








-
Atractyiodes DC. (Compositae) consists of eight perennial herbaceous species that are native to the temperate regions of East Asia, primarily inhabiting forest understories, grassy slopes, and meadows. Atractylodes lancea (Thunb.) DC. (A. lancea), Atractylodes chinensis (DC.) Koidz. (A. chinensis), Atractylodes macrocephala Koidz. (A. macrocephala) and Atractylodes koreana (Nakai) Kitam. (A. koreana) thrive in the wild across Northern China, stretching from the central sections of the Yangtze River to the northeast areas leading into Inner Mongolia. Meanwhile, Atractylodes japonica Koidz. (A. japonica) is indigenous to Japan. Both A. japonica and A. koreana are found growing naturally in the Korean Peninsula[1]. These plants hold significant value as essential herbal remedies in traditional medicine practices of Chinese, Japanese, Korean, and Thai cultures[2], and are extensively utilized as important ingredients in healthcare products, and functional foods[3]. The plants are distinguished by their unsplit to pinnately divided leaves, embellished with small spines or teeth tipped with spines. They produce separate male and female flowers, displaying corollas in shades of white or purplish-red, as well as achenes that are obovoid to ovoid and flattened[4].
Medicinal plant affinities indicate that species within the same genus are closely related, sharing similar genetic material bases, as well as that species with close affinities have resembling external morphology, tissue structure, chemical composition, and pharmacological effects. Modern research has shown that A. chinensis has a closer relationship with A. coreana while being more distantly related to A. lancea. Similarly, A. macrocephala exhibits a closer kinship with A. japonica and a more distant connection to other Atractylodes species[5]. A. lancea, A. chinensis, and A. macrocephala are prominently featured in multiple editions of the Chinese Pharmacopoeia as medicinal herbs known as Cangzhu (A. lancea and A. chinensis) and Baizhu (A. macrocephala). These herbs boast a rich historical background in treating an array of illnesses[2]. However, in traditional Japanese medicine (Kampo), A. lancea, A. chinensis, or their interspecific hybrids are all referred to as Cangzhu ('Sojutsu' in Japanese), while both A. macrocephala and A. japonica are used as Baizhu ('Byakujutsu' in Japanese). Comparable medicinal properties and versatile applications underscore the intricate nature of the taxonomic connections associated with the source plant. In fact, the Atractylodes genus was first recognized as far back as 1838, but the interrelationships among and within the genus have consistently sparked debates. Deciphering the classification and evolutionary history of Atractylodes remains a formidable task owing to its tendency for hybridization and the overlap in both morphological and chemical characteristics[6]. Additionally, the scarcity of wild resources has prompted the widespread adoption of artificial cultivation, leading to the degradation of germplasm resources, highlighting the necessity for the sustainable preservation of this precious resource by identifying its quality formation mechanisms and evaluation methods. Herein, our focus extends beyond the previous review of the phytochemistry and pharmacology of Rhizoma atractylodis to encompass additional facets. There include the identification of germplasm resources, the biosynthesis pathway of active ingredients and product quality formation and control. We emphasize the utilization of emerging analytical and genetic advancements in technology and their remarkable implementations in assuring the quality of Rhizoma atractylodis. Moreover, we address the challenges inherent in Rhizoma atractylodis research and shed light on emerging trends within the field.
-
Species are constantly mutating to produce new traits, and whether the newly produced traits can be passed on from generation to generation depends on the ecological environment in which the species is located[7]. A stable germplasm is the result of natural selection under the long-term action of the ecological environment, and the germplasm of wild herbs corresponds to the environment. Atractylodes are widely distributed in China. A. lancea is mainly distributed in humid and warm regions, while A. chinensis is mainly distributed in dry and cold regions, with large environmental differences; A. coreana is mainly distributed in the northeastern region, with obvious cross-relationships with A. chinensis, with small environmental differences; A. macrocephala and A. japonica have no cross-relationships, and are located in different environments. Modern research has shown that A. chinensis is more distantly related to A. lancea and closer to A. coreana, and some studies have even suggested that A. coreana is a variant of A. chinensis adapted to dry hillside environments[8]. There are many debates about the classification of Atractylodes and their species.
The morphological diversity within this genus is quite extensive, and traditional methods of identification make it challenging to establish relationships. A. macrocephala has branched stems from the base, whereas A. carlinoides has pinnatifid, rosulate basal leaves, which are easy to distinguish from other species. And A. carlinoides and A. macrocephala have been identified as the earliest diverging taxa. However, differentiating between the other species can be challenging from a morphological standpoint, especially when the plants are in their early stages with unbranched stems and undivided leaves[9]. A. lancea, which is known as Maocangzhu in traditional Chinese medicine (TCM), primarily thrives across the provinces of Jiangsu, Hubei, Anhui, and Henan in China. The Maoshan region in Jiangsu is recognized as a significant geoherbalism medicinal herb area. Notably, the cut section of A. lancea exhibits vermilion oil spots and develops a white frost precipitate upon prolonged placement, as observed from its external appearance. Nevertheless, a reduced presence of this frosting characteristic is noticeable in the cross-section of A. lancea from the Maoshan region. Consequently, it becomes crucial and imperative to differentiate the variations in the chemical composition of Atractylodes. The composition of compounds in Atractylodes demonstrates significant variability based on their natural habitat. The differences in the geographical distribution and the content of active ingredients of the Atractylodes are shown in Table 1. Over the past few decades, numerous approaches have been employed to explore the origin, genetic diversity, and evolutionary connections of Atractylodes, including morphological characteristics, chemometrics, plastid DNA sequences, and nuclear DNA datasets.
Table 1. Distribution, main active ingredients, and content of genus Atractylodes in China.
Crude drug Species Geographic
distributionVolatile oil
content (%)Atractylodin
(%)β-eudesmol
(%)Atractylone
(%)Atractylenolide
I (%)Atractylenolide
II (%)Atractylenolide
III (%)Atractylodis
RhizomaA. lancea Mainly in Jiangsu, Zhejiang, Hubei, Sichuan and other places south of the Qinling Mountains. 5-9 0.31 4.4 0.39 0.016 0.013 0.007 A. chinensis Mainly in northeast, north and northwest areas north of the Qinling Mountains. 3-5 0.34 0.25 0.35 0.034 0.021 0.016 Atractylodis
Macrocephalae
RhizomaA. macrocephala It is distributed in a small amount in Jiangxi, Zhejiang, Anhui and other regions. 1-3 0.003 / 5.89 0.025 0.018 0.038 A. japonica Northeast China and Inner Mongolia, Hebei and other regions. 1-3 0.14 0.025 1.29 0.033 0.030 0.049 A. coreana A few areas in Qian Shan, Jilin and Inner Mongolia. 2-3 0.28 / 0.08 / 0.020 / Physical and chemical identification
-
The morphological variation of Atractylodes is relatively large, and it is difficult to identify them by traditional methods. At present, the clinical use of Cangzhu is mostly based on A. lancea, and it is recognized in the industry that A. lancea is of good quality, so it can be seen that there are differences in quality between A. lancea and A. chinensis. As they are plants of the same family, their microscopic characteristics and chemical composition are similar, and it is difficult to identify them, leading to serious confusion in the market for A. lancea and A. chinensis. Therefore, we can rely on the development of modern chemometrics or sequencing technology to understand the evolution of the chemical clustering or genome structure of Atractylodes and analyze the phylogenetic relationship.
The theory of 'quality evaluation through morphological identification' in Chinese medicine is to judge the authenticity of Chinese medicines based on the characteristics of their appearance (shape, color, smell, and taste), to clarify the essence of their quality, which is the essence and characteristic of the identification of Chinese medicines in the past[10]. The taste of A. lancea and A. chinensis are both pungent and bitter, and the standard is 'the strongest taste is the best', which shows that taste is an important index for the quality evaluation of A. lancea and A. chinensis. An electronic tongue technique was used to determine the quantitative values of the taste of 23 lots of A. lancea, and chemometric methods to construct a qualitative identification model to identify A. lancea and A. chinensis and their origins, and indicate that the content of volatile oil of Cangzhu herbs was correlated with the flavor of the herbs[11]. Another quality evaluation standard of A. lancea is 'firm in quality, with many cinnabar dots on the cross-section and a strong aroma', which is an accepted industry standard for identifying the quality grade of A. lancea in the Chinese medicine circulation field, and has been used for thousands of years, in which cinnabar dots are brownish red or orange oil dots scattered on the yellowish white or greyish white cross-section of A. lancea, and the number of cinnabar dots directly affects the color of the cross-section and powder of the herb. Color difference meter was used to determine the trichromatic spatial values of the powder of A. lancea and A. chinensis, combined with HPLC to determine the contents of four active ingredients in atractylenolide II, β-eudesmol, atractylodin, and atractylone, and analyzed the correlation between the contents of the active ingredients and the chromatic values, and constructed a qualitative identification model based on the trichromatic spatial values of A. lancea and A. chinensis, to provide a reference for the quality control of Rhizoma atractylodis[12]. In addition, the color of the secretory cavity at A. lancea differs greatly among geographic origins. In a study conducted by Jiang et al.[13], laser-capture microdissection-based metabolomics was employed to analyze three natural accessions of A. lancea with distinct color patterns. Through multivariate statistics analysis across various secretory cavity types, three polyacetylenes were found to be significantly associated with the red secretory cavity of A. lancea. Mass spectrometry imaging further confirmed that these polyacetylenes are likely the causal compounds responsible for the cinnabar-like color of the secretory cavities in A. lancea (Fig. 1). These findings will provide valuable insights for understanding the biosynthetic pathways of polyacetylenes in A. lancea. Furthermore, the characteristics of frost formations on A. lancea and their surfaces were analyzed using various microscopy techniques, including light microscopy, polarizing microscopy, stereomicroscopy, and metalloscopy. A. lancea and its frost formations were also subjected to GC-MS analysis, revealing that the main components responsible for the frost formation on the plant's surface were hinesol and β-endemol. These results highlight the scientific significance of considering the precipitated frosts of medicinal materials as an indicator of quality and efficacy[14].
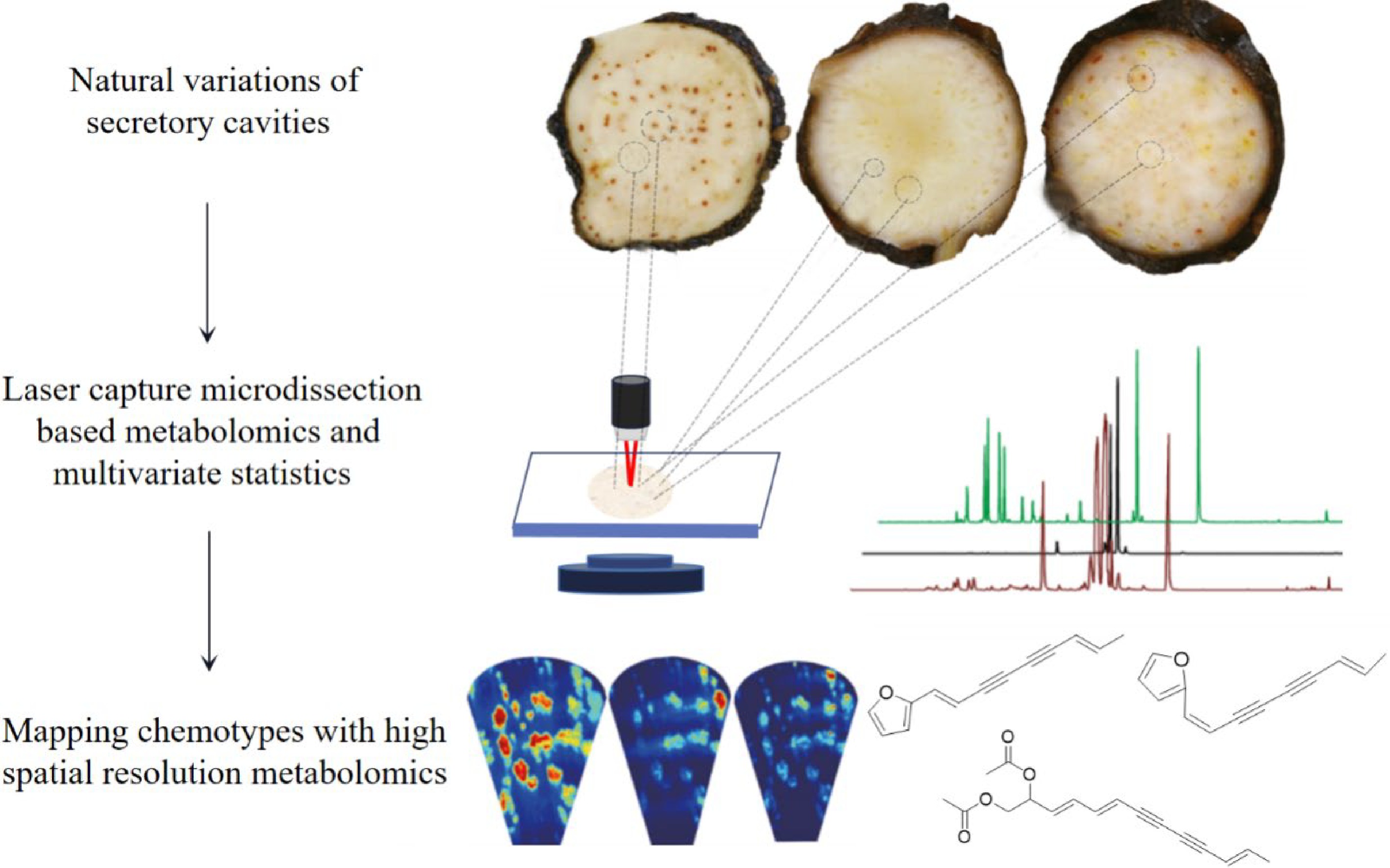
Figure 1.
Combination of natural variations and spatial metabolomics to reveal key metabolite markers associated with important pharmaceutical properties[18].
Chromatographic fingerprinting (HPLC or GC), can elucidate the essential chemical characteristics of the species[15]. Nevertheless, it's worth emphasizing that the preliminary purification process for chromatographic fingerprinting may not capture all the metabolites found in the raw medicinal plant. To overcome these limitations, NMR spectroscopy offers a promising solution. In recent years, the application of NMR-based metabolomics technology has demonstrated its efficacy in the examination of herbal remedies[16, 17]. A recent study performed 1H NMR metabolic profiling of Atractylodes species, revealing that the rhizome extracts' metabolic profiles, analyzed through principal component analysis (PCA), effectively differentiated five Atractylodes species. This differentiation was confirmed by comparing the results with DNA barcoding authentication.
Comparative analysis of molecular biology in Atractylodes
-
The chloroplast genome, being a complete DNA sequence distinct from the nuclear genome, exhibits a high degree of conservation and stability during the evolutionary process. It has found extensive utility in recent years for plant species identification and systematic evolutionary research. Through comparative analysis of the chloroplast genome, notable variations in both sequence and structural elements can be identified within and between plant species[19]. The chloroplast genome has a conservative tetrad structure: a large single copy (LSC) region, a small single copy (SSC) region, and two reverse repeat (IR) regions. The genome size ranges from 120 to 160 kb[20]. Chloroplast genome analysis is usually carried out from the aspects of sequencing to determine molecular structure, annotation to obtain coding genes, phylogeny, evolution, and comparison. At present, many articles on chloroplast genomes of medicinal plants have been published successively, including Salvia miltiorhoza[21], S. yangii[22], S. przewalskii, S. bulletyana and S. japonica[23], chloroplast genomes of Salvia species have been published successively. In June 2021, the complete chloroplast genome of four cross-generic (Cadaba and Maeerua) medicinal plants of Caparaceae (i.e., C. farinosa, C. glutenalosa, M. crassifolia, and M. oblongifolia) was published[24]. In addition, the complete chloroplast genomes of medicinal plants such as Sophora japonica[25], Artemisia annua[26], Peony[27], and Aristolochia[28] have also been published.
Wang et al.[28] published the chloroplast data of six species of Atractylodes, with a total size between 152,294 bp (A. carlinoides) and 153,261 bp (A. macrocephala). The chloroplast genome of Atractylodes has a typical tetragonal structure, including a pair of IR regions (25,132−25,153 bp), LSC regions (83,359−84,281 bp), and SSC regions (18,634−18,707 bp), and there is almost no difference between the chloroplast genomes of the six Atractylodes. Phylogenetic analysis showed that Atractylodes species independently formed a monophyletic group and formed a sister relationship with other species (Fig. 2)[28], further reflecting a monophyly genetic relationship among the six Atractylodes species. The study of the chloroplast genome of Atractylodes provides valuable information for variety identification and high-efficacy plant breeding.
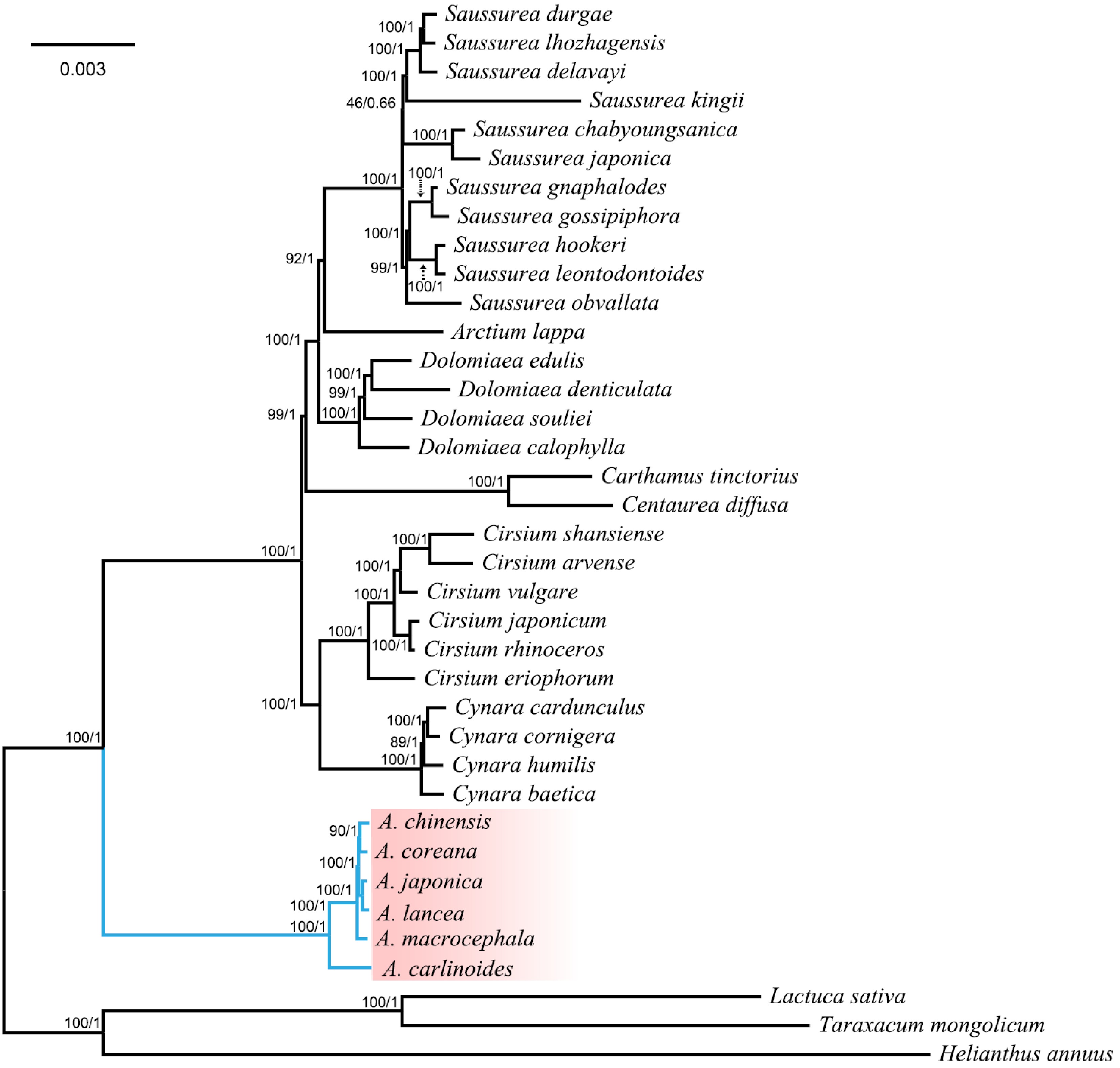
Figure 2.
Phylogenetic tree constructed using the maximum likelihood (ML) and Bayesian inference methods based on the whole chloroplast genomes from 37 different species. The numbers above the branches represent the ML bootstrap values/BI posterior probabilities[28].
To accurately differentiate A. macrocephala, A. lancea, and their hybrids, a molecular approach combining multiplex ligation-dependent probe amplification (MLPA) and Real-time Quantitative PCR (qPCR) was developed. This method utilizes highly species-specific MLPA probes designed specifically for the ITS2 region within the nuclear genome, ensuring no cross-reaction among the species. Within 3 h, the melting curve analysis of the amplified products allows for the rapid and precise identification of the three species[29]. The chloroplast whole genome sequences of four A. chinensis from different origins, screened specific DNA barcodes (ccsA and trnC-GCA_petN) to identify the germplasm resources of A. chinensis and analyzed the genetic diversity of the populations, showing that A. chinensis has high genetic diversity at the species level, small genetic differences among haplotypes, and forms two distinct branches from other taxa of the same genus except A. macrocephala[30]. To assess the genetic diversity of A. chinensis in various geographical regions of northern China, Simple Sequence Repeats (SSR) were employed. The genetic relatedness among 19 populations of A. chinensis was determined using a neighbor-joining tree, which classified them into three distinct clusters. Additionally, DNA fingerprinting was conducted using ten SSR markers. The results indicated that genetic diversity was not strictly correlated with geographic origin. Populations from different provinces were found to be grouped within the same cluster, suggesting that other factors beyond geographic location contribute to the genetic variation in A. chinensis populations[31].
It is crucial to acknowledge that phylogenetic ambiguity might arise due to hybridization and introgression among species within the A. lancea complex[32,33]. Furthermore, plastid genes can manifest distinct phylogenetic relationships compared to nuclear genes owing to plastid capture. To attain more comprehensive phylogenetic insights, the utilization of both nuclear and plastid datasets is imperative[34]. This entails extensive sampling of all taxa and the careful selection of more suitable markers and methodologies.
-
The active ingredients in medicinal plants are of great interest for their important roles in disease treatment, such as artemisinin for anti-malaria, tanshinone and tannic acid for anti-cardiovascular diseases, paclitaxel for anti-cancer, and ginsenosides for immunomodulation[35]. Secondary metabolites are typically the active ingredients found in medicinal plants. In the case of Rhizoma atractylodis, sesquiterpenoids have been identified as the key bioactive compounds. These sesquiterpenoids exhibit a wide range of beneficial effects, including anti-tumorigenic, anti-inflammatory, neuroprotective, anti-diabetic, anti-hyperlipidemic, anti-microbial properties, and are effective against gastrointestinal diseases[2].
Concerning biosynthesis in Rhizoma atractylodis, sesquiterpene volatile oils have attracted the most attention. Rhizoma atractylodis is rich in multiple sesquiterpenes and according to the structure can be divided into three cyclized structures: monocyclic sesquiterpenes, bicyclic sesquiterpenes, and tricyclic sesquiterpenes[36]. Cyclization in the biosynthesis of monocyclic sesquiterpenes involves cyclization through either the farnesyl cation or its isomer, while bicyclic sesquiterpenes are formed through germacrene modifiers. Tricyclic sesquiterpenes are produced via the double cyclization of the guaianyl cation. The excavation of their synthesis pathway genes is a prerequisite for the in-depth understanding and regulation of the biosynthesis of active ingredients in A. lancea. Sesquiterpenoids are composed of three molecules of isoprene polymerized into a skeleton containing 15 carbon atoms and have a complex biosynthetic pathway. In general, the biosynthesis of sesquiterpene volatile oils is mainly through the mevalonate (MVA) pathway in the cytoplasm and mitochondria of the plant, and to a lesser extent through the methylerythritol-4-phosphate (MEP) pathway in the plastid, as shown in Fig. 3[37]. In the MVA pathway, a variety of terpenoids are produced by the synergistic action of 3-hydroxy-3-methylglutarate monoacyl coenzyme A reductase (HMGR), farnesyl pyrophosphate synthase (FPPS) and different terpene synthases (TPS)[38,39]. The 1-deoxy-D-xylose-5-phosphate synthetase (DXS) is the first key rate-limiting enzyme in the MEP pathway. The functions of some sesquiterpene biosynthesis-related genes have been studied, and HMGR and DXR have been reported in the upstream MVA and MEP pathways of A. lancea sesquiterpenes.
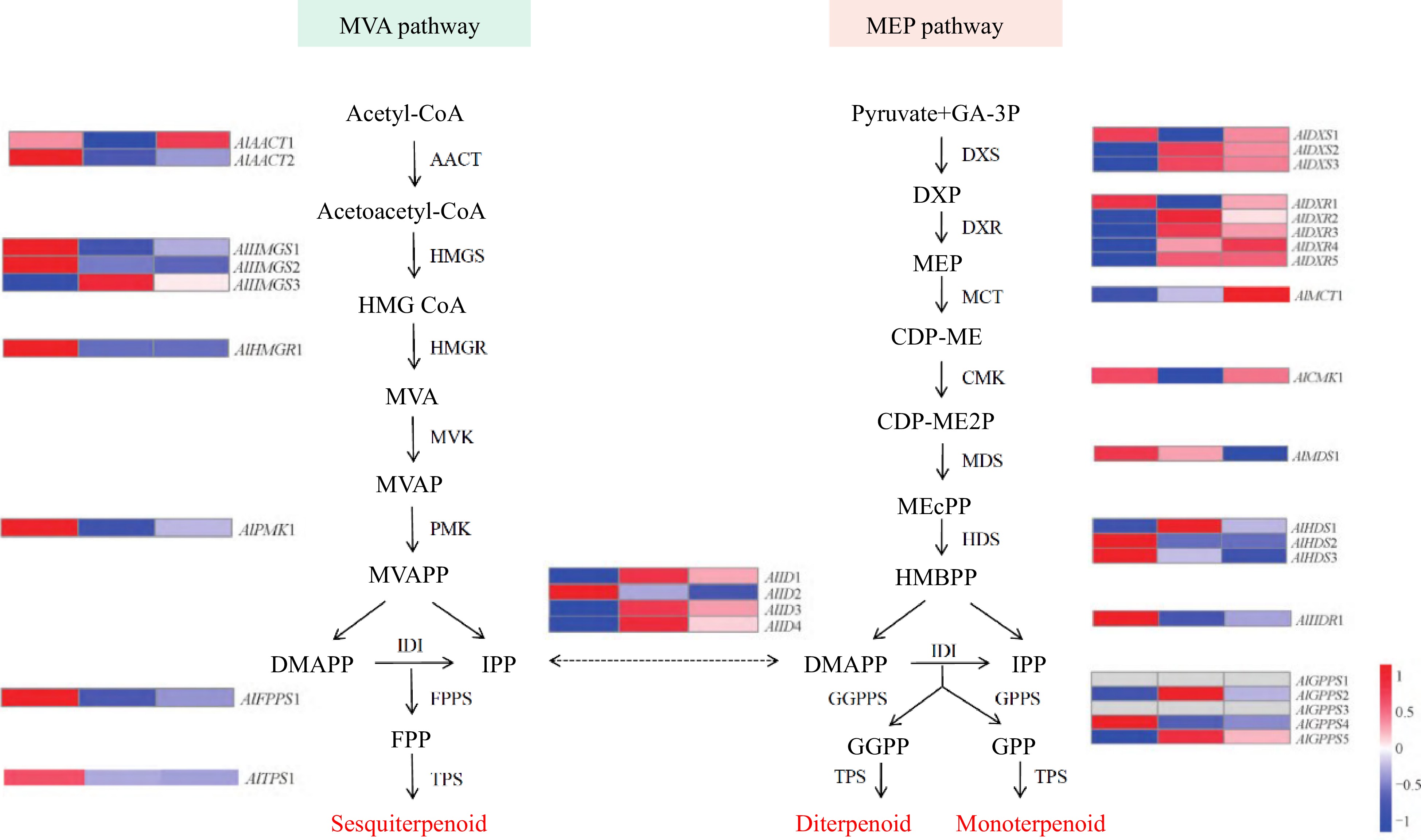
Figure 3.
Biological pathway for sesquiterpenoid synthesis in A. lancea. AACT: Acetyl-CoA-acetyltransferase; HMGS: Hydroxy methyl glutaryl-CoA synthase; HMGR: 3-hydroxy-3-methylglutaryl-coenzyme A reductase; MVK: Mevalonate kinase; PMK: Phosphomevalonate kinase; DXS: 1-deoxy-D-xylulose-5-phosphate synthase; DXR: 1-deoxy-Dxylulose-5-phosphate reductoisomerase; MCT: 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase; MDS: 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase; HDS: 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase; IDI: Isopentenyl-diphosphate delta-isomerase; GPPS: Geranyl diphosphate synthase; FPPS: Farnesyl diphosphate synthase; GGPPS: Geranylgeranyl pyrophosphate synthase; TPS: Terpenoid synthase.
Transcriptome sequencing, specifically RNA-seq, employs high-throughput sequencing technology to investigate the crucial genes involved in the biosynthesis of medicinal ingredients and explore their respective biosynthetic pathways and regulatory mechanisms. This approach aims to enhance the content of active ingredients in medicinal plants. Over time, transcriptome sequencing and differential expression analysis have emerged as effective methods for exploring functional genes since the advent of second-generation sequencing techniques[40]. In a study conducted by our group, the whole genome of A. lancea was sequenced, providing a valuable resource for further research. Cao et al.[41] utilized the A. lancea genome as a reference to sequence the transcriptome of A. lancea roots at different developmental stages. The dynamics of transcriptional regulation during the maturation of A. lancea were analyzed using the weighted gene coexpression network analysis (WGCNA) method. These findings contribute to the exploration of key enzyme genes involved in the biosynthesis of sesquiterpenoids in A. lancea. Moreover, they provide a foundation for the subsequent analysis and regulation of the synthesis and accumulation of active ingredients in the plant.
The transcriptome analysis of various tissues of the non-model plant A. lancea was investigated using short-read sequencing technology by Ahmed et al.[38]. The researchers identified 62,352 high-quality unigenes with an average sequence length of 913 bp in the A. lancea transcripts. They were able to identify enzymes involved in terpenoid biosynthesis as well as five known sesquiterpenoids through the MVA pathway and plastidial MEP pathways. This transcriptomic resource of A. lancea provides valuable insights into the classes of terpenoids and other chemical compounds with medicinal and economic importance. Furthermore, the study classified A. lancea into two chemotypes: the Dabieshan and Maoshan chemotypes, although this classification is unreasonable. GC-MS analysis revealed higher hinesol and β-eudesmol contents in the Dabieshan chemotype compared to the Maoshan chemotype. To investigate the molecular mechanisms of sesquiterpenoid biosynthesis in A. lancea, a combination of next-generation sequencing (NGS) and single-molecule real-time (SMRT) sequencing technologies was employed. The researchers identified 42 differentially expressed genes (DEGs) involved in terpenoid biosynthesis in the two chemotype groups and identified nine full-length TPS genes. Subcellular localization analysis revealed that the AITPS1 and AITPS2 proteins were localized in the nucleus and endoplasmic reticulum, respectively. AITPS1 catalyzed the biosynthesis of elemol, while AITPS2 exhibited β-farnesene synthase activity. This study provides valuable information regarding the differences in terpenoid accumulation in the two chemotypes of A. lancea and establishes a foundation for further elucidating the molecular mechanisms of sesquiterpenoid biosynthesis[38]. These findings on biosynthetic pathways contribute to the understanding of biosynthesis and targeted enrichment of different types of sesquiterpenes. They also provide essential information for improving the yield of valuable and active sesquiterpenes through plant metabolic engineering.
Quality formation of Rhizoma atractylodis
-
The accumulation of active compounds in medicinal plants is influenced by a synergistic interplay between genetic and environmental factors[39,40]. Genetic factors play a pivotal role in secondary metabolism, and their comprehensive exploration is essential for understanding the molecular mechanisms underpinning medicinal substance biosynthesis[41]. This understanding not only facilitates the enhancement of medicinal herb quality but also addresses the scarcity of medicinal plant resources. Concurrently, environmental factors exert a significant influence on the synthesis and accumulation of volatile oils in Rhizoma atractylodis, assuming a crucial role in shaping the development of ecologically dominant authentic herbs.
Genetic factors
-
The wide range of volatile oil components in A. lancea signifies that its synthesis is intricately governed by multiple genetic factors. Currently, most genetic studies on the synthesis and accumulation of volatile oils in A. lancea have concentrated on the overexpression of individual or multiple key enzyme genes and the regulation of transcription factors within the synthesis pathway[42].
Overexpression of HMGR, a key enzyme, has been shown to significantly increase sesquiterpene content in various medicinal plants, such as Artemisia annua and Arabidopsis thaliana[43]. The overexpression of the HMGR gene could be promoted by modifying the genetic characteristics of A. lancea with appropriate amounts of calcium chloride and improving the accumulation of volatile oil components in A. lancea[44]. The accumulation of H2O2 signaling molecules also significantly increased the activity of HMGR and sesquiterpene β-eudesmol content in A. lancea, and there was a high positive correlation between the two. These results indicated that the activation of HMGR promoted the biosynthesis of β-eudesmol[45]. In addition, Jiang et al.[46] found that the content of various sesquiterpenoids in A. lancea at different times was significantly and positively correlated with the expression of the key enzyme gene FPPS, and found that overexpression of FPPS could increase the yield of sesquiterpene volatile oil in A. lancea, proving that FPPS is an important factor affecting the biosynthetic pathway of sesquiterpenoids. The full-length cDNA sequence of the DXS gene from Artemisia annua by gene cloning, and the amino acid sequence comparison revealed that its similarity with the DXS gene from Artemisia annua in the same family was as high as 93%[47]. Chen et al.[48] cloned and analyzed two DXR genes from A. lancea and found that they were highly similar to those of wild chrysanthemum and Artemisia annua in the Asteraceae family and were highly conserved, providing theoretical guidance to further reveal the regulatory mechanism of the key enzyme gene DXR in the sesquiterpenoid volatile oil synthesis pathway of A. lancea.
The biosynthetic pathway of polyacetylene volatile oils in A. lancea is less studied, and several intermediates are not well defined[49]. Among them, acetyl coenzyme A carboxylase (ACC) and acetyl coenzyme A synthase (ACS) are the key and rate-limiting enzymes in the synthesis pathway. The expression of ACC is easily induced by environmental factors, such as salicylic acid and fungi, which can enhance or reduce its expression[50]. Tao et al.[51] found that a novel plant signaling molecule, nitric oxide, could mediate the activation of the key enzyme ACC by endophytic fungal elicitors through salicylic acid signaling channels to promote secondary metabolism in A. cepacia cells and enhance the production of cynarin. Since the key enzyme gene ACC promotes the accumulation of secondary metabolites in plants[52], the synthesis pathways of polyacetylene and fatty acid components are similar and both key and rate-limiting enzymes contain ACC, it is hypothesized that genetic engineering of A. lancea can improve the accumulation of polyacetylene volatile oil and hence the quality. There are few reports on the factors affecting ACS.
The co-regulation of multiple genes can simultaneously break multiple restrictions in metabolic pathways, often improving metabolic efficiency more than individual genes. In recent years, many researchers have begun to explore the use of multiple gene regulation to enhance the content of active ingredients in medicinal plants. The co-regulation of HMGR and FPPS, HMGR and DXR, FPPS, cytochrome P450s (CYP450s), and TPS has been demonstrated in the synthesis of volatile oil in A. lancea. The expression of HMGR and FPPS genes in the rhizome of A. lancea was significantly and positively correlated with the contents of atractylenol, β-eudesmol, and atractylenol, but the sesquiterpene atractylenol and β-eudesmol were significantly higher than the polyethylene-based atractylenol, which was consistent with the localization of HMGR and FPPS genes in the sesquiterpene pathway. Similarly, the medicinal plant Artemisia annua from the same family overexpression of the HMGR and FPPS genes, both individually and simultaneously, significantly increased the sesquiterpene component artemisinin content, and that co-expression increased the yield by 80%[53]. Zhou et al.[54] found that the signal molecules gibberellin and abscisic acid directly enhanced the expression of HMGR and DXR, the key enzymes encoding sesquiterpene volatile oil synthesis in the rhizome of A. lancea, resulting in increased enzyme activity and promoting the synthesis and accumulation of sesquiterpene volatile oil. The downstream genes FPPS and TPS were less studied than the upstream genes in the sesquiterpene volatile oil synthesis pathway of A. lancea. A large number of unigenes functions annotated as CYP450s in the transcriptome data of A. lancea suggested that the synthesis of sesquiterpene volatile oils is usually accomplished through the collaboration of the cytochrome P450 gene family and TPS gene family. It has also been demonstrated that cytochrome P450 monooxygenase (CYP71AV1) and cytochrome P450 oxidoreductase (CPR) genes jointly regulate the biosynthesis of sesquiterpenoids[55], and simultaneous overexpression of endogenous CYP71AV1 and CPR genes in Artemisia annua resulted in a significant increase in the production of the sesquiterpenoid artemisinin[56].
Transcription factors also hold a significant role in plant secondary metabolism. They do so by binding to cis-acting elements within the promoter regions of relevant genes, thereby either activating or suppressing the expression of those genes. Currently, the most studied transcription factors in A. lancea are AP2-ERF, WRKY, bHLH, and other families. It has been found that the AP2-ERF family of transcription factors is specific to the leaves of A. lancea and plays an important role in leaf growth and development, and it can be speculated that the regulation of AP2-ERF family transcription factors can affect the synthesis and accumulation of metabolites in A. lancea[57]. This is corroborated by the contribution of the AP2-ERF family transcription factor TAR1 to the synthesis and accumulation of the sesquiterpene artemisinin in the medicinal plant Artemisia annua, a member of the same family[58]. The WRKY family transcription factors may act as skeletal sesquiterpenes in the formation of diverse sesquiterpenes in Artemisia annua, leading to an early shoot and a significant increase in sesquiterpene content[59], which is consistent with the role of WRKY-type transcription factors in Artemisia annua[60]. Among the bHLH family transcription factors, the most thoroughly studied and isolated is the MYC family transcription factor, which is involved in the regulation of the jasmonic acid signaling pathway and also responds to gibberellin signals, regulating the formation of sesquiterpene volatile oils in response to external stimuli[53]. Huang et al.[61] found that the AP2-EREBP family transcription factors were present in both leaves and rhizomes, whereas the WRKY family was mostly from leaves and the bHLH family was mostly in rhizomes, as shown by a comparison of transcription factors in leaves and rhizomes. It is evident that the growth and secondary metabolism of A. lancea are regulated by several transcription factor families, and further studies are needed to identify and elucidate the underlying genetic mechanisms.
miRNAs, known as plant microRNAs, are short RNA molecules that do not code for proteins. They play a role in RNA silencing and regulate various target genes involved in plant development, metabolism, and responses to different environmental conditions. Their impact on terpene synthesis primarily occurs through the regulation of rate-limiting enzyme genes in the terpene synthesis pathways. For instance, in Artemisia annua, the target genes of miR414, including HMGR, P450, FPS, and amorpha-4,11-diene synthase, are mainly involved in terpene synthesis[62]. Similarly, in A.lancea, four differentially expressed miRNAs (MIR159, MIR172, MIR393, MIR399), have been found to affect the biosynthesis of terpenoid backbones[63]. Although it has not been confirmed whether these four miRNAs directly influence terpenoid synthesis, further investigation is necessary to determine if they contribute to the compositional differences in volatile oils observed in A. lancea from different origins.
In conclusion, if the genetic characteristics of volatile oil polygenic regulation can be reasonably exploited, the quality of A. lancea herbs can be efficiently improved through genetic engineering and other biological means, thus effectively alleviating the increasingly severe market demand for medicinal plants. However, the secondary metabolic pathways remain intricate, with many pathways not yet comprehensively elucidated and limited to the examination of a select few functional genes. The advancement of molecular biology is poised to facilitate deeper investigations into secondary metabolic pathways and their regulatory mechanisms. Notably, in recent years, the integration of genomics, transcriptomics, protein genomics, and bioinformatics has significantly streamlined the study of secondary metabolic pathways in medicinal plants, ushering in a new era of convenience and effectiveness in this field of research.
Environmental factors
-
Ecological suitability analysis shows that environmental factors such as temperature, light, water, fertilizer, and biological factors can affect the synthesis and accumulation of active ingredients in Rhizoma atractylodis to varying degrees.
Temperature is one of the most important factors influencing the secondary metabolic activities of plants and can cause changes in the content of various components. High temperatures affect the fluidity of plant cell membranes and enzyme activities, disrupting the dynamic balance between the production and removal of reactive oxygen species and other harmful substances in the plant, inhibiting photosynthesis and secondary metabolism, and even preventing normal growth and development[64]. Low temperature also has a significant impact on plant metabolism and development and is one of the most important environmental factors affecting the geographical distribution of plants. It was found that climatic zones suitable for the growth and development of A. lancea are not necessarily suitable for the accumulation of volatile oil, and that appropriate high temperatures are beneficial to the accumulation of volatile oil, but not to normal growth and development[65]. The moderate temperature was more favorable to the accumulation of total biomass, the high temperature was more favorable to the accumulation of volatile oil[66]. Zhang et al.[67] found that the suitable monthly average temperature for A. lancea was 32 °C, while the highest temperature in summer reached 38 °C in open field cultivation, resulting in a lower volatile oil composition in open field cultivation than in forest cultivation, which indicates that forest cultivation has some regulatory effect on extreme high-temperature environments.
Light is the only source of photosynthetic energy for plants, and light intensity, light duration, irradiation direction, and spectral combination are important factors influencing organ differentiation, morphological establishment, yield improvement, and quality. The biomass and volatile oil content of A. lancea at 80% light intensity were significantly higher. At 7% and 20% light intensity, which were below the minimum light compensation point for photosynthesis, the plants could not carry out normal physiological and biochemical reactions, resulting in the lowest biomass and volatile oil content[68]. This indicates that full light intensity is not the best condition for the increase of biomass and accumulation of secondary metabolites in A. lancea, and that either too high or too low light intensity will cause the accumulation of reactive oxygen species in the plant, and may even cause membrane lipid peroxidation resulting in membrane incompleteness and chloroplast malformation, thus reducing the efficiency of photosynthesis. In a study by Guo et al.[69], it was discovered that moderate shading at 80% intensity creates an optimal shade condition that supports consistent and sustainable production of premium A. lancea. Under these shade conditions, there is an effective enhancement in the accumulation of overall volatile oils, primarily achieved by regulating the biosynthesis of sesquiterpenoids like hinesol and β-eudesmol. This regulation is achieved through the integration and coordination of photosynthesis, phytohormones, and the promotion of DXR, and FPPS gene expression. Light duration also influenced the accumulation of volatile oil in A. lancea, with volatile oil content at different photoperiods of 10 h < 16 h < 12 h < 14 h. The volatile oil content at 14 h photoperiod increased by 19, 70% compared with that of the control group (12 h)[70]. Different light qualities caused several physiological responses associated with stress resistance in A. lancea, which in turn affected volatile oil accumulation[71]. In particular, at 10% red light and 90% blue light, the higher content of cystoids in the leaves of A. lancea is most suitable for the growth and development of A. lancea, and the accumulation of reactive oxygen species in A. lancea is reduced at this time, thus resulting in significantly higher biomass and volatile oil content than the other spectral combinations. The gene expression of FPPS and TPS in the rhizome of A. lancea also showed a certain correlation with the different types of light quality, with the gene expression in the blue light group being lower and significantly higher than that in the control group when the ratio of red and blue light was 9:1[72]. Different light directions can lead to differences in the distribution of plant growth factors that cause phototropic movements, which also have important implications for yield, but there are few studies on the effects of light directions on plant secondary metabolism.
Water is most important as a nutrient transport carrier and dissolution medium. Too much or too little water can put the plant in a flood or drought adversity, causing secondary metabolic processes in the plant to be affected, which in turn affects the accumulation of medicinal components. Under drought stress conditions, the assimilation process in plants was disrupted due to the negative impact on photosynthesis. The tolerance to stress was compromised as the antioxidative enzyme system was inhibited. Additionally, the synthesis of bioactive components was impeded by the downregulation of gene expression related to sesquiterpene synthase[73]. Guo et al.[69] found that severe drought reduced the accumulation of volatile oil components in the nutritional, late fruiting, and whole growth periods, but had no significant effect on the reproductive growth period. Volatile oil components mainly decreased in the nutritional and reproductive growth periods during mild drought, while some volatile oil components such as atractylenol and β-eudesmol increased in the late fruiting and whole growth periods, but total volatile oil decreased. When A. chinensis was subjected to severe drought stress, the accumulation of secondary metabolites was greatly inhibited, although the body could activate its defense system by regulating the protective enzymes to resist the external environment against itself[74]. Water stress also regulated the expression of sesquiterpenoid-related genes, increasing the expression of HMGR and OvTPS2 genes and a decrease in the expression of FPPS and OvTPS6 genes, indicating that water is a limiting factor for secondary metabolic processes in plants[75].
In summary, genuine medicinal herbs of A. lancea do not grow in the most suitable environment, and the impact of environmental stress on the secondary metabolites of genuine medicinal herbs of A. lancea is of great significance. It is advantageous for the accumulation of total biomass of A. lancea to be grown at room temperature, while higher temperatures are more conducive to the accumulation of volatile oils. Therefore, the optimal environmental temperature for the accumulation of overall biomass in A. lancea is at room temperature, with occasional periods of higher temperatures. In situations where temperature cannot be controlled, forest cultivation offers more temperature stability. Under 80% light intensity conditions, both the biomass and volatile oil content of A. lancea significantly increase. In geographical locations that can provide 14 h of daylight and optimize light intensity for A. lancea, this may be a crucial factor in supporting the continuous production of high-quality A. lancea. Meanwhile, mild, occasional drought conditions can also contribute to the accumulation of certain effective secondary metabolites in A. lancea.
The application of fertilizers significantly affects the growth and development of herbs and the accumulation of metabolites. A reasonable application can not only ensure the supply of soil nutrients but also improve the soil, which can significantly improve the yield and quality of herbs per unit area. Inorganic fertilizers that are widely used at present mainly include nitrogen, phosphorus, and potassium. Hu et al.[76] found that 15 g/m2 was the most suitable amount of phosphorus for A. lancea, while low or high phosphorus would also affect photosynthesis, causing an imbalance in the metabolism of reactive oxygen species in A. lancea leaves and disrupting the osmotic pressure inside and outside the cells. An insufficient supply of potassium affected the normal metabolism of proteins in A. lancea plants and prevented photosynthesis in the above-ground parts of the plant, resulting in a reduction in the content of β-eudesmol and hinesol[77]. In agricultural production, fertilizer mixes are often more effective than individual applications. During the rapid nutritional growth period, a follow-up application of nitrogen, phosphorus, and potassium fertilizers can improve the synthesis, transport, and accumulation efficiency of the volatile oil components in the rhizomes of A. lancea, and the selection of suitable fertilizer ratios and application periods is important for the improvement of the quality and yield of A. lancea[78].
In nature, organisms are intricately related to each other and interact with each other. Among them, the endophytic bacteria present in the plant are the most closely linked to the plant, and their interaction has an important impact on the changes in physiological indicators, biomass, and the accumulation of active ingredients. The endophytic bacterium A. fortuitum could increase the volatile oil content of secondary metabolites in A. lancea, selectively reduce β-eudesmol with volatile oil as the only carbon source, and regularly increase the content of atractylenol and atractylenone, thus improving the medicinal quality[79]. Li[80] screened the endophytic bacterium and indicated that the endophytic bacterium was related to the formation of atractylenol. Ren[81] found that endophytic bacteria were the main component of volatile oil accumulation in A. lancea by studying the accumulation of metabolites of the herb. In addition, endophytic bacteria can increase the activity of various enzymes, improve the photosynthetic rate, and promote growth and development, as well as induce salicylic acid, jasmonic acid, nitric oxide, and hydrogen peroxide to promote the accumulation of secondary metabolites in A. lancea. In contrast, A. lancea could directly produce reactive oxygen species after inoculation with the plant endophyte Pseudomonas fluorescens, causing oxygen bursts that oxidized anaerobic sesquiterpenes to oxygenated sesquiterpenes and promoted the accumulation of non-oxygenated terpenoids in A. lancea, thereby increasing the diversity of sesquiterpenoids[82]. The accumulation of metabolites was also affected by the disturbance of endophytic bacteria by environmental changes, and it was found that climatic changes in different seasons caused a certain degree of succession in the endophytic community of A. lancea, which in turn induced the accumulation of volatile oil[83]. The mycorrhizal fungi (AM) fungus can promote the rapid repair of cell membranes and normal growth of A. lancea plants when they are subjected to high temperatures and drought stress[84]. Zhang et al.[85] found that AM fungal treatment could increase the number of soil microorganisms in the inter-rhizosphere of A. lancea, improve the functional diversity, metabolic activity, and utilization of single carbon substrate, improve the inter-rhizosphere soil environment, and then improve the yield and quality. AM fungi could also improve the survival rate, growth, and development, increase the activities of disease-resistant and antioxidant-related enzymes in the leaves, and accumulate the secondary metabolites hinesol, β-eudesmol, and atractylodin, indicating the theoretical feasibility of using AM fungi in the cultivation of A. lancea[86]. In addition, soil microorganisms contribute to the enhancement of A. lancea's growth and development, bolstering the plant's capacity to withstand stresses and expediting the accumulation of secondary metabolites. Notably, A. lancea has the capability to attract and foster specialized plant growth-promoting bacteria (PGPBs) from the soil. These PGPBs exhibit a marked and favorable correlation with A. lancea's secondary metabolites and soil nutrient content, making them potential prime candidates for employment as valuable biological resources in A. lancea cultivation and quality enhancement[87].
It can be seen that the key to achieving the high quality of TCM is to study the relationship between various ecological factors and the formation and accumulation of pharmacodynamic components, clarify the leading factors affecting the ecological environment, and further clarify the response process of key enzyme and hormone system changes affecting the synthesis of pharmacodynamic components to the ecological factors. The most important basis of this research idea is to elucidate the synthetic pathway of pharmacodynamic components. However, the lack of understanding of the biosynthetic pathway of most secondary metabolites including pharmacodynamic components is the main and most direct limiting factor that many studies have not been further in-depth.
Quality control of Rhizoma atractylodis
-
The existing TCM quality control mode mostly adopts the combination of traditional empirical identification (the appearance, color, smell, and other morphological characteristics)[88] and chemical composition analysis. With the continuous development of analytical technology, the quality evaluation method has gradually developed from the traditional identification mode (character, microscopic) to the accurate quantitative index component analysis (spectroscopic and chromatographic techniques). Taking the quality control standards of Rhizoma atractylodis in Chinese Pharmacopoeia as an example, the quality control items mainly include name, source, character, identification, inspection, content determination, etc. Among them, identification (such as TLC identification) and content determination items (atractylodin) mostly use spectrum or chromatography to qualitatively or quantitatively analyze the index components, aimed at ensuring the safety, authenticity, effectiveness, stability, and controllability of CMM. In the Japanese Pharmacopoeia, the differentiation between these two raw materials is achieved by evaluating their morphological characteristics[89] and detecting the sesquiterpenoid atractylon[90]. By comparing the differences in characteristics, microscopic identification, and volatile oil content of cultivated and wild A. lancea, A. chinensis, A. coreanae, and A.japonica, it found that the characteristics of cultivated Atractylodes were different from those of wild species. It is suggested that 'Knotty-lumpy' and 'Pale Reddish Brown to Reddish Brown' should be added to the character description of A. lancea in Chinese Pharmacopoeia[91].
The active components of Atractylodes are mainly volatile oil components, including terpenoids, alkynes, aromatics, and other components[92]. The volatile oil components of A. lancea and A. chinensis are mainly terpenes, accounting for 77.3% and 4.9% of the total volatile oil respectively, and the active components of both are atractylodin and eudesmol[93]. The chemical composition of A. macrocephala and A. japonica is similar, both dominated by atractylone, which can account for 60% and 5.18% of the total volatile oil, respectively[89]. A. coreana is less studied, and the main active ingredient is atractylone[94]. According to the Chinese Pharmacopoeia, the atractylodin in the herb Rhizoma atractylodis should not be less than 0.30%, but the quality evaluation of the herb also often uses atractylodin, β-eucalyptol, and atractylone as quality standards for evaluating Rhizoma atractylodis. The Chinese Pharmacopoeia only uses the extract as the quality evaluation standard of A. macrocephala, and the quality evaluation of the herb also often uses atractylone, atractylenolide I, atractylenolide II, and atractylenolide III as indicators for quality evaluation[2]. A long-term study by Lanping Guo showed that the volatile oil of A. lancea was highly variable among different origins and different individuals from the same origin. The variation in the volatile oil fractions and contents of the main active ingredients in A. lancea was correlated with its geographical distribution. The contents of the six main components of the volatile oil (elemol, atractylon, hinesol, β-endemol, selina-4(14), 7(11)-dienone, and atractylodin) showed a continuous decrease from south to north. The volatile oil of A. lancea is mainly of two chemical types, one is represented by Hubei type (HBA), mainly located in Hubei, Anhui, Shaanxi, and southern Henan, and is mainly composed of hinesol and β-endemol, with no or very small amounts of atractylon and selina-4(14), 7(11)-dienone. The other type is mainly represented by Jiangsu Maoshan, mainly located in the northern regions of Jiangsu, Shandong, Hebei, and northern Henan, and mainly composed of atractylon and atractylodin, referred to as Maoshan type (MA)[95]. This division method not only reflects the continuous variation of A. lancea volatile oil from south to north, reflects the correlation between the variation of A. lancea volatile oil and its geographical distribution, but also highlights the fact that atractylon and selina-4(14), 7(11)-dienone emerge from nothing during the continuous variation process. In the classification of chemical types, it is more convincing to use the presence or absence of certain chemical components in medicinal materials as the classification standard than to classify them according to their content, and it is operable in practice.
Herbal medicine and famous medicine experts of all dynasties in my country generally believe that A. lancea is an authentic medicinal material. However, at present, A. chinensis is the main product (more than 80%) in the Chinese market[95]. The germplasm resources of A. lancea are hybridized and degenerated, and a relatively high content of volatile oil has not been included in the pharmacopeia. Modern omics technology and chemometric methods have pushed the identification of the source of Rhizoma atractylodis to a new level. Many studies of comprehensive chemical pattern recognition of Rhizoma atractylodis were established by GC-MS[96] or UPLC[97] or HPLC[98] fingerprint, principal component analysis, cluster analysis, and discriminant analysis to distinguish genuine A. lancea, ungenuine A. lancea, and other Atractylodes. Thus, offering a valuable benchmark for the quality control and assessment of Atractylodes.
Therefore, it is suggested that in terms of planting, we should focus on optimizing the germplasm resources of A. lancea. In terms of quality control, the determination of volatile oil should be included in the Chinese Pharmacopoeia, and then combined with fingerprints or characteristic maps to determine the quality of A. lancea. A two-pronged approach will allow genuine medicinal materials to better serve TCM clinics.
-
The extensive use of Rhizoma atractylodis in various fields globally has made it a highly researched topic. The depletion of wild resources has emphasized the need to enhance the quality of cultivated varieties. Studies have revealed significant differences in a total of 76 volatile components, primarily including terpenes, aromatics, and esters, between wild and cultivated varieties of A. chinensis[99]. However, the variations in components among different Atractyiodes species and plant parts remain unclear, posing challenges in recognizing adulteration. Significant progress has been achieved over the past decade, notably in the fields of phytochemistry, quality assurance, metabolism/metabolomics, and biosynthesis, thanks to ongoing developments in analytical technology. The analysis of Rhizoma atractylodis has become more systematic, integrated, personalized, and intelligent, aligning with its multi-source and multi-component nature. DNA sequencing holds the potential to accurately identify different Atractyiodes species, even in extractive forms or Chinese medicine preparations. MSI can aid in imaging various Rhizoma atractylodis ingredients. Such as the cinnabar spot, at any location within the plant body, enabling the investigation of active constituents and revealing quality differences. The 'Flavor and Meridian tropism' as described in TCM theory, varies among different Atractyiodes species, but the underlying chemical basis remains unknown. Blosensor methods, such as electronic tongue and electronic nose, prove useful in distinguishing Atractyiodes species for quality control purposes.
As a secondary metabolite, the synthesis and accumulation of volatile oil from A. lancea are long and inefficient in living organisms. With in-depth research on the key enzymes and genes of the biosynthetic pathway of volatile oil in A. lancea, the use of genetic engineering, metabolic engineering, and synthetic biology can improve the efficiency and quality of volatile oil synthesis and save production costs. The findings indicate that the concentrations of β-eudesmol, hinesol, atractylon, and atractylodin in A. lancea are predominantly modulated by genetic factors. Clonal propagation emerges as a promising strategy to cultivate populations enriched with essential oil compounds. In a study conducted by Wang et al.[100], a tissue culture seedling with mutated characteristics from A. lancea was cultivated and identified as an exceptional germplasm resource. This mutant exhibited early stem growth and an elevated sesquiterpenoid content in contrast to the wild-type counterpart. The compound contents in A. lancea display significant variability influenced by their habitat. Another study by Yun et al.[101] focused on establishing and optimizing the in vitro cultivation of adventitious root in A. chinensis to produce atractylon and β-eudesmol. The researchers successfully optimized key parameters in the induction and suspension cultures of adventitious roots to maximize the efficiency of the culture process. At present, there are more sesquiterpenoids than polyacetylenes in the study of volatile oil components of A. lancea, and the synthetic pathways for sesquiterpenoids are more mature, whereas those for polyacetylenes are not yet well established. The synthesis mechanism of the atractylodin, as an index component of the Chinese Pharmacopoeia, will greatly limit the modernization of A. lancea if its synthesis mechanism is not fully understood.
The synthesis and accumulation of volatile oil, the main medicinal component of Rhizoma atractylodis, are influenced by a combination of genetic factors and environmental factors. The extent to which geographical variation is driven by genetic or environmental factors remains uncertain[102]. When the ecological environment changes, Rhizoma atractylodis will activate its defense system to protect itself from aggression. This includes changes in enzyme activity, metabolism, and osmotic balance regulation, which not only protect against external damage but also promote the accumulation of metabolites under certain conditions[103]. And reasonable regulation of environmental factors such as temperature, light, water, fertilizer, and biology on the growth and development of Atractylodes is necessary to effectively enhance the accumulation of secondary metabolites, thereby enhancing medicinal quality and yield. In conclusion, taking into account both genetic and environmental factors comprehensively, inducing key enzymes and gene overexpression in plants through external environmental factors can effectively promote the synthesis and accumulation of compounds in plants. This can help explore the mechanism of the superposition of genetic and environmental factors on the synthesis and accumulation of metabolites in medicinal plants. Despite the ups and downs in the modernization of Atractylodes, the tide is unable to return.
-
The authors confirm their contribution to the paper as follows: study conception and design: Guo L, Huang L, Gao W, Zhang W; data collection: Bai Q, Cui G; analysis and interpretation of results: Lyu C, Sun J; draft and revised manuscript preparation: Zhang W, Zhang X. All authors reviewed the results and approved the final version of the manuscript.
-
The complete chloroplast genome sequences of Atractylodes were submitted to GenBank (
www.ncbi.nlm.nih.gov ). This work was supported by the major projects of the National Natural Science Foundation of China (81891014) and special funds for the Construction of Modern Agricultural Industry Technology System of China (CARS-21).
-
The authors declare that they have no conflict of interest. Lanping Guo is the Editorial Board member of Medicinal Plant Biology who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer review handled independently of this Editorial Board member and the research groups.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang W, Bai Q, Cui G, Zhang X, Lyu C, et al. 2023. Recent progress and ongoing challenges in Rhizoma atractylodis research: biogeography, biosynthesis, quality formation and control. Medicinal Plant Biology 2:19 doi: 10.48130/MPB-2023-0019 

Recent progress and ongoing challenges in Rhizoma atractylodis research: biogeography, biosynthesis, quality formation and control
- Received: 09 June 2023
- Accepted: 26 September 2023
- Published online: 26 December 2023
Abstract: Rhizoma atractylodis, including Atractylodes lancea and Atractylodes chinensis, is widely utilized worldwide as a Chinese meteria medica (CMM) due to its rich content of various sesquiterpenoids and other medicinal components. Benefiting from the rapid and ongoing advancements in analytical technologies, significant strides have been achieved in advancing the analysis of Rhizoma atractylodis towards a more systematic, comprehensive, personalized, and intelligent approach. In this review, we highlight the progress made in the biogeography, biosynthesis, quality formation, and control of Rhizoma atractylodis over the past decade. Special attention is given to the incorporation of advanced genetics tools, modern omics technology, and chemometric methods that facilitate the accurate utilization of Rhizoma atractylodis. The elucidation of biosynthetic pathways, the formation of quality, and the control of active components continue to pose challenges in the study of Rhizoma atractylodis. Additionally, this review presents novel directions and opportunities aimed at enhancing ongoing research efforts.
-
Key words:
- Rhizoma Atractylodis /
- Germplasm resources /
- Biosynthesis /
- Molecular biology /
- Biogeography