sulfur in methionine, and is a compound of selenium in organic form in plants[11]. Research has demonstrated that SeMet participates in the regulation of plant growth processes, thereby significantly improving plant growth and nutritional quality of fruits[12,13]. Although SeMet is highly bioactive and offers critical advantages[14], its application is not thoroughly investigated, and its impact on kale quality is still uncertain.
Flavonoids, a ubiquitous class of polyphenolic compounds in plants, have emerged as a prominent research focus in medicine and food science due to their potent natural antioxidant properties[15,16]. They have the functions of scavenging free radicals, up-regulating the expression of antioxidant enzymes, and regulating the MAPK signaling pathway, which effectively enhances the antioxidant capacity and alleviates oxidative stress-induced damage in plants[17,18]. According to their skeleton structure, they can be divided into seven subclasses: flavonols, flavones, isoflavones, anthocyanidins, flavanones, flavanols, and chalcones. It should be emphasized that flavonoid biosynthesis is modulated by diverse external factors through both molecular mechanisms and physiological processes[19]. The accumulation and transport of flavonoids are transcriptionally controlled by the MBW (MYB-BHLH-WD40) complex, in which the MYB transcription factor is the main determinant[20,21] by binding directly or indirectly to the promoters of genes involved in flavonoid synthesis and transport, thereby affecting flavonoid accumulation[22,23]. In Arabidopsis thaliana, the specific regulation of flavonol biosynthesis is carried out by AtMYB12 through the activation of promoters for essential genes such as CHS and F3H[24]. Notably, when AtMYB12 was heterologously expressed in kale, it also led to high flavonol accumulation[25]. Additionally, AtMYB3, AtMYB7, AtMYB4, and AtMYB32 in Arabidopsis thaliana are capable of directly binding to the promoters of genes associated with flavonoid biosynthesis, leading to the repression of downstream gene expression[26]. In other plants, SbMYB3, CsMYB60, PsMYB12, VvMYBPA2, VvMYBA2, and ZmC1 all play roles in regulating flavonoid biosynthesis, indicating that MYB transcription factors during evolution are highly conserved in function[27,28]. Furthermore, exogenous active substances such as melatonin, phytohormone, and sodium selenite also regulate flavonoid accumulation and antioxidant capacities[29,30].
In this investigation, the effects of different SeMet levels on kale's nutritional quality were assessed, with metabolomics and transcriptomics revealing the main differential metabolites and key genes, and further analysis was conducted on how SeMet regulates kale's nutritional quality at the molecular level. These findings will establish a theoretical foundation for understanding SeMet's role in kale nutrition and enable a comprehensive analysis of its quality formation mechanisms.
-
In this experiment, 'Green Rosa' (a new leafy kale cultivar) was selected as the experimental material due to its desirable commercial traits, including high yield, stress resistance, stress tolerance, superior taste, marketability, and consistently high germination rates.
The SeMet (CAS: 3211-76-5), serving as the Se donor, was sourced from Aladdin Biochemical Technology Co., Ltd (Shanghai, China).
Seeds of full and uniform size were selected, germinated for 3 d in moistened medium, and transferred into 7 cm × 7 cm black containers filled with an equal mix of nutrient soil and vermiculite. All seedlings were cultivated in controlled growth chambers set to 22 ± 2 °C with a 16 h light period (8 h dark period), a light level of 100 μmol·m−2·s−1, and a humidity ranging from 30% to 50%.
Experimental design
-
To explore the appropriate Se concentration for kale, five concentration gradients of SeMet solutions of 0, 5, 10, 20, and 30 mg·L−1 containing 0.01% Tween-20 were prepared based on previous studies[13,14]. Twenty-five-day-old kale seedlings were selected, and ten seedlings were treated with each concentration. Each seedling was sprayed with 5 mL of the respective SeMet solution to ensure uniform leaf coverage and prevent dripping once every 5 d, a total of four times. The biomass, selenium content, chlorophyll, flavonoids, and other related physiological indices were measured post-treatment.
To investigate the time effect of Se treatment on kale seedlings, the contents of Se, total phenolics, and flavonoids were measured at 0, 1, 3, 5, and 7 d after Se treatment, and water containing 0.01% Tween-20 was used as the control (CK). Kale seedlings treated with 10 mg·L−1 SeMet solution and H2O for 5 d were collected, instantly frozen using liquid nitrogen, and kept at −80 °C for future transcriptomic and metabolomic analysis.
Biomass measurement
-
Kale seedlings were cut from the base of the stem, and their fresh biomass was quantified using a microbalance (Sartorius, Beijing, China). Ten grams fresh leaves were randomly weighed, and the dry matter was measured after drying in an oven at 65 °C.
Total phenols and flavonoids measurement
-
According to the method[31], a 2 g fresh sample was extracted with 20 mL pre-cooled 1% HCl-methanol solution at 4 °C for 20 min, then centrifuged at 8,000 rpm for 10 min at 4 °C. The clarified supernatant was taken to measure absorbance at 280 and 325 nm, and total phenolics and flavonoids contents were calculated using gallic acid and rutin calibration curves, respectively.
Selenium content measurement
-
Selenium was extracted from kale leaves using a pressure tank digestion method and quantified by inductively coupled plasma mass spectrometry (ICP-MS)[15] (PerkinElmer NexlON®1,000, Waltham, USA). To eliminate potential interference from surface contaminants, all leaf samples were carefully washed with deionized water prior to selenium quantification.
Total antioxidant capacity measurement
-
The evaluation of kale leaves' antioxidant capacity was conducted using DPPH, ABTS, and FRAP assay following the instruction manual of Suzhou Keming Biotechnology Co., Ltd (Jiangsu, China).
Metabolite extraction and analytical procedures
-
Freeze-dried kale leaves were pulverized to a fine powder, with 50 mg aliquots subsequently combined with 1.2 mL of chilled 70% methanol solution containing internal standards. After vortexing, the mixtures underwent centrifugation at 12,000 × g for 3 min at 4 °C. The clarified supernatant was subsequently filtered through a 0.22 μm nylon membrane before chromatographic analysis using a UPLC-MS/MS platform (SCIEX ExionLC™AD, Framingham, USA). Analytical separation was accomplished on an Agilent SB-C18 column (2.1 × 100 mm, 1.8 μm particle size) employing a binary mobile phase system: component A (aqueous 0.1% formic acid) and component B (acetonitrile with 0.1% formic acid). The elution protocol consisted of an initial 5% B composition, progressively ramped to 95% B over 9 min, then maintained for 1 min before column re-equilibration. The gradient program was adjusted to 5% B through a 1.1 min transition (10.0−11.1 min), followed by system stabilization at 5% B until 14 min. The mobile phase was delivered at 0.35 mL·min−1 through a column thermostatted to 40 °C, with 2 μL sample injections.
Mass spectrometric conditions were configured with: ion source heated to 500 °C, electrospray ionization voltages: +5,500 V for positive ion detection and −4,500 V for negative mode, auxiliary gas pressures set at 50 psi (GSI), 60 psi (GSII), and 25 psi for curtain gas. Metabolite identification was conducted using the MetWare database with secondary MS/MS fragmentation data. Quantitative measurements utilized MRM detection with metabolite-specific declustering potentials and collision energies optimized for each analyte. Following data acquisition, chromatographic peak alignment and integration were performed across samples. Metabolic pathways were annotated through KEGG enrichment analysis (hypergeometric test) using the KEGG Pathway database (
www.kegg.jp/kegg/pathway.html ).RNA-seq
-
Total RNA was extracted from the samples, the purified mRNA fragments were converted to cDNA, and sequencing libraries were constructed using adapter ligation and PCR amplification. High-throughput sequencing was performed using Illumina technology with 150 bp paired-end reads. Initial data processing involved adapter removal and quality control through fastp (v0.23.2). Processed reads were mapped to the GWH reference genome (Assembly ID: 68029) through HISAT2 (v2.2.1), with transcript abundance quantified using fragments per kilobase million (FPKM) measurements. Statistical analysis of differential expression was conducted through DESeq2, applying significance criteria of adjusted p < 0.05 and absolute log2 fold-change > 1. Biological interpretation included KEGG pathway analysis and Gene Ontology term enrichment evaluation using hypergeometric testing (significance threshold p < 0.05). Sequencing datasets are publicly accessible through NCBI's SRA repository under BioProject PRJNA1220178 (
www.ncbi.nlm.nih.gov/sra/ PRJNA1220178 ).Real-time PCR
-
Total RNA extraction from kale leaves was performed using TRNzol Universal Reagent (TIANGEN, Beijing, China). cDNA synthesis was carried out with StarScript III All-in-one RT Mix containing gDNA Remover (GenStar, Beijing, China) following the manufacturer's protocols. Amplification reactions were executed in triplicate using 2 × RealStar Fast SYBR qPCR Mix (GenStar, Beijing, China) with specific primer pairs (Supplementary Table S1). Select β-ACTIN (BroT26.C01g38600) as the housekeeping gene, and relative gene expression quantification was achieved through the comparative threshold cycle (2−ΔΔCT) calculation method.
Vector construction and transient transformation
-
To generate BroMYB34-overexpressing transgenic plants, the coding sequence (CDS) of BroMYB34 was cloned and inserted into the CaMV35S-driven pSuper1300-GFP. The recombinant plasmid was subsequently transformed into Agrobacterium strain LBA4404 through a freeze-thaw method. For transient transformation, uniformly grown kale leaves were infiltrated with Agrobacterium suspensions carrying the BroMYB34 overexpression construct via sterile syringe. Leaves were incubated in the dark for 24 h, followed by 1–5 d of normal cultivation to assess total phenolic and flavonoid contents and related gene expression. Gene-specific primers employed for vector construction are detailed in Supplementary Table S1.
Dual luciferase reporter gene assay
-
The promoter regions of BroT26.C02gCHS, BroT26.C03gCHS3, and BroT26.C09gCHS3, along with the CDS of BroMYB34, were directionally cloned into the pGreenII 0800-LUC reporter and pGreenII 62-SK effector vectors, respectively. The recombinant constructs were transformed into Agrobacterium strain GV3101 (pSoup) and infiltrated into tobacco leaves. After 48 h of dark incubation, luminescence signals were quantified using the Dual-Luciferase Reporter Assay System (Promega, WI, USA), with firefly luciferase (LUC) activity normalized against Renilla luciferase (REN) internal control. Simultaneously, bioluminescence imaging was performed using a cooled CCD imaging system (BIO-OI, Guangzhou, China) to visualize spatial expression patterns. The relevant primers for target amplification are provided in Supplementary Table S1.
Yeast one hybrid
-
The 50 bp core promoter region containing the MYB-binding site of BroCHSs was inserted into the pHIS2 reporter vector, while the coding sequence (CDS) of BroMYB34 was subcloned into the pGADT7 bait vector. Co-transformation of both constructs into Y187 yeast cells, then selected on SD/-Leu-Trp and SD/-Leu-His-Trp medium containing 3-AT to assess interaction-dependent growth. Primer sequences for cloning are detailed in Supplementary Table S1.
Statistical analysis
-
All experimental data were biologically replicated three times and analyzed using Microsoft Office Excel. Statistical analysis was carried out by DPS software, the significance between two treatments was assessed by Student's t-test (* p < 0.05; ** p < 0.01), and Duncan's multiple range test (p < 0.05) was used for comparisons among multiple treatments.
-
The study explored the effects of SeMet on the growth of kale seedlings. Results showed that Se content accumulated continuously in kale as the SeMet concentration increased (Supplementary Fig. S1a). However, no significant differences were observed in the fresh weight and leaf dry matter content of kale seedlings after treatment with SeMet at different concentrations, indicating that exogenous organic selenium neither promoted nor inhibited the growth of kale seedlings (Fig. 1a, b). Similarly, SeMet treatment had no significant impact on chlorophyll a, chlorophyll b, or carotenoid levels in kale seedlings (Supplementary Fig. S1b–d). In terms of quality, soluble sugar and soluble protein contents increased and then decreased as SeMet concentration increased, and the 10 mg·L−1 treatment was the highest. However, the total glucosinolate content in kale was significantly reduced, whereas vitamin C content decreased initially, reaching the lowest at 20 mg·L−1 treatment and the highest at 30 mg·L−1 treatment (Supplementary Fig. S2). Notably, a significant increase in total phenolics and flavonoid contents was observed at 10 mg·L−1 SeMet. Compared with 0 mg·L-1 treatment, the total phenolics and flavonoids contents of 10 mg·L−1 treatment were significantly increased by 19.54% and 18.88%, respectively (Fig. 1c, d). In general, 10 mg·L−1 SeMet is the optimal concentration for exogenous application to kale, significantly improving the nutritional quality of kale seedlings.
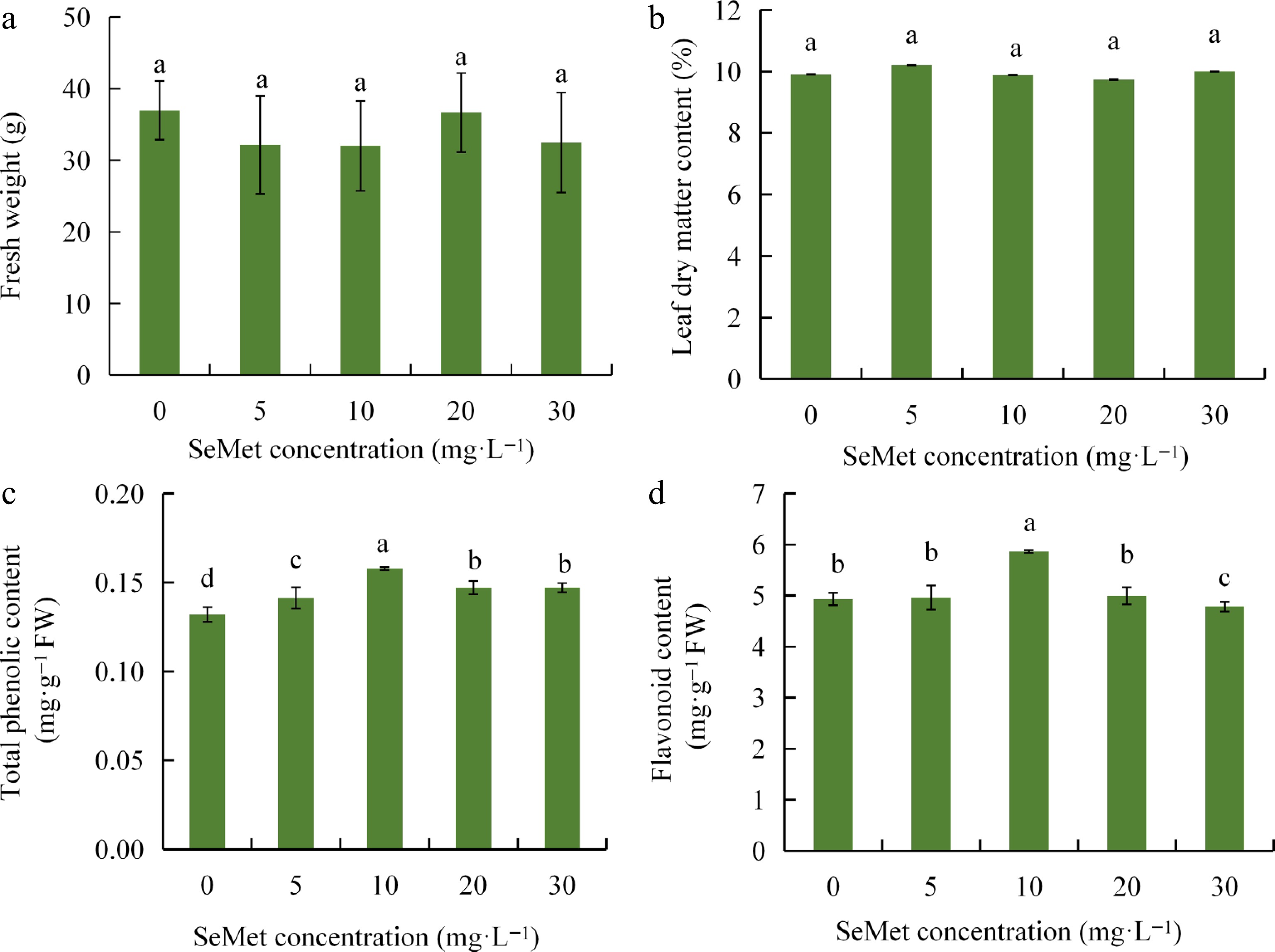
Figure 1.
Effects of different exogenous concentrations of SeMet on the growth and quality of kale seedlings. (a) Effect of SeMet on fresh weight of kale seedlings. (b) Effect of SeMet on leaf dry matter of kale seedlings. (c) Effect of SeMet on total phenolic content of kale seedlings. (d) Effect of SeMet on the flavonoid content of kale seedlings. Data shown as mean ± SD (n = 3). Means with different letters differ significantly (Duncan's test, p < 0.05).
To study the cumulative effect of selenium in kale leaves, the study determined Se content in leaves following exogenous application of 10 mg·L−1 SeMet for 0, 1, 3, 5, 7 d. Results showed that Se content gradually accumulated over time (Fig. 2a), while the total phenolic content significantly increased after 3 d, and flavonoid content significantly increased throughout the 1 to 7-day treatment period. Compared with the CK treatment, the total phenolics and flavonoid contents reached their highest levels at 5 d of exogenous SeMet treatment, increasing by 1.45 fold and 1.28 fold, respectively (Fig. 2b, c). Flavonoids are well-established regulators of plant antioxidant capacity. DPPH, ABTS, and FRAP radical scavenging capacities were measured to evaluate the impact of exogenous SeMet on the antioxidant capacity of kale seedlings. Results from all three experiments showed that SeMet significantly enhanced the antioxidant capacity of kale seedlings during 1 to 7 d, with the highest DPPH, ABTS, and FRAP scavenging capacities observed at 5 d of exogenous SeMet treatment, increasing by 1.43 fold, 1.07 fold and 1.16 fold, respectively (Fig. 2d−f). To further elucidate the correlation between flavonoids and antioxidant capacity, the quantitative relationship between flavonoids content and antioxidant capacity after different days of CK and SeMet treatment was analyzed. The results demonstrated that both total phenolics and flavonoid contents exhibited highly significant correlations with DPPH, ABTS, and FRAP assays. Specifically, Pearson correlation coefficients were 0.74, 0.68, and 0.78 for total phenolics and 0.85, 0.80, and 0.86 for flavonoids, respectively (Fig. 2g). These results suggest that SeMet regulates flavonoid accumulation as well as enhances the antioxidant capacity of kale seedlings.
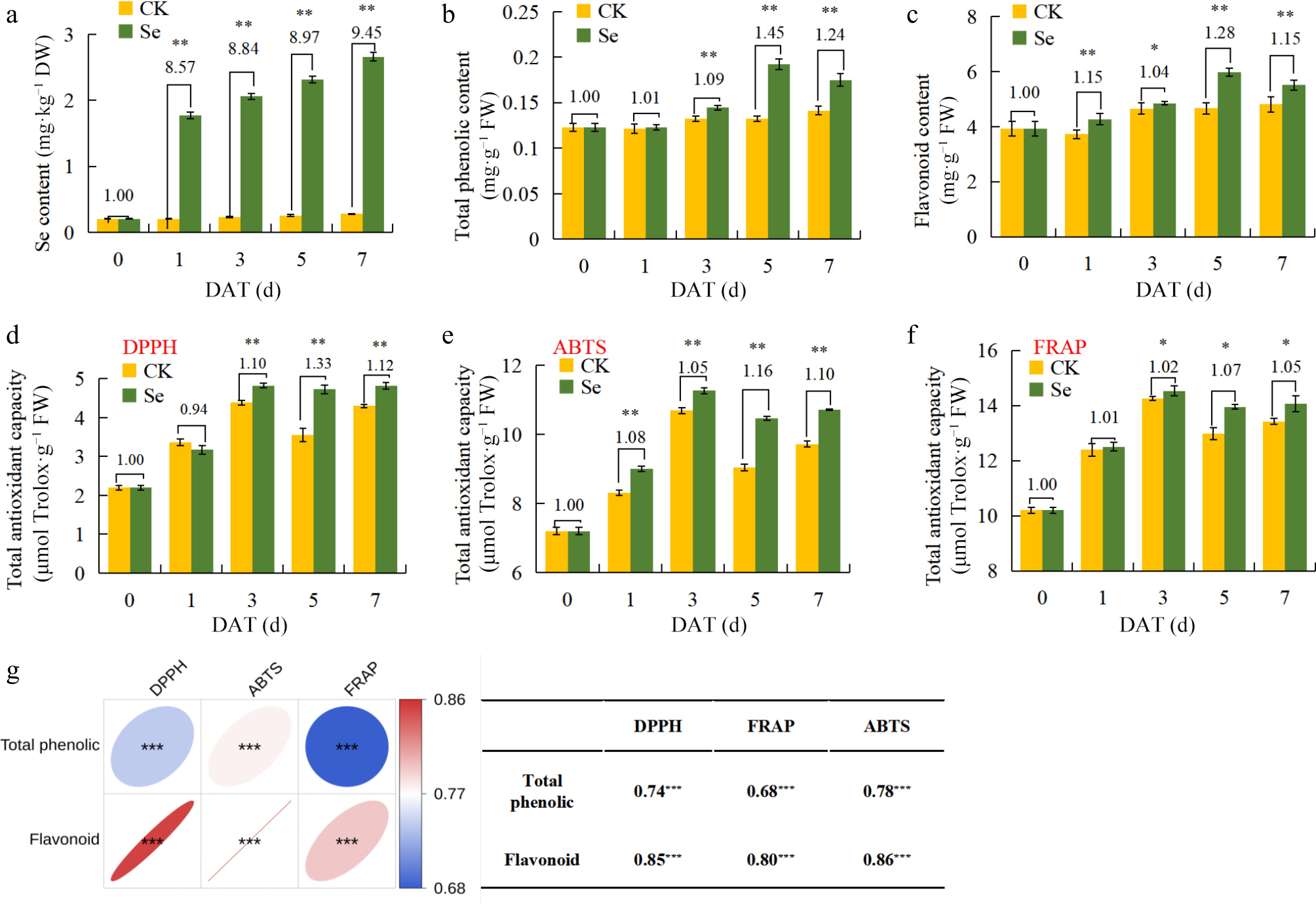
Figure 2.
Effects of kale seedlings after exogenous SeMet treatment on different sampling days. (a) Changes in Se content in kale seedlings with different sampling days. (b) Changes in total phenolic content in kale seedlings with different sampling days. (c) Changes in flavonoid content in kale seedlings with different sampling days. (d) Changes of DPPH radical scavenging capacity in kale seedlings with different sampling days. (e) Changes of FRAP radical scavenging capacity in kale seedlings with different sampling days. (f) Changes of ABTS radical scavenging capacity in kale seedlings with different sampling days. (g) The correlation between flavonoids and antioxidant capacity. The kale seedlings were treated with 10 mg·L−1 for 0 d and 5 d, and the H2O treatment was the CK. Statistical analysis was performed using Student's t-test on triplicate measurements (mean ± SD), with significance levels denoted as follows: * p < 0.05; ** p < 0.01. The DAT means day after treatment.
Metabolome analysis of SeMet on kale seedlings
-
The metabolome of kale leaves treated with 10 mg·L−1 SeMet for 5 d was analyzed, with H2O treatment as the control. A total of 2,317 metabolites were identified in this study, among which alkaloids, amino acids and their derivatives, flavonoids, terpenoids, and phenolic acids accounted for 12.47%, 11.61%, 11.83%, 11.61%, and 11.18% of all metabolites, respectively (Fig. 3a, Supplementary Data S1).
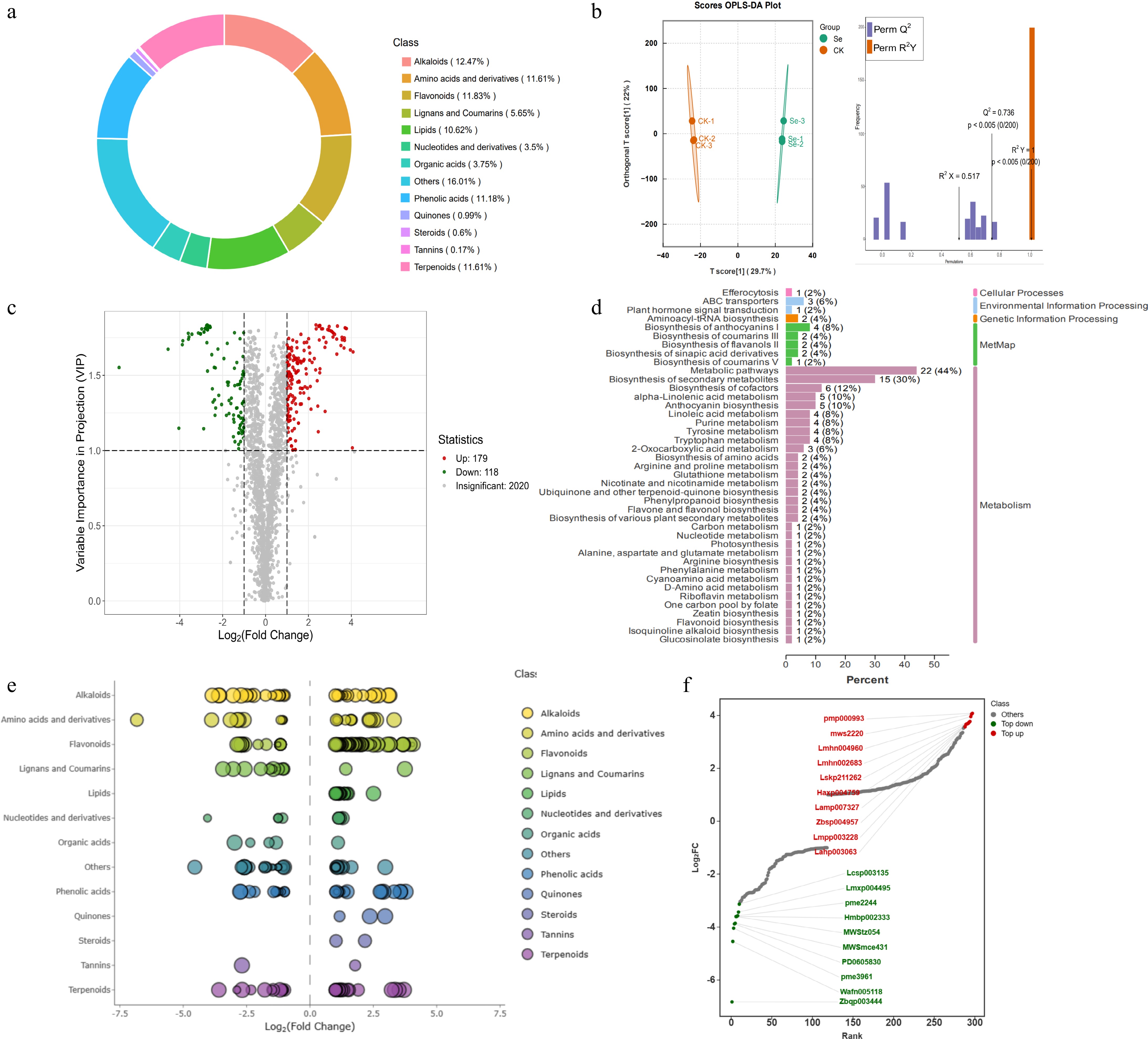
Figure 3.
Metabolome analysis of kale seedlings in response to exogenous SeMet. (a) Classification of metabolites in the metabolome. (b) OPLS-DA analysis of metabolites between the Se and CK groups. (c) Volcano plots for DEMs between the Se and CK groups. Metabolite up-regulation is color-coded in red, while down-regulation is shown in green. (d) KEGG pathway enrichment analysis of DEMs. (e) Scatter plots visualization of DEMs showing log2FC values on the x-axis and variable importance in projection (VIP) scores represented by dot size. (f) Dynamic distribution of the top 10 up-regulated and down-regulated DEMs with Log2FC. Metabolite up-regulation is color-coded in red, while down-regulation is shown in green.
Orthogonal partial least squares discriminant analysis (OPLS-DA) revealed significant separation between SeMet-treated and CK groups, as evidenced by the model parameters of R2X = 0.517 and R2Y = 1 indicated strong explanatory power for the X and Y matrices, respectively, while Q2 = 0.736 confirmed effective predictive capability, suggesting excellent model performance (Fig. 3b). Using variable importance in projection (VIP) ≥ 1 and |log2foldchange| > 1 as thresholds, a total of 297 differential metabolites (DEMs) were screened, comprising 179 up-regulated and 118 down-regulated compounds (Fig. 3c, Supplementary Data S2). Functional annotation of DEMs via KEGG and Metmap showed that most metabolites were related to metabolic processes, with a large number annotated in metabolic pathways and biosynthesis of secondary metabolites (Fig. 3d). Classification of all the DEMs showed that flavonoids had the highest content, with most accumulating under SeMet treatment (Fig. 3e). In addition, the top ten substances of |log2FC| in DEMs are counted. Among up-regulated metabolites, pmp000993, mws2220, Lmhn004960, Lamp007327, Zbsp004957, and Lahp003063 were identified as flavonoids (Fig. 3f). These results highlight flavonoids as key metabolic components potentially involved in SeMet regulation of kale quality.
The flavonoids in all the DEMs were further analyzed. The classification of flavonoids primarily included flavonols, accounting for 41.25% of all differential flavonoids, including 27 up-regulated and four down-regulated substances. The remaining classification comprised flavones (25%), anthocyanidins (12.5%), flavanones (6.25%), flavanonols (2.5%), flavanones (2.5%), isoflavones (2.5%), chalcones (1.25%), and other flavonoids (6.25%) (Fig. 4a, b). All differential flavonoids are shown in violin diagrams (Supplementary Fig. S3). Eight metabolites were annotated to the KEGG pathways, which are related to metabolic pathways, biosynthesis of secondary metabolites, flavone and flavonol biosynthesis, flavonoid biosynthesis, and anthocyanin biosynthesis (Fig. 4c). As can be seen from Fig. 4d and Supplementary Data S3, among all 80 differential flavonoid metabolites, 15 flavonoids exhibited statistically significant differences (p < 0.05).
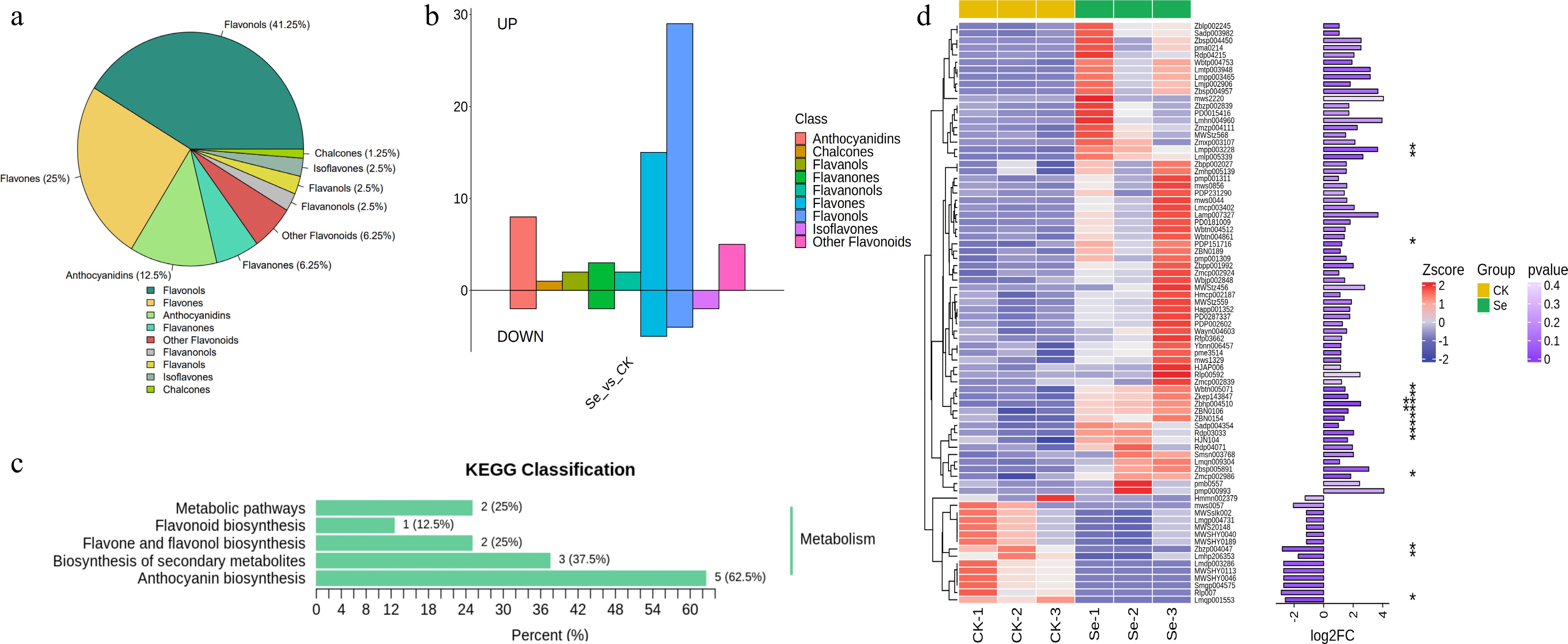
Figure 4.
Analysis of flavonoid substances in the metabolome. (a) Classification of flavonoid substances in DEMs. (b) Statistics of up-regulated and down-regulated flavonoid substances. (c) KEGG annotation of flavonoids in DEMs. (d) The cluster heat map of flavonoid substances in DEMs between the Se and CK groups. The length of the column on the right column represents the log2FC value of the substance, with significance levels denoted as follows: * p < 0.05; ** p < 0.01.
Transcriptome analysis of SeMet on kale seedlings
-
To investigate SeMet-responsive gene expression patterns, transcriptome analysis was performed on seedlings treated with 10 mg·L−1 SeMet for 5 d. Principal component analysis (PCA) revealed distinct clustering of gene expression profiles between Se and CK (Fig. 5a). Using screening criteria of |log2FC| ≥ 1 and false discovery rate (FDR) < 0.05, 484 differentially expressed genes (DEGs) were identified, comprising 228 up-regulated genes and 256 down-regulated genes (Fig. 5b, Supplementary Data S4). GO enrichment analysis of DEGs was performed, and the GO enrichment cycle was plotted according to the six GO terms with the lowest q-value across three categories of biological process, cellular component, and molecular function. This analysis showed extremely significant enrichment of DEGs in GO: 0009812 (flavonoid metabolic process) and GO: 0009813 (flavonoid biosynthetic process) of biological processes (Fig. 5c). As shown in Fig. 5d, KEGG pathway enrichment demonstrated predominant enrichment of DEGs in flavonoid biosynthesis. Figure 5e displays a heat map of all flavonoid-related genes, including 11 up-regulated and eight down-regulated genes. These results indicate that SeMet has a great effect on flavonoid synthesis and metabolism at the transcriptome level. In addition, further analysis of transcription factors among the DEGs regulated by SeMet showed a total of 20 differentially expressed transcription factors, including seven up-regulated and 13 down-regulated genes. It covers HSF, Tify, Trihelix, MYB, AP2/ERF, NAC, bHLH, C2H2, and WRKY type transcription factors (Fig. 5f), all of which potentially contribute to SeMet-induced modulation of flavonoid biosynthesis pathways. The RNA-seq expression patterns were validated by qRT-PCR analysis of ten representative genes, with expression trends matching the FPKM values (Supplementary Fig. S4).
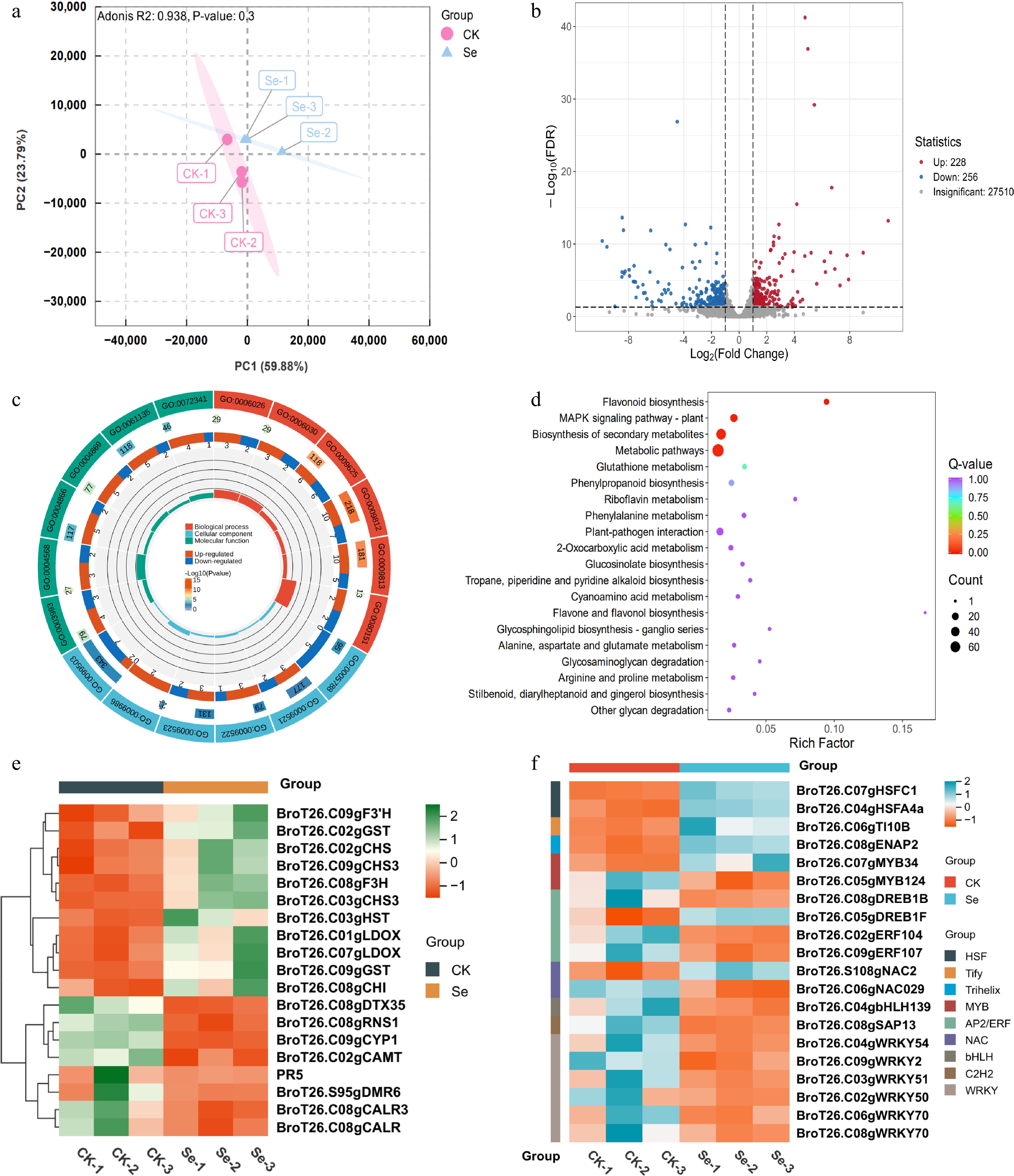
Figure 5.
Transcriptome analysis of kale seedlings in response to exogenous SeMet. (a) PCA plot of DEGs between SeMet-treated and control (CK) groups. (b) Volcano plot visualization of DEGs between SeMet-treated and CK groups. DEGs are color-coded with red indicating up-regulation and blue representing down-regulation. (c) GO enrichment analysis of DEGs. The first circle is KEGG_level_1 entries, with different colors representing different KEGG categories. The second circle is the pathway-specific gene counts and corresponding p-values of this category. The third circle is stacked bar plots showing up- (red) and down-regulated (blue) gene proportions. The fourth circle is the percentage of DEGs relative to total genes in each pathway category. (d) KEGG pathway enrichment analysis of DEGs. (e) Clustered heatmap of flavonoid-related DEGs. (f) Clustered heatmap of transcription factor DEGs.
BroMYB34 as the upstream transcription factor to activate the expression of BroCHSs
-
Chalcone synthase (CHS), as the pivotal rate-limiting enzyme, controls metabolic flux through its rate-limiting catalytic activity in the pathway's first committed step. Transcriptome profiling revealed three genes encoding CHS (BroT26.C02gCHS, BroT26.C03gCHS3, and BroT26.C09gCHS3) exhibiting significant up-regulation following SeMet treatment (Fig. 5e). To investigate the upstream transcription factors regulating BroCHSs, comprehensive cis-element analysis of their promoter regions was performed. The promoters contained multiple conserved regulatory motifs, including hormone-responsive elements (ABRE, as-1, TCA-box, P-box), low-temperature response elements (LTR), light-responsive elements (G-box), and developmental regulation-related elements (L-box, HD-Zip1, A-box). Notably, all three BroCHS promoters have MYB-binding sites, consistent with the established roles of MYB transcription factors in modulating flavonoid biosynthesis (Fig. 6a). This conserved cis-regulatory structure suggests BroCHS genes may be coordinately regulated by MYB-type transcriptional activators during SeMet-induced flavonoid accumulation.
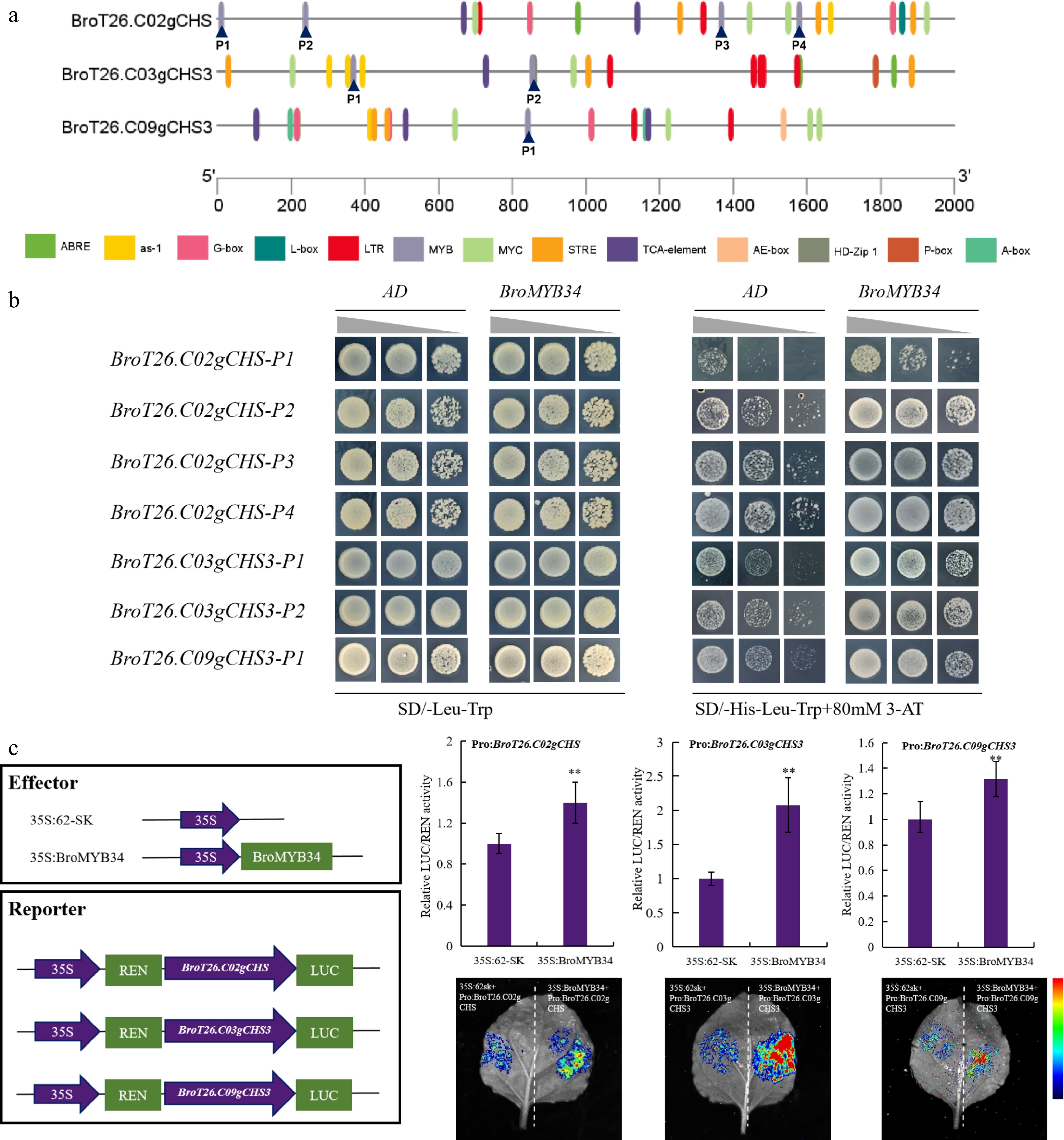
Figure 6.
BroMYB34 could bind with the promoter of BroCHSs to activate its expression. (a) The cis-elements analysis of BroCHSs promoters. The PlantCare was used for analysis, and TBtools was used for mapping. (b) The Y1H experiment is used to determine the binding between BroMYB34 and BroCHSs' promoters. (c) Dual luciferase reporter experiments demonstrated that BroMYB34 is the upstream activator of BroCHSs. The relative LUC/REN activity was performed using Student's t-test on triplicate measurements (mean ± SD), with significance levels denoted as follows: * p < 0.05; ** p < 0.01. An empty vector was used as the reference and set to 1.
Transcriptome profiling identified two candidate MYB transcription factors (BroT26.C07gMYB34 and BroT26.C05gMYB124), among which BroMYB34 exhibited a co-upregulation pattern with all three BroCHS genes under SeMet treatment (Fig. 5e, f; Supplementary Fig. S4), demonstrating their coordinated transcriptional regulation during SeMet-mediated responses. To confirm the relationship between the BroMYB34 and BroCHS genes, the promoter sequences of BroCHSs were analyzed and four MYB-binding sites were identified in BroT26.C02gCHS, two in BroT26.C03gCHS3, and one in BroT26.C09gCHS3 (Fig. 6a). Additionally, Y1H assays were performed using core sequences within 50 bp of these MYB-binding sites. The results showed that both proBroCHSs + AD and proBroCHSs + BroMYB34 grew normally on SD/-Leu-Trp medium, whereas proBroCHSs + BroMYB34 exhibited significantly better growth than proBroCHSs + AD on SD/-His-Leu-Trp medium supplemented with 80 mM 3-AT, indicating direct binding of BroMYB34 to the promoters of BroCHSs (Fig. 6b). To functionally validate this regulatory relationship, dual-luciferase reporter assays were conducted in tobacco. The BroMYB34 effector vector (35S::BroMYB34-62SK) was co-infiltrated with three individual reporter vectors containing BroCHSs promoters (Pro: BroCHS-LUC0800) driving firefly luciferase. Quantitative analysis revealed that overexpression of BroMYB34 significantly induced the relative LUC/REN activity of BroT26.C02gCHS, BroT26.C03gCHS3, and BroT26.C09gCHS3 promoters compared to empty vector controls, the results of imaging analysis were consistent with those of quantitative analysis (Fig. 6c). These findings demonstrate that BroMYB34 acts as a trans-activator to bind and enhance the transcriptional activity of all three BroCHSs promoters, establishing its role as a key upstream regulator in SeMet-mediated flavonoid biosynthesis.
BroMYB34 positively regulates flavonoid biosynthesis
-
To verify the biological function of BroMYB34, an overexpression vector of BroMYB34 (OE-BroMYB34) was constructed and transferred into the cotyledon of kale by transient transformation, with the empty vector as the control (EV). qRT-PCR analysis revealed that OE-BroMYB34 transcripts were significantly up-regulated from 1 to 5 d after treatment, peaking at 3 d with a 3.23-fold increase compared with EV (Fig. 7a). Concomitantly, the expression of BroT26.C02gCHS, BroT26.C03gCHS3 and BroT26.C09gCHS3 exhibited initial up-regulation followed by gradual decline in expression patterns, while maintaining significantly higher levels than EV throughout the 5 d after treatment (Fig. 7b−d). Compared with EV, the total phenolics and flavonoids contents in the leaves of OE-BroMYB34 were significantly higher (Fig. 7e, f), which was correlated with significantly improved antioxidant capacity at 4 d after treatment, as evidenced by 62.1%, 20.0%, 30.4% higher DPPH, FRAP, ABTS radical scavenging capacity relative to controls (Fig. 7g). These results demonstrate that BroMYB34 could induce the expression of key genes in flavonoid synthesis to promote flavonoid biosynthesis, and improve the antioxidant capacity of kale.
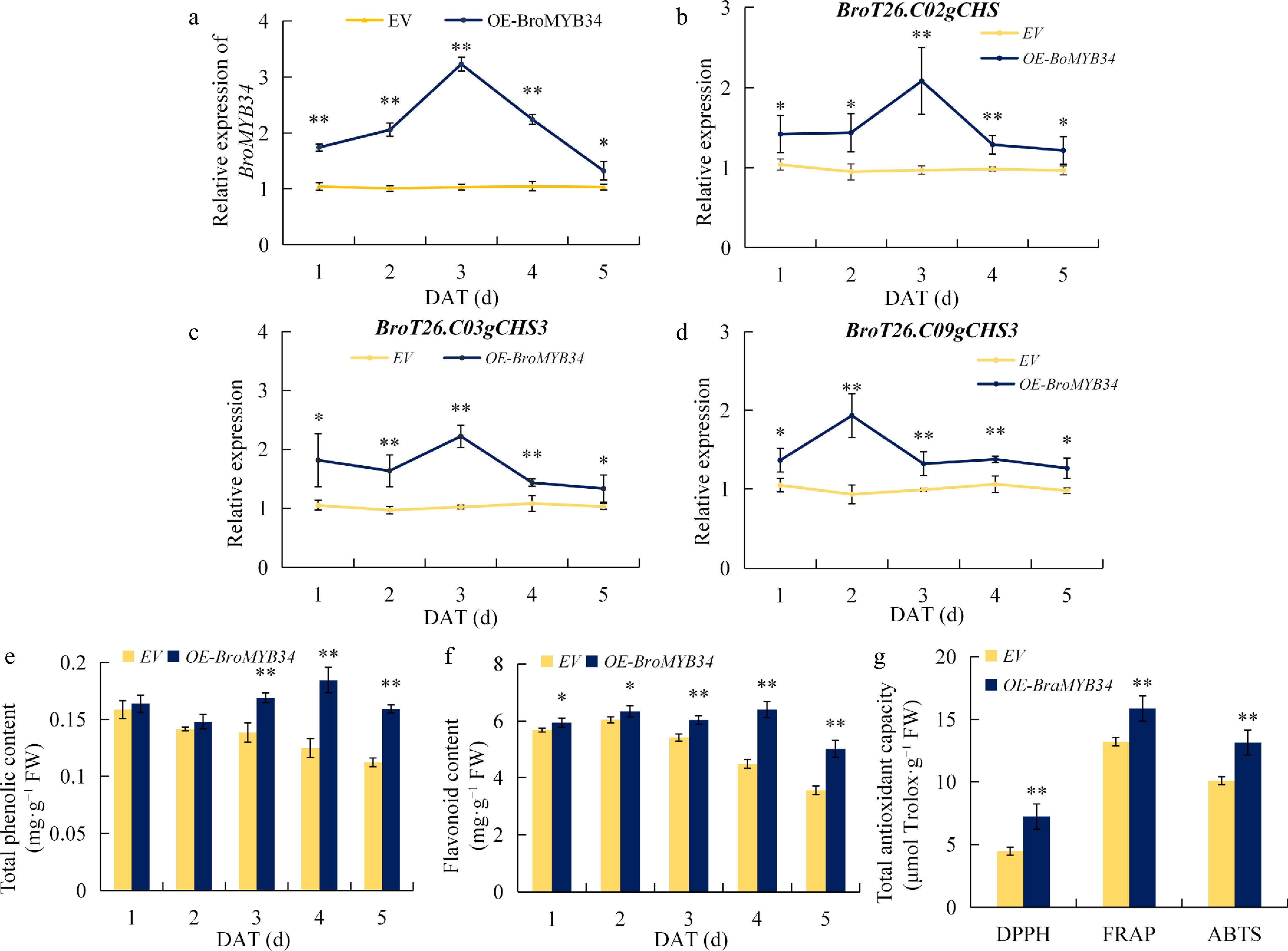
Figure 7.
Overexpression of BroMYB34 improves the flavonoid biosynthesis of kale. (a) BroMYB34 expression in EV and OE-BroMYB34 kales. (b)−(d) BroT26.C02gCHS, BroT26.C03gCHS3, and BroT26.C09gCHS3 expression in EV and OE-BroMYB34 kales. (e) Total phenolic contents in EV and OE-BroMYB34 kales. (f) Favonoid content in EV and OE-BroMYB34 kales. (g) Total antioxidant capacity in EV and OE-BroMYB34 kales. Empty vector (EV) and OE-BroMYB34 transient transgenic kales were sampled for 1−5 d. Statistical analysis was performed using Student's t-test on triplicate measurements (mean ± SD), with significance levels denoted as follows: * p < 0.05; ** p < 0.01. The DAT means day after treatment.
-
Kale is a highly nutritious vegetable, rich in nutrients and offering significant health benefits to humans[1,16]. SeMet, an organic form of selenium, exhibits high biological activity and low toxicity compared to inorganic selenium, making it a suitable exogenous selenium donor[9]. To better explore the effects of SeMet on the quality of kale, the optimal concentration of exogenous SeMet for kale was first screened, then transcriptional and metabolic differences were analyzed, aiming to reveal the mechanism by which SeMet regulates flavonoid biosynthesis in kale.
Exogenous SeMet promotes the enrichment of Se content and the synthesis of secondary metabolites in kale
-
SeMet, a form of organic selenium with antioxidant and antiviral biological functions, has been widely applied in food processing and livestock production and has gradually been applied to plant. Previous studies have shown that exogenous application of SeMet could significantly improve chlorophyll content, enhance photosynthesis, and increase plant biomass[13], whereas these effects depend on the plant species and variety[32]. In this study, it was found that exogenous SeMet significantly enhanced the Se content in kale with rising concentration, but different SeMet concentrations had no effect on fresh weight and dry matter content (Fig. 1a, b), indicating that kale has a strong ability for selenium enrichment and tolerance. However, the application of selenium exhibits a dose-dependent effect, and high concentrations can impede plant growth and development[33]. Notably, total phenolics, flavonoids, soluble sugars, and soluble proteins contents in kale increased firstly and subsequently decreased as SeMet concentration increased, peaking at the 10 mg·L−1 (Fig. 1c, d; Supplementary Fig. S2), indicating that 10 mg·L−1 is the optimal concentration of SeMet for exogenous application, and SeMet can promote the synthesis of secondary metabolites in kale. Compared with inorganic selenium, SeMet is an organic form of selenium that can be directly utilized by plants, exhibiting high bioavailability[11]. Consequently, lower concentrations typically suffice to promote plant growth, development, and quality formation. In contrast, sodium selenite must be converted into organic selenium within plants before being utilized[34], requiring a higher concentration (e.g., 40 mg·L−1) in kale[7], which may lead to greater toxicity and growth inhibition risks. Thus, using SeMet as a selenium donor in production may offer greater advantages.
In addition, the application SeMet has a time effect. Se content gradually accumulated in kale from 0 to 7 d under 10 mg·L−1 SeMet treatment, whereas total phenolics and flavonoids contents reached the highest at 5 d (Fig. 2a−c), indicating continuous Se enrichment in kale while its inductive effect diminishes over time. Appropriate selenium concentrations can improve activities of plant enzymes (SOD, CAT, APX, GR) and metabolism of non-enzymatic antioxidants (ascorbic acid, carotenoids, flavonoids, tocopherols), thereby regulating ROS metabolism to alleviate oxidative stress or boost antioxidant capacity[35]. Since the value of flavonoid in the SeMet-regulated process was analyzed, the antioxidant capacity of kale after exogenous SeMet treatment was assessed. The results showed that exogenous 10 mg·L−1 SeMet treatment significantly enhanced the antioxidant capacity of kale (Fig. 2d−f), consistent with the results for inorganic selenium[30]. These findings suggest that SeMet may regulate flavonoid accumulation to enhance the antioxidant capacity in kale seedlings. Both SeMet and inorganic selenium promote the accumulation of flavonoids in kale, with effects closely linked to enhanced antioxidant capacity[7] (Fig. 2g). This suggests such regulation may represent a general mechanism by which selenium modulates flavonoid biosynthesis.
Integrated metabolome and transcriptome analysis reveals flavonoids playing an important role in the improvement of the nutritional quality of kale by SeMet
-
In this study, there were 297 DEMs identified under 10 mg·L−1 SeMet treatment for 5 d compared with CK, and most of these were flavonoids, which are closely related to secondary metabolism biosynthesis, flavonoid biosynthesis, flavone and flavonol biosynthesis and anthocyanin biosynthesis pathways (Figs 3 and 4). In plants, flavonoids typically occur in the form of glycosides or other carbohydrates, often bound within plant tissues, with glucosides being the most prevalent[36]. Similarly, the metabolomic profiling revealed that the most significantly up-regulated DEMs in the flavonoid pathway corresponded to glycosylated derivatives. Flavonoids are vital secondary metabolites that play an important role in plant growth and development, stress regulation, and quality formation[37,38]. The transcriptome was used to mine the key DEGs that SeMet regulates in kale. A total of 484 DEGs were detected under SeMet treatment. GO and KEGG enrichment analyses indicated that these DEGs had significant correlations with flavonoid-related pathways (Fig. 5); they represented key structural genes in flavonoid biosynthesis and regulated flavonoid accumulation under SeMet treatment. Previous studies have reported that both organic and inorganic selenium can affect the flavonoid biosynthesis[7,9]; whereas, the application of SeMet in different plants induces significantly divergent downstream effects on plant metabolism. Exogenous SeMet differentially regulates genes involved in carotenoid and sugar metabolism, as well as antioxidant defense pathways, thereby increasing soluble solid content and vitamin C levels in tomatoes[39]. Additionally, SeMet regulates amylose synthesis and optimizes the processing quality and flavor of rice[13]. This study demonstrates that flavonoids are the main component through which SeMet enhances the nutritional quality of kale.
BroMYB34 as an upstream activator of BroCHSs to promote flavonoid synthesis and antioxidant capacity
-
Transcription factors participate in plant growth processes through sequence-specific promoter interactions[40]. MYB transcription factors form a major regulatory network in plants, constituting one of the largest and most functionally diverse TF families, functioning as master regulators of secondary metabolites synthesis as well as biotic and abiotic stress responses[41,42]. MYB proteins feature a conserved N-terminal DNA-binding domain and a variable C-terminal regulatory region, which play important roles in DNA binding[43,44] and the regulation of protein activity, respectively. Flavonoid biosynthesis in plants is coordinately regulated by conserved transcriptional networks, with MYB-type transcription factors serving as central regulators that directly bind to cis-acting elements in structural gene promoters of the flavonoid biosynthesis pathway and regulate the expression of downstream genes[20,45,46]. CHS catalyzes the formation of the main carbon skeleton of flavonoids, mediating the first committed step in the flavonoid synthesis pathway[47]. This study identified BroMYB34, a SeMet-responsive MYB transcription factor (Fig. 5f), as a key activator of flavonoid biosynthesis in kale. The Y1H assays demonstrated that BroMYB34 binds to the promoters of three BroCHS homologs (BroT26.C02gCHS, BroT26.C03gCHS3, BroT26.C09gCHS3). In contrast, dual-luciferase reporter assays confirmed its ability to transcriptionally activate the expression of all three BroCHS genes (Fig. 6). There are both transcriptional activation and inhibitory regions in the transcription structure of MYB transcription factors, which positively or negatively regulate flavonoid biosynthesis[26,48]. BroMYB34 was instantaneously overexpressed in kale; the temporal expression of BroMYB34 peaked at 3 d (Fig. 7a), while the flavonoid content peaked at 4 d (Fig. 7e, f). This delay likely reflects a hierarchical regulatory mechanism that BroMYB34 modulates downstream gene expression to activate flavonoid metabolic pathways, ultimately influencing flavonoid accumulation, suggesting the presence of a noticeable delay between the activation of the upstream regulatory factor and the substantial accumulation of the downstream metabolite. In this study, functional characterization of BroMYB34 indicated that its overexpression induced CHS expression, leading to enhanced flavonoid biosynthesis and consequent improvement of the antioxidant capacity of kale (Fig. 7).
In conclusion, this study demonstrated that exogenous application of 10 mg·L−1 SeMet to kale seedlings contributed to the accumulation of Se content in kale and established BroMYB34 as a positive regulator that transcriptionally activates the key genes for flavonoid synthesis, thereby stimulating flavonoid biosynthesis and enhancing antioxidant capacity in kale. In future research, an accurate SeMet application plan suitable for field production can be further established to enhance the nutritional value of kale, providing a feasible strategy for the development of selenium-rich functional vegetables via selenium biofortification. This study preliminarily reveals the mechanism by which SeMet regulates flavonoid biosynthesis in kale seedlings (Fig. 8). Subsequently, through targeted breeding or gene-editing approaches, superior varieties with high flavonoid content can be developed to meet rising consumer demand for antioxidant 'super vegetables'.
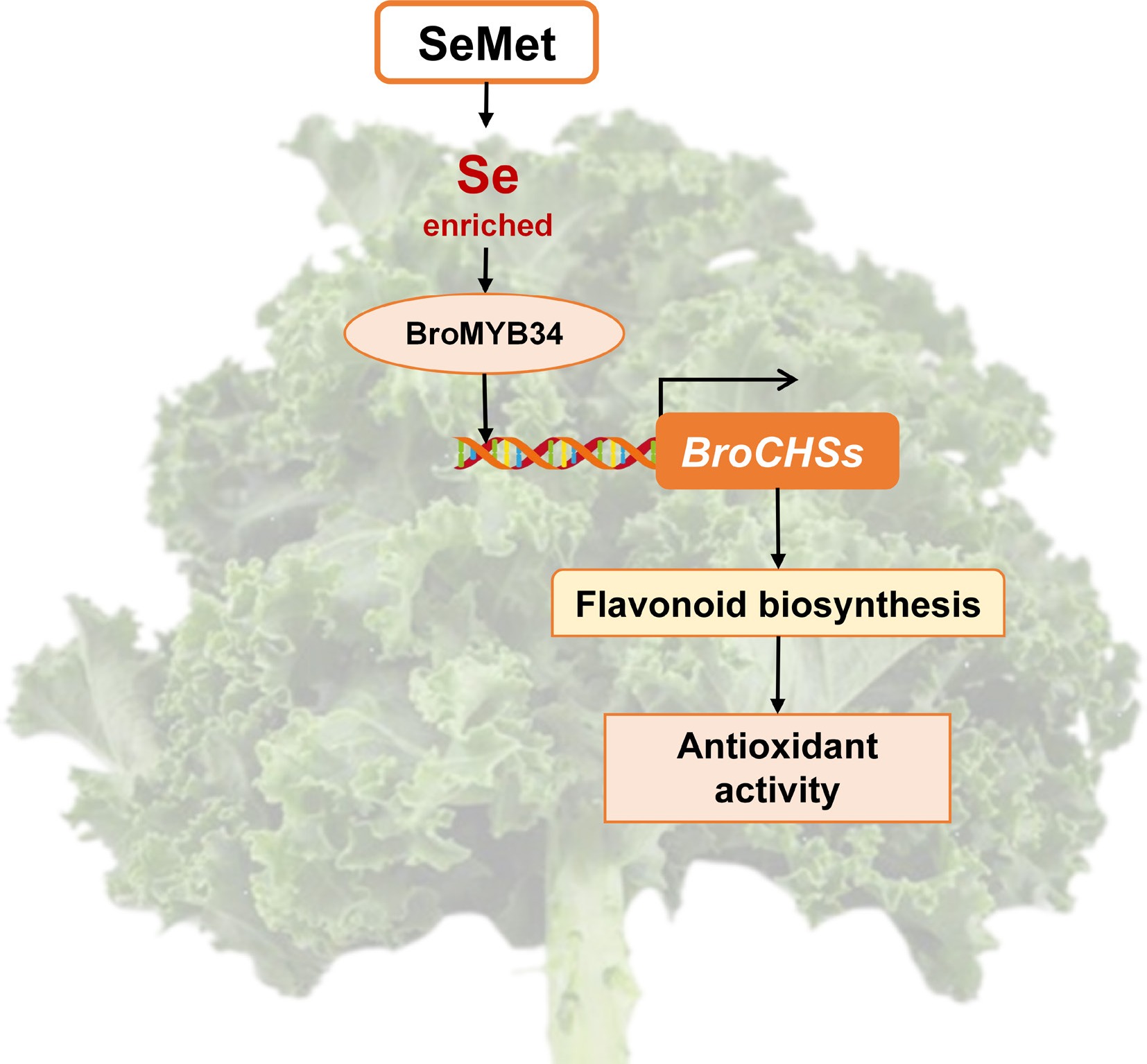
Figure 8.
Schematic diagram of the mechanism of exogenous SeMet regulating flavonoid accumulation to improve antioxidant activity in kale seedlings.
This study was supported by the Nature Foundation of Shandong Province, China (Grant No. ZR2022QC113), the Formation Mechanism and Regulation of Fruit and Vegetable Quality in Saline-alkali Land (Grant No. GYJ2023004), the Agricultural Science and Technology Innovation Project of Shandong Academy of Agricultural Sciences (Grant No. CXGC2025B20), the Modern Agricultural Industrial Technology System Funding of Shandong Province, China (Grant No. SDAIT-05), the Prospect of Shandong Seed Project, China (Grant No. 2023LZGC014), the China Agriculture Research System (Grant No. CARS-23-G14), and the Shandong Province Technology-based Small and Medium-sized Enterprise Innovation Capacity Improvement Project (Grant No. 2024TSGC0331).
-
The authors confirm their contributions to the paper as follows: study conception and design: Wang F; data collection: Fu X, Liu Y, Li X, Shu R, Yao T, Jiao J, Zhao X; analysis and interpretation of results: Fu X, Wang L, Wang F; draft manuscript preparation: Fu X, Li J, Zhang Y; supervision and validation: Li J, Zhang Y, Gao J, Wang F; funding acquisition: Wang L, Gao J, Wang F. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xin Fu, Yuxiang Liu
- Supplementary Table S1 The primer sequence.
- Supplementary Fig. S1 Effects of different exogenous concentration SeMet on the Se and pigment content of kale seedlings.
- Supplementary Fig. S2 Effects of different exogenous concentration SeMet on the quality of kale seedlings.
- Supplementary Fig. S3 Violin diagram of all the different flavonoids in the metabolome.
- Supplementary Fig. S4 Identification of DGEs in transcriptome by qRT-PCR.
- Supplementary Data S1 The summary of all the metabolite in the metabolome.
- Supplementary Data S2 The summary of all the DEMs in the metabolome treated by SeMet in kale.
- Supplementary Data S3 The summary of all the different flavonoid in the metabolome treated by SeMet in kale.
- Supplementary Data S4 The summary of all the DEGs in the transcriptome treated by SeMet in kale.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fu X, Liu Y, Wang L, Li X, Shu R, et al. 2025. Selenomethionine regulates flavonoid biosynthesis through the BroMYB34–BroCHSs module in kale (Brassica oleracea var. acephala). Vegetable Research 5: e037 doi: 10.48130/vegres-0025-0030 

Selenomethionine regulates flavonoid biosynthesis through the BroMYB34–BroCHSs module in kale (Brassica oleracea var. acephala)
- Received: 15 April 2025
- Revised: 01 July 2025
- Accepted: 06 August 2025
- Published online: 20 October 2025
Abstract: Kale is a super vegetable that is rich in a variety of nutrients and has antioxidant and anti-cancer properties. Selenomethionine (SeMet) is a type of organic selenium compound that enhances fruit quality; however, there is a lack of research on its application in kale. The study identified 10 mg·L−1 as the optimal SeMet concentration for kale, and selenium gradually accumulated after SeMet treatment, which significantly improved the antioxidant capacity of kale. Metabolome and transcriptome analysis of exogenous SeMet in kale revealed flavonoids as the main differential metabolites, and differential genes were most significantly enriched in flavonoid-related pathways. In addition, SeMet significantly induced the expression of MYB-type transcription factor BroMYB34. BroMYB34 binds to the BroCHSs promoter, a key gene for flavonoid synthesis, to activate its transcription. Transient overexpression of BroMYB34 could significantly increase the expression of BroT26.C02gCHS, BroT26.C03gCHS3, and BroT26. C09gCHS3 stimulates flavonoid synthesis to enhance the antioxidant capacity of kale. Overall, the findings of this study preliminarily reveal the molecular mechanism of SeMet promoting flavonoid biosynthesis of kale seedlings.
-
Key words:
- Kale /
- Selenomethionine /
- Flavonoid /
- MYB transcription factor /
- CHS