










-
Age-related gene expression is essential for the growth and development of woody perennials, including the control of the growth rate[1], rooting ability[2], body architecture formation[3], wood formation[4−11], the juvenile period[12,13], and reproductive phase changes[12,13]. As trees age, many genes change their expression[14−19], indicating their functions and roles vary with development. Conifers are long-lived woody perennials that require a long time to bloom[20,21]. The conifer Larix kaempferi is characterized by rapid growth and is appreciated for its economic value. The juvenile period of L. kaempferi is about 10 years, which limits genetic improvement by sexual hybridization and production of improved varieties. In addition, to maintain the excellent traits of L. kaempferi mother trees, vegetative reproduction is commonly used. However, it often takes a long time to identify the excellent traits, and when these traits are identified, the rooting ability of cuttings decreases greatly. Thus, investigating the mechanisms underlying the effects of aging on L. kaempferi development is of theoretical and practical importance.
In our previous study, a comparative transcriptomic analysis of the uppermost main stems of 1-, 2-, 5-, 10-, 25-, and 50-year-old active L. kaempferi trees was performed to reveal the molecular aspects of aging on wood formation[5] and phase change[16], and it was found that many genes alter their expression with age[16]. Furthermore, seven transcription factors have age-related expression patterns[15]. L. kaempferi DEFICIENS-AGAMOUS-LIKE 1 (LaDAL1/LaAGL2-1), L. kaempferi AGAMOUS-Like 2-2 (LaAGL2-2), LaAGL2-3, and L. kaempferi SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1-1 (LaSOC1-1) show increased transcription at 5 years, LaAGL11 shows increased transcription at 3 years, and L. kaempferi APETALA 2-2 (LaAP2-2) shows decreased transcription at 1 year[15]. Further analysis reveals that both LaDAL1 and LaAP2-1 have age-dependent expression patterns in both active and dormant stages[14]. Similarly, age-dependent patterns have also been revealed for homologs of LaDAL1 in Picea abies[22], Pinus koraiensis[18], and Pinus tabuliformis[17], indicating that these DAL1 homologs have conservative roles and can be used to explore the age-mediated control of tree growth and development by studying their regulatory mechanisms. However, these studies have only detected the age-dependent expression patterns, and further studies are needed to understand how age regulates their expression via the underlying mechanisms.
Transcription and precursor messenger RNA (pre-mRNA) splicing are critical in the control of gene expression. In eukaryotes, pre-mRNA is transcribed by RNA polymerase II in a complicated process[23]. Genes in higher eukaryotes usually contain several introns. These introns are often of considerable length, sometimes extending to many thousands of bases. As such, they can account for 90% of the length of a typical pre-mRNA[24]. The vast majority of eukaryotic introns are not self-splicing, and the formidable task of identifying and splicing together exons among all the intronic RNA is performed by the spliceosome, a large ribonucleoprotein machine[25,26]. After splicing, the two concomitant exons are joined, and the intron is released as a lariat RNA[25,27] (Fig. 1). In previous studies, various age-dependent expression patterns have been revealed at the mature mRNA level[15]. However, it remains unclear which step from pre-mRNA transcription to splicing is regulated by age.
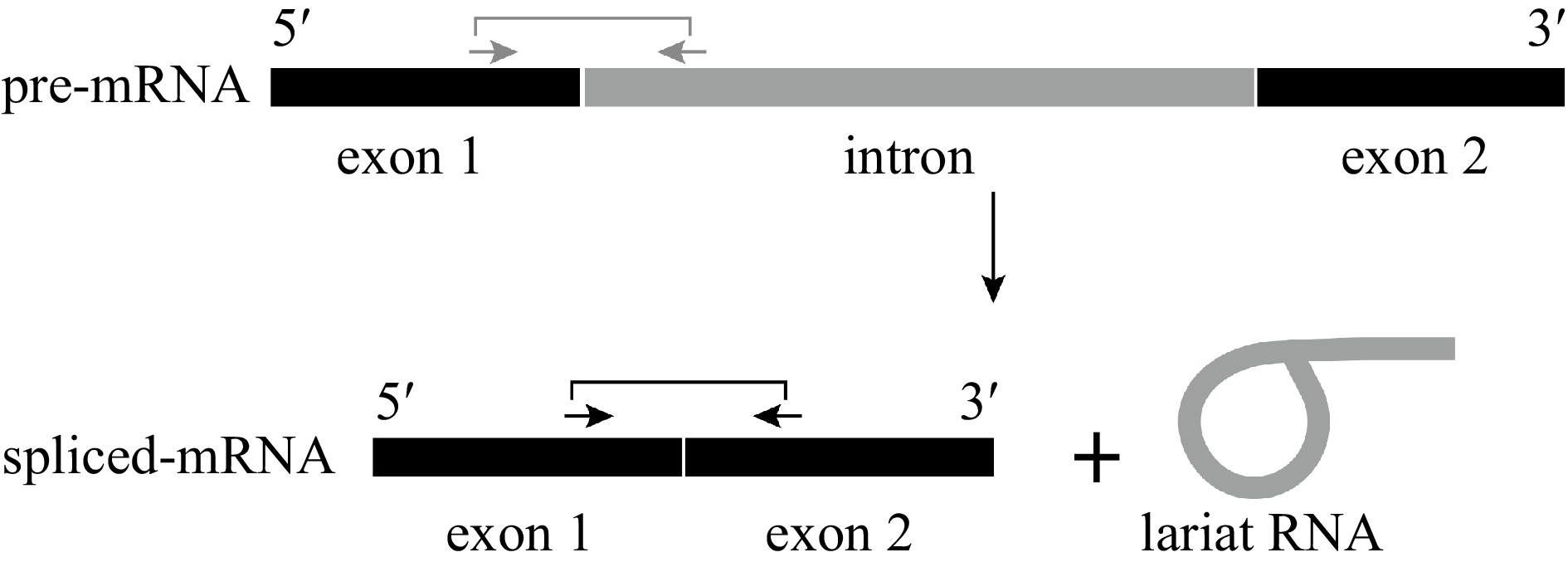
Figure 1.
Process of mRNA splicing. RNA splicing involves cutting an intron (gray) from pre-mRNA and joining together the two neighboring exons (black). The spliced exons form the functional RNA, and the intron is usually degraded. Arrows indicate the positions of the primers used to detect the pre-mRNA (gray) and spliced mRNA (black).
Here, we hypothesized that the expression of age-related genes may be regulated at the levels of transcription and mRNA processing. To gain further insights into the age-mediated regulation of gene expression in L. kaempferi, we studied pre-mRNA transcription and splicing efficiency of seven age-related genes during tree aging. Specific primers were designed to detect pre-mRNA and spliced mRNA changes in trees of different ages. Comparisons of pre-mRNA and spliced mRNA change fold can be instrumental in the analysis of splicing efficiency. Our results provide novel insights into the age-mediated regulation of gene expression in tree growth and development.
-
The DNA sequences of age-related transcription factors were obtained from the L. kaempferi genome[28]. The gene structure was analyzed using Gene Structure Display Server 2.0[29]. Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was used to examine the expression patterns of pre-mRNA and spliced mRNA. Specific primers were designed to detect pre-mRNA and spliced mRNA (
www.ncbi.nlm.nih.gov/tools/primer-blast/ , accessed on 20 March 2022) (Table 1 & Supplemental Table S1). For each gene, three pairs of primers were designed against the 5′ end, middle, and 3′ end.Table 1. Polymerase chain reaction primers for the amplification of age-related transcription factors.
Gene (accession: mRNA/DNA) Primer Sequence (5'-3') Position Size (bp) LaDAL1 (MN790744/WOXR02001943.1) I Forward-CGATGCAGAAGTGGCGCTAA first exon 94 Reverse-GTCAACAGCGCAAAGAAAGGA first intron II Forward-GCTCTCAGTGCTGTGCGAT first exon 249 Reverse-CCGAGATCTTCCCCCAACAAA fourth exon III Forward-AGACCAGATTGAGGAGCTTCG fifth exon 90 Reverse-GGGACGGAATAGCGTGCATTA fifth intron IV Forward-GGTTGAGCTCCTTCAGCGAT third exon 163 Reverse-GCGAAGCTCCTCAATCTGGT fifth exon V Forward-GTACTAACGGGCCTTGGGAT seventh exon 172 Reverse-TTCCAGCTTCAAAAGTGCCAAT seventh intron VI Forward-ACGCAGGTGATGCTAGACCA fifth exon 165 Reverse-CCAAGGCCCGTTAGTACCAG seventh exon LaAGL2-2 (MN790745/WOXR02003181.1) I Forward-GAGTTTGCTAGTGCCGGGTA first exon and first intron 207 Reverse-GTTGGGGGAAGATCTGGGTC first intron II Forward-GGGCTGCTGAAGAAAGCCTA first exon 110 Reverse-TTCATGCCGGCACTAGCAAA second exon III Forward-GTTGGGGGAAGATCTGGGTC fourth exon 242 Reverse-TGCGTTGTGTCGTATTTAGGTC fourth intron IV Forward-TGCAGCAACTCGAACATCAAC fourth exon 76 Reverse-TGGCCTAGCATAACCTGCG fifth exon V Forward-AATTCAAGCCTCCCGACTGT sixth exon 207 Reverse-TCCGGGGACTACATATTGGC sixth intron VI Forward-TGCTCTCTTACACCCGCAAC sixth exon 141 Reverse-CCACCACCCTTGCACGTAT seventh exon LaAP2-1 (MN790757/WOXR02007023.1) I Forward-CCCCGGAGTTCTGAGGAAAC first exon 197 Reverse-TTGCTAGAGGCCTCGTGTTC first intron II Forward-GCTCGCAATATCGTGGAGTG first exon 117 Reverse-TAGCAGCGGCATGAGCAGTA third exon III Forward-CAGCTATCAAGTTTCGAGGCG fourth exon 200 Reverse-ACTGCCATCCAAATGACTACC fourth intron IV Forward-TTCGAGGCGTTGAAGCTGAT fourth exon 131 Reverse-TCCACGAGAGAAACCAGTGC fifth exon V Forward-CTGAAGCTCACATGAGGGAGG ninth exon 158 Reverse-TCCGCTCAGTCCATCTTTATGC ninth intron VI Forward-CCTGACCATCTGGGTAACTGT ninth exon 159 Reverse-TACTGGAGTTGTTGGTCCGC tenth exon Plant materials
-
In our previous work, we found that the mature mRNA levels of LaDAL1, LaAGL2-2, LaAGL2-3, LaAGL11, and LaSOC1-1 increased from 1 to 7 years, whereas the mature mRNA levels of LaAP2-1 and LaAP2-2 decreased from 1 to 5 years[15]. Based on these results, in this work, we used 1-, 3-, 5- and 13-year-old active L. kaempferi trees that were collected on 4 July 2019[15]. These trees were grown from seeds and included the vegetative and reproductive phases of L. kaempferi. They were collected from a seed orchard in Dagujia (42°22′ N, 124°51′ E), Liaoning Province, in Northeast China. During sampling, main stems or lateral branches from the upper crowns were collected. After removal of buds or needles, the left stems from at least six trees from each age category were pooled, frozen in liquid nitrogen, and stored at −80 °C until RNA extraction.
RNA extraction, cDNA synthesis, and qRT-PCR
-
These experiments were carried out as previously described[15]. L. kaempferi translation elongation factor-1 alpha 1 (LaEF1A1) (GenBank accession no. JX157845), which was used as the internal control, is expressed stably during tree aging[5], and the primers 5′-GACTGTACCTGTTGGTCGTG-3′ and 5′-CCTCCAGCAGAGCTTCAT-3′ were used[30]. The relative expression ratio was expressed using the 2−ΔΔCt method[31]. The sample with the minimum or maximum age was used for normalization and was set to a value of 1. The qRT-PCR was performed with four technical replicates, and the data are shown as the mean ± standard deviation.
-
The LaDAL1 DNA sequence was 211,832 bp in length and contained eight exons and seven introns, with an open reading frame (ORF) of 774 bp, which encoded 257 amino acids (aa) (Fig. 2a). The exon length ranged from 42 to 182 bp, and the intron length ranged from 92 to 85,043 bp. The first, second, fourth, and fifth introns were more than 10 kb in length and were super-long introns (Fig. 2a). Fifteen CpG islands were predicted in the super-long introns (Fig. 2a).
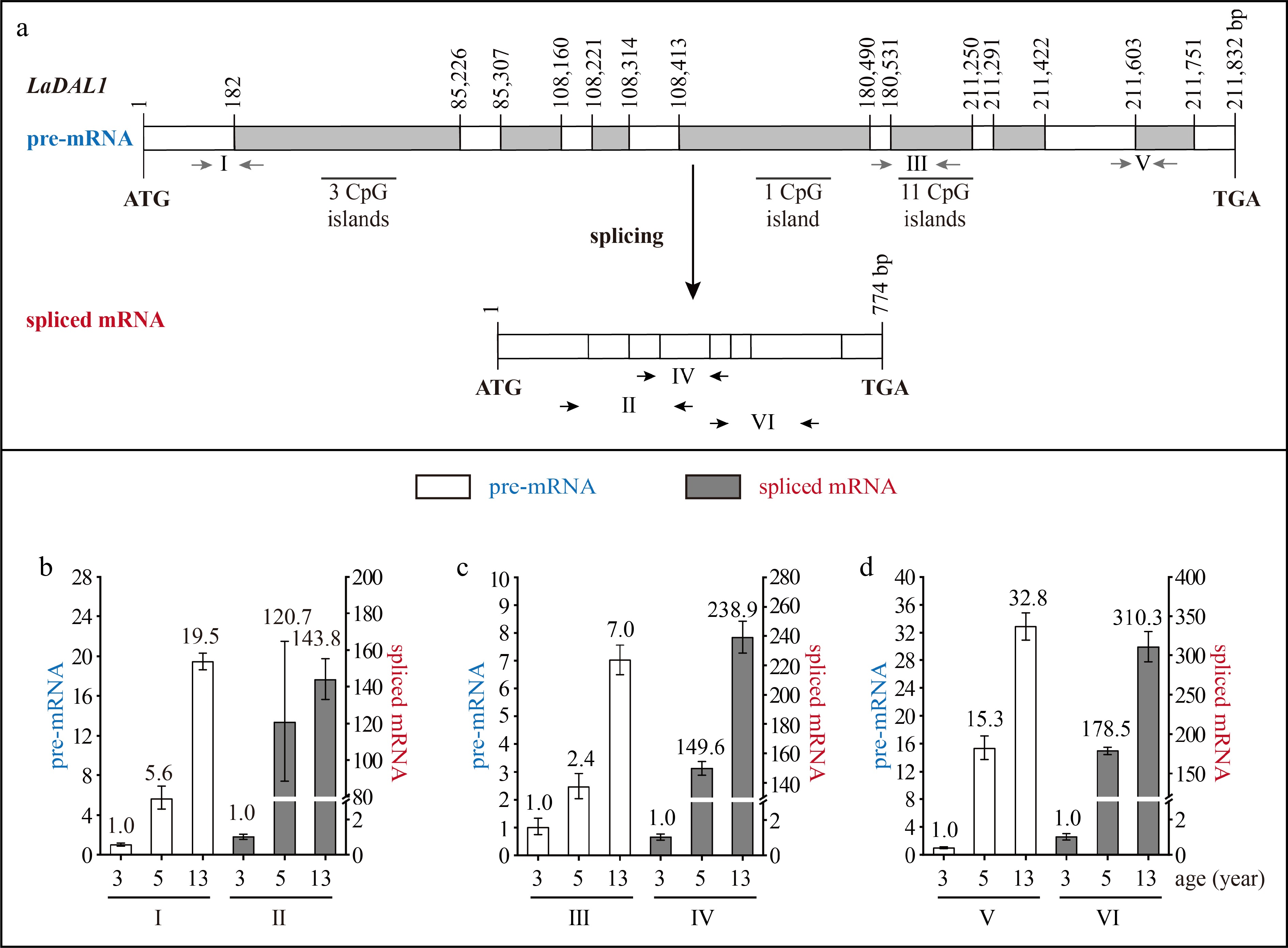
Figure 2.
Structure and expression pattern of LaDAL1. (a) Schematic representation of the gene structure of LaDAL1. White indicates exon; gray indicates intron; gray arrows indicate the positions of the primers used to measure the pre-mRNA; and black and horizontal arrows indicate the positions of the primers used to measure the spliced mRNA. After the prediction of DNA methylation, the CpG islands were revealed. (b)–(d) Expression patterns of LaDAL1 pre-mRNA and spliced mRNA during tree aging detected by three different primer pairs. The lateral branches of 3-, 5-, and 13-year-old active Larix kaempferi trees (n ≥ 6, sampled on 4 July 2019) were used to examine the expression patterns, which were assayed by qRT-PCR with LaEF1A1 as the internal control. The capitalized Roman numerals (I–VI) in panels (a)–(d) represent the different primers. The p-values of the differences between 5- and 3-year-old trees were calculated. One-way ANOVA Duncan’s test was used for statistical analysis.
LaAGL2-2 and LaAGL2-3 were two splice variants, because their transcript sequences were mapped to the same DNA sequence. This DNA sequence was 222,007 bp in length and contained seven exons and six introns (Fig. 3a). The transcript sequence of LaAGL2-2 was 783 bp in length, encoding a polypeptide of 260 aa, and that of LaAGL2-3 was 678 bp in length, encoding a polypeptide of 225 aa (Fig. 3a). The LaAGL2-3 spliced mRNA retained part of the sequence of its sixth intron in which there was a termination codon. The first, second, fourth, and fifth introns were super-long introns, and three CpG islands were predicted in the introns (Fig. 3a).
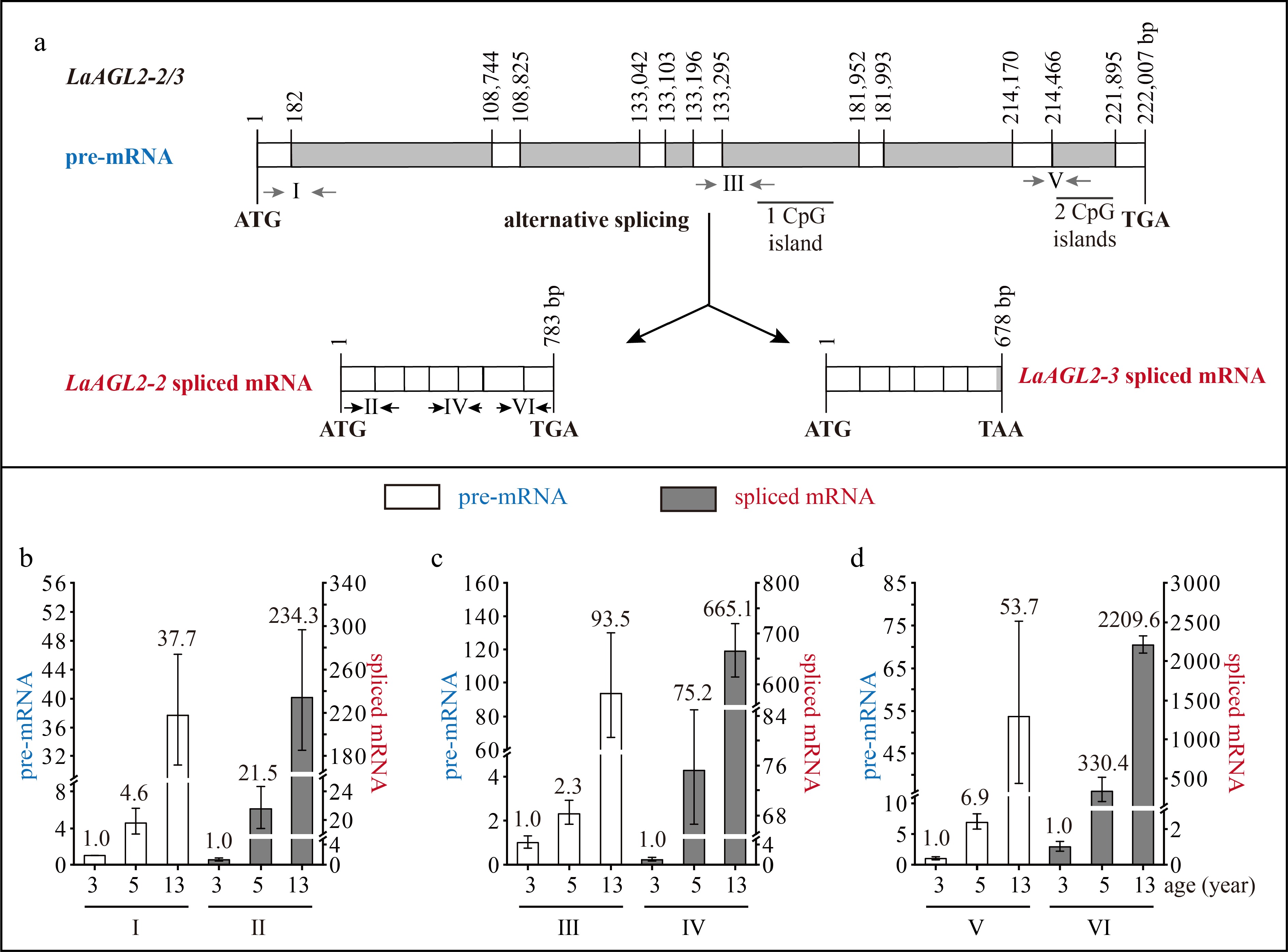
Figure 3.
Structure and expression pattern of LaAGL2-2/3. (a) Schematic representation of the gene structure of LaAGL2-2/3. White indicates exon; gray indicates intron; gray arrows indicate the positions of the primers used to measure the pre-mRNA; and black and horizontal arrows indicate the positions of the primers used to measure the spliced mRNA. After the prediction of DNA methylation, the CpG islands were revealed. (b)–(d) Expression patterns of LaAGL2-2/3 (b, c) and LaAGL2-2 (d) pre-mRNA and spliced mRNA during tree aging detected by three different primer pairs. The lateral branches of 3-, 5-, and 13-year-old active Larix kaempferi trees (n ≥ 6, sampled on 4 July 2019) were used to detect the expression patterns, which were assayed by qRT-PCR with LaEF1A1 as the internal control. The capitalized Roman numerals (I–VI) in panels (a)–(d) represent the different primers. The p-values of the differences between 5- and 3-year-old trees were calculated. One-way ANOVA Duncan’s test was used for statistical analysis.
The LaAP2-1 DNA sequence was 59,981 bp in length and contained ten exons and nine introns, with an ORF of 1,935 bp, which encoded 644 aa (Fig. 4a). The exon length ranged from 26 to 692 bp, and the intron length ranged from 107 to 55,979 bp. The fourth intron was a super-long intron, and one CpG island was predicted in the exon (Fig. 4a).
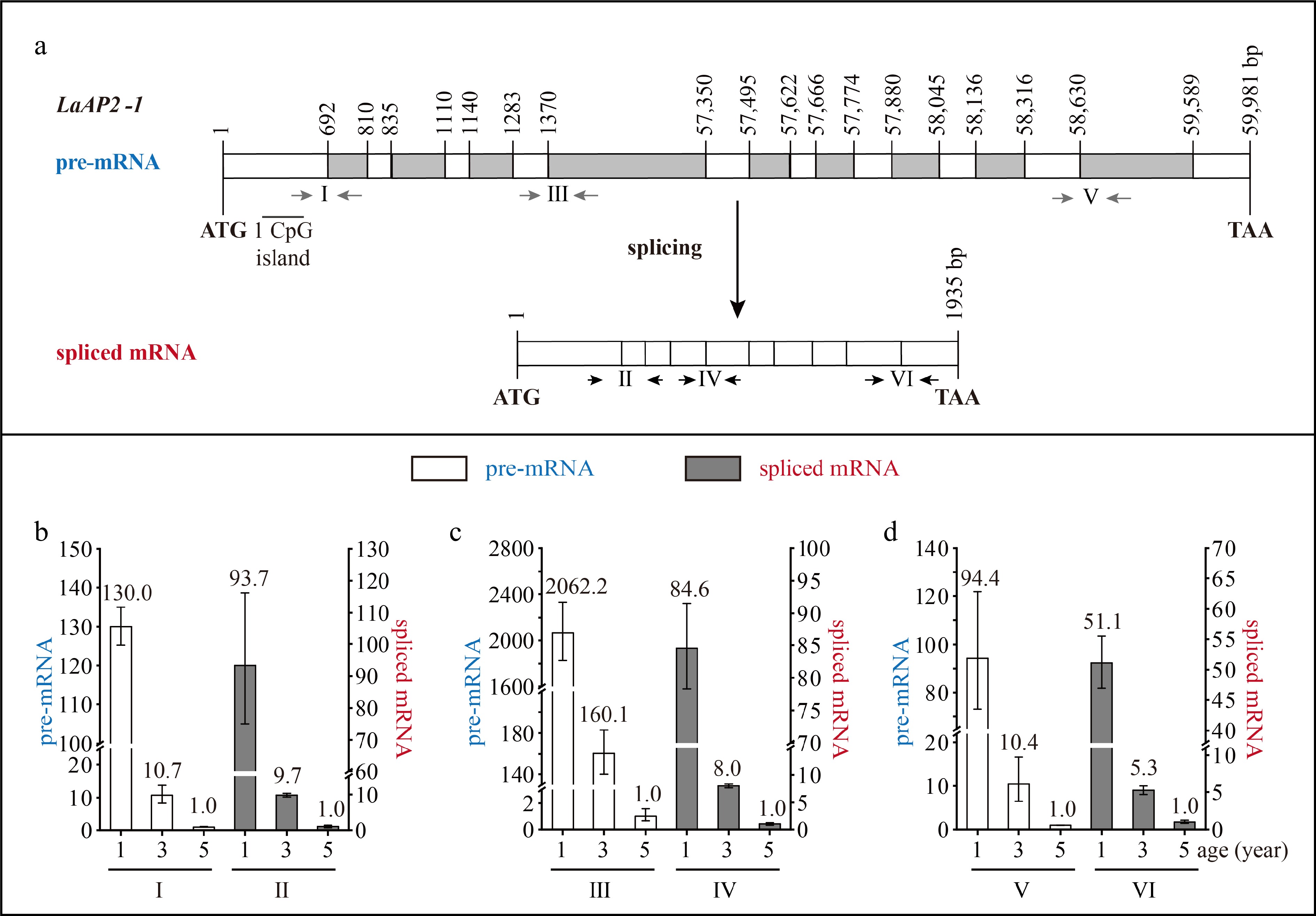
Figure 4.
Structure and expression pattern of LaAP2-1. (a) Schematic representation of the gene structure of LaAP2-1. White indicates exon; gray indicates intron; gray arrows indicate the positions of the primers used to measure the pre-mRNA; and black and horizontal arrows indicate the positions of the primers used to measure the spliced mRNA. After the prediction of DNA methylation, the CpG islands were revealed. (b)–(d) Expression patterns of LaAP2-1 pre-mRNA and spliced mRNA during tree aging detected by three different primer pairs. The lateral branches of 1-, 3-, and 5-year-old active Larix kaempferi trees (n ≥ 6, sampled on 4 July 2019) were used to detect the expression patterns, which were assayed by qRT-PCR with LaEF1A1 as the internal control. The capitalized Roman numerals (I–VI) in panels (a)–(d) represent the different primers.
Based on these data, we speculated that the existence of the super-long introns and CpG islands in these age-related genes may control their transcription by affecting site recognition and splicing by the spliceosome. Notably, in the P. tabuliformis DAL1 (PtDAL1) DNA sequence, super-long introns and CpG islands also exist, and two segments at the 5′ end of the first ultra-long intron show a gradual decline of CHG methylation with increasing age, which is negatively correlated with its expression[32], indicating that increased PtDAL1 transcription during tree aging may be controlled by CHG methylation in this intron. These results not only support our speculation but also provide new insights on the mechanism underlying the formation of age-dependent expression patterns.
Transcription of age-related genes is regulated by age
-
The LaDAL1 pre-mRNA level in 5-year-old trees was about 5.6-times higher as detected by primer I (Fig. 2b), 2.4-times higher as detected by primer III (Fig. 2c, p < 0.05), and 15.3-times higher as detected by primer V (Fig. 2d, p < 0.05) than that in 3-year-old trees. Compared with 5-year-old trees, a higher level of LaDAL1 pre-mRNA was detected in 13-year-old trees (Fig. 2b−d ). The LaAGL2-2/3 pre-mRNA level in 5-year-old trees was about 4.6-times higher as detected by primer I (Fig. 3b, p < 0.05) and 2.3-times higher as detected by primer III (Fig. 3c, p < 0.05) than that in 3-year-old trees. Compared with 5-year-old trees, a higher level of LaAGL2-2/3 pre-mRNA was also detected in 13-year-old trees. The LaAP2-1 pre-mRNA level in 1-year-old trees was about 103.0-times higher as detected by primer I (Fig. 4b, p < 0.05), 2,062.2-times higher as detected by primer III (Fig. 4c, p < 0.05), and 94.4-times higher as detected by primer V (Fig. 4d, p < 0.05) than that in 5-year-old trees.
Increased LaAGL11 and LaSOC1-1 pre-mRNA levels (Supplemental Figs S1 & S2), as well as a decreased LaAP2-2 pre-mRNA level, were also detected (Supplemental Fig. S3). These results showed that the pre-mRNAs of these seven age-related genes also have age-dependent expression patterns and that the age regulation of their transcription does occur. Notably, many CpG islands were predicted in the introns of five genes (LaDAL1, LaAGL11, LaAGL2-2/3, and LaSOC1) (Figs 2a & 3a, Supplemental Figs S1a & S2a), which are highly expressed in the reproductive stage. However, no CpG islands were predicted in the introns of two genes (LaAP2-1 and LaAP2-2) (Fig. 4a & Supplemental Fig. S3a), whose transcript levels decreased with tree aging, indicating that other regulatory mechanisms of the transcription of these age-related genes exist in addition to DNA methylation[32] and that age may control the expression of these up- and down-regulated genes with different mechanisms.
Pre-mRNA splicing may contribute to the formation of age-dependent expression patterns
-
Pre-mRNA is the initial product of transcription, and its level can be detected when the primer used for qRT-PCR is designed within the intron, which is removed when pre-mRNA is extensively edited by splicing. Splicing is a process in which pre-mRNA is used as a substrate to produce spliced mRNA under the action of the spliceosome[33]. In this process, the splicing efficiency determines the production rate of spliced mRNA. Based on this, we measured the pre-mRNA levels of seven age-related genes and their splicing efficiencies.
The levels of LaDAL1 pre-mRNA and spliced mRNA increased with age at different rates (Fig. 2b−d). For example, the LaDAL1 pre-mRNA level in 5-year-old trees was about 5.6-times higher than that in 3-year-old trees (Fig. 2b, p < 0.05), whereas the spliced mRNA level in 5-year-old trees was about 120.7-times higher than that in 3-year-old trees (Fig. 2b, p < 0.05). Differences in the increasing rates in LaDAL1 pre-mRNA and spliced mRNA levels also existed in 13- and 3-year-old trees. Compared with pre-mRNA, spliced mRNA showed a higher increase in its level in 5- and 13-year-old trees (Fig. 2b−d). Differences in the increasing rates in spliced mRNA levels were also detected in LaAGL2-2/3 (Fig. 3b−d) and LaAGL11 (Supplemental Fig. S1b−d). These results indicate that the production of spliced mRNA from the pre-mRNA may be enhanced with tree aging.
The levels of LaAP2-1 pre-mRNA and spliced mRNA decreased with age at different rates (Fig. 4b−d). For example, the LaAP2-1 pre-mRNA level in 1-year-old trees was about 2,062.2-times higher than that in 5-year-old trees (Fig. 4b, p < 0.05), whereas the spliced mRNA level in 1-year-old trees was about 84.6-times higher than that in 5-year-old trees (Fig. 4b, p < 0.05). Differences in the decreasing rate in LaAP2-1 pre-mRNA and spliced mRNA levels were also detected in 3- and 5-year-old trees. Compared with spliced mRNA, pre-mRNA showed a smaller decrease in its level in 3- and 5-year-old trees (Fig. 4b−d), indicating that the production of LaAP2-1 spliced mRNA from its pre-mRNA may be reduced in 3- and 5-year-old trees.
During tree aging, a greater increase in the LaAGL2-2 transcript level was detected compared to that in the LaAGL2-3 transcript level, because the LaAGL2-2 and LaAGL2-3 spliced mRNA levels in 13-year-old trees were about 234.3-times higher than those in 3-year-old trees as detected by primer II (Fig. 3b, p < 0.05), whereas the LaAGL2-2 spliced mRNA level in 13-year-old trees was about 2209.6-times higher than that in 3-year-old trees as detected by primer VI (Fig. 3d, p < 0.05), which can only amplify LaAGL2-2 (Fig. 3a). These data indicate that the alternative splicing of LaAGL2-2/3 is controlled by age. It has been shown that light-mediated[34] and temperature-mediated[35] pre-mRNA alternative splicing in plants can regulate gene expression to promote morphogenesis and stress resistance. The role and significance of the alternative splicing of LaAGL2-2/3 during aging require further study.
-
Based on these data, we conclude that age affects the pre-mRNA transcription of seven age-related genes. Furthermore, our findings show that pre-mRNA splicing plays a role in controlling LaDAL1, LaAGL11, LaAGL2-2/3, and LaAP2-1, and their increased or decreased spliced mRNA levels during tree aging result from the increased or decreased transcription and the enhanced or reduced pre-mRNA splicing efficiency, respectively.
This work was supported by the National Natural Science Foundation of China (32271904). The authors thank Yao Zhang (Chinese Academy of Forestry) for help in sampling. We would like to thank A&L Scientific Editing (
www.alpublish.com ) for its linguistic assistance during the preparation of this manuscript.-
The authors declare that they have no conflict of interest. Wanfeng Li is the Editorial Board member of Forestry Research who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and his research groups.
- Supplemental Fig. S1 The structure and expression pattern of LaAGL11.
- Supplemental Fig. S2 The structure and expression pattern of LaSOC1-1.
- Supplemental Fig. S3 The structure and expression pattern of LaAP2-2.
- Supplemental Table S1 Three age-related Larix kaempferi transcription factors and their qRT-PCR primers.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li X, Cheng D, Qi L, Zhan J, Li W. 2023. Regulation of age-dependent expression patterns of five transcription factors in Larix kaempferi. Forestry Research 3:18 doi: 10.48130/FR-2023-0018 

Regulation of age-dependent expression patterns of five transcription factors in Larix kaempferi
- Received: 04 May 2023
- Accepted: 20 July 2023
- Published online: 31 July 2023
Abstract: To reveal the regulatory mechanisms underlying age-dependent expression patterns, we characterized seven age-related genes, LaDAL1, LaAGL2-2, LaAGL2-3, LaAGL11, LaSOC1-1, LaAP2-1, and LaAP2-2 in terms of transcription and intron splicing in Larix kaempferi. Based on the exon–intron structures, we quantified the pre-mRNA levels and mature mRNA levels of these seven genes using quantitative reverse transcription polymerase chain reaction experiments. We found that the pre-mRNA levels manifested age-related patterns, indicating that their transcription was primarily regulated by age. By comparing the increasing or decreasing rates of the pre-mRNA and spliced mRNA levels, we found that their splicing efficiencies also changed with age. These results clearly show that both pre-mRNA transcription and splicing of five age-related genes are regulated by age, indicating that age-dependent expression patterns are controlled at both transcriptional and post-transcriptional levels, and unveiling the underlying regulatory molecular mechanisms should focus on the transcription factors, epigenetic regulation, and RNA splicing. These data provide new insights into the age-mediated regulation of gene expression in woody perennials in terms of longevity.
-
Key words:
- APETALA 2 /
- Gene expression /
- Larch /
- MADS-box /
- Tree /
- Post-transcriptional regulation