









-
The global prevalence of obesity has imposed a huge burden on modern society[1]. Obesity-related chronic low-grade inflammation is a significant factor in the advancement of non-alcoholic fatty liver disease (NAFLD) and its potential progression to non-alcoholic steatohepatitis (NASH)[2]. NASH has emerged as a prominent contributor to liver-related morbidity and mortality on a global scale[3]. Considering the limited availability of approved pharmacological treatments for obesity and NAFLD, significant attention has been directed towards promoting healthy lifestyle interventions, such as dietary modifications and regular physical activity, as crucial strategies.
Low-grade inflammation is extensively associated with the disturbances observed in energy metabolism pathways in individuals with obesity. Dysbiosis of the microbiota has been recognized as a significant contributor to obesity and its associated metabolic disorders. Moreover, it has been demonstrated to play a role in the advancement of systemic inflammation[4]. Clinical observations have provided evidence indicating a reduction in the diversity of gut microbiota and a decrease in the Bacteroidetes/Firmicutesratio among obese individuals. An increase in the phylum Firmicutes, specifically the genus Clostridium, was observed[5]. In HFD-fed rodents, Lactobacillus and Bifidobacterium resulted in improvements in hepatic steatosis and fibrosis. This improvement was found to be correlated with a reduction in the abundance of Desulfovibrio in feces[6]. Akkermansia muciniphila has the potential to improve inflammation and mitigate metabolic endotoxemia, adipose tissue inflammation, insulin resistance, and fat mass accumulation[7, 8]. Altered gut microbiota has been found to be linked to a reduction in the expression of short-chain fatty acids (SCFAs), which play a crucial role in preserving the integrity of the intestinal epithelial barrier, thus resulting in an elevation of bacterial translocation and inflammation, accompanied by a reduction in the expression of hormones that suppress hunger[9]. Lipopolysaccharides (LPS) generated by gram-positive bacteria within the intestinal lumen have been found to damage intestinal permeability[10], increase circulating LPS levels, and augment systemic inflammatory response[11]. It has been observed that endotoxemia is present in HFD-fed mice, and microbiota-derived LPS triggers the activation of obesity-related innate immune response[12]. Microbiota-derived endotoxin or LPS is associated with the activation of Kupffer cells, thereby influencing the progression of NAFLD[13]. Thus, the regulation of gut microbiota is important in terms of early prevention of obesity and NAFLD.
Some edible and medicinal plants can be used individually or in combination to regulate energy metabolism and promote weight loss. Polyphenols, flavonoids, oligosaccharides, polysaccharides, and various other metabolites are widely recognized as major functional components. Anti-obesity mechanisms of plants include suppressing appetite, reducing the digestion and absorption of lipids and carbohydrates, inhibiting the growth of white adipose tissue, inhibiting adipogenesis and lipogenesis, regulating lipid metabolism in adipocytes, increasing energy expenditure, regulating gut microbiota, and reducing obesity-related inflammation[14].
Tea is widely recognized as a popular and healthy beverage worldwide. According to the ISO 20715-2023 standard, tea can be categorized into six major types: green tea, white tea, yellow tea, oolong tea, black tea, and dark tea. Previous studies have shown that tea could provide health benefits against obesity, cardiovascular disorders, insulin resistance, diabetes, and various types of cancers by exerting antioxidant and anti-inflammatory activities. Catechins, especially (−) Epigallocatechin-3-gallate (EGCG), are recognized as one of the most important bioactive components[15,16]. EGCG could also regulate bile acid signaling and increase the abundance of A. muciniphila, thereby altering body weight and bile acid composition[17]. During tea manufacturing, catechins undergo a gradual conversion into theaflavins, thearubigins, and theabrownine. This conversion is facilitated by polyphenoloxidase from tea leaves and enzymes produced by microorganisms[18,19]. Meanwhile, the concentration of tea polysaccharides experienced a significant increase, whereas the molecular mass exhibited a noticeable decrease throughout the process of pile fermentation of dark tea[20]. Recently, there has been growing interest in the potential of fermented tea enriched with polysaccharides or theabrownine as a regulator of gut flora and metabolic health. Fu-brick tea polysaccharides exhibited a dose-dependent inhibition of body weight gain and mitigated gut dysbiosis in mice with diet-induced obesity. Lactobacillus, Parabacteroides, A. muciniphila, Bifidobacterium, and Roseburia exhibited enrichment in the gut, leading to significant microbial shifts that played a role in adipose browning and thermogenesis. This, in turn, alleviated host adiposity, glucose homeostasis, dyslipidemia, and the associated hepatic steatosis[21]. Fu-brick tea polysaccharides also inhibit Desulfovibrio and stimulate the synthesis of short-chain fatty acids (SCFAs), which are recognized as potential prebiotic agents[22]. Theabrownine from Puerh tea suppresses microbes associated with bile-salt hydrolase (BSH) activity, leading to an elevation of ileal conjugated bile acids (BAs) and suppression of the intestinal FXR-FGF15 signaling pathway. As a consequence, there is an elevation in hepatic synthesis and elimination of bile acids, a decline in hepatic cholesterol levels, a decrease in lipogenesis, an enhancement in energy metabolism in both white and brown adipose tissue, and ultimately amelioration of obesity induced by a high-fat diet[23,24].
JHXY is a novel type of Chinese tea produced in the Fujian province of China. 'Yongchun Foshou', a traditional tea plant used to produce oolong tea, was selected as the raw material for JHXY. After processing into oolong tea, it was roasted at 120 °C for 2−4 h, and was subsequently exposed to fermentation facilitated by environmental E. cristatum. This step is traditionally regarded as essential in the production of Fu-brick tea. Following an adequate fermentation process, the counts of E. cristatum in JHXY products can reach up to 1 × 106 cfu/g and the polysaccharide content experienced a significant increase from 1.20% to 3.90%. Conversely, the content of polyphenols and ester catechins experienced a decrease from 13.33% to 8.70% and from 11.9 mg/g to 4.23 mg/g, respectively. The amount of theabrownine in JHXY was found to be lower compared to that in ripened Puerh tea. The distinctive chemical composition of JHXY leads to a unique regulatory mechanism on gut microbiota, distinguishing it from ripened Pu-erh tea.
Our study endeavors to examine the potential of JHXY in offering protection against obesity and early hepatic liver fat accumulation. A two-week diet consisting of high sucrose and high fat was administered, which was then followed by a ten-week period of interventions. Weight gain, energy intake, and plasma lipids were assessed in the study. Following the euthanization of the rats, the adipose tissue weights were recorded, and the liver was examined for pathological changes. Gut microbiota were analyzed. The discussion revolved around determining the optimal dosage, taking into consideration the evidence pertaining to metabolic health and intestinal homeostasis.
-
The leaves of 'Yongchun Foshou' were primarily made into oolong tea and the relative processes were taken in reference to ISO 20716:2022. Specifically, the fresh leaves underwent sun fixation for 0.5 h, followed by four rounds of rocking and cooling. After stir fixation at 280 °C for 8 min, the tea leaves were rolled into shapes and dried at 130 °C for 30 min to get 'Yongchun Foshou' oolong tea. To produce "Jinhua Xiangyuan", 'Yongchun Foshou' oolong tea was roasted at 120 °C for 2−4 h, then steamed and piled up for 14−21 h, maintaining a core temperature of 42−50 °C and an air humidity of 68%−81%. After undergoing pile fermentation, the tea materials were steamed for 28−126 s at 104−120 °C and then pressed into standard-sized brick tea. Then the brick tea was placed in the fermentation room with 1.8−2.5 cm gaps. The fermentation process lasted for approximately 18−22 d. During this period, the room temperature and relative humidity were controlled at 25−33 °C and 58%−82%, respectively. Then, the room temperature was increased by 1.5−2 °C per day until it reached the final temperature of 42 °C, in order to allow the bricks to dry for 14−16 d until completion (Fig. 1). The water content of the final products was approximately 4%−5% (CN107410589A). The main components in JHXY raw materials and products are shown in Table 1. Polysaccharides, total polyphenols, and flavonoids were detected using standard methods.
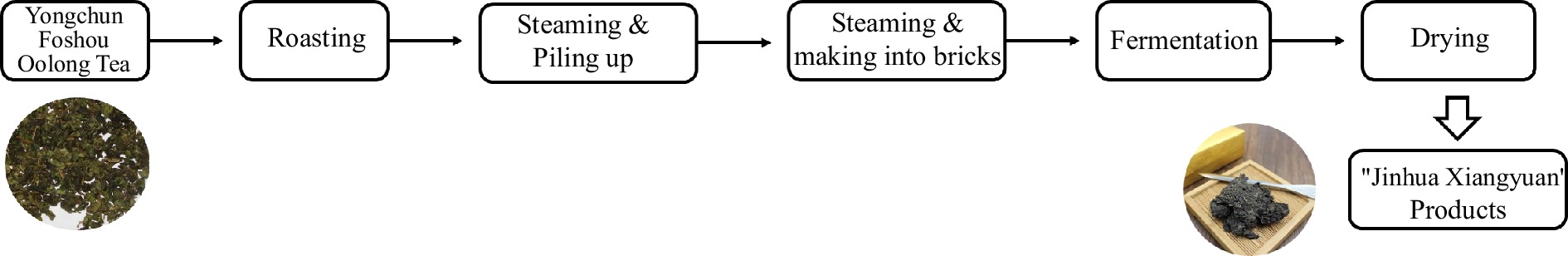
Figure 1.
The process of JHXY products.
Table 1. Main components in JHXY raw materials and products.
Raw materials Products C (mg/g) 0.92 0.49 EC (mg/g) 1.96 0.64 EGC (mg/g) 5.72 3.53 ECG (mg/g) 1.08 0.43 EGCG (mg/g) 10.82 3.80 TFs (mg/g) 1.20 0.50 Tea polysaccharides (%) 1.20 3.90 Reducing sugar (%) 6.26 1.25 Caffeine (%) 1.81 1.58 Total flavonoids (%) 4.37 2.90 Polyphenol (%) 13.33 8.70 Experimental rat treatment
-
Eighty seven six-week-old male SD rats were purchased from Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). Rats were housed in a specified pathogen-free environment under monitored conditions of a 12/12-h light/dark cycle, a temperature of 23 °C ± 2 °C, and a humidity level of 40% ± 10%. Following a 1-week acclimation period, 12 out of 87 subjects were randomly selected as the blank control group (BC) and were fed a normal maintenance diet. The other 75 rats were fed a high-sucrose, high-fat diet (HFD) containing 15% glucose and 15% fat from Spaifu Biotechnology Co. Ltd., Beijing, China, for 2 weeks. 55 out of 75 rats with higher weight gain were randomly divided into five groups: HFD (MC), HFD + JHXY tea infusion at different doses (LI: 0.4 g/kg·bw, MI: 0.8 g/kg·bw, HI: 1.6 g/kg·bw), and HFD + Simvastatin group (PC) was used as the positive control (10 mg/kg·bw).
The aforementioned six groups were fed an experimental diet with appropriate interventions for 10 weeks. Body weight and daily energy intake were monitored weekly, while relative serum indexes were detected every two weeks. Animals were sacrificed after a 10-week intervention. All the animals were euthanized under anesthesia after fasting for 12 h. The WAT index and BAT index were calculated using the following formulas: WAT index = WAT weight / body weight × 100%; BAT index = BAT weight / body weight × 100%.
Rats serum index
-
Levels of total cholesterol, triglycerides, high-density lipoprotein cholesterol, and low-density lipoprotein cholesterol were determined using the Cobas 6000 modular chemistry analyzer (Roche Co., Mannheim, Germany).
Histopathology assay
-
Sections of hepatosomatic samples were fixed in a 10% formalin buffer and processed for paraffin embedding. Sections were stained with hematoxylin-eosin (HE) for histological observation. To visualize the neutral triglycerides and lipids in hepatic samples, frozen sections were prepared and fixed with 4% paraformaldehyde for 15 min. After treating with 60% isopropanol, the sections were further treated with Oil Red O (Sigma-Aldrich) dissolved in 60% isopropanol. Photos were taken, and the absorbance at 520 nm was then measured (Imaging mass spectrometry for lipidomics).
16S ribosomal RNA V3-V4 region sequencing
-
Fecal samples were collected before sacrifice by tail suspension. Fecal DNA extraction was performed following the protocol of the MOBIO PowerSoil® DNA Isolation Kit 12888-100. The extracted DNA was stored at −80 °C in a Tris-EDTA buffer solution until further use and subsequently analyzed by 16S ribosome RNA gene sequencing[25]. Raw sequences were processed into operational taxonomic units (OTUs). OTUs were then matched with Greengenes databases and alpha and beta diversity were also performed[26]. Spearman analysis was conducted to explore the correlation. A p value < 0.05 of rho was used to perform the correlation network using the OmicStudio tools at
www.omicstudio.cn .Statistical analysis
-
Data were presented as the mean ± SD and were analyzed using ANOVA analysis. All statistical analyses were conducted using the SPSS 19.0 statistical software package, and a p value of < 0.05 was considered statistically significant. Rats were randomized in a blinded manner. Histopathology observations were made using a random selection process.
-
During the 10-week intervention, rats in the MC group exhibited a greater increase in body weight compared to the rats in the BC group. Rats in the LI, MI, and HI groups exhibited a slower rate of body weight gain compared to the MC group from the 4th to the 8th weeks. In the 10th week, the rats in the MC group exhibited a statistically significant increase in body weight (492.17 ± 34.41 g, p < 0.05) when compared to the BC group (561.73 ± 34.41 g). The body weight of rats in the LI group were measured to be 527.02 ± 16.91 g (p < 0.05), while those in the MI group were 514.36 ± 23.89 g (p < 0.01), and in the HI group were 507.36 ± 22.83 g (p < 0.01) (Fig. 2a, b). Therefore, the administration of JHXY at a dosage range of 0.4−1.6 g/kg·bw has been shown to effectively mitigate the excessive weight gain induced by the HFD.
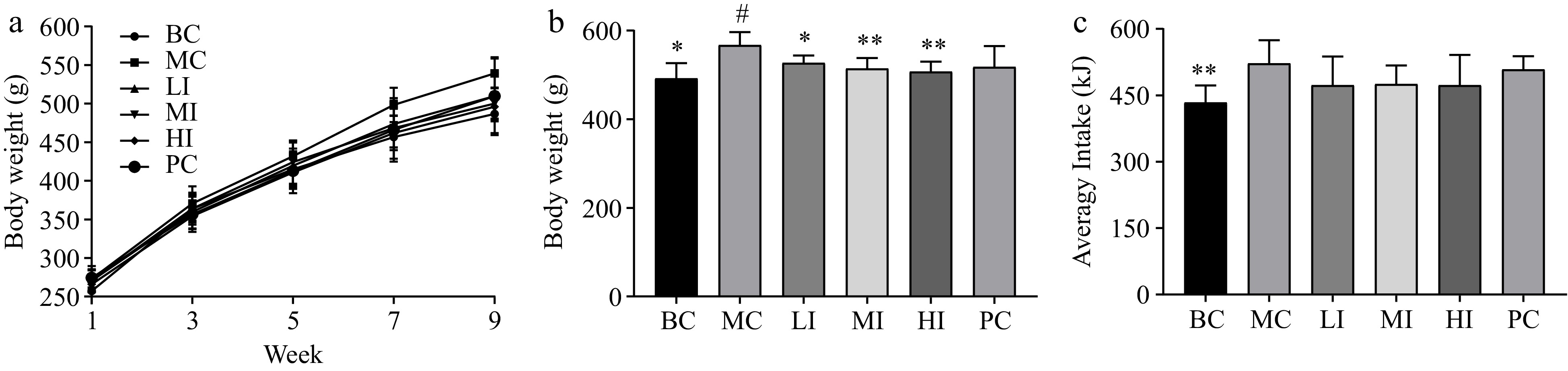
Figure 2.
Effects of JHXY on body weight and food intake. (a) The weight gain of rats in the BC, MC, LI, MI, HI, and PC groups. (b) The rats' body weight in the 10th week. The differences were denoted as follows: # p < 0.05, compared to the BC group; * p < 0.05, ** p < 0.01 compared to the MC group. (c) The average food intake of rats in BC, MC, LI, MI, HI, and PC.
However, the JHXY tea infusion did not exhibit a significant reduction in food intake. This finding suggests that the weight control effect of JHXY cannot be solely attributed to appetite suppression, as depicted in Fig. 2c.
Effects of JHXY on fat mass index and distribution
-
As shown in Fig. 3, the BAT index in the MC group (0.09% ± 0.01%) exhibited a significant decrease when compared to the BC group (0.11% ± 0.01%, p < 0.05). The JHXY intervention at a dosage of 0.8−1.6 g/kg·bw effectively mitigated the reduction in the BAT index observed in the MI (0.12% ± 0.01%, p < 0.05) and HI (0.11% ± 0.01%, p < 0.05) groups. The BAT index in the MI group exhibited the highest value in comparison to the other groups (Fig. 3a). The WAT index in the MC group (5.68% ± 0.36%) exhibited a statistically significant increase when compared to the BC group (3.82% ± 0.20%, p < 0.05). Additionally, the WAT index exhibited a significant decrease in both the MI (4.56% ± 0.28%, p < 0.05) and HI (4.54% ± 0.26%, p < 0.05) groups, when compared to the MC group. The WAT index in the HI group exhibited the lowest value in comparison to the other groups (Fig. 3b).
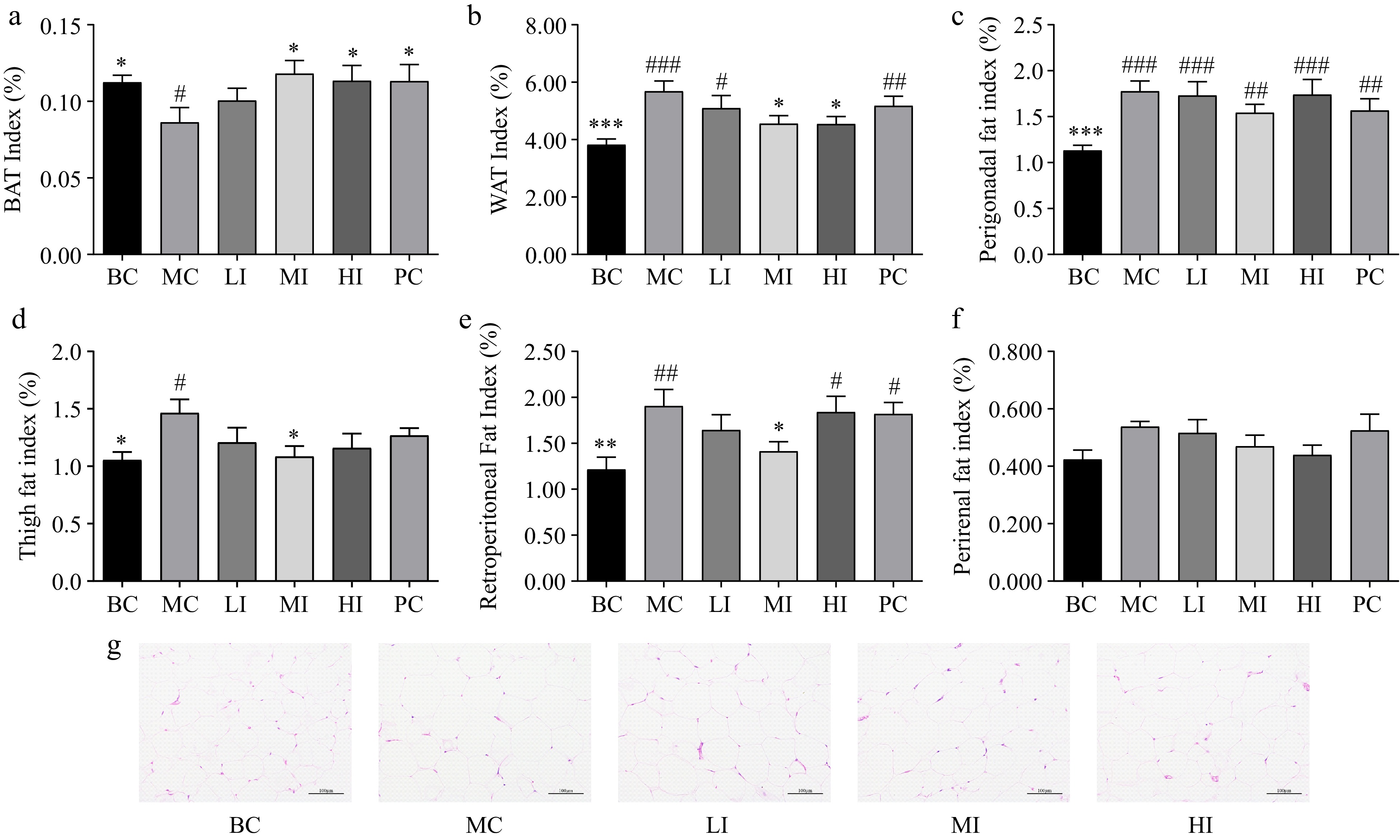
Figure 3.
Effects of JHXY on fat mass index and distribution. (a) - (f) Effects of JHXY tea on brown adipose tissue (BAT) index, white adipose tissue (WAT) index, perigonadal fat index (PGFI), thigh fat index (TFI), retroperitoneal fat index (RPFI), and perirenal fat index (PRFI) after sacrifice. The differences were denoted as follows: # p < 0.05, ## p < 0.01, ### p < 0.001 versus the BC group; * p < 0.05, ** p < 0.01, *** p < 0.001 versus the MC group. (g) The histology of the WAT from rats treated with BC, MC, LI, MI, and HI for 10 weeks. (Magnification, 200×)
JHXY exhibited a more pronounced impact on the reduction of visceral fat mass in comparison to the reduction of thigh fat mass (Fig. 3). In the MI group, there was a significant decrease in retroperitoneal fat index (RPFI, Fig. 3e), perigonadal fat index (PGFI, Fig. 3c), and thigh fat index (TFI, Fig. 3d). In the HI group, JHXY exhibited a significant reduction of white fat, while no significant effect was observed on fat deposits in specific regions of the body. The JHXY intervention demonstrated more significant reduction in visceral fat, compared to thigh fat (Fig. 3d). Among the tested subjects, JHXY exhibited the highest effect at a dosage of 0.8 g/kg·bw.
Effects of JHXY on blood lipid profiles
-
In comparison to the MC group, the JHXY intervention groups exhibited a reduction in plasma lipid levels, including TC, TG, and LDL-C, while the level of HDL-C increased. A relationship that is dependent on the dose was observed, although there was no statistically significant difference between the MI and HI groups (Fig. 4a−d). Hepatocyte morphology and the presence of fat droplets in hepatocytes were examined by HE staining (Fig. 5a) and oil O staining (Fig. 5b). The sizes of fat droplets and the counts of hepatic cells in the MC group exhibited a statistically significant increase compared to the BC group. The JHXY intervention at a dosage of 0.8 g/kg·bw demonstrated the highest potential in mitigating the effects caused by an HFD in the MI group. 1.6 g/kg·bw of JHXY intervention in the HI group did not yield superior results compared to 0.8 g/kg·bw of JHXY (Fig. 3g).
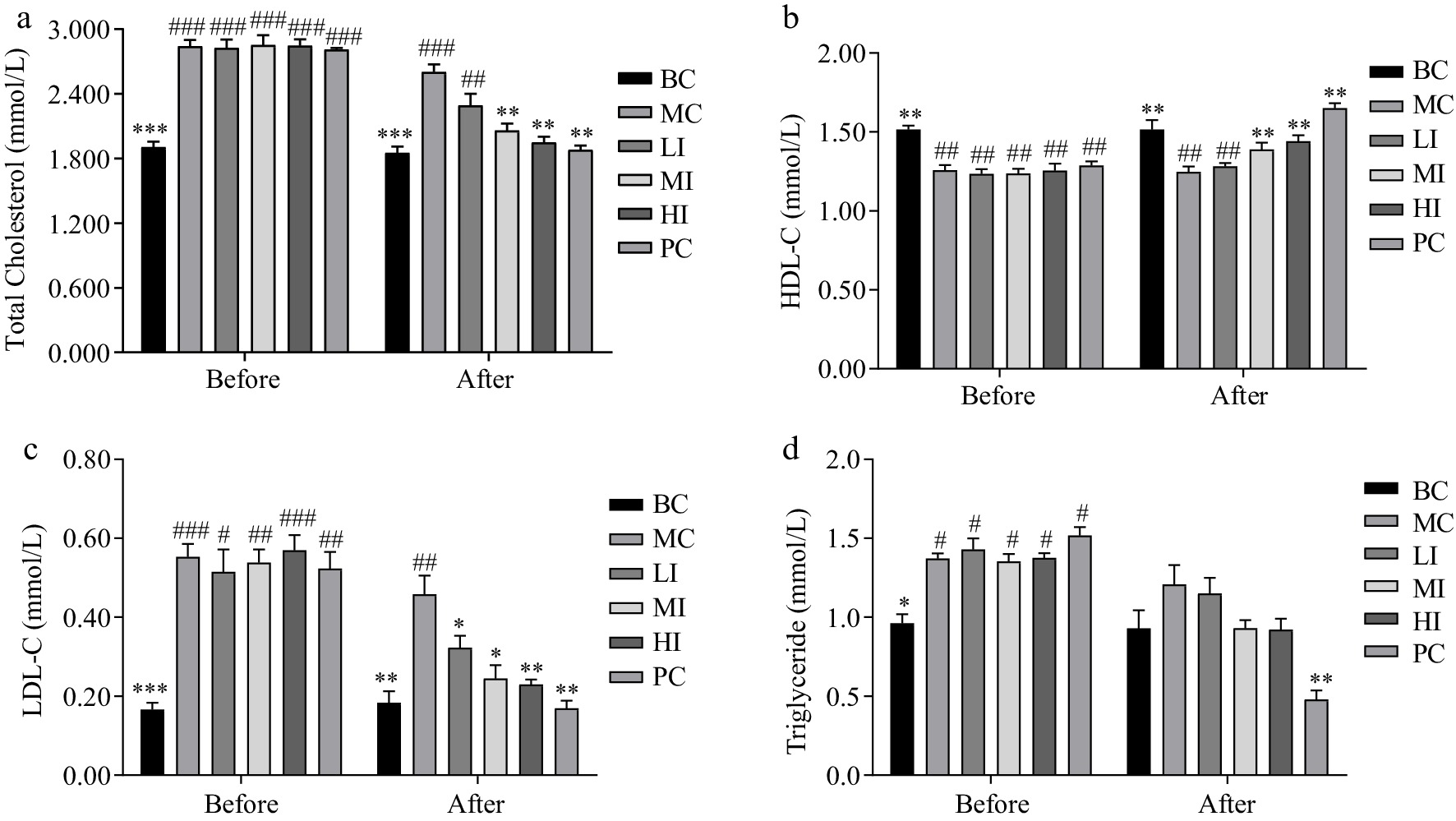
Figure 4.
Effects of JHXY on blood lipid profiles. (a) TC, (b) HDL-C, (c) LDL-C, and (d) TG levels before and after JHXY intervention. The differences were denoted as follows: # p < 0.05, ## p < 0.01, ### p < 0.001 compared to the BC group; * p < 0.05, ** p < 0.01, *** p < 0.001 compared to the MC group.
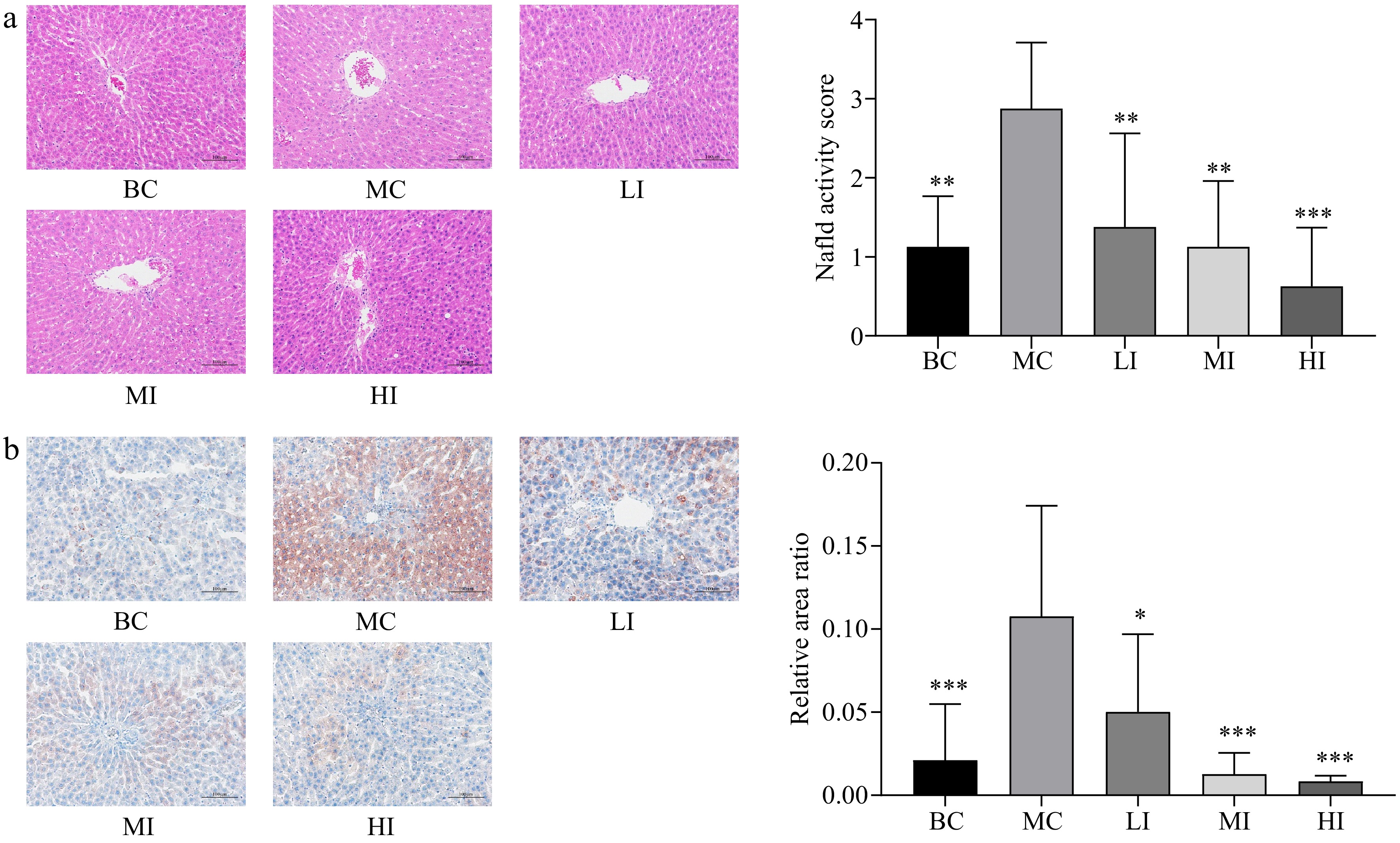
Figure 5.
Histology and oil red staining of the liver. (a) The histology of the liver from rats treated with BC, MC, LI, MI, and HI for 10 weeks. (Magnification, 200×); NAFLD scores were analyzed. The differences were denoted as follows: * p < 0.05, ** p < 0.01, *** p < 0.001 compared to the MC group. (b) The liver was stained with oil O red to observe the effects of different treatments on rats. (Magnification, 200×); the relative area ratio of positive areas was analyzed using Image J software. The differences were denoted as follows: * p < 0.05, ** p < 0.01, *** p < 0.001 versus the MC group.
Effects of JHXY on gut microbiota
-
The α-diversity indices, namely Chao1 and Shannon, were used to assess species diversity. The results indicated that there were no statistically significant differences observed among the different groups (Fig. 6a). As depicted in Fig. 6b−c, the dominant phylum in each group was Firmicutes, followed by Actinobacteriota and Bacteroidota. The Firmicutes ratio exceeded 80%. Compared to the control group, HFD significantly enhances the relative abundance of Desulfobacterota (p < 0.01). On the contrary, the administration of middle and high doses of JHXY tea infusion results in a significant reduction in the abundance of Desulfobacterota (p < 0.05). At the genus level, Lactobacillus exhibited the highest abundance. A significantly higher abundance of Bifidobacterium (p < 0.001) and a lower abundance of Lactobacillus were observed in the MC group compared to the BC group. Middle and high dose JHXY intervention for a duration of 10 weeks has been found to significantly enhance the abundance of Lactobacillus. The abundance of Bifidobacterium in the HI group exhibited a statistically significant increase when compared to the MC group (p < 0.05). Blautia exhibited a significant increase in the LI group as compared to the MC group.
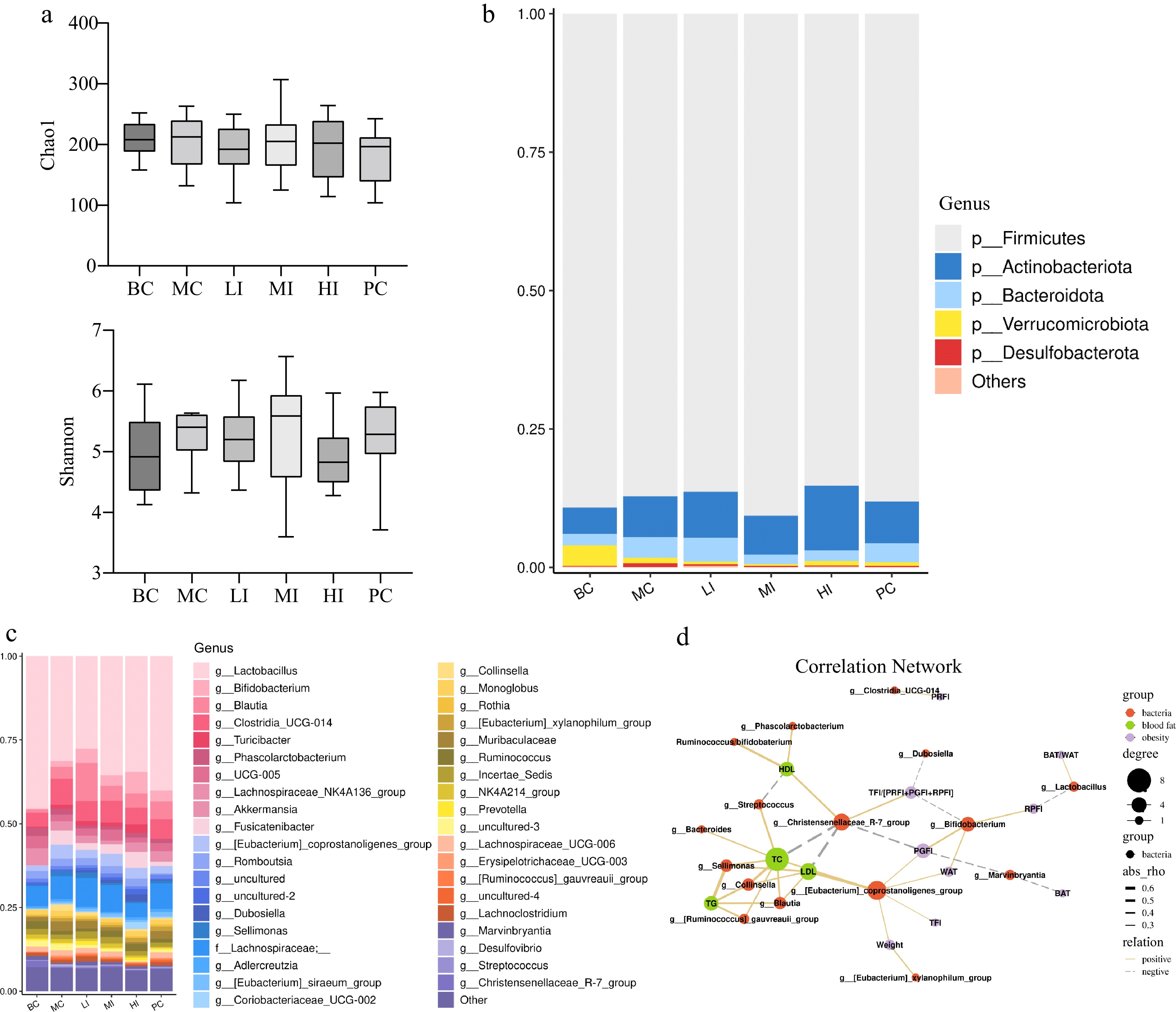
Figure 6.
Effects of JHXY on gut microbiota. (a) The α-diversity of gut microbiota in various groups. (b) Relative abundance of bacteria at the phylum level. (c) Relative abundance of bacteria at the genus level (top 40 genera analysis). (d) Spearman correlation analysis of microorganisms and physical indexes (solid line indicates positive correlation, dotted line indicates negative correlation).
The metabolic pathways were predicted using PICRUSt 2, and a statistical analysis revealed significant differences in 22 pathways (p < 0.05). Compared to the control group (MC group), the administration of a moderate dose of JHXY led to a significant decrease in the expression of amino acid-related enzymes, as well as the down regulation of the lipopolysaccharide biosynthesis pathway and lipopolysaccharide biosynthesis proteins signaling pathway (p < 0.05). Nevertheless, the administration of high-dose JHXY has been shown to enhance the bacterial toxin production, biosynthesis of siderophore group nonribosomal peptides pathways, and exert inhibitory effects on the sulfur relay system pathway (p < 0.05).
Observed interactions of anti-obesity, hypolipidemia effects and gut microbiota
-
Correlation analysis between gut flora and blood biochemical indices was performed, based on data from the BC, MC, and JHXY intervention groups (Fig. 6d). Blood lipid and obesity-related indexes were categorized into two distinct clusters. Key bacteria were classified into three types, namely those linked to blood lipid profile, fat indexes, or both. The abundance of Collinsella, Blautia, Sellimonas, and Ruminococcus_gauvreauii_group exhibited a positive correlation with TC, TG, and LDL-C. The abundance of Lactobacillus exhibited a positive correlation with the ratio of BAT/WAT, while displaying a negative correlation with the relative proportion of RPFI. Bifidobacterium aboundance was negatively related to TFI/(PRFI+PGFI+RPFI), but positively related to WAT, PGFI, and RPFI. The abundance of the Christensenellaceae_R-7_group and [Eubacterium]_coprostanoligenes_group was associated with both lipid profile and obesity indicators, but showed opposite effects. The abundance of the [Eubacterium]_coprostanoligenes_group were positively related to LDL-C, TC, weight gain, TFI, WAT, and PGFI. The abundance of Christensenellaceae_R-7_group was positively related to protective indicators such as HDL-C and TFI/(PRFI+PGFI+RPFI), while being negatively related to risk indicators such as TC, LDL-C, and PGFI. The ratio of Ruminococcus to Bifidobacterium was positively related to HDL-C.
-
In the present study, the efficacy of JHXY tea in mitigating hyperlipidemia and controlling body weight was investigated in obese rats induced by a high-sucrose high-fat diet. Additionally, there were observed reduction in hepatosteatosis and stimulation of adipose browning. In the previous studies, it was observed that both oolong tea and Fu-brick tea exhibited a protective effect against obesity phenotypes induced by a high-fat diet. This protective effect was accomplished through the promotion of adipose browning and thermogenesis, in conjunction with the modulation of the gut microbiota[21, 27,28]. Our study has substantiated the preservation of effects following the innovative production process of JXHY. This process enables the fermentation of E. cristatum after the primary processing of oolong tea.
Correlation networks have unveiled diverse associations between gut microbiota and blood lipids or obesity. Lactobacillus counts exhibited a positive correlation with the ratio of BAT to WAT. Previously, high-fat diet-fed mouse models demonstrated that the administration of the probiotic bacterium Lactobacillus amylovorus KU4 (LKU4) resulted in increased mitochondrial function, as well as enhanced expression of thermogenic genes in WAT[29]. Hence, the increase in Lactobacillus population following the consumption of JHXY could potentially exert a significant impact on the process of white fat browning and energy expenditure. Lachnospiraceae_UCG-006 exhibited an increase following the administration of pear pomace to HFD-fed mice and its correlation with metabolic advantages was observed[30]. Further investigation is warranted to explore the mechanisms by which JHXY exerts its effects through Lachnospiraceae_UCG-006.
The Christensenellaceae_R7_group exhibited a positive correlation with protective indicators, including HDL-C and TFI/(PRFI+PGFI+RPFI), while displaying a negative correlation with risk indicators. Previously, the MEC Adiposity Phenotype Study (MEC-APS) revealed that the weight loss effects observed in individuals were attributed to the anti-inflammatory properties of specific dietary components. The relationship between the E-DII score and total fat mass was found to be mediated by LPS, and a negative correlation was observed between the E-DII score and the Christensenellaceae R-7 group[31]. In the context of polycystic ovary syndrome, a significant correlation has been observed between Christensenellaceae and BMI in women, indicating a strong connection between the microbiome and metabolic disease[32]. In this study, the administration of JHXY resulted in the attenuation of the lipopolysaccharide pathway and the enhancement of the proliferation of Christensenellaceae_R-7_group in the MI group. The weight loss effects observed in this study may be attributed to the anti-inflammatory properties of JHXY.
The investigation focused on examining the dose-effect relationship of JHXY in order to determine the optimal dosage that can effectively balance the metabolic health benefits and maintain intestinal homeostasis. PICRUSt2 prediction revealed a significant weakening (p < 0.05) of the lipopolysaccharide biosynthesis pathway and the lipopolysaccharide biosynthesis proteins signaling pathway in the 0.8 g/kg·bw group. In the group receiving a dosage of 1.6 g/kg·bw, it was observed that the sulfur relay system pathway was attenuated, whereas the bacterial toxins pathway was augmented (p < 0.05). The abundance of Desulfovibrio exhibited a dose-dependent decrease after JHXY intervention, which can be attributed to the inhibition of the sulfur relay system pathway. Thus, a dosage of 0.8 g/kg·bw was found to be more advantageous for maintenance of the gastrointestinal homeostasis. Based on body surface area conversion algorithm, it was 7−8 g JHXY for adults per day, which was brewed in 200−400 mL of boiled water for at least 1 min before ingestion.
The role of Bifidobacterium and A. muciniphila in this study was found to be ambiguous. A significant positive correlation between the presence of Bifidobacterium and obesity was identified in CCA analysis. In previous studies, it was observed that the levels of Bifidobacterium and A. muciniphila were higher in lean individuals, and exhibited an inverse relationship with plasma lipopolysaccharide levels[33,34]. However, in this study, it was observed that the levels of Bifidobacterium and A. muciniphila exhibited a general increase in a dose-dependent manner in the JHXY group. Interestingly, the MI group showed a significant inhibition of the LPS-related pathway, whereas no such inhibition was observed in the HI group. The ratios of Ruminococcus/Bifidobacterium, Ruminococcus/A. muciniphila, Ruminococcaceae/Bifidobacterium, and Ruminococcaceae/A. muciniphila were found to reach the highest values in the MI group, in comparison to the LI and HI groups. Acetate and lactate were identified as significant metabolites produced by Bifidobacterium. These metabolites served as substrates for the synthesis of propionic acid and butyrate in Ruminococcus and Ruminococcaceae. Butyrate plays a crucial role in safe guarding the integrity of the intestinal mucosal barrier and preventing the translocation of toxins into the circulatory system. Considering excessive levels of acetate and lactate could lead to fatty acid synthesis and mediate obesity-induced inflammation and systemic insulin resistance[11, 35], the balance between butyric acid synthesis bacteria and Bifidobacterium could be important. On the other hand, mucin-degrading A. muciniphila has recently been recognized as safe and beneficial for glycolipid metabolism. It has also been found to contribute to the protection of gut barrier function, but only at low counts and not at high counts[36]. Thus, the intricate relationship and balance between A. muciniphila and butyric acid synthesis bacteria could also be necessary for achieving intestinal homeostasis in the case of long-term consumption.
Previous studies have demonstrated that the consumption of oolong tea and Fu-brick tea typically leads to an increase in the abundance of Lactobacillus and A. muciniphila, while simultaneously reducing the abundance of Desulfovibrio. Polyphenols derived from Fu-brick tea were found to specifically enhance the abundance of A. muciniphila. However, during processing, the level of polysaccharides of JHXY significantly increased, while the levels of tea polyphenols decreased. Thus, polysaccharides should also be emphasized as important functional components of JHXY. Synergistic effects may potentially exist among JHXY polysaccharides, polyphenols, and other unidentified constituents[22, 37,38]. Theabrownin, isolated from ripened Puerh tea, strongly affects bile acid and fatty acid metabolism through the intestinal FXR-FGF15 signaling pathway[23]. JHXY theabrownin was lower than ripened Puerh tea, which may explain the lack of apparent interaction with bile acid systems.
Numerous studies have reported that various types of Chinese tea, particularly green tea, exhibit potential advantages in terms of weight management and glycolipid metabolism. The distinctiveness of JHXY in comparison to other varieties of Chinese tea lies in its lower concentrations of ester catechins and caffeine, which are attributed to the fermentation process. As a result, JHXY could be more gentle toward the gastrointestinal tract.
-
Our study has gathered substantial evidence supporting the recognition of a novel type of tea, known as "Jinhua Xiangyuan". The administration of JHXY at doses of 0.8 and 1.6 g/kg·bw resulted in a significant reduction in the levels of TG, TC, and LDL-C in obese rats. Additionally, JHXY led to a dose-dependent increase of HDL-C level. The treatment of JHXY at a dosage of 0.8 and 1.6 g/kg·bw resulted in a reduction in both the size and number of fat droplets in hepatic cells. Hepatocyte morphology was observed to be restored in the group treated with JHXY at a dosage of 0.8 g/kg·bw. The abundance of Bifidobacterium exhibited a significant increase, whereas the abundance of Bacteroidetes demonstrated a significant decrease across all JHXY treatment groups. Correlation analysis revealed a positive association between the Ruminococcus/Bifidobaterium ratio and HDL-C. The abundance of Christensenellaceae_R-7_group was negatively correlated with TC and LDL-C, while positively correlated with HDL-C. The abundance of Lactobacillus was positively correlated with BAT/WAT ratio. PICRUSt2 prediction revealed a significant weakening (p < 0.05) of the lipopolysaccharide biosynthesis pathway and the lipopolysaccharide biosynthesis proteins signaling pathway in the 0.8 g/kg·bw group. In the group receiving a dosage of 1.6 g/kg·bw, it was observed that the sulfur relay system pathway was attenuated, whereas the bacterial toxins pathway was augmented (p < 0.05). The findings of our research indicate that JHXY has the potential to be a beneficial option for individuals who are overweight or suffer hyperlipemia. This is particularly relevant for those who are unable to tolerate green tea or have an aversion to the taste of ripened Pu-erh tea. For individuals aiming to improve intestinal homeostasis, it is suggested that they consume a daily dosage of 7−8 g of JHXY, which was brewed in 200−400 mL of boiled water for at least 1 min before ingestion. This dosage and preparation method are considered optimal for adults in need of improving intestinal homeostasis. Polysaccharides should be highlighted as significant functional constituents of JHXY.
-
The authors confirm contribution to the paper as follows: study conception and design: Ying J, Xiao J, Chen Z, Li S, Niu X; data collection: Xiao J, Hou C, Fan Y, Qin J; analysis and interpretation of results: Xiao J, Ying J, Qin J, Xiang S, Hou C, Zhu X, Wang L, Hao B; draft manuscript preparation: Xiao J, Ying J, Chen Z, Xiang S. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are not publicly available due to management requests, but are available from the corresponding author on reasonable request.
-
All procedures were carried out in strict accordance with the guidelines and regulations approved by the Animal Care and Use Committee of the Chinese Center for Disease Control and Prevention (CDC).
This work was funded by Applied Basic Research Project of COFCO (2017-C1-T06), Yunnan Province Science and Technology Project (202102AE090038).
-
The authors declare that they have no conflicts of interest. Funding for this research was provided by COFCO, Ltd. The study results and data presented in this publication have been developed by and/or for Xiamen Tea Import and Export Co., Ltd. These results and data are considered proprietary and exclusive to Xiamen Tea Import and Export Co., Ltd, who reserves the right to use them for potential health claim requests.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xiao J, Chen Z, Xiang S, Zhu X, Hou C, et al. 2023. Anti-obesity and hypolipidemic effects of "Jinhua Xiangyuan" tea infusion in high-fat diet-induced obese rats. Beverage Plant Research 3:25 doi: 10.48130/BPR-2023-0025 

Anti-obesity and hypolipidemic effects of "Jinhua Xiangyuan" tea infusion in high-fat diet-induced obese rats
- Received: 10 July 2023
- Revised: 14 September 2023
- Accepted: 15 September 2023
- Published online: 11 October 2023
Abstract: "Jinhua Xiangyuan" (JHXY) is a unique type of Chinese tea made from roasted oolong tea and subsequently fermented by Eurotium cristatum. In this study, we investigated the potential benefits of JHXY on obese rats induced by a high-sucrose high-fat diet. JHXY infusion was administered orally at 1.6 g/kg·bw (HI), 0.8 g/kg·bw (MI), and 0.4 g/kg·bw (LI) per day for 10 weeks. Weight gain was significantly suppressed without affecting appetite. In the MI and HI groups, JHXY led to a significant reduction in triglycerides (TG), total cholesterol (TC), and low-density lipoprotein cholesterol (LDL-C), accompanied by an increase in high-density lipoprotein cholesterol (HDL-C) levels. A significant reduction in white adipose tissue (WAT) index and an increase in brown adipose tissue (BAT) index were observed. In MI group, the morphology of hepatocytes was restored. The abundance of Bifidobacterium was elevated, whereas the abundance of Desulfovibrio was significantly reduced. Correlation analysis revealed that the ratio of Ruminococcus to Bifidobacterium was positively correlated with HDL-C; the abundance of the Christensenellaceae_R-7_group was negatively correlated with TC and LDL-C, while positively correlated with HDL-C; the abundance of Lactobacillus was positively correlated with BAT/WAT ratio. In the MI group, the lipopolysaccharide biosynthesis pathway was predicted to be significantly weakened (p < 0.05); in the HI group, the sulfur relay system pathway was attenuated, whereas the bacterial toxins pathway was augmented (p < 0.05). Our research showed that JHXY has the potential to enhance metabolic health and maintain intestinal homeostasis. Optimal dosage should be a focal point in future research.