










-
LEA proteins were first identified in cotton seeds at the late embryonic development more than 30 years ago[1]. Following this discovery, these proteins have also been found in roots, stems, leaves, flowers and other tissues of plants. According to the similarity of amino acid sequences and conserved motifs, LEA proteins were divided into at least eight different groups (LEA1(PF03760), LEA2(PF03168), LEA3(PF03242), LEA4(PF02987), LEA5(PF00477), LEA6(PF10714), dehydrin (DHN) (PF00257) and seed maturation protein (SMP) (PF04927) in the Pfam database[2]. LEA proteins are widely distributed in higher plants[3−5]. To date, genome-wide identification and analysis of the LEA family genes have only been carried out in a few sequenced plant species genomes, such as Arabidopsis thaliana[4], Oryza sativa[6], maize[7], Solanum Tuberosum[8], cucumber[9], Solanum lycopersicum[10], Brassica napus[6], cassava[11], Populus trichocarpa[12], Prunus mume[13], Pinus tabulaeformis[14] and Canavalia rosea[15], linseed flax[16], peanut[17], Dendrobium officinale[5], Brassica campestris[18], Moso Bamboo[19], watermelon and melon[20], as well as in algae, fungi, bacteria[21]and even invertebrates[22].
Intrinsically disordered in their natural form, a large proportion of LEA proteins are predominantly characterized by their remarkable hydrophilicity, owing to the substantial presence of charged amino acid residues. These residues, along with glycine or other small amino acids such as alanine, serine, and threonine, exhibit a remarkable lack or, in some instances, minimal amounts of cysteine and tryptophan[23, 24]. LEA genes are considered pivotal players in the intricate processes of plant growth and development, as they are thought to assume an important role in alleviating the adverse effects of various stress conditions on cells. A study has unveiled intriguing insights into the dynamic nature of these genes, revealing that they exhibit an intriguing pattern of accumulation during the final stages of seed maturation, which coincides with the onset of dehydration. Specifically, these genes were found to be present in high concentrations during this phase, suggesting a potential role in safeguarding the developing seed against environmental perturbations. The results of the study also shed light on the impressive adaptive capabilities of LEA genes, as they were shown to be significantly induced under abiotic stress conditions, such as cold, heat, and drought stress[19, 25]. Several scientific investigations have propounded a compelling postulate that the LEA genes may potentially be implicated in a defense mechanism against a plethora of deleterious abiotic stressors. These mechanisms include, but are not limited to, the fortification and maintenance of the structural integrity of the membrane[26, 27], scavenging of free radicals[28, 29], sequestering ions[30] or biotin[31].
Somatic embryogenesis, a process of profound significance in the arsenal of modern biotechnological methodologies, has been widely recognized as a highly efficient technique for generating complete embryos that can subsequently develop into fully-fledged plants. It is characterized by two distinct pathways, namely the somatic embryogenesis direct pathway and the somatic embryogenesis indirect pathway. These pathways offer an unparalleled degree of relative genetic stability, reproducibility, and remarkable efficiency, rendering them as one of the most prominent technical means of achieving successful plant regeneration, aside from the syncytial embryogenesis pathway[32]. The significance of somatic embryogenesis extends far beyond the mere confines of basic research on cell differentiation, gene expression, hormone signaling, and metabolic regulation[33]. In addition to their profound implications for these domains of scientific inquiry, somatic embryogenesis also furnish an efficacious avenue for the large-scale clonal propagation of forest trees, the protracted conservation of finely-tuned germplasm, and the consequential transformation of genes[34].
Liquidambar L. species are literally multi-objective and multi-purpose tree species that possess a myriad of economic, ecological, and medicinal values. These trees are not only globally renowned for their outstanding forest resource status but also the main tree species in China's National Reserve Forest Construction Plan (2018−2035). L. formosana is recognized for its wide range of distribution, impressive adaptability to various ecological niches, and remarkable suitability as timber, L. styraciflua is characterized by a fast growth rate. With L. styraciflua as the female parent and L. formosana as the male parent, the hybrid sweetgum offspring demonstrates a salient and significant advantage over its parental counterparts, in fact, studies have confirmed that the growth velocity of this hybrid sweetgum exceeds that of L. styraciflua[35]. At present, hybrid sweetgum is mainly propagated and utilized by somatic embryogenesis, but its propagation efficiency needs to be further improved, the mechanism of somatic embryogenesis is still unclear, and there is a lack of research on the mining of key regulatory genes and their functions.
In this study, the genome-wide identification of the LsfLEA family genes were performed on the hybrid sweetgum genome and transcriptome using the Hidden Markov Model (HMM), and its sequence features, phylogenetic relationships, conserved motifs and gene structure were investigated. Meanwhile, the expression profiles of hybrid sweetgum LEA genes were analyzed in a series of developmental processes in non-embryogenic callus (NEC), embryogenic callus (EC), globular embryos (GE), heart-shaped embryos (HE), torpedo-shaped embryos (TE), cotyledonary embryos (CE), roots (R), stems (S), and leaves (L).This study provides new insights into the LEA family genes of hybrid sweetgum, which will contribute to the study of the function of genes related to somatic embryogenesis of hybrid sweetgum and the interpretation of the mechanism.
-
The SF15SH-5a embryonic cell line was induced from hybrid seeds obtained from L. formosana and L. styraciflua trees, as described previously[36]. The non-embryogenic callus (NEC) cells and embryogenic callus (EC) cells were previously reserved in our laboratory. To analyze the tissue- specific expression profile of the identified LsfLEA genes, each 90-mm plastic petri plate was inoculated with 10 clumps of EC on fresh induction medium of the same composition in three replicates and callus were transferred to fresh medium every 3−4 weeks, the medium was transferred to fresh media of the same composition monthly. The samples were incubated in an incubator at a constant temperature of (25 ± 2) °C and cultured in the dark. The EC obtained after induction for a period of time was transferred into the maturation medium in the same way as described above, embryos with different morphologies were collected as samples. At the stage of cotyledon embryo, they were moved to the germination medium in the tissue culture room of Beijing Forestry University (China), and cultured into plants. The roots and leaves were collected as materials for subsequent experiments. Composition of the medium for each stage are given in Table 1. All samples were immediately frozen in liquid nitrogen and stored at -80°C for subsequent gene expression analysis. Three independent biological replicate experiments were performed.
Table 1. Medium used in the experiment.
Medium Composition Induction/
proliferationBasic medium[37] + 1 mg 2, 4-D + 0.5 mg 6-BA + 40 g/L sucrose + 1 g/L enzymatic hydrolyzed casein 2.6−2.7 g/L vegetable gel, pH adjusted to 5.6~5.7 Mature Basic medium + 40 g/L sucrose + 6 g/L vegetable gel +5% PEG, pH was adjusted to 5.6~5.7 Germination Basic medium + 40 g/L sucrose + 2.6-2.7 g /L vegetable gel, pH was adjusted to 5.6~5.7 Identification and characterization of LEA genes in hybrid sweetgum
-
The HMM model's advantages over BLAST are significant. Its ability to handle probabilistic models with hidden states, flexibility in modeling different types of sequences, and accurate modeling of sequence variation within related sequences make it an invaluable tool for studying biological sequences. The hidden Markov models of LEA gene subfamilies (PF03760 (LEA1), PF03168 (LEA2), PF03242 (LEA3), PF02987 (LEA4) and PF00477) were downloaded from the Pfam database (
http://pfam.xfam.org/ ) (LEA5), PF10714 (LEA6), PF00257 (DHN) and PF04927 (SMP). Tbtools[38] was used to perform HMM search on the hybrid sweetgum genome database (our laboratory has successfully completed the whole genome sequencing of hybrid sweetgum, and we are eagerly anticipating the imminent publication of this invaluable data) to obtain all putative LEA genes of hybrid sweetgum. All identified candidate genes were analyzed by the Pfam database (http://pfam.xfam.org/ )[2] and the NCBI conserved domain search database (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )[39] to verify and ascertain the presence of conserved domains within the LEA genes. The LsfLEA genes that lacked a conserved domain and complete CDS sequence were removed. Additionally, any redundant LsfLEA genes were also deemed superfluous and therefore, justifiably excluded.To thoroughly investigate the complex and multifaceted properties of the LsfLEA genes, a comprehensive array of analytical techniques were employed. To begin with, the molecular weight (MW), theoretical pI, instability index, aliphatic index and grand average of hydropathicity (GRAVY) were scrupulously predicted utilizing the powerful and sophisticated ProtParam tool (
http://web.expasy.org/ )[40]. In addition, with an eye towards elucidating the subcellular localization of the LsfLEA proteins, we leveraged the remarkable predictive capabilities of the PSORT Prediction tool (www.genscript.com/wolf-psort.html )[41]. We conducted a rigorous and exhaustive analysis of their cis-acting elements, leveraging the PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html ) online website[42]. Moreover, protein hydrophobicity analysis utilizing ExPASy ProtScale (https://web.expasy.org/protscale ), we conducted a detailed analysis of their signal peptides, leveraging the powerful and intuitive online website Signa1P Server (https://services.healthtech.dtu.dk/service.php?SignalP-5.0 ). Phylogenetic analysis of 79 LsfLEA and 51 AtLEA[4] amino acid sequences was performed using the 1,000 bootstrapped Maximum Likelihood(ML) method[43] of MEGA 7.0 software. Chromosome localization and visualization were done using TBtools[38] software.Structural and protein conserved sequence analysis of LsfLEA genes
-
Structural analysis of the identified LsfLEA genes in the hybrid sweetgum genome was performed using Tbtools[38] software. The conserved structural domains of the LsfLEA family genes were analyzed using the MEME (Multiple Expectation Maximization for motif Elicitation) Suite (
http://meme-suite.org/ )[44]. Different subfamilies of LEA genes were labeled with different colors. Fourty conserved structural domains (Motif 1-Motif 40) were identified in the LsfLEA subfamily genes and labeled with different colors and numbers.Chromosome location and gene duplication of the LsfLEA genes
-
DNA sequences from the LEA family genes were mapped onto the genome of hybrid sweetgum. Distribution of the genes on the chromosomes or scaffolds was calculated by the TBtools[38] software. Duplicate events of the PtrLEA genes were calculated and visualized for mapping by using the TBtools[38] software.
Analysis of cis-acting elements of LsfLEA genes
-
To explore cis-elements in the gene promoter of LsfLEA, a 2 kb genomic sequence upstream of the transcription start site (TSS) of each gene was analyzed, the PlantCare database was extracted and searched to identify and count cis-elements associated with abiotic stress responses.
Heatmap of the differentially expressed 79 LsfLEA genes during hybrid sweetgum SE and different tissues
-
Combined with transcriptome data[36] and whole genome data, TBtools[38] software was used to draw the heatmap of gene expression in different periods and different tissues of hybrid sweetgum. Heatmap indicate the gene expression level by Log2[FPKM] with a color scale, each row represents a single gene, the IDs are indicated to the outside of fan, and each circle represents a sample.
Visualization of LEA key genes in different stages of somatic embryogenesis by qRT-PCR
-
These six genes were selected from the hybrid sweetgum LEA genes, specifically LsfLEA1-1 (EVM0027957), LsfLEA3-3 (EVM0021267), LsfLEA6-2 (EVM0005932), LsfDHN-2 (EVM0003818), LsfLEA1-3 (EVM0000925), LsfSMP-1 (EVM0023588). To validate the efficacy of these genes, we employed qRT-PCR gene expression analysis, and compared the gene expression levels with gene FPKM values obtained from transcriptomic data[36]. Using the plant RNA extraction kit, we extracted total RNA in accordance with the kit's instructions. In order to assess the concentration and quality of RNA samples, we utilized a NanoDrop 2,000 spectrophotometer (Thermo Scientific, Wilmington, DE, USA) and 1% agarose gel electrophoresis. The extracted total RNA was then subjected to reverse transcription using the TransScript® First Strand Synthesis Kit. For the purpose of qRT-PCR analysis, we designed specific primers utilizing the Primer Premier 5.0. The detailed primer sequences can be found in Table 2. The qRT-PCR analysis was conducted with the QuantStudio 6 Flex real-time PCR system. Each PCR reaction mixture consisted of 10 μL 2×PerfectStart ®Green qPCR SuperMix(Yisheng Biotechnology Shanghai Co., Ltd.), 1 μL cDNA, 0.4 μL forward primer, 0.4 μL reverse primer, and 8.2 μL ddH2O to achieve a final volume of 20 μL. The amplification conditions for qRT-PCR were as follows: 45 cycles of 94 °C for 30 s, 94 °C for 5 s, and 60 °C for 30 s. Apple EF1-α gene[45] was utilized as an internal reference control. The relative gene expression was calculated using the 2−ΔΔCᴛ method, with three biological replicates and three technical replicates being carried out.
Table 2. Primer sequence information of qRT-PCR.
Gene_ name Gene_id Primer sequence LsfLEA1-1 EVM0027957 F: TATTGGGGTGATGACGGTCC
R: TTCCTCTGCGTGGCCATATCLsfLEA3-3 EVM0021267 F: ATACTCGGCGGCATCACAAG
R: ACGTCAATCTCTTCCGCACGLsfLEA6-2 EVM0005932 F: TGGAGGACTACAAGCGTCAAG
R: CACCACCGGAAAGAGTGGGLsfDHN-2 EVM0003818 F: CCAATTGGGTTGGAAACCGTC
R: TCACCGAGCTAGAGCTTGAACLsfLEA1-3 EVM0000925 F: AGGAGAAGTTGAGCGACATGG
R: TTTGCTTGAGCTTCCTTCGCLsfSMP-1 EVM0023588 F: ATCGTGATCTGACCGGCATC
R: CATCAACCACCACCACATGCEF1-r-07041 / F: ACTGCACGGTCATTGATGCT
R: AAAGCATGCTCACGGGTCTGPlasmid construction and subcellular localization analysis
-
Based on the LsfLEA1-3 CDS, cloning primers LsfLEA1-3-F: ATGCATTCTGCAAAGGAGAAGT; LsfLEA1-3-R: CTACATATACTCACGTCGAGGAGG were designed by Primer 5. The cDNA from embryogenic callus of hybrid sweetgum was used as a template to amplify the LsfLEA1-3 CDS. The 50 µL PCR system included: 2 ×Phanta®Max Master Mix 25 µL, cDNA 2 µL, each primer 2.5 µL, ddH2O 18 µL, and the PCR program as follows: pre-denaturation at 95 °C for 3 min; denaturation at 95 °C for 15 s, annealing at 56 °C for 15 s, extension at 72 °C for 1 min, 34 cycles; extension at 72 °C for 5 min. Then that product was ligated into the pEasy-Blunt3 vector (TransGen Biotech) and sequenced. They were transformed into E. coli DH5α, cultured overnight for 12 h, and the monoclonal clones were selected for PCR identification, and the bacterial solution with the correct band was sequenced by using 1300-F:GACGCACAATCCCACTATCC;GFPfusion-R: CTCCACTGACAGAAAATTTG as primers. Primer sequences at both ends respectively introduced restriction enzymes Kpn1 and the corresponding protection bases, primer sequences for LsfLEA1-3-1300-F: 5'-CGGGGGACGAGCTCGGTACCATGCATTCTGCAAAGGAGAAGTTGA-3' and LsfLEA1-3-1300-R: 5'-CTAGAGGATCCCCGGGTACCCATATACTCACGTCGAGGAGGGTG-3', and the carrier of linearization, adopt the method of homologous recombination connect it. The LsfLEA1-3 was amplified and ligated into an expression pCAMBIA1300 vector with a GFP fluorescent label and a CaMV35 S promoter. Then, the successfully constructed plasmid was transferred to Agrobacterium GV3101 through the conventional freezing-thawing method. Next, empty pCAMBIA1300-GFP or pCAMBIA1300- LsfLEA1-3-GFP GV3101 instantly infiltrated the strain into 5-week-old N. benthamiana leaves with expression buffer (10 mM MES pH 5.6, 10 mM MgCl2, 200 µM acetosyringone). After the infiltrated N. benthamiana was cultured in the dark for 12 h and then in low light for 24 h, the fluorescence signals were captured with a confocal laser‐scanning microscope (TCS SP8; Leica).
-
Based on the HMM search of LEA1 (PF03760), LEA2 (PF03168), LEA3 (PF03242), LEA4 (PF02987), LEA5 (PF00477), LEA6 (PF10714), SMP (PF04927) and DHN (PF00257), a total of 79 LEA genes were identified from the whole genome of hybrid sweetgum, which were meticulously documented in Table 3. Upon close analysis, it became evident that the LsfLEA genes were divided into 8 subfamilies, each unique in their conserved domain, as displayed in Fig. 1. The LEA2 subfamily genes were notably the most significant, consisting of an astonishing 57 members, while the numbers of LEA1, LEA3, LEA4, LEA5, LEA6, DHN, and SMP subfamilies were more modest, with 5, 3, 2, 3, 2, 6, and 1 member, respectively. Upon further investigation, it was discovered that the predicted 79 LsfLEA genes encoded peptides with an extensive range of amino acid lengths, spanning from a diminutive 89 (LsfLEA6-1, LsfLEA6-2) to an impressive 323 (LsfLEA4-2) amino acids. Additionally, the molecular weight ranged from a minute 9.37028 (LsfLEA6-2) to 36.9282 (LsfLEA4-2) kDa. Further analysis of the calculated grand average of hydropathicity indexes revealed a total mean ranging from a minuscule −1.533 to a noteworthy 0.407. Furthermore, the hydrophilicity values of 65 LsfLEA genes (82%) were lower than 0, indicating the hydrophilic nature of most LsfLEA genes. The prediction of subcellular localization indicated a diverse and intricate distribution of LsfLEA genes, with LsfLEA1 genes found in both the nucleus and cytoplasm. Most LsfLEA2 genes were placed in chloroplasts, nucleus, cell walls, and membranes, while all LsfLEA3 genes were presented in chloroplasts. All LsfLEA4, LsfLEA5, and LsfLEA6 genes were found in the nucleus, while LsfDHN genes were contained in both the cytoplasm and nucleus. These intricate and diverse details are documented in Table 3.
Table 3. The characteristics of LsfLEA genes.
Name Gene ID Family Pfam ID Length kD pI GRAVY Subcellular localization LsfLEA1-1 EVM0027957 LEA1 PF03760 191 19.36702 6.8 −0.865 Cytoplasm. Nucleus. LsfLEA1-2 EVM0018353 LEA1 PF03760 120 13.7979 9.25 −0.863 Nucleus. LsfLEA1-3 EVM0000925 LEA1 PF03760 169 18.18858 8.77 −0.731 Nucleus. LsfLEA1-4 EVM0022193 LEA1 PF03760 164 16.51795 8.83 −0.946 Cytoplasm. Nucleus. LsfLEA1-5 EVM0016558 LEA1 PF03760 107 12.32725 9.33 −0.769 Nucleus. LsfLEA2-1 EVM0002770 LEA2 PF03168 239 26.42419 10 −0.094 Cell membrane. Cell wall. LsfLEA2-2 EVM0027034 LEA2 PF03168 205 22.97259 9.84 −0.017 Chloroplast. LsfLEA2-3 EVM0005964 LEA2 PF03168 177 19.27253 10.27 0.042 Chloroplast. LsfLEA2-4 EVM0015809 LEA2 PF03168 190 21.8101 10.03 −0.482 Chloroplast. LsfLEA2-5 EVM0019694 LEA2 PF03168 214 23.44119 9.91 −0.121 Cell membrane. Nucleus. LsfLEA2-6 EVM0004956 LEA2 PF03168 210 23.12978 9.97 −0.186 Chloroplast. LsfLEA2-7 EVM0027836 LEA2 PF03168 219 24.18402 9.73 −0.195 Chloroplast. LsfLEA2-8 EVM0001803 LEA2 PF03168 237 27.18655 9.15 −0.103 Chloroplast. LsfLEA2-9 EVM0020637 LEA2 PF03168 178 19.33671 10.25 0.086 Chloroplast. LsfLEA2-10 EVM0015055 LEA2 PF03168 184 19.86985 9.92 −0.223 Chloroplast. Nucleus. LsfLEA2-11 EVM0017871 LEA2 PF03168 212 23.53223 10 −0.157 Chloroplast. Nucleus. LsfLEA2-12 EVM0018801 LEA2 PF03168 260 28.41312 9.01 0.133 Chloroplast. LsfLEA2-13 EVM0013310 LEA2 PF03168 198 21.34478 9.37 0.407 Cell membrane. LsfLEA2-14 EVM0010683 LEA2 PF03168 215 23.44427 9.88 −0.083 Cell membrane. Nucleus. LsfLEA2-15 EVM0023593 LEA2 PF03168 201 22.72988 10.05 0.109 Chloroplast. Nucleus. LsfLEA2-16 EVM0028234 LEA2 PF03168 188 20.34671 10.27 −0.001 Chloroplast. LsfLEA2-17 EVM0003550 LEA2 PF03168 151 16.49626 5.58 0.077 Chloroplast. Nucleus LsfLEA2-18 EVM0008005 LEA2 PF03168 265 29.16785 9.35 −0.066 Chloroplast. LsfLEA2-19 EVM0007435 LEA2 PF03168 267 29.08778 8.38 −0.215 Cell membrane. Cell wall. Nucleus. LsfLEA2-20 EVM0028118 LEA2 PF03168 223 25.25915 10.05 −0.122 Chloroplast. Mitochondrion. LsfLEA2-21 EVM0001898 LEA2 PF03168 215 23.68493 10.14 −0.021 Nucleus. LsfLEA2-22 EVM0018273 LEA2 PF03168 232 24.94073 9.3 0.113 Cell membrane. Chloroplast. LsfLEA2-23 EVM0025800 LEA2 PF03168 189 20.97945 10.01 −0.074 Chloroplast. Nucleus. Peroxisome. LsfLEA2-24 EVM0013583 LEA2 PF03168 264 29.43273 8.93 −0.048 Cell wall. Nucleus. LsfLEA2-25 EVM0020355 LEA2 PF03168 306 34.40006 9.47 −0.383 Cell membrane. Chloroplast. LsfLEA2-26 EVM0011840 LEA2 PF03168 214 23.38122 10.14 −0.056 Chloroplast. LsfLEA2-27 EVM0023465 LEA2 PF03168 215 23.46828 9.91 −0.064 Cell membrane. Nucleus. LsfLEA2-28 EVM0021483 LEA2 PF03168 262 29.00554 6.51 −0.288 Cell wall. LsfLEA2-29 EVM0024390 LEA2 PF03168 213 23.36631 9.94 −0.035 Nucleus. LsfLEA2-30 EVM0024154 LEA2 PF03168 215 23.32817 9.88 −0.031 Chloroplast. LsfLEA2-31 EVM0007128 LEA2 PF03168 244 27.2663 10.2 −0.063 Chloroplast. LsfLEA2-32 EVM0007725 LEA2 PF03168 259 28.26897 9.17 0.09 Chloroplast. LsfLEA2-33 EVM0019270 LEA2 PF03168 223 24.39271 9.61 0.041 Cell wall. Chloroplast. LsfLEA2-34 EVM0011791 LEA2 PF03168 210 23.90993 9.78 0.129 Cell membrane. Chloroplast. LsfLEA2-35 EVM0015677 LEA2 PF03168 191 20.8606 9.28 0.33 Chloroplast. LsfLEA2-36 EVM0011399 LEA2 PF03168 209 22.83086 9.98 0.197 Chloroplast. LsfLEA2-37 EVM0020583 LEA2 PF03168 151 16.45712 5.04 0.054 Cell membrane. Nucleus. LsfLEA2-38 EVM0011512 LEA2 PF03168 213 23.27806 9.97 −0.17 Chloroplast. Nucleus LsfLEA2-39 EVM0000290 LEA2 PF03168 251 28.02687 9.56 −0.076 Chloroplast. LsfLEA2-40 EVM0006920 LEA2 PF03168 269 29.83144 10.03 −0.246 Nucleus. LsfLEA2-41 EVM0027367 LEA2 PF03168 253 27.51712 10.3 −0.089 Cell membrane. Cell wall. LsfLEA2-42 EVM0005315 LEA2 PF03168 311 34.5181 9.07 −0.186 Cell wall. Nucleus. LsfLEA2-43 EVM0025389 LEA2 PF03168 178 19.27267 10.25 0.108 Chloroplast. LsfLEA2-44 EVM0000222 LEA2 PF03168 186 21.21822 7.87 −0.163 Cell membrane. Chloroplast. LsfLEA2-45 EVM0018178 LEA2 PF03168 212 23.1178 9.91 −0.134 Cell membrane. Cell wall. LsfLEA2-46 EVM0005757 LEA2 PF03168 178 19.26954 10.06 0.087 Chloroplast. LsfLEA2-47 EVM0016553 LEA2 PF03168 209 22.80349 10.14 −0.098 Chloroplast. LsfLEA2-48 EVM0016432 LEA2 PF03168 210 23.00723 9.83 0.226 Mitochondrion. Nucleus. LsfLEA2-49 EVM0021824 LEA2 PF03168 308 34.60045 9.64 −0.337 Cell membrane. Chloroplast. LsfLEA2-50 EVM0023803 LEA2 PF03168 214 23.29603 9.88 −0.102 Cell membrane. Cell wall. LsfLEA2-51 EVM0017004 LEA2 PF03168 197 21.68226 10.22 −0.131 Chloroplast. LsfLEA2-52 EVM0012352 LEA2 PF03168 217 24.03337 10.21 0.105 Chloroplast. LsfLEA2-53 EVM0023248 LEA2 PF03168 225 24.9939 9.55 −0.009 Cell membrane. Chloroplast. LsfLEA2-54 EVM0006284 LEA2 PF03168 213 23.40402 9.56 −0.175 Chloroplast. Cytoplasm. Nucleus. LsfLEA2-55 EVM0008344 LEA2 PF03168 315 34.64837 4.75 −0.403 Nucleus. LsfLEA2-56 EVM0022790 LEA2 PF03168 253 28.07601 10.17 −0.1 Cell membrane. Cell wall. LsfLEA2-57 EVM0012837 LEA2 PF03168 191 21.15586 9.58 0.26 Chloroplast. LsfLEA3-1 EVM0028438 LEA3 PF03242 101 10.79223 9.88 −0.222 Chloroplast. Nucleus. LsfLEA3-2 EVM0019119 LEA3 PF03242 94 10.47005 9.37 −0.418 Chloroplast. LsfLEA3-3 EVM0021267 LEA3 PF03242 121 13.48961 10.28 −0.668 Chloroplast. LsfLEA4-1 EVM0019164 LEA4 PF02987 212 23.759 5.42 −1.533 Nucleus. LsfLEA4-2 EVM0021387 LEA4 PF02987 323 36.9282 9.53 −1.53 Nucleus. LsfLEA5-1 EVM0020631 LEA5 PF00477 94 9.93673 5.48 −1.26 Nucleus. LsfLEA5-2 EVM0022920 LEA5 PF00477 114 12.14812 5.53 −1.278 Nucleus. LsfLEA5-3 EVM0002005 LEA5 PF00477 114 12.07605 5.91 −1.251 Nucleus. LsfLEA6-1 EVM0004641 LEA6 PF10714 89 9.40021 4.75 −0.939 Nucleus. LsfLEA6-2 EVM0005932 LEA6 PF10714 89 9.37028 5.77 −1 Nucleus. LsfSMP-1 EVM0023588 SMP PF04927 259 26.66671 4.66 −0.253 Nucleus. LsfDHN-1 EVM0028566 Dehydrin PF00257 290 32.91761 5.23 −1.388 Nucleus. LsfDHN-2 EVM0003818 Dehydrin PF00257 174 18.52736 8.01 −1.126 Cytoplasm. LsfDHN-3 EVM0011412 Dehydrin PF00257 174 18.62131 9.45 −1.31 Cytoplasm. LsfDHN-4 EVM0018312 Dehydrin PF00257 193 21.77947 5.56 −1.388 Nucleus. LsfDHN-5 EVM0008852 Dehydrin PF00257 1891 21.36301 5.46 −1.348 Nucleus. LsfDHN-6 EVM0003289 Dehydrin PF00257 155 16.54385 9.1 −1.369 Cytoplasm. 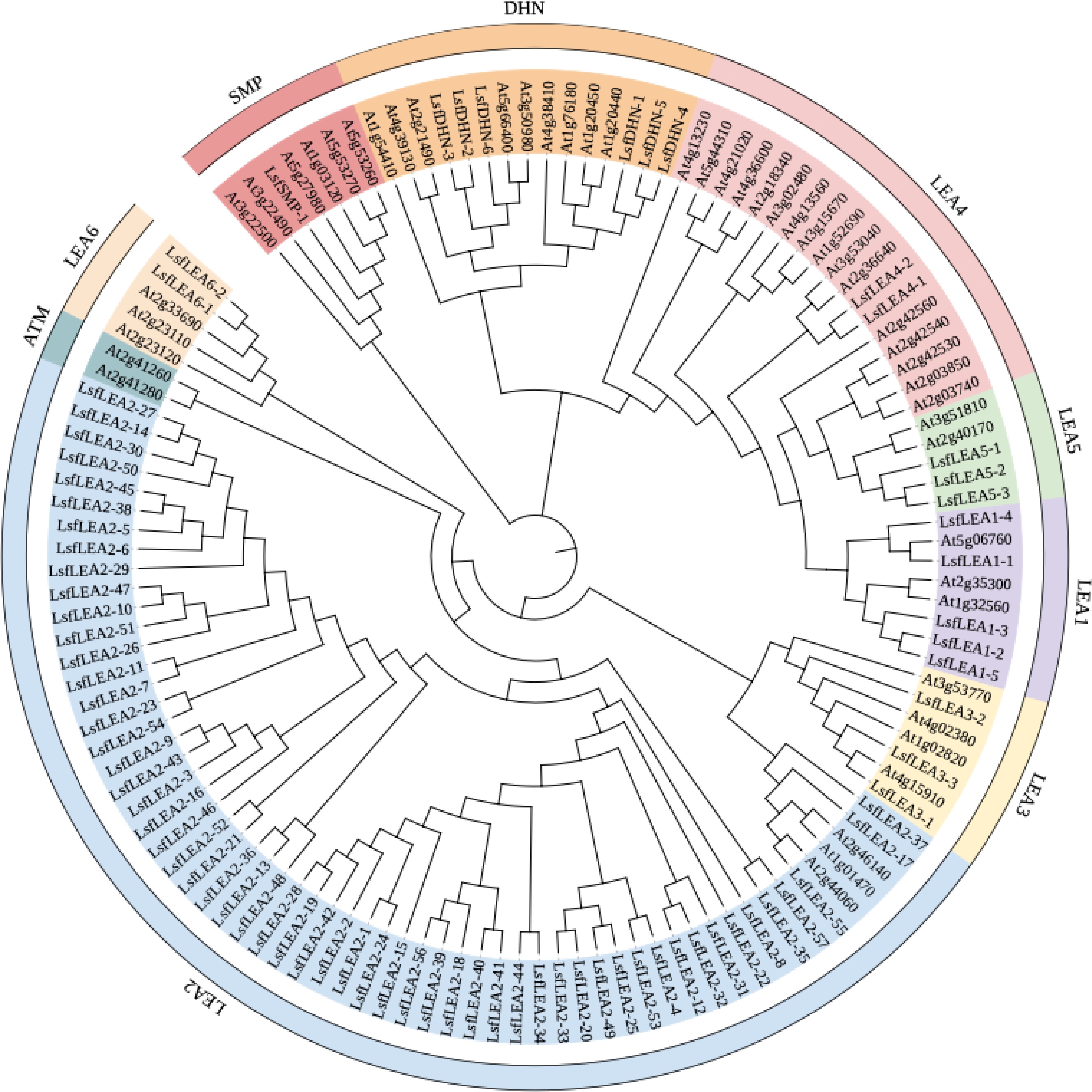
Figure 1.
Maximum likelihood phylogenetic tree of 79 LsfLEA genes and 51 AtLEA genes. The phylogenetic tree was constructed using MEGA 7 software with 1000 bootstrap replicates. We used the online site iTOL for evolutionary tree beautification, and the 9 major groups are marked with different colored backgrounds.
Analysis of LEA gene structure and protein conserved sequence of hybrid sweetgum
-
The number of introns in the LsfLEA gene is small, with the majority exhibiting a mere one or two introns. Upon scrutinizing conserved motifs, it was uncovered that the vast majority of homologous members within the same group shared indistinguishable conserved motifs (Fig. 2a). When undertaking an exhaustive analysis of gene structure, it was noticed that there existed a degree of differentiation in the gene structure of LsfLEA genes, with the exception of those in the same subfamily that possessed virtually identical structures. An exemplar of this was seen with LsfLEA2-9, LsfLEA2-45, LsfLEA2-54, LsfLEA2-26, LsfLEA2-5 and LsfLEA2-51, which all exhibited similar gene structures (Fig. 2b). The members of the same group are presumed to have relatively similar biological functions. These results suggest that the composition of conserved structural domain motifs differs among different LsfLEA subfamilies, but the composition of conserved structural domain motifs of LsfLEA genes is extremely similar within the same subfamily, and the motifs of these structural domains are also conserved. Motif sequences and amino acid sequences of 79 LEA genes in hybrid sweetgum are given in Table 4 & Supplemental Table S1.
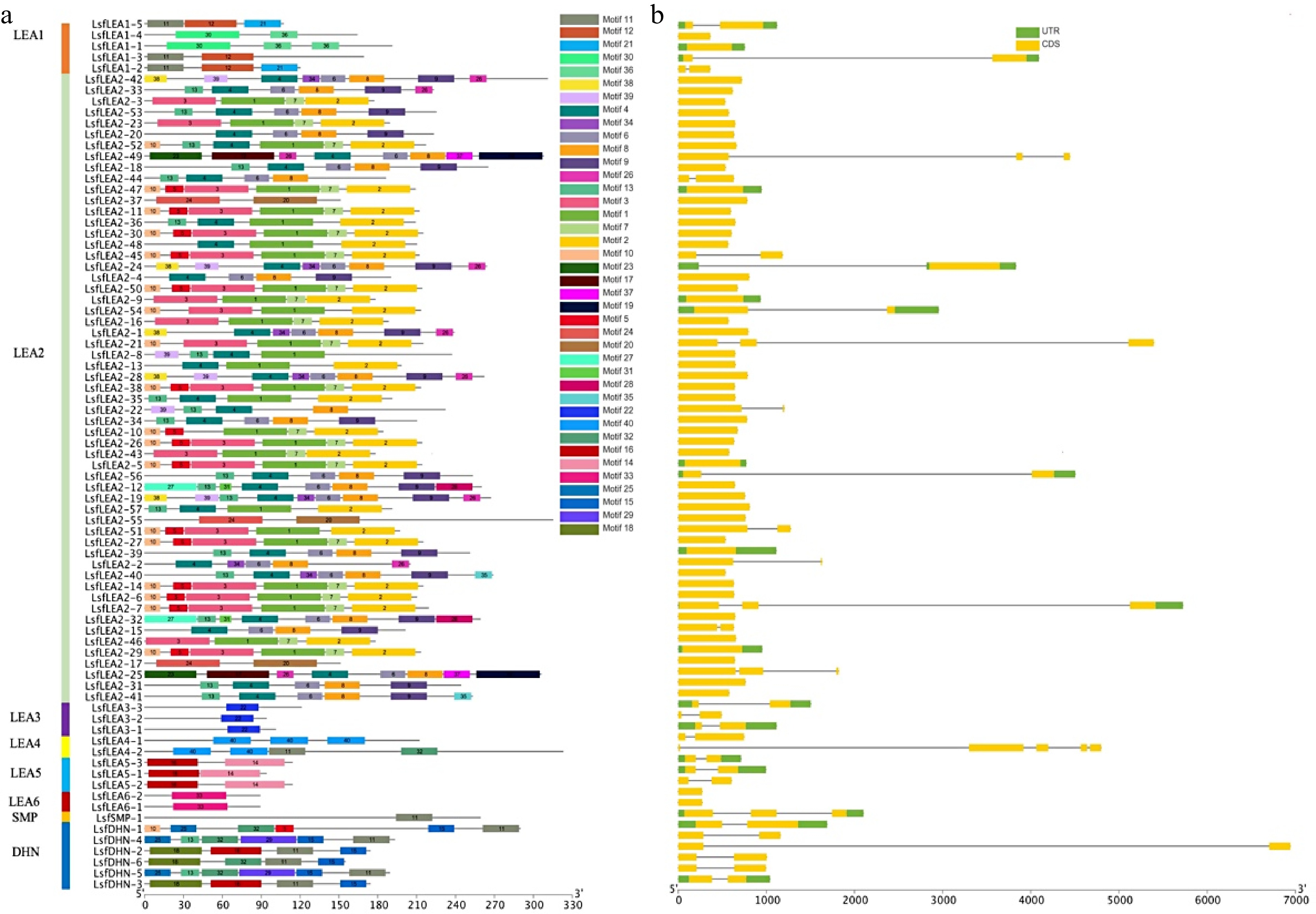
Figure 2.
(a) Motif analysis of LsfLEA genes from hybrid sweetgum. The conserved motifs of each group on the right side were identified by the MEME web server. Different motifs are represented by different colored boxes (b) Gene structure distribution map of LsfLEA.
Table 4. The motif sequences of LsfLEA genes.
Motif number Motif sequences 1 NTRFVAQVTVKNTNFGPYKFDNTTATFTYGGMTVGZVVIPKGKAGARSTK 2 SDISSGILKLSSQAKLSGKVELMFVMKKKKSAEMNCTLTFBLSAKALQAL 3 RRIKIAAYIVAFVVFQSIIIAIFAVTVMKYRTPKFRLGKVTVETLTSTPA 4 VVALJLWLVFRPKRPKFTVNSVSVSSFNL 5 DEESATLQSKEFRRK 6 RNPNKKJSIYYDSIEASVYY 7 KIBVTVDLNSDALTS 8 QQILSSGSLPPFYQGRKNTTVLSTTLAG 9 FBVKVDARVRFKVGRWKTKRVGVRVSCDG 10 MAEKNQQVYPAAP 11 SAAEKKKGMKEKIKEKLPGYKAKAEEEKE 12 HEGRKAKEAQAKMEMHEAKARHAEEKLNAKQSHLYGHQVHE 13 PKRERRRGCCCCGCL 14 KGGQTRKEQLGTEGYQEMGRKGGLSTTDQSGGERAAELGVDIDESKF 15 HHPEEKKGILEKIKEKLPGQH 16 GNREQREELEARARQGETVVPGGTGGGSLEAQEHLAEGRS 17 PHHYHCSPIHHSRESSTSRFSASLKNAHHHKHSAPWKKMHRVVDVDDDD 18 RDEYGNPIQLTDERGNPVQLTDEYGNPMHLSGVATTHGTTA 19 LNLTFVMRSRAYILGKLVKSKFYKRIRCSVTLRGNQLGKLQNLTDSCIYQ 20 NDITMLDVGLKVPHNVLVTLVKDIGADWDIDYELEVGLTIDLPIIGNFTI 21 HIIGTMNQWWVSMDTMGRSLLELRPLYLY 22 WVPDPVTGYYRPENRAGEIDVAELRD 23 MHAKSDSEVTSLDASSPPRSPRRPLYYVQSPSQHDVEKMSY 24 NFMVEKVANIKKPEASVEDVDLKDVSRECIDYNAKVAVKNPYGHALPICE 25 MADQHFQCHEKEPRDRGMFDF 26 GTMVGGPRECKVRL 27 MADSAIRKTEDSPPSSKPSPNPTSKPVRHVVFSEIPCRPHK 28 IQQSQVDIGQEPKCSVKMFSFRLHTFLFI 29 EGGEEKKKKKGLKEKIKEKMACQGEEEVTEIPVDKCDNIVDAET 30 GMDKTKATMQZKVEKMTARDPIZKEMATZKKEAKKTEAELNKQEAREHNA 31 CCAWGCMIGF 32 EPQEKKEVEKPTLVEELRRSGSSSSSSSD 33 GLPLESSPYVKNSDLEDYKRKGYGTEGHLDPKPGRGAGGTDAP 34 SSQITGNWNVSFYV 35 CKVDLRIKIWKWTF 36 GHPTGGHQMSAMPGEGTWQPTW 37 YQVPLYGGVSVLGGARNHYEN 38 MEDQKKPVIGYPVQFYHP 39 NTAAAHVVQPPPVVHHQQQ 40 GKVGEYKDYAAEKAKETKDSALEKAREYKD Chromosomal localization of the hybrid sweetgum LEA genes
-
The chromosomal locations of 79 hybrid sweetgum LEA genes were queried in the hybrid sweetgum genome database, and the chromosomal localization of the LsfLEA genes and gene duplication events were located by TBtools software. As the data poured in, we were able to pinpoint the precise chromosomal location of a staggering 75 genes, leaving only four genes (LsfLEA2-12, LsfLEA5-3, LsfLEA2-49, and LsfDHN-5) dispersed across unassembled scaffolds (Fig. 3a). The distribution of the LsfLEA genes across different chromosomes was marked by a noteworthy level of diversity and heterogeneity (Fig. 3b). It has been observed that the chromosomes LG12 and LG14 are equipped with the highest number of LsfLEA genes, 13 LsfLEA genes each. It is worthy of note that LG10 has 11 LsfLEA genes distributed across its length. In contrast, the chromosomes LG06 and LG16 have been found to be wanting when it comes to the number of LsfLEA genes present on them, boasting only one LsfLEA gene each. What's truly intriguing is that the LsfLEA2 subfamily genes have been identified on as many as seven chromosomes. These chromosomes are LG01, LG05, LG06, LG09, LG12, LG14, and LG16. On the other hand, the LsfLEA1 subfamily genes have been found to be distributed across only two chromosomes, LG03 and LG10. The LsfLEA3 subfamily genes are confined to only two chromosomes as well, LG07 and LG13. Moreover, it is fascinating to observe that the LsfLEA4 subfamily genes are present only on the LG10 chromosome. Further analysis has revealed that the LsfLEA5 subfamily genes have been distributed across the LG10 and LG11 chromosomes, while the LsfSMP family genes have been identified solely on the LG07 chromosome. The LsfDHN family genes, meanwhile, are distributed across the LG02, LG08, and LG15 chromosomes. In addition, we found that the highest number of subfamilies were located on chromosome LG10, including four subfamilies of LEA1, LEA2, LEA4 and LEA5 genes. The evaluation of collinearity within the hybrid sweetgum species revealed an interesting discovery, characterized by a grand total of nine gene duplication events. (Fig. 3c).
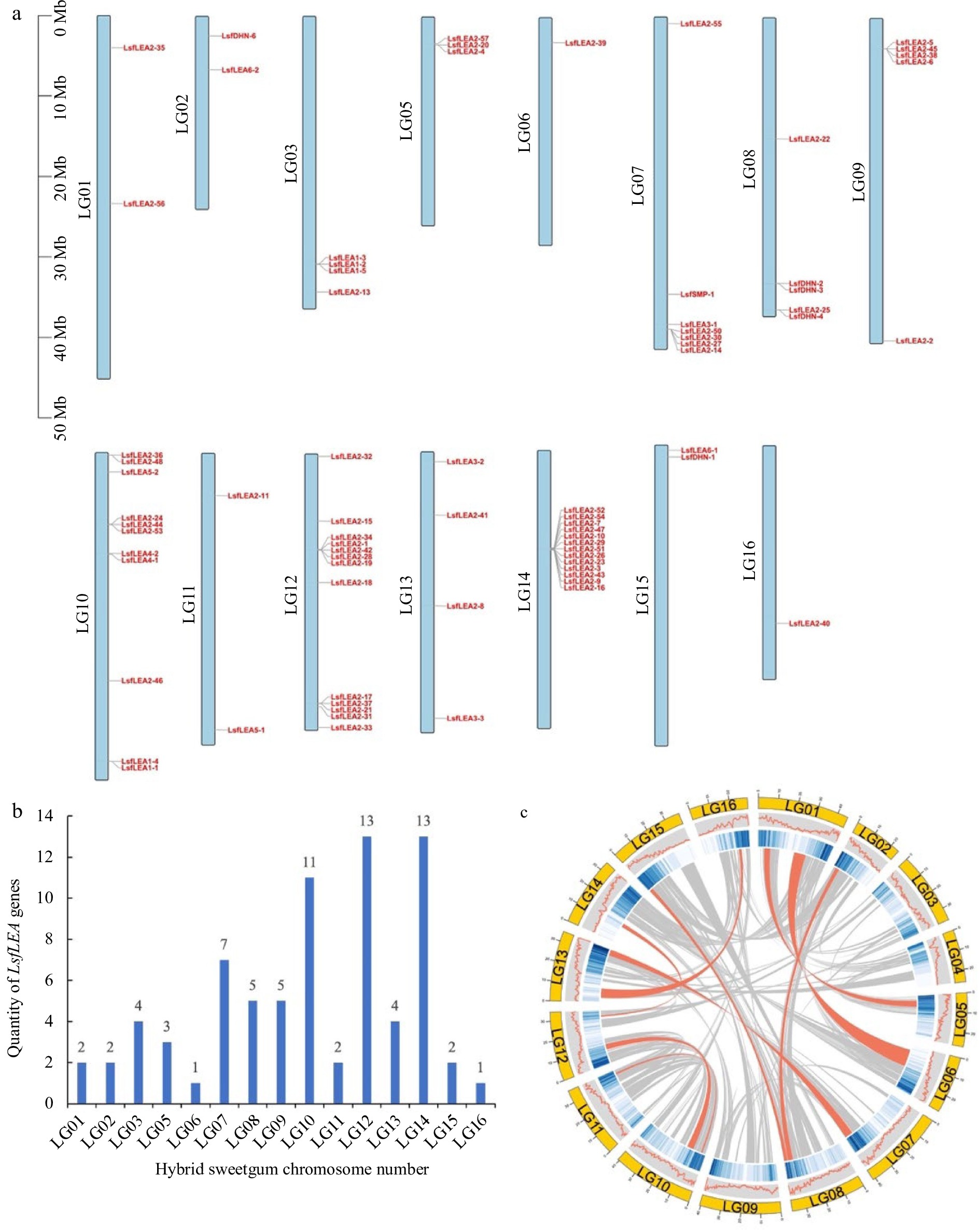
Figure 3.
Chromosomal localization of the LsfLEA genes and gene duplication events. (a) Chromosomal localization of hybrid sweetgum LsfLEA gene. (b) Numbers of LsfLEA genes on each chromosome in hybrid sweetgum. (c) Duplicate pairs of the LsfLEA genes in hybrid sweetgum. The red lines represent collinear pairs of the LsfLEA genes.
Analysis of cis-acting elements of LEA genes in hybrid sweetgum
-
The presence of multiple distinct cis-elements in gene promoters may indicate that these genes have different functions. We identify cis-elements that regulate phytohormone response in the promoters of LsfLEA genes (Fig. 4). These elements included abscitic acid (ABA)-(ABRES), auxin-(AuxreS), methyl jasmonate (Mejares), and salicylic acid response elements (SARES). Upon further investigation, we found that the promoters of some LsfLEA genes boasted ABREs and MeJAREs, which could be activated by ABA and MeJA, respectively. We also uncovered a plethora of cis-elements linked to the plant's response to stress, including drought response elements (DREs), light response elements, MYB binding sites (MBS), and MYC binding sites (MYC). Perhaps the most fascinating discovery of all was that the LsfLEA genes promoter was heavily enriched with light-responsive elements, accounting for a staggering 87% of all cis-elements identified. The nucleotide sequences of 79 LEA genes in hybrid sweetgum are given in Supplemental Table S2.
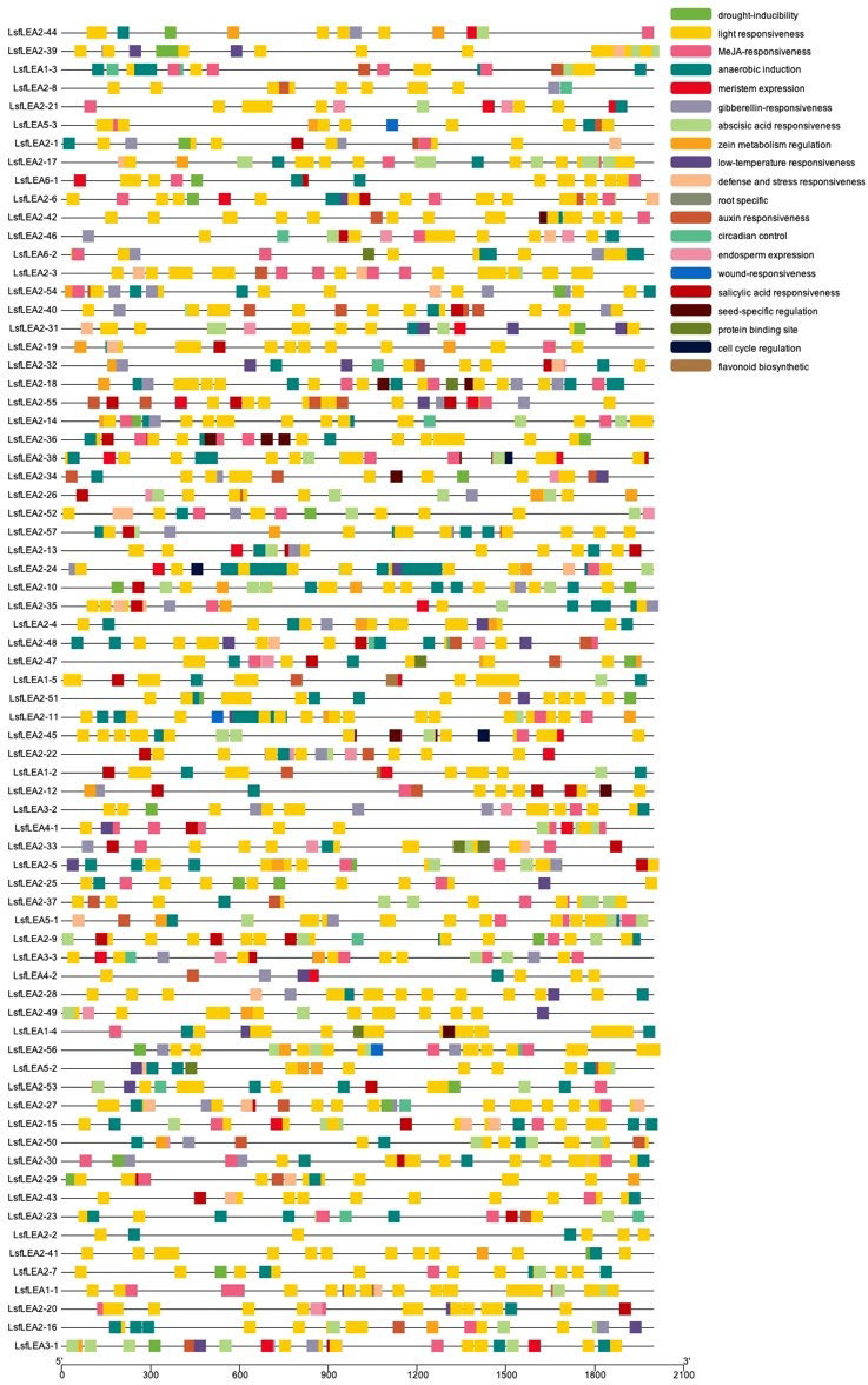
Figure 4.
Cis-acting elements analysis of LsfLEA genes. The patterns in different colors on the right represent different cis-elements.
Expression profile analysis of hybrid sweetgum LEA genes in different stages of somatic embryogenesis and different tissues
-
Non-embryonic callus, embryonic callus and embryos of different forms of hybrid sweetgum were taken as samples. After cotyledon embryos were formed into seedlings, root and leaf samples were taken as samples (Fig. 5). Gene expression levels were measured in a total of nine periods. To investigate the expression pattern of the LsfLEA gene in different tissues, RNA-Seq analysis of the LsfLEA genes were performed (Fig. 6). The findings are indicative of the fact that the LsfLEA1 genes are predominantly and primarily expressed in NEC, EC, GE, HE, TE, and CE stages, with conspicuously low or practically negligible expression in the root (R), stem (S), and leaf (L). It is worth noting that LsfLEA1-5, on the other hand, is not expressed in any of the tissues. In stark contrast to the tissue specificity of LsfLEA1-5, the expression of the LsfLEA2 genes were not delineated to any particular tissues, yet intriguingly, LsfLEA2-35 is not expressed in any of the tissues. Moreover, the LsfLEA3 genes were detected to be expressed in all stages, and the expression of LsfLEA3-1 in the root (R), stem (S), and leaf (L) was conspicuously and remarkably high. Conversely, the expression of LsfLEA3-3 was most pronounced in GE, HE, TE, and CE. Notably, the LsfLEA4 genes were demonstrated to be expressed minimally or not at all in any of the stages. The expression of LsfLEA5 genes in GE, HE, TE, and CE were manifestly and significantly higher than that observed in other stages. Finally, the expression level of the LsfDHN genes were determined to be higher in GE, HE, and TE, with the expression of LsfDHN-6 being markedly and notably higher than other LsfLEA genes in all tissues. Additionally, among the LsfSMP subfamily, the expression of LsfSMP-1 was highly and remarkably pronounced in GE, HE, and TE, while being very low or even completely absent in all other tissues.
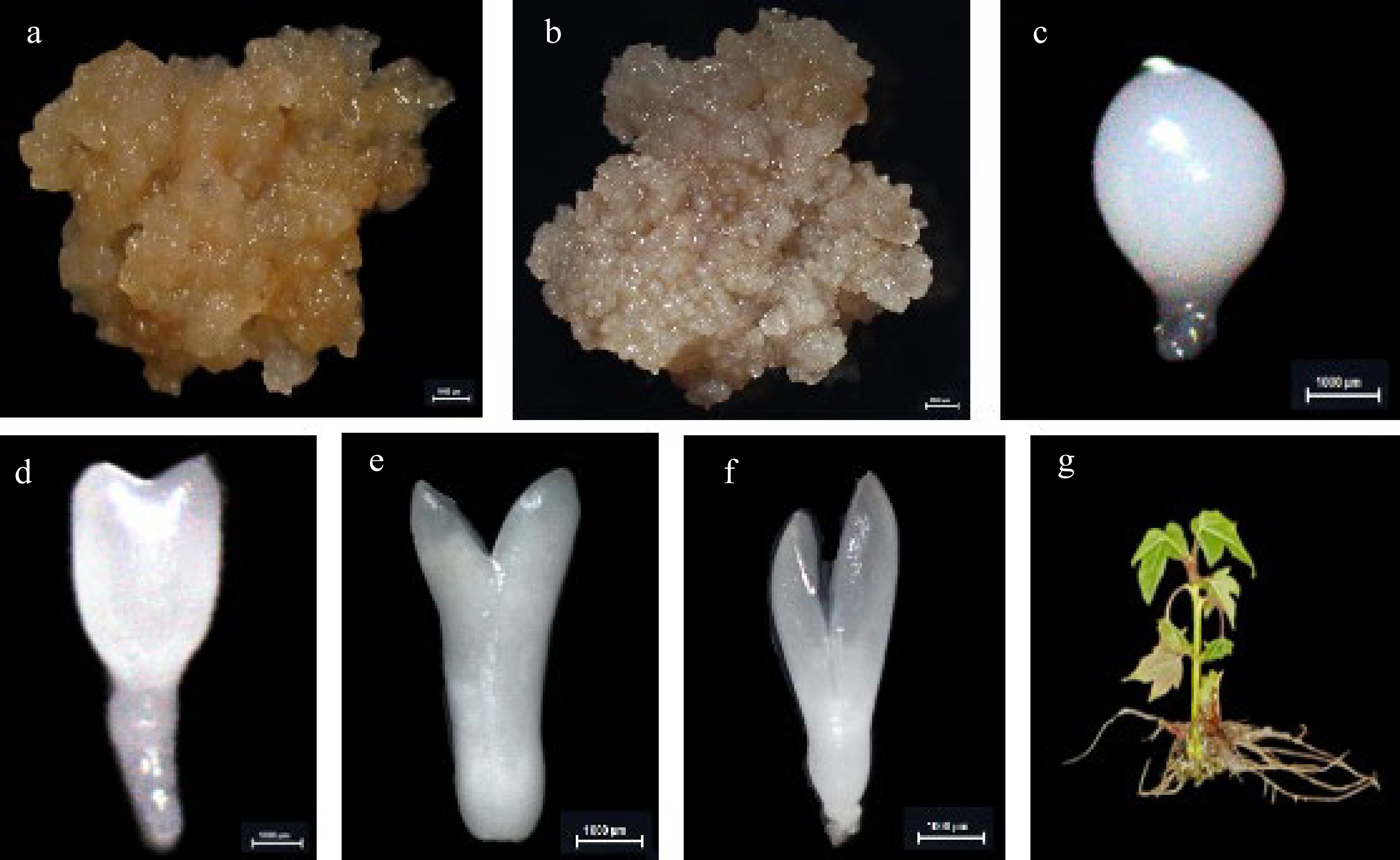
Figure 5.
The cultures during hybrid sweetgum somatic embryogenesis and different tissues. (a) NEC: non-embryogenic callus. (b) EC: friable-embryogenic callus. (c)-(f) Somatic embryos at different developmental stages globular embryo (GE), heart-shaped embryo (HE), torpedo-shaped embryo (TE), cotyledonal embryo (CE). (a)-(f) Bars = 1000 μm. (g) Different tissues in root (R), stem (S), and leaf (L) of hybrid sweetgum.
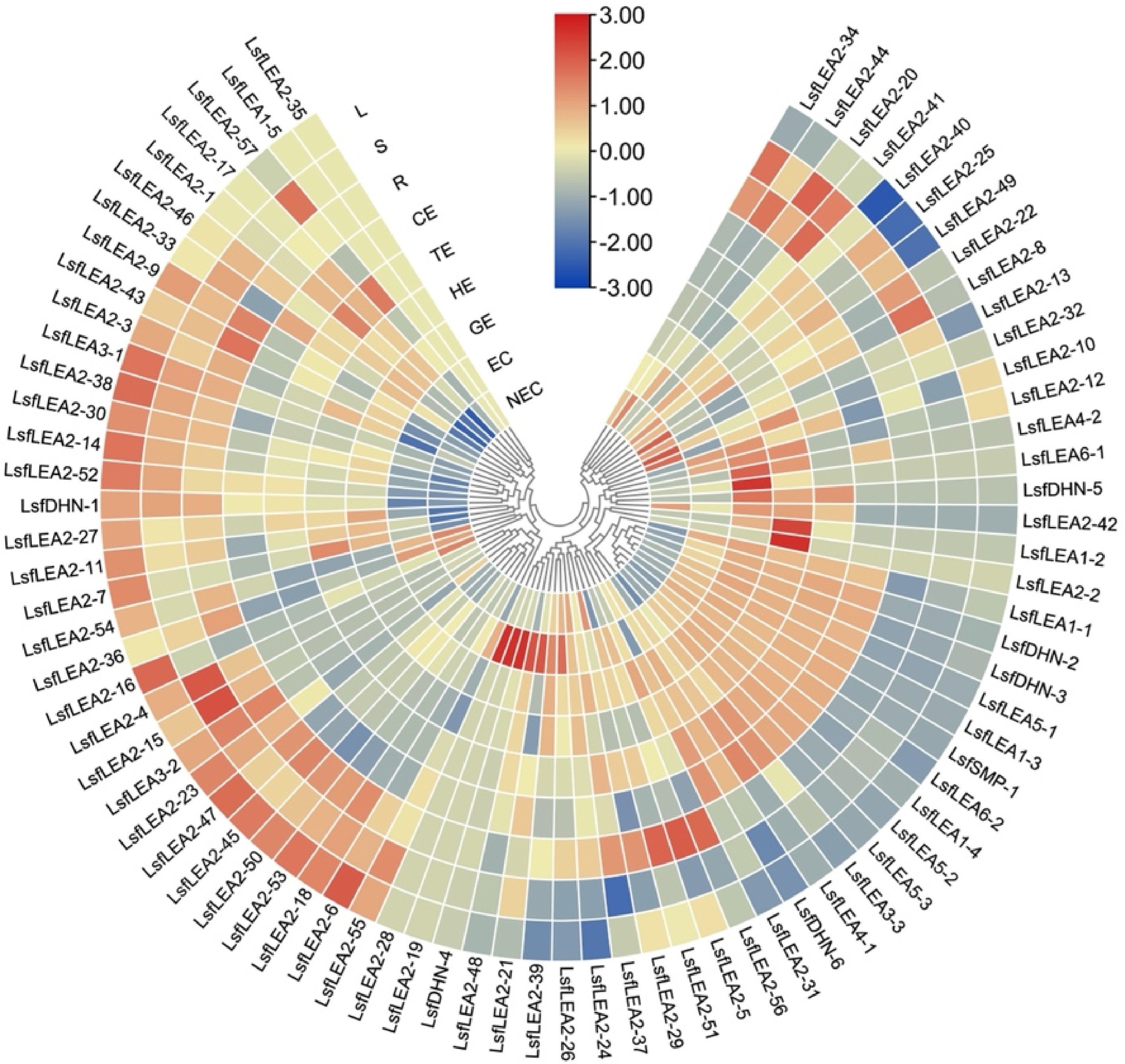
Figure 6.
Heatmap of the differentially expressed 79 LsfLEA genes during hybrid sweetgum SE and different tissues. The heatmap was clustered by TBtools software. The heatmap indicates the gene expression level by Log2[FPKM] with a color scale, each row represents a single gene, the IDs are indicated to the outside of fan, and each circle represents a sample. NEC: non-embryogenic callus; EC: friable-embryogenic callus; GE: globular embryo; HE: heart-shaped embryo; TE: torpedo-shaped embryo; CE: cotyledonal embryo; R: root; S: stem; L: leaf.
Validation of hybrid sweetgum LEA genes by qRT-PCR in different stages of somatic embryogenesis and different tissues
-
The results of the quantitative real-time polymerase chain reaction (qRT-PCR) analysis presented in Fig. 7 exhibit a remarkable variability in the expression levels of the hybrid sweetgum LEA family genes during somatic embryogenesis and different tissues. Intriguingly, the transcriptional levels of LsfLEA3-3, LsfLEA1-1, LsfLEA1-3, and LsfDHN-2 displayed negligible or scanty expression at the non-embryogenic callus (NEC) stage, but were conspicuously augmented during somatic embryo development, eventually culminating in the attainment of the highest expression level at this stage, which exhibited a precipitous decline in the nutritional organs (R, L, S). These results suggest that these genes may promote somatic embryo development and confer a pivotal function in the procurement and sustenance of somatic embryogenic competency. Moreover, LsfLEA6-2 exhibited persistent or accentuated up-regulation throughout somatic embryogenesis, whereas it was concomitantly down-regulated in NEC and nutritional organs (R, L, S), thereby underscoring the pivotal role played by these molecular markers in the context of somatic embryogenesis in hybrid sweetgum. In sharp contrast, the gene expression of LsfSMP-1 was observed to be comparatively lower during NEC and somatic embryo development (GE, HE, TE, and CE stages), but up-regulated in nutritional organs (R, L, S), pointing towards the active participation of this gene in the overall process of somatic embryogenesis and its subsequent development. Meanwhile, qRT-PCR verification of genes related to somatic embryogenesis also demonstrated the high correlation between RNA-seq and qRT-PCR data.
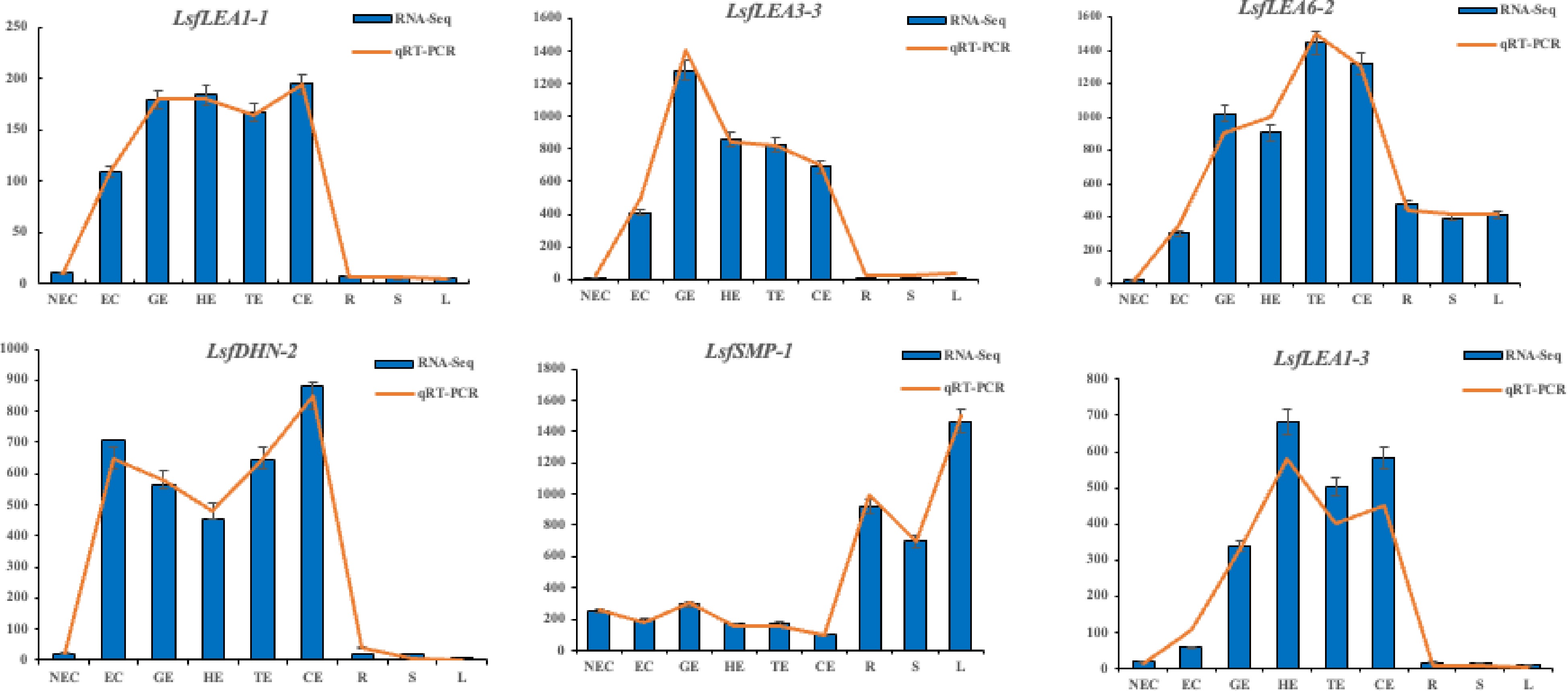
Figure 7.
Transcriptome expression (FPKM value) and qRT-PCR value of six LEA family genes in hybrid sweetgum at different stages during somatic embryogenesis. Relative expression values were calculated using the 2−ΔΔCᴛ method with apple EF1-α as a housekeeping gene. NEC: non-embryogenic callus; EC: friable-embryogenic callus; GE: globular embryo; HE: heart-shaped embryo; TE: torpedo-shaped embryo; CE: cotyledonal embryo; R: root; S: stem; L: leaf.
Subcellular localization of LsfLEA 1-3
protein -
To verify the prediction results of LsfLEA1-3 protein subcellular localization, to begin with, the successful cloning of the full-length CDS of the LsfLEA1-3 gene was accomplished, followed by the ligation of said gene into the T-vector for sequencing purposes. Following this step, the LsfLEA1-3 gene was inserted into the pCAMBIA1300 vector, ensuring that these genes were fused with the GFP protein driven by the 35 S promoter upon expression in N. benthamiana leaves. Subsequently, the subcellular localization of the expressed proteins was observed and analyzed in a rigorous manner. The localization results obtained were markedly striking, with the GFP protein devoid of any gene insertion being expressed in various organelles in N. benthamiana, whilst the GFP fused with LsfLEA1-3 protein was exclusively expressed within the nucleus. This phenomenon effectively proves the expression and functional operation of LsfLEA1-3 protein within the nucleus (Fig. 8).
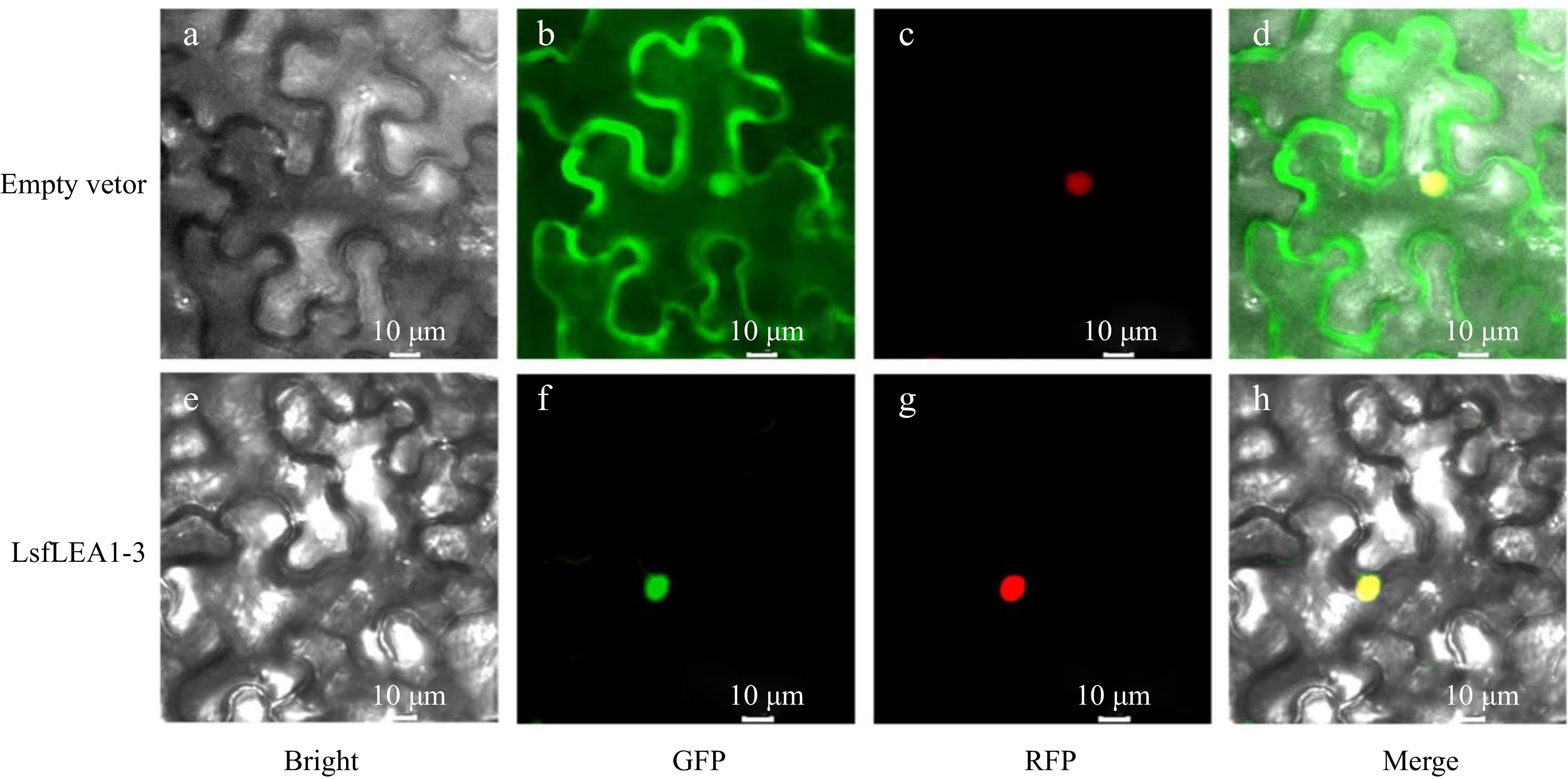
Figure 8.
Subcellular localization of LsfLEA 1-3 protein in N. benthamiana leaves. (a) GFP empty vector in bright field. (b) Empty vector in GFP green fluorescence. (c) Empty vector in RFP red fluorescence. (d) Empty vector in merge. (e) LsfLEA 1-3 protein in bright field. (f) LsfLEA 1-3 protein in GFP green fluorescence. (g) LsfLEA 1-3 protein in RFP red fluorescence. (h) LsfLEA 1-3 protein in merge.
-
The LEA family genes have been the subject of investigation across many different plant species, however, this gene family in the hybrid sweetgum whole genome has been sorely lacking. In this study, 79 LsfLEA family genes were identified and separated into eight unique subfamilies. Conserved structural domain analysis revealed that each member of the LEA subfamily contains its own unique set of conserved domains, which have been observed in the likes of Arabidopsis thaliana[4], Oryza sativa[6] and Triticum aestivum[46, 47], and these results suggest that LsfLEA proteins may have group-specific functions.
Furthermore, upon closer inspection, the conserved motifs observed in each LEA subfamily suggest that their members may have originated from gene amplifications within the subfamily[48]. Our results also indicate that the LsfLEA genes contain very few introns, which is consistent with other plant species such as Brassica napus[5], Camellia sinensis[49], and maize[50], the low number of introns facilitates the rapid expression of the LsfLEA genes and functional protein production under abiotic stress.
Subcellular localization results of the LsfLEA protein revealed its widespread presence across several subcellular compartments, namely the mitochondria, nucleus, cytoplasm, cell membrane, and chloroplast (Table 3). Interestingly, these observations are corroborated by previous reports on Arabidopsis thaliana[4] and Solanum lycopersicum[10], lending further support to the notion that LEA proteins from hybrid sweetgum are ubiquitously distributed across cells and their corresponding tissues, serving indispensable roles in the operation of all cellular compartments during ontogeny and amidst stress circumstances[51].
Expression pattern analysis of LsfLEA genes
-
It is a well-established fact that the LEA genes constitute a pivotal component in facilitating the growth, development, and response to stressors in the plant kingdom[52]. Previous results have demonstrated that LEA genes are involved in the plants' reaction to abiotic adversities, most notably encompassing drought, low-temperature constraints, and high salinity stress[53]. Somatic embryogenesis, a developmental process that involves the formation of embryos from somatic cells, has gained recognition as a rapid and efficient means of propagation for woody plant species. Moreover, LEA genes were considered to be involved in somatic embryogenesis in cotton[54], white spruce[55] and sweet orange[56]. We identified the cis-elements of LsfLEA genes, which have been implicated in plant growth and development and stress response, including ABREs, AuxREs, GAREs, MeJAREs, SAREs, DREs, MBS, and MYC (Fig. 4). Similar cis-elements have been detected in the LEA gene promoter in Arabidopsis thaliana[4]and Prunus mume[13]. It has been widely established through previous investigations that the LEA protein exhibits a remarkable capability of being reconstituted by the CBF/DREB transcription factors, which has been found to play a pivotal role in the plants' intricate response mechanisms to the daunting challenges presented by the insidious cold and drought stressors that often threaten their survival and vitality[57]. The up-regulation of LEA gene expression has been found to be intricately linked with the overexpression of LBDREB. This phenomenon highlights the significant role played by LBDREB in regulating the expression of the LEA gene. It is important to note that the overexpression of LBDREB has been observed to elicit a robust response in the LEA gene expression, leading to its up-regulation[58]. In the intricate and multifaceted world of cellular biology, the regulation of proteins plays a crucial role in maintaining proper function and balance within the system. One such protein, LEA, has been found to be subject to the controlling influence of various transcription factors, including BHLH, MYB, and BZIP. These intricate regulators, with their multifarious modes of operation, serve to modulate the expression and activity of LEA protein, thereby contributing to the dynamic equilibrium of the cellular milieu[59, 60]. AtABI5 regulates LEA protein accumulation during dark-induced leaf senescence[60]. The transcription factor TABHLH49 emerging as a key player in the regulation of the wheat dehydration protein WZY2. The modulation of WZY2 levels by TABHLH49 has been shown to confer significant tolerance to the plant under conditions of water scarcity[59]. The transcription factor LcMYB2, which has been shown to play a crucial role in the activation of gene expression by directly binding to the promoter of AtLEAA14. These results suggest that the LsfLEA genes may be extensively involved in the response to abiotic stress in hybrid sweetgum through regulation of transcription factors. The LsfLEA genes have been found to encompass the majority of LsfLEA1, LsfLEA4, LsfSMP, as well as members of the LsfLEA3, LsfLEA5, and LsfDHN subfamilies. This discovery has revealed the intricate and multifaceted nature of the LsfLEA gene family, with its diverse and nuanced subfamilies contributing to a complex network of genetic regulation and expression. The presence of these various subfamilies highlights the importance of a holistic approach to the study of the LsfLEA genes, as well as the need for a deep understanding of the intricate molecular mechanisms underlying their function. In our quest for understanding the intricate workings of somatic embryogenesis, we endeavored to unravel the complex expression pattern of the LsfLEA genes across diverse stages of development. Our findings revealed a relative upsurge in the expression levels of LsfLEA1, LsfLEA5, and LsfSMP genes in the roots. Additionally, our investigation unraveled a relatively high expression level of LsfLEA1 and LsfSMP genes in the GE, HE, and TE stages of development. From these results, we infer that LsfLEA genes are intricately involved in the elaborate processes of somatic embryogenesis and development. It is noteworthy that these genes may fulfill divergent biological roles at different developmental stages, underscoring the complexity of this phenomenon.
-
In this study, the genome of hybrid sweetgum has revealed the presence of 79 LsfLEA genes, partitioned among eight subfamilies. The phylogenetic relationship, exon-intron structure, conserved domains and cis-acting elements of LsfLEA genes were analyzed. Transcriptome and qRT-PCR results showed that the expression of LsfLEA genes had certain specificity, which preliminarily revealed the function of LsfLEA genes in somatic embryogenesis. Subcellular localization results confirmed that the LsfLEA1-3 gene was localized in the nucleus. These results provide a pivotal reference point for future bioinformatic analyses and in-depth investigations into the profound and intricate function of LsfLEA genes in somatic embryogenesis.
This work was supported by the National Natural Science Foundation of China (No. 32271836), National Forestry and Grassland Administration Promotion Project of China (2020133102), the Project Fund (Somatic embryogenesis and efficient propagation technology in trees) Provided by Beijing Advanced Innovation Center for Tree Breeding by Molecular Design and the Fundamental Research Funds for the Central Universities(2019ZY39).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 The Amino acid sequence of LsfLEA genes.
- Supplemental Table S2 The Nucleotide sequence sequence of LsfLEA genes.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li Y, Qi S, Chen S, Li H, Zhang T, et al. 2023. Genome-wide identification and expression analysis of late embryogenesis abundant (LEA) genes reveal their potential roles in somatic embryogenesis in hybrid sweetgum (Liquidambar styraciflua × Liquidambar formosana). Forestry Research 3:12 doi: 10.48130/FR-2023-0012 

Genome-wide identification and expression analysis of late embryogenesis abundant (LEA) genes reveal their potential roles in somatic embryogenesis in hybrid sweetgum (Liquidambar styraciflua × Liquidambar formosana)
- Received: 12 March 2023
- Accepted: 04 May 2023
- Published online: 29 May 2023
Abstract: Late embryogenesis abundant (LEA) proteins are widely distributed in higher plants that play significant roles in embryonic development and abiotic stress response. Hybrid sweetgum is an important forest tree resource around the world, and somatic embryogenesis is an efficient way of reproduction and utilization. However, a systematic analysis of the LEA family genes in hybrid sweetgum is lacking, this is not conducive to the efficiency of its somatic embryogenesis. From the whole genome of the hybrid sweetgum, utilizing hidden Markov models, an identification of a total of 79 LEA genes was successfully conducted. They were classified into eight different groups based on their conserved domains and phylogenetic relationships, with the LsfLEA2 group of genes being the most abundant. The gene structure and sequence characteristics and chromosomal localization, as well as the physicochemical properties of LEA proteins were meticulously carried out. Analysis of the cis-acting elements shows that most of the LsfLEA genes are associated with light-responsive-elements. In addition, some genes are associated with biosynthetic pathways, such as abscisic acid response, growth hormone response, methyl jasmonate response, somatic embryogenesis, meristematic tissue expression. Furthermore, we systematically analyzed the expression patterns of hybrid sweetgum LEA genes in different stages of somatic embryogenesis and different tissues, in LEA family genes we also found significant specificity in gene expression during somatic embryogenesis. This study provides new insights into the formation of members of the LsfLEA family genes in hybrid sweetgum, while improving the understanding of the potential role of these genes in the process of hybrid sweetgum somatic embryogenesis and abiotic stress response. These results have a certain guiding significance for the future functional study of LsfLEA family genes, and provide a theoretical basis for exploring the regulatory mechanism of LsfLEA genes in the somatic embryo development stage of hybrid sweetgum.