










-
Industrialization and urbanization have increased the emission of heavy metals, which has become a global problem due to the adverse effects on biosystems and public health[1−4]. Heavy metals, e.g. cadmium (Cd), lead (Pb), chromium (Cr), copper (Cu), manganese (Mn) and zinc (Zn), are major sources of soil pollution and they receive increasing attention[5−8]. It has been widely reported that heavy metal stress negatively affects plant growth, decreases photosynthesis and disturbs biochemical and physiological processes[4,9,10]. Heavy metal stress usually increases the production of ROS, such as
${\text{O}^-_2} $ Plants act as bioaccumulators that extract and concentrate heavy metals from the soil and water[13,15,16], and they have a series of defense mechanisms to cope with heavy metals. Plants commonly allocate heavy metals into the roots and stems, restrict transportation to the leaves and protect the photosynthetic cells from heavy metal damage[7,13,17,18]. In addition, plants can modify gene expression[19], upregulate antioxidant enzymes, such as APX, CAT and POD, scavenge ROS and alleviate the oxidative damage induced by heavy metals[14,20]. Many studies have previously investigated plants' responses to heavy metals; however, the knowledge is limited concerning quantitative assessment at regional and global levels, especially in dioecious plants.
Renner[21] reported that there are 15,600 dioecious angiosperm species that account for 5%−6% of all plant species. Under unfavorable conditions, the greater reproduction costs of females to produce flowers, seeds and fruits result in higher sensitivity and worse performance compared to males[22−25]. In addition, the different responses in females and males may lead to sex ratio biases that potentially affects the structure and stability of ecosystems[23,24,26]. Thus, especially dioecious plants may be at risk and vulnerable to environmental changes due to the sex-specific responses in growth, physiology and morphology under stress conditions, which further reinforces the spatial sexual segregation[27−29].
Populus species generally have fast growth rates and they are distributed in the Northern Hemisphere. The small genome size, clonal propagation and commercial values have made Populus species excellent model plants to study trees' responses to environmental stresses[29−33]. In addition, Populus species are dioecious, and separate female and male individuals may have different responses under unfavorable conditions[34−37]. For example, P. cathayana males have higher plasticity in photosynthetic activity, and females show more severe damage to cellular ultrastructure under Pb stress[38]. P. deltoides females suffer greater negative effects under Cd stress and show higher levels of leaf symptoms, lipid peroxidation and damage to the cellular ultrastructure[39]. Despite some previous research activity, there is still limited quantitative information on the region patterns of the responses of dioecious Populus trees to heavy metals.
In the present study, we performed a meta-analysis with 946 sets of comparative data from 20 publications to evaluate the responses of biomass accumulation and allocation, photosynthetic capacity, oxidative stress and antioxidants in Populus females and males under heavy metal exposure, such as Cu, Mn, Zn, Pb and Cd. We aimed to answer the following questions: (1) Whether Populus females and males also exhibit different responses to heavy metals on a regional scale? If yes, (2) whether Populus males perform better and have higher resistance compared to females under heavy metal exposure?
-
Peer-reviewed articles, other academic papers, and book chapters reporting the effects of heavy metals on antioxidant enzyme activities and on the concentration of heavy metals in different organs of male and female poplars published before May 2023 were searched in Web of Science and China National Knowledge Infrastructure (CNKI). We used the following keywords ('sexual' OR 'male and female' OR 'sex-related') AND ('heavy metal' OR 'Cd stress' OR 'Zn stress' OR 'Mn stress' OR 'Pb stress' OR 'Cu stress' OR 'aluminum' OR 'uranium') AND ('enzymatic activity' OR 'reactive oxygen species' OR 'ROS' OR 'MDA' OR 'CAT' OR 'POD' OR 'SOD' OR 'NPT' OR '
${\text{O}^-_2} $ The data are mainly extracted from the main text and tables of the primary studies, The GetData Graph Digitizer (version 2.26,
www.getdata-graph-digitizer.com ) was used to extract data from figures. After extraction and compilation, we had collected a total of 946 sets of comparative data (32 for MDA, 36 for${\text{O}^-_2} $ Data analysis
-
We calculated the effect of the heavy metal stress on the growth, photosynthesis capacity, antioxidant capacity, and heavy metal content in different organs of poplar trees. Natural log response ratios (lnRR) for each pairwise comparison were derived using the following equation[40−42]:
$ {\rm{LnRR}} = \left( {\frac{{\overline X t}}{{\overline X {\text{c}}}}} \right) $ (1) Where
$\overline X_t $ $\overline X_{\rm c} $ $ Wr=(Nc\times Nt)/(Nc+Nt) $ (2) where Wr is the weight associated with each lnRR, and Nt and Nc are the number of repeats in heavy metal and control groups, respectively.
We calculated the weighted mean effect sizes (lnRR++) for the growth, photosynthesis, antioxidant capacity, and heavy metal concentration in different organs. We employed linear mixed-effects models exclusively focusing on the intercept. The response variable for these models was represented by lnRR. Furthermore, we incorporated the identity of primary studies from which the data were collected as a random-effects factor. This factor enabled us to address any potential non-independence of observations derived from the same primary study[44,45]. The implementation of the linear mixed effect model and meta-regression was conducted using the restricted Maximum Likelihood Estimation (MLE) method within the lme4 software package[46]. To aid the interpretation of results, we back-transformed lnRR++ and the associated 95% confidence intervals (CI) using the equation of (elnRR++-1) × 100%[43]. All relevant statistical analyses were performed in R version 4.3.1[47]. All figures were prepared using Origin 9.0 (OriginLab) software.
-
Heavy metals were found to have positive effects on the concentrations of Cu, Mn, Zn, Pb and Cd as well as on oxidative stress and antioxidant enzyme activities, but negative effects on growth and photosynthetic capacity (Fig. 1). The concentrations of heavy metals significantly increased in different organs, except the Cu concentration of roots (Fig. 1a). Under heavy metal exposure, the total biomass, leaf biomass, stem biomass, root biomass, height and R/S ratio decreased by 29.78%, 33.41%, 27.22%, 35.30%, 34.83% and 6.50%, respectively. Furthermore, total chl, Pn, gs, E and Ci decreased by 23.30%, 26.03%, 40.49%, 23.76% and 18.24%, respectively (Fig. 1b). Oxidative stress variables TBARS, H2O2,
${\text{O}^-_2} $ 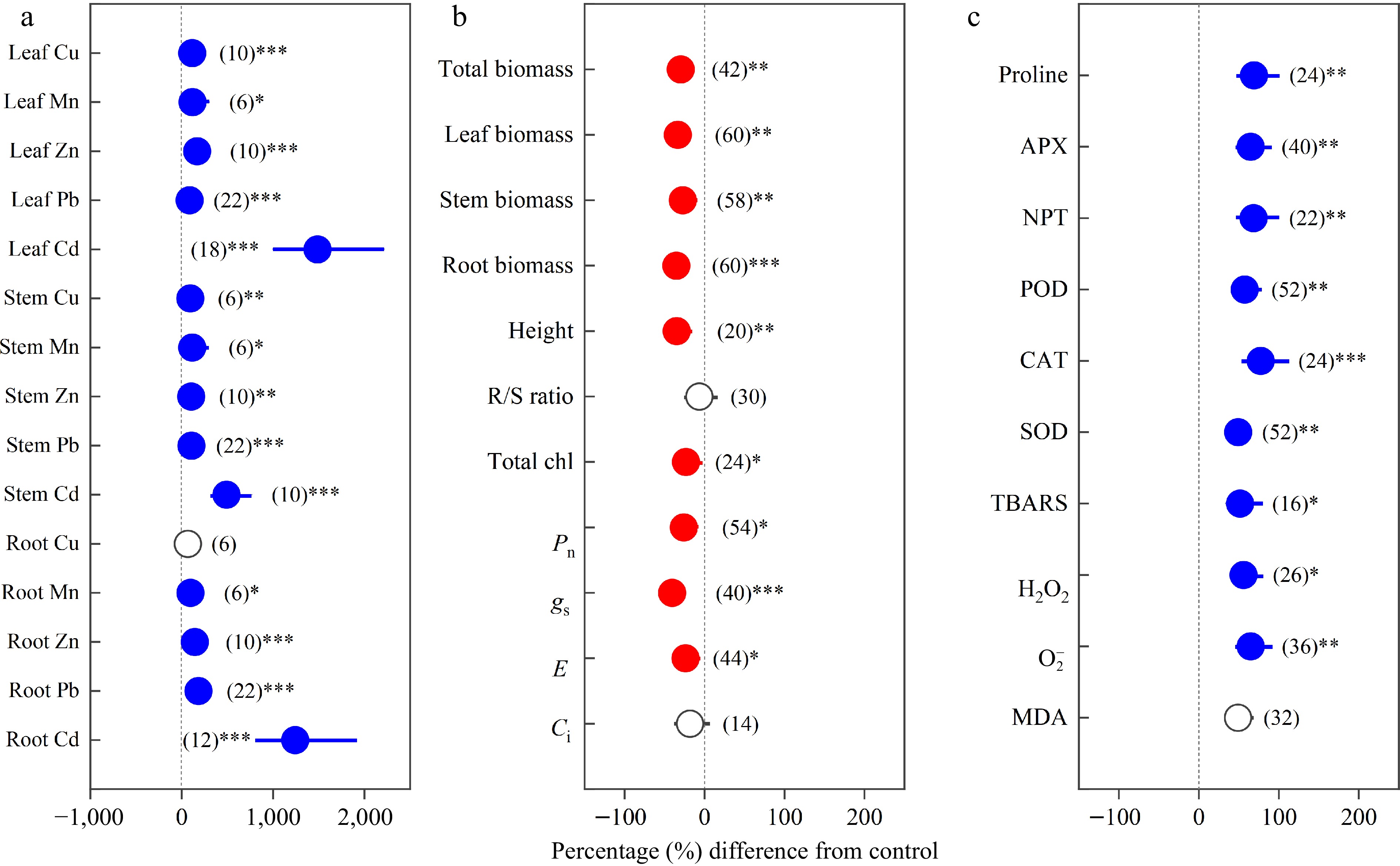
Figure 1.
Overall effects of heavy metals on the concentrations of (a) Cu, Mn, Zn, Pb, Cd in leaf, stem and root; (b) total biomass, leaf biomass, stem biomass, root biomass, height, R/S ratio, total chl, Pn, gs, E and Ci; (c) proline, APX, NPT, POD, CAT, SOD, TBARS, H2O2, ${\text{O}^-_2} $ and MDA. Values are means with 95% confidence intervals. The number of observations for each variable is shown in parentheses. The blue color indicates significant positive effects, and the red color indicates significant negative effects.*p < 0.05,**p < 0.01,*** p < 0.001.
Sexual differences in heavy metal concentrations, growth and antioxidants
-
Heavy metals significantly increased leaf Cu concentration in males, leaf Cd concentrations in both sexes, stem Zn concentrations in both sexes, and root Pb and Cd concentrations in both sexes (Fig. 2). There were no significant differences in the concentrations of heavy metals between females and males.
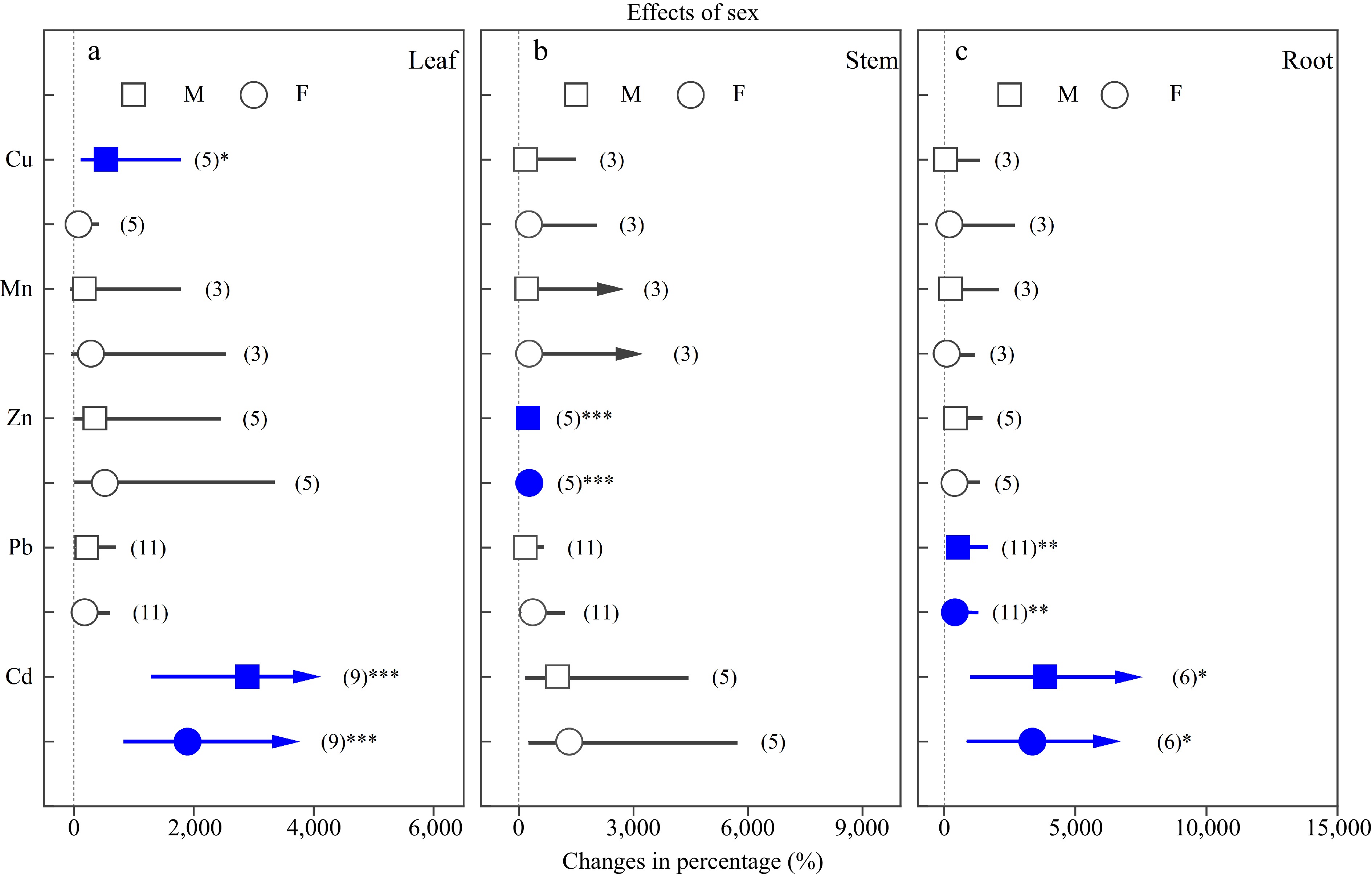
Figure 2.
Impacts of sex on the effects of heavy metals on the concentrations of Cu, Mn, Zn, Pb and Cd in (a) leaf, (b) stem and (c) root. Values are means with 95% confidence intervals. The blue color indicates significant positive effects.*p < 0.05,**p < 0.01,*** p < 0.001.
Root biomass, R/S ratio and height decreased in females by 36.43%, 17.15% and 33.87%, respectively, while these parameters decreased in males by 21.63%, 1.11% and 9.85%, respectively (Fig. 3). In addition, total chl, Pn, gs and E in females decreased by 29.21%, 32.71%, 48.77% and 36.80%, respectively, while in males these parameters decreased by 16.63%, 28.00%, 47.79% and 39.52%, respectively (Fig. 4). Compared with males, Populus females showed significantly greater declines (p < 0.05) in root biomass, R/S ratio, height and total chl under heavy metal exposure.
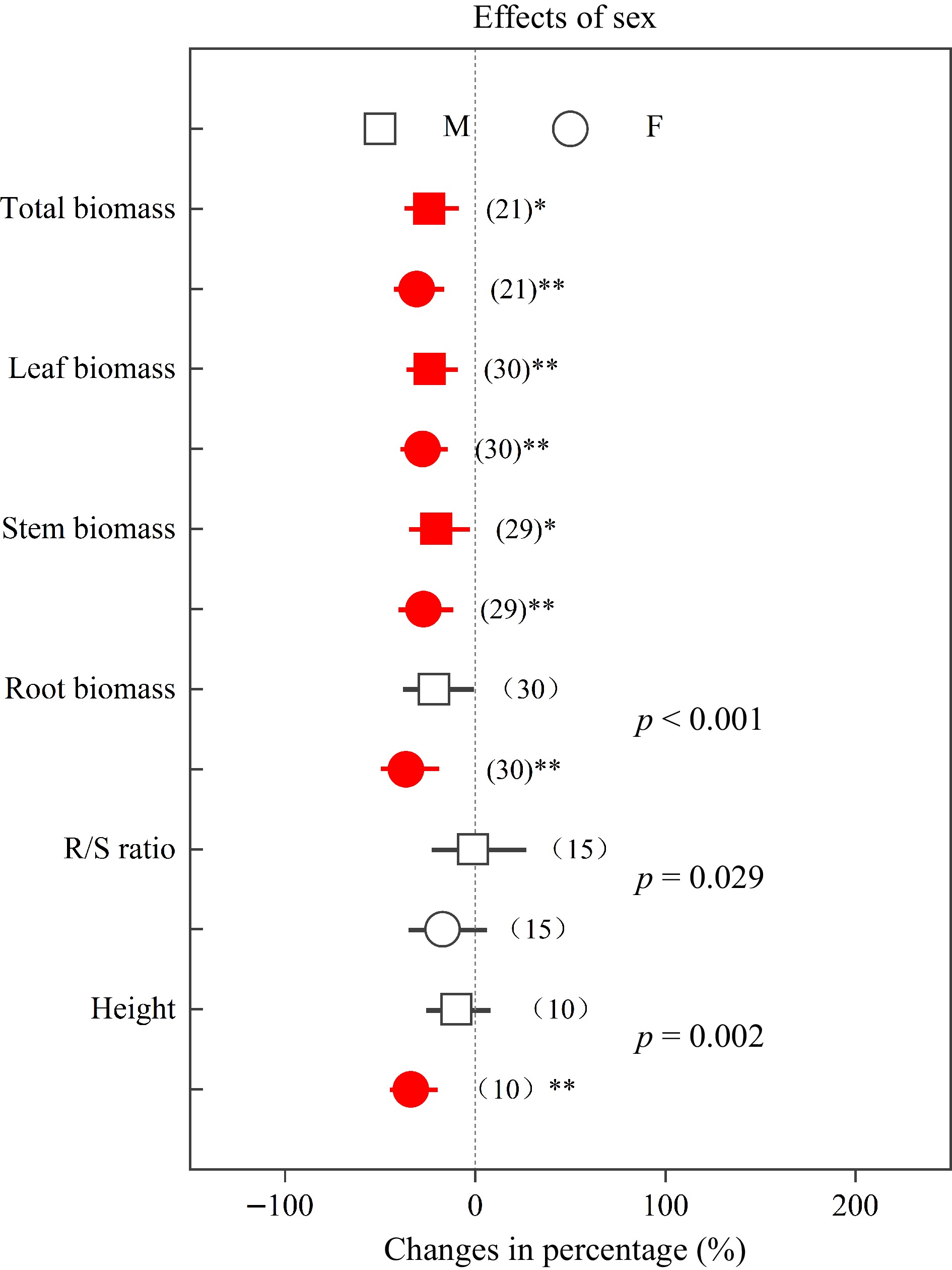
Figure 3.
Impacts of sex on the effects of heavy metals on total biomass, leaf biomass, stem biomass, root biomass, R/S ratio and height. Values are means with 95% confidence intervals. The p-values indicate differences between Populus sexes, and the red color indicates significant negative effects.*p < 0.05,**p < 0.01,*** p < 0.001.
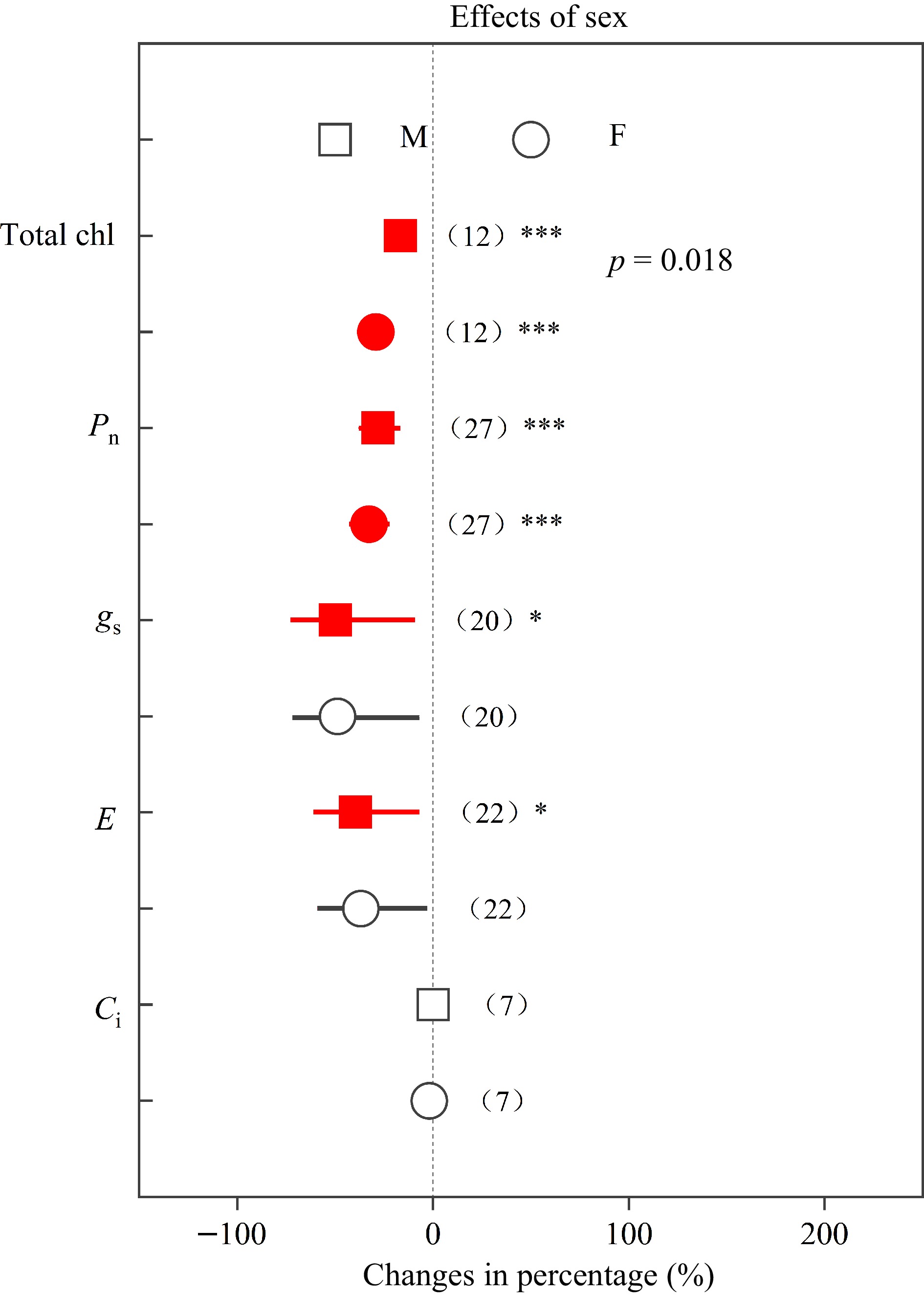
Figure 4.
Impacts of sex on the effects of heavy metals on total chl, Pn, gs , E and Ci. Values are means with 95% confidence intervals. The p-values indicate differences between Populus sexes, and the red color indicates significant negative effects.*p < 0.05,**p < 0.01,*** p < 0.001.
Heavy metals significantly increased oxidative stress in both sexes, except TBARS in males.
${\text{O}^-_2} $ 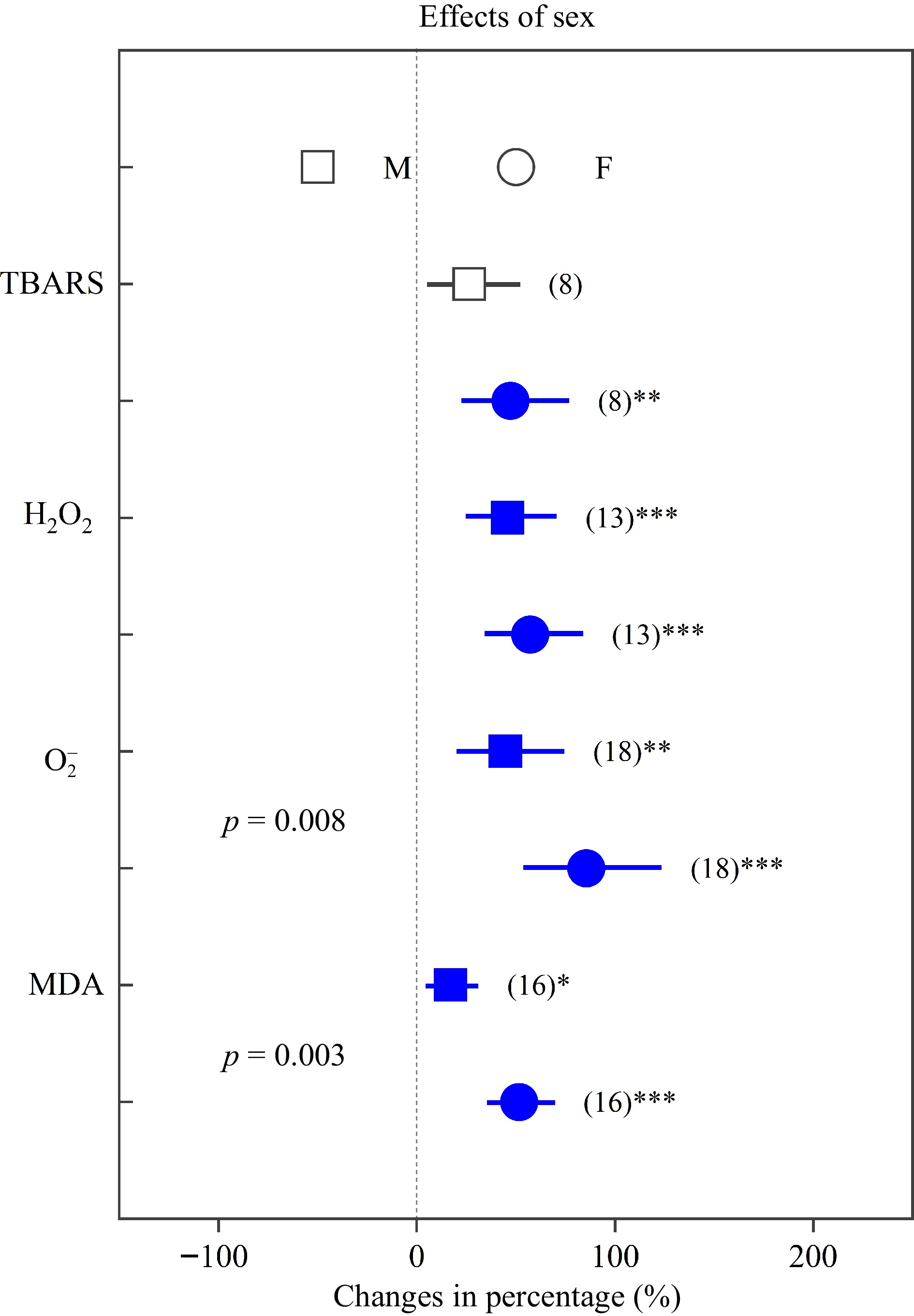
Figure 5.
Impacts of sex on the effects of heavy metals on the concentrations of TBARS, H2O2, ${\text{O}^-_2} $, and MDA. Values are means with 95% confidence intervals. The p-values indicate differences between Populus sexes, and the blue color indicates significant positive effects.*p < 0.05,**p < 0.01,*** p < 0.001.
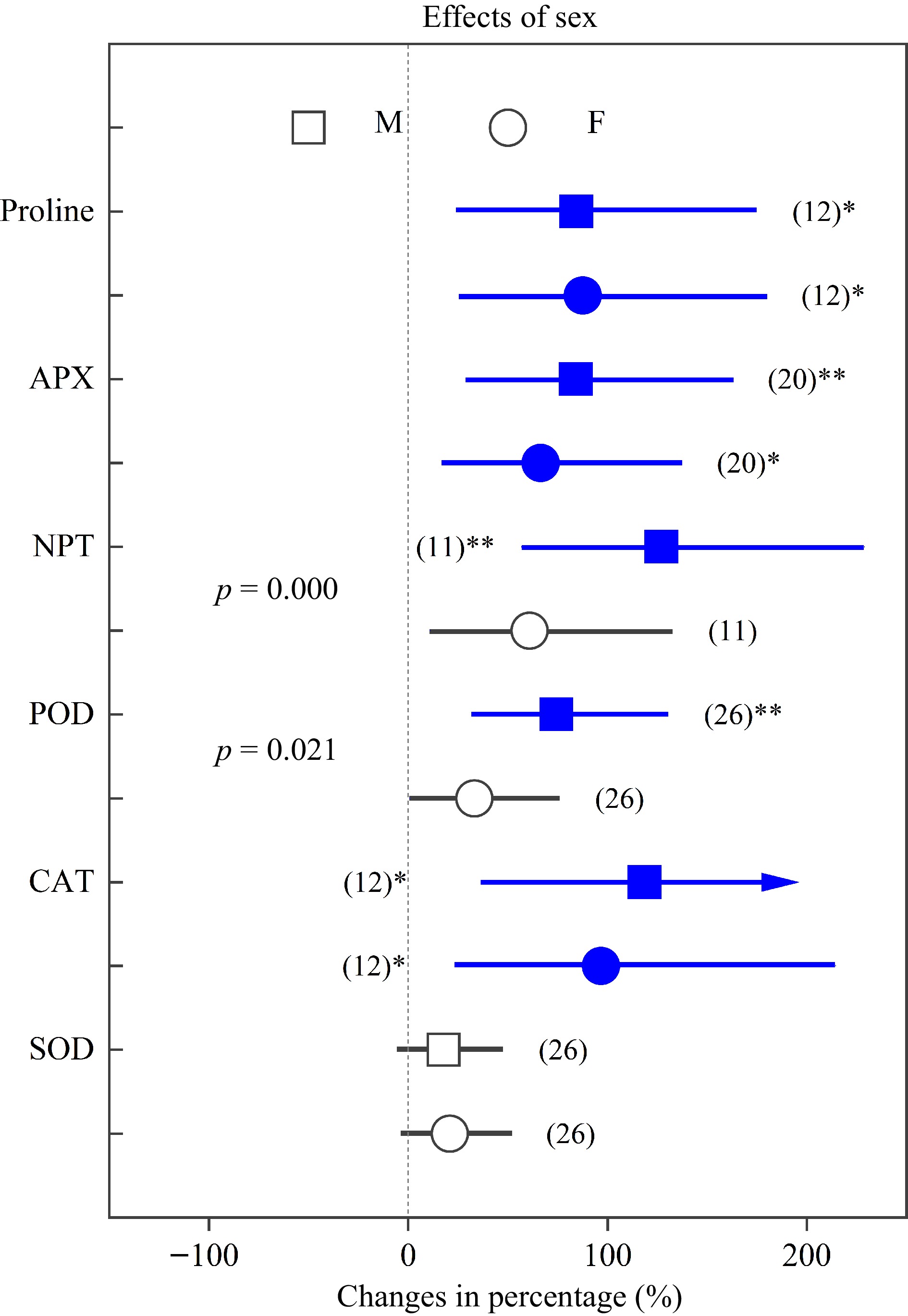
Figure 6.
Impacts of sex on the effects of heavy metals on the concentrations of proline, APX, NPT, POD, CAT and SOD. Values are means with 95% confidence intervals. The p-values indicate differences between Populus sexes, and the blue color indicates significant positive effects.*p < 0.05,**p < 0.01,*** p < 0.001.
-
In the present study, heavy metal treatment significantly increased average concentrations of Cu, Mn, Zn, Pb and Cd in different organs, except Cu in roots (Fig. 1a), which agreed with previous reports showing that Populus trees, as the bioaccumulators, can extract heavy metals from contaminated soils[38,48,49]. In addition, Populus can inhibit heavy metal transport to leaves and allocate more into stems and roots[39,50,51]. Our results were in line with the above statements. Leaf Cu concentration in males increased significantly more than that in females under Cu exposure. The reason was that the transpiration stream is the main way for Cu transport from roots to shoots[50,52,53]. At the same time, the average stem and root Cu concentrations of males increased by 170.28% and 63.20%, respectively, while these of females increased by 258.33% and 207.47%, respectively, implying males having a better ability to inhibit Cu transportation.
We found that Pb concentrations of roots in both sexes significantly increased, while Pb concentrations of leaves and stems showed no significant differences compared with the control treatments. These results indicated that the Pb accumulation was higher in roots, which is consistent with previous studies[54−56]. Cd is a highly toxic heavy metal for both plants and humans. A recent study have found that Cd translocation and reallocation was sex-dependent and that females showed greater upward transport of Cd, whereas males had greater downward transport, indicating males had greater capacity to restrict Cd transportation and protect the photosynthetic cells from heavy metal damage[8]. Some earlier studies have reported that Cd absorption and accumulation are affected by nitrogen levels[8,57] and plant-plant interactions[18], which indicate the complex nature of heavy metal absorption and accumulation and may explain the non-significant differences in the concentrations of heavy metals between Populus females and males shown by the present meta-analysis.
Growth and photosynthetic capacity affected by heavy metals
-
Similarly as documented elsewhere[7,8,20,50,53,55,56], we found that heavy metals have negative effects on plant growth, total chl, and photosynthetic capacity in both sexes. On the other hand, root biomass and height in females decreased by 36.43% and 33.87%, respectively, while these parameters decreased in males less, namely by 21.63% and 9.85%, respectively (Fig. 3). Compared with males, females had significantly greater decreases in root biomass, R/S ratio and height, indicating that females may be more sensitive and suffer greater negative effects, which was in accordance with earlier studies[50,53,55]. Previous studies have demonstrated that plant roots play key roles in the absorption of nutrients and water[58−60]. Females with smaller root biomass and R/S ratio may have a lower capacity to absorb resources, and this could explain the more negative effects observed in females under heavy metal stress. Previous studies have reported that Pb and Cd stress induced more severe damage and decreased number of chloroplasts in females[38, 53], which may explain that females had a significantly greater decline in total chl (Fig. 4), and implying that Populus males have more efficient heavy metal responses. Thus, the photosynthetic pigments of males may be better protected, while the photosynthesis machinery of females is more sensitive to heavy metal stress[50].
Oxidative stress and antioxidants affected by heavy metals
-
It is well known that heavy metals can result in an enhanced ROS accumulation, and they can cause oxidative damage to cellular membranes and proteins[21,35,61]. We discovered in the present study that heavy metals induce oxidative stress, for instance, the levels of TBARS, H2O2,
${\text{O}^-_2} $ ${\text{O}^-_2} $ ${\text{O}^-_2} $ On the other hand, plants usually upregulate their antioxidant enzyme activities to cope with heavy metals. We found that the levels of APX, NPT, POD, CAT and SOD increased by 64.81%, 68.40%, 57.34%, 77.30% and 49.01%, respectively, which closely correlated with oxygen-scavenging and indicated important roles for antioxidant enzymes when dealing with abiotic stress[32,60,62,63]. In addition, NPT and POD increased significantly more in males than females. The above statements collectively demonstrated that Populus males have higher antioxidant enzyme activities, better abilities to scavenge ROS and a more effective antioxidant defense system under heavy metals[7,8,50,51].
Previous studies have demonstrated that sexual dimorphism in Populus may lead to niche segregation, bias in sex ratios, and spatial segregation of the sexes (SSS) across different environmental gradients[22,28,29]. Less stress and resource-rich areas are usually with female-biased sex ratios, whereas males are more abundant under adverse and stressful conditions[22]. Thus, dioecious plants are more vulnerable under the future climate change due to SSS across environmental gradients[27]. The sex-specific responses and adaptive strategies of Populus may result in a situation that one sex is more prone to future climate change than the other one. These results are important for understanding sexual dimorphism, spatial sexual segregation and sex ratio biases, which may be reinforced in Populus with the increasing heavy metal pollution in the future.
-
In the present meta-analysis, we quantified the responses of growth, photosynthetic capacity, oxidative stress and antioxidants in Populus females and males to heavy metals at a region scale. Heavy metals have negative effects on Populus growth and photosynthetic capacity, and increased oxidative stress and antioxidants. Although we did not discover significant differences in the concentrations of heavy metals between females and males, there were still some sex-specific responses to heavy metals. Females suffered more negative effects, as they showed significantly more increased
${\text{O}^-_2} $ -
The authors confirm contribution to the paper as follows: study conception and design: Yu L, Li C; data collection: Yu L, Tang S, Kang J; analysis and interpretation of results: Yu L, Korpelainen H, Li C; draft manuscript preparation: Yu L, Korpelainen H, Li C; All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available from the corresponding author upon reasonable request.
The authors thank Dr. Chaoxiang Yuan for assistance in data analysis. This work was supported by the Natural Science Foundation of China (32001287) and the Talent Program of the Zhejiang University (0022112).
-
The authors declare that they have no conflict of interest. Chunyang Li is the Editorial Board member of Forestry Research who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and his research groups.
- Supplemental Fig. S1 Frequency distribution of the data for heavy metal concentrations of (a) Leaf Cu, (b) Leaf Mn, (c) Leaf Zn, (d) Leaf Pb, (e) Leaf Cd, (f) Stem Cu, (g) Stem Mn, (h) Stem Zn, (i) Stem Pb, (j) Stem Cd, (k) Root Cu, (l) Root Mn, (m) Root Zn, (n) Root Pb, and (o) Root Cd.
- Supplemental Table S1
- Supplemental Fig. S2 Frequency distribution of the data for heavy metal concentrations of (a) Total biomass, (b) Leaf biomass, (c) Stem biomass, (d) Root biomass, (e) Height (f) R/S ratio, (g) Total chl, (h) Pn, (i) gs, (j) E, and (k) Ci.
- Supplemental Fig. S3 Frequency distribution of the data for heavy metal concentrations of (a) TBARS, (b) H2O2, (c) O2-, (d) MDA, (e) Proline, (f) APX, (g) NPT, (h) POD, (i) CAT, and (j) SOD.
- Supplemental Table S1 A summary of the sample sizes, and ranges of publication year and effect size (lnRR) for each soil variable.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu L, Tang S, Kang J, Korpelainen H, Li C. 2023. Responses of dioecious Populus to heavy metals: a meta-analysis. Forestry Research 3:25 doi: 10.48130/FR-2023-0025 

Responses of dioecious Populus to heavy metals: a meta-analysis
- Received: 15 August 2023
- Accepted: 06 October 2023
- Published online: 24 October 2023
Abstract: A total of 946 sets of comparative data were collected from 20 publications and a meta-analysis performed to evaluate the responses of growth, photosynthetic capacity, oxidative stress and antioxidants in Populus females and males under exposure to heavy metals, like Cu, Mn, Zn, Pb and Cd. It was found that heavy metals have negative effects on Populus growth and photosynthetic capacity, as the average total biomass, leaf biomass, stem biomass, root biomass and height decreased by 29.78%, 33.41%, 27.22%, 35.30% and 34.83%, respectively. Furthermore, total chl, Pn, gs, E, Ci decreased by 23.30%, 26.03%, 40.49%, 23.76% and 18.24%, respectively. In addition, heavy metals increased oxidative stress and antioxidant enzyme activities: the average values of TBARS, H2O2, ${\text{O}^-_2} $ and MDA increased by 51.39%, 55.79%, 64.67% and 48.92%, respectively, and proline, APX, NPT, POD, CAT and SOD increased by 68.91%, 64.81%, 68.40%, 57.34%, 77.30% and 49.01%, respectively. However, there were sex-specific responses to heavy metals: females suffered more negative effects, as they had significantly greater decreases in root biomass, R/S ratio, height and total chl, and significantly smaller increases in NPT and POD activities than males. The present meta-analysis shows the responses of Populus females and males to heavy metals on a regional scale, which is crucial for understanding the patterns of sexual dimorphism and sex ratio biases in Populus with increasing heavy metal pollution in the future.
-
Key words:
- Heavy metals /
- Poplars /
- Sexual dimorphism /
- Reactive oxygen species