








-
Ornamental plants, including herbaceous flowers, ornamental grasses, shrubs, and woody plants, form an important and rapidly growing sector of the green industry[1]. In addition to desirable aesthetic attributes, these crops enhance environmental quality, provide ecosystem services, increase property values, and are major economic drivers with annual direct sales of $156 billion in 2018[2]. Consumers and the industry demand new and attractive elite cultivars with enhanced biotic and abiotic resistance. Complementary to the traditional cross-hybridization, ploidy manipulation, and mutation breeding techniques, genetic modification via genetic engineering and gene editing hold tremendous promise for ornamental trait improvement. These bioengineering technologies can introduce unique genetic variations that are not available in current genetic resources and obtain improved traits in one or a few generations.
To date, about 50 ornamental species have been genetically transformed[1], and about 20 ornamental species have been gene-edited[3]. However, only three transgenic ornamental species (i.e., carnation, rose, and petunia) have been deregulated and approved for commercialization in a limited number of countries[1]. For example, the 'Moon' carnation varieties, with various flower colors, are commercially available in Australia, Norway, Japan, Colombia, Malaysia, European Union, and the U.S. The blue rose 'Applause' has been approved for marketing in Australia, Japan, and the U.S. and is restricted to greenhouse production for the export purposes in Colombia. A Petunia-CHS co-suppression transgenic event with altered flower color (from purple to white) has been approved for commercialization in China but is not on the market yet (
www.isaaa.org/gmapprovaldatabase ). The accidentally released 'orange' petunia varieties containing a maize A1-DFR transgene[4] have been approved for sale in Canada and deregulated recently in the U.S. (www.aphis.usda.gov/aphis/newsroom/stakeholder-info/SA_By_Date/SA-2021/SA-01 ). Detailed information for genetically engineered crops is available in the International Service for the Acquisition of Agri-biotech Applications (ISAAA) database (www.isaaa.org/gmapprovaldatabase ).Considering the large number of ornamental plant species, genetic engineering and gene editing in these crops are limited and lag behind major crops. There are multiple reasons for the slow development of bioengineered ornamental plants, e.g., limited resources, high deregulation cost, etc. Scientifically, plant transformation and regeneration difficulty are the main bottlenecks in applying genetic engineering and genome editing for trait improvement in specialty crops, including ornamental crops[5−7]. Fortunately, recent evidence shows that the use of growth and developmental regulator genes WUSCHEL (WUS), BABY BOOM (BBM), and growth-regulating factors (GRFs)–GRF-interacting factors (GIFs) can greatly improve transformation efficiency and speed up the process by promoting regeneration[6,8] (Fig. 1). We anticipate that these genes and other growth and developmental regulator genes could improve transformation efficiency and facilitate the application of new breeding technologies in ornamental plants. This review discusses the recent advances in the use of WUS, BBM, GRFs, and GRFs–GRFs in plant regeneration and their potential for ornamental crop transformation.
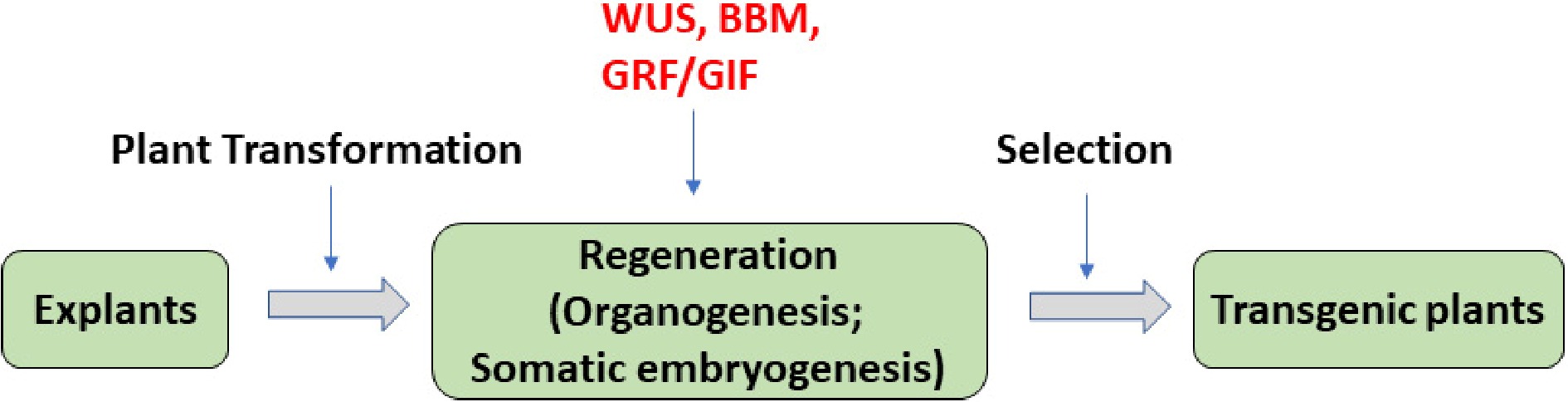
Figure 1.
The regeneration-promoting effect of WUS, BBM, GRFs, and GRFs–GRFs genes in plant transformation. In standard plant transformation systems, transgene is delivered into selected explants, plant tissue culture is used to induce plant regeneration, then the final transgenic plants are selected from regenerated plants. Among them, plant regeneration is often the bottleneck of the process. Genes WUS, BBM, GRFs, and GRFs–GRFs (red) could promote plant regeneration via either organogenesis or somatic embryogenesis in various plant species.
-
Critical needs and priorities for the nursery/floriculture industries include enhanced disease resistance, non-invasiveness, valuable commercial trait improvement, and advanced breeding technology development. With the completion of whole-genome sequencing for > 70 ornamental plants[9], transgenic breeding and gene editing offer tremendous potential and are within reach for many ornamental crops to address these critical needs worldwide. The release of GMO rose, petunia and carnation into the market[1] has paved the way for more genetically engineered and gene-edited ornamental crops. Nevertheless, the deregulation cost is often extremely high considering the market size and value for each ornamental species and variety. Recently, Bratlie et al.[10] proposed a relatively flexible regulatory framework to reasonably lower the regulatory hurdles for certain uses of GMO crops in the best interest of innovation and agroeconomics. Regulatory flexibility could apply to nursery/floricultural crops, making them ideal 'test cases' for trait engineering and improvement.
However, for multiple reasons, plant transformation is a primary bottleneck in applying advanced biotechnologies for crop trait improvement in nursery/floriculture crops. Agrobacterium- and biolistic-mediated transformation approaches are routinely used in ornamental plant transformation[5] and are highly genotype-dependent and ineffective in many plants, including ornamentals. Many ornamental genotypes are not transformable or show poor transformability due to high genetic diversity, polyploidy, complex reproduction systems, aging and maturity, desire for clonal stability, and limited input resources. Lack of effort and funding support exaggerates the situation since numerous trial-and-error experiments need to be conducted to establish an efficient callus induction and plant regeneration system for a given cultivar. Conventionally, different explants, medium components, and auxin/cytokinin ratios should be tested for every plant species or genotype, which is laborious, time-consuming, inefficient, and requires specialized skill and experience. Efforts to develop novel strategies to improve transformation efficiency and shorten the transformation process are needed in ornamental crops and elite cultivars.
-
Plant cell totipotency is the foundation of plant tissue culture and regeneration, allowing plants to be developed from a single cell via somatic embryogenesis or organogenesis[11]. Thus, plant genes, especially transcription factors (TFs) involved in plant embryo development and meristem maintenance, are logical targets for engineering to improve plant transformation and regeneration. WUS is a homeodomain transcription factor functioning as the primary regulator of stem cell fate and meristem maintenance in plants[12,13]. When screening for disrupted meristem maintenance, WUS was first discovered in an Arabidopsis ethyl methanesulfonate (EMS) mutant[14]. Chemical-induced activation of WUS expression caused enhanced somatic embryo formation in Arabidopsis[15]. Overexpression studies confirmed the embryogenesis-enhancing effect of WUS[16−18] and its organogenesis-promoting outcome in different species[19−20]. In addition, BBM is a member of the AP2/ERF family and a key regulator of plant cell totipotency[21]. BBM was identified during the in vitro microspore embryogenesis of Brassica napus[22]. Overexpression of BBM induced hormone-independent somatic embryogenesis in Arabidopsis and B. napus[22]. Through transgenic BBM overexpression, improvements in somatic embryogenesis enabled genetic transformation in previously transformation-recalcitrant sweet pepper[23]. Heterologous overexpression of Arabidopsis and B. napus BBMs in tobacco enhanced regeneration via organogenesis and induced somatic embryogenesis on a cytokinin-containing medium[24]. Thereafter, both WUS and BBM genes have been well studied for their effects on meristematic induction and maintenance and plant regeneration and transformation in various species (Table 1).
Table 1. The effects of WUS, BBM, GRFs, and GRFs–GRFs on plant development and genetic transformation.
Gene* Promoter Explants Effects Ref. AtWUS Estrogen-inducible A. thaliana root High somatic embryo formation frequency [15] Estrogen-inducible Nicotiana tabacum leaf Shoot formation from root tip [20] 35S Gossypium hirsutum hypocotyl Shoot formation from root tip [16] vsp1 Medicago truncatula seedling radicle 47.75% increase in embryogenic callus formation [18] ZmWUS2 ZmPLTP Zea mays immature embryo Enhanced callogenesis and embryogenesis [66] Nos A. thaliana (seedling), Solanum lycopersicum (seedling), N. tabacum (seedling/mature plant), Solanum tuberosum (mature plant), Vitis. vinifera (mature plant) de novo meristem induction [38] AtWUS-GR, AtSTM-GR 35S A. thaliana (floral dip) Triggered ectopic organogenesis [18] AtWUS, CHAP3A (PmLEC1) Estrogen-inducible Picea glauca immature embryo Did not induce somatic embryogenesis [59] eGFP-GhWUS1a, eGFP-GhWUS1b Estrogen-inducible G. hirsutum hypocotyl Inhibited embryogenic callus formation [60] AtBBM, BnBBM 35S, inducible N. tabacum leaf Enhance the regeneration capacity [24] BcBBM 35S Populus tomentosa calli Plant regeneration through somatic embryogenesis [25] BnBBM 35S, HnUbB1 A. thaliana (floral dip) B. napus haploid embryo Spontaneous formation of somatic embryos and cotyledon-like structures [22] BnBBM
EgAP2-1 (BBM)35S Capsicum. annuum cotyledon Made recalcitrant pepper transformable [23] 35S A. thaliana (floral dip) Enhanced regeneration capacity [63] GmBBM1 35S A. thaliana (floral dip) Induced somatic embryos on vegetative organs [64] TcBBM 35S A. thaliana (floral dip) Enhanced/hormone-independent somatic [65] AtBBM-GR 35S A. thaliana (floral dip) Improved plant regeneration for extended periods of time in tissue culture [62] HvWUS, HvBBM ZmAxig1, ZmPLPT Hordeum vulgare Co-expression increased transformation efficiency by 3 times [61] ZmBBM+ZmWUS2 ZmUbi, Nos Z. mays immature embryo, mature embryo, seedling leaf segment; Oryza sativa calli; Sorghum bicolor immature embryo; Saccharum officianrum calli Enabled transformation of recalcitrant varieties and/or increased transformation efficiency [26−28] ZmAxig1, ZmPLTP Z. mays immature embryo Established rapid callus-free transformation [29] ZmPLTP S. bicolor immature embryo Reduced genotype dependence, accelerated regeneration, increased transformation efficiency [67] AtGRF5/BvGRF5-L 2×35S Beta. vulgaris cotyledon, hypocotyl Enabled transformation of recalcitrant varieties. Increased transformation efficiency [33] AtGRF5/HaGRF5-L 2×35S Helianthus annuus cotyledon Improved transgenic shoot formation GmGRF5-L PcUbi4-2 Glycine. max primary node Improved transgenic shoot formation BnGRM5-L PcUbi4-2 B. napus hypocotyl Promoted callus production ZmGRF5-L1/2 BdEF1 Z. mays immature embryo) Increased transformation efficiency ~3 times TaGRF4-GIF1 ZmUbi Triticum aestivum immature embryo Increased regeneration efficiency 7.8 times; shortened protocol [34] O. sativa calli from seeds Increased regeneration efficiency 2.1 times ClGRF41-GIF1/VvGRF4-GIF1 35S Citrus limon etiolated epicotyl Increased regeneration efficiency ~4.7 times CIGRF42-GIF1 35S Citrullus lanatus cotyledon Increased transformation efficiency ~9 times [68] *At, A. thaliana; Zm, Z. mays; Pm, Picea mariana; Gh, G. hirsutum; Bn, B. napus; Bc, B. campestris; Eg, Elaeis guineensis; Gm, G. max; Tc, Theobroma cacao; Hv, H. vulgare; Bv, B. vulgaris; Ta, T. aestivum; Cl, 1C. limon, 2C. lanatus; Vv, V. vinifera. Constitutive or ectopic expression of these two genes commonly results in pleiotropic effects and subsequently abnormal plants[6]. Multiple strategies have been developed to use morphogenic genes to enhance transformation efficiency while maintaining normal plant growth[6,7]. One strategy is to utilize transitory morphogenic gene expression through chemically inducible systems to control their expression. Upon adding or removing the external stimuli, expression of the regulator genes can be turned on/off, or the function of regulator genes can be post-translationally controlled, limiting transgene-induced plant dysfunction[15,24]. Another strategy is to use site-specific recombinase systems such as Cre/LoxP and FLP/FRT to remove the regulator genes from the transgenic plant genome following plant regeneration. The expression of the recombinase genes and subsequent transgene excision can be controlled by environment-responsive (e.g., heat shock or desiccation) or development-/tissue-specific (e.g., meristematic) promoters. This strategy was first deployed in the Chinese white poplar[25]. The overexpressed B. campestris BcBBM gene was used to generate transgenic plants that exhibited abnormal phenotypes, including dwarfism and small wrinkled leaves[25]. However, heat shock-induced expression of FLP excised the BcBBM gene in these transgenic plants, resulting in transgenic plants with normal phenotypes[25].
Recently, an optimized procedure for using the maize ZmWUS and ZmBBM for plant transformation has enabled the successful transformation in many transformation-recalcitrant monocot genotypes[26]. Lowe et al.[26] reported that the combined expression of a weakly expressed ZmWUS and a strongly expressed ZmBBM significantly increased the transformation frequency in multiple maize inbred lines and several cultivars of sorghum, rice, and sugarcane. Moreover, a drought-inducible excision of transgenes before regeneration directly produced transgenic maize plants with normal phenotypes[26]. The effectiveness of this strategy was also confirmed in previously non-transformable maize and sorghum varieties[27,28]. In yet another iterative improvement, the maize auxin-inducible AXIG1 promoter was used to drive ZmWUS2 expression, while the maize phospholipid transfer protein (PLTP) gene promoter, which has strong expression in maize embryos and leaves, was used to drive ZmBBM expression. This approach resulted in direct somatic embryo development in various maize varieties and avoided the need for callus formation and excision of the transgenes to generate phenotypically normal transgenic plants[29].
-
GRFs and GIFs belong to a small family of highly conserved, plant-specific TFs in all land and charophyte plants[30]. They form a regulatory module with microRNA miR396 to control many aspects of plant growth and development, including the development of leaf, stem, seed, root, and flower, as well as plant responses to environmental stress conditions[31]. Recent research on the members of this gene family makes them a promising area of focus for biotechnology-based plant improvement for yield traits, given their roles in promoting cell proliferation and expansion[32]. Two independent studies using GRFs or GFR-GIF chimeras from different species demonstrated their enhancing effects on plant regeneration and transformation in various dicot and monocot species[33−34]. Interestingly, ectopic expression of these genes did not affect typical plant growth and development, leading to the regeneration of transgenic lines with normal phenotypes.
In a proof-of-concept study, Kong et al.[33] used the overexpressed Arabidopsis AtGRF5 for Agrobacterium-mediated sugar beet (B. vulgaris ssp. vulgaris) transformation and found that the transgenic calli surprisingly produced many shoots. Further tests with AtGRF5 and its sugar beet ortholog BvGRF5-like confirmed that GRF5 overexpression enhanced shoot organogenesis and improved transgenic plant regeneration in orthodox and recalcitrant sugar beet varieties[33]. Kong et al.[33] also extended their studies from GRF5 to different members of the GRF family and from sugar beet to various plant species. Overexpression of AtGRF5, AtGRF6, AtGRF9 or a putative B. napus ortholog BnGRF5-like only increased transgenic callus formation in canola. However, overexpression of AtGRF5 or its orthologs in soybean (GmGRF5-like) and sunflower (HaGRF5-like) increased transgenic shoot production in both soybean and sunflower[33]. Additionally, Kong et al.[33] demonstrated that overexpression of maize ZmGRF5-LIKE1 or ZmGRF5-LIKE2 enhanced maize transformation through somatic embryogenesis.
Debernardi et al.[34] created a wheat GRF4-GIF1 chimeric gene and tested its effects on the genetic transformation of wheat, the most difficult-to-transform cereal crop[35]. They found that the wheat GRF4-GIF1 chimera increased regeneration efficiency by 7.8-fold and shortened the transformation process by a month in the wheat varieties tested. The regeneration- and transformation-promoting effects of GRF4-GIF1 chimera were confirmed in previous transformation-recalcitrant genotypes, including commercial durum, bread wheat, and a triticale line. The use of the wheat GRF4-GIF1 chimera also increased the robustness and efficiency of previously developed wheat transformation protocols such as the John Innes Centre method[35] and the Japan Tobacco method[36]. Additionally, GRF4-GIF1 chimera enhanced the regeneration efficiency of citrus (C. limon) or grape (V. vinifera), indicating the chimera's effectiveness in dicots[34].
-
Recent advances in the studies of WUS, BBM, GRF5, and GRF-GIF chimeras have overcome the regeneration and transformation bottleneck in many plant species, including monocots and dicots with shortened transformation time even though these plant species use varied explants for transformation[26,27,33,34] (Table 1). The transformation-promoting effects of these genes have also been demonstrated in producing gene-edited plants[34,37,38]. With the help of these genes, marker-free transgenic plants can be generated – sometimes without the use of plant hormones such as cytokinin[34]. Thus, the translational studies of these genes in ornamental plants could provide tremendous opportunities for developing transgenic or gene-edited ornamentals. These genes promote regeneration in plant transformation in various plants, irrespective of explant type (Fig. 1; Table 1).
Effects of ectopic expression of these genes from Arabidopsis and maize could be readily tested in target ornamental plant species. While a total of 9, 29, and 13 GRF family members have been identified experimentally and in silico in Arabidopsis, poplar, and rice, respectively[31], only a few of them have been tested. Thus, the remaining GFR family members could be tested alone or in combination with different GIFs for their effects on regeneration and transformation. Since there is evidence that endogenous genes sometimes function better than homologs from other species[33], the homologs of these genes in ornamentals could be tested for their effects on regeneration and transformation of the same species. In addition, it is worthwhile to fine-tune the expression of WUS and BBM in combination with different GRF-GIF chimeras for any additive or synergistic effects on ornamental transformation and regeneration.
Conditional or inducible expression could be further explored in ornamental crops to minimize or eliminate the side effects of the continuous expression of these genes – especially WUS and BBM – in transgenic plant growth and development. Synthetic promoters could be used to regulate their expression[39,40]. In addition, transient expression[6,41] or protein delivery of these genes could be explored in explants cultured on callus induction medium, protoplasts[42], or suspension cells[43] of ornamentals. Such transformative approaches could enhance the opportunity to deliver non-GMO engineered ornamental cultivars, reducing regulatory hurdles and enhancing public acceptance.
In planta (ex vitro) transformation is ideal for ornamentals, especially for woody plants[7]. It has been demonstrated that gene-edited or transgenic plants could be rapidly created through de novo meristem induction from various soil-grown dicot plants[38]. More specifically, using Agro-injection to deliver WUS2 and BBM or IPT, transgenic shoots were produced in the mature plants of N. benthamiana, potato, and grape. Using Agro-injection to deliver WUS2 and BBM or IPT together with Cas9/gRNA, gene-edited transgenic shoots were also produced in the mature plants of these species[38]. This strategy allowed developmental regulator genes to extend in planta transformation to a broad range of plant species. Exploring additional developmental regulator genes and extending similar approaches to ornamental and woody plants is highly encouraged for ornamental crop improvement.
In addition, more morphogenic genes have been identified in plant meristem development and embryogenesis. Some of these genes, such as KNOTTED-1 (KN1)/SHOOT-MERISTEMLESS (STM)[44] and LEAFY COTYLEDON (LEC)[45], have been shown to increase the transformation efficiency in different plants. Other genes have not been extensively studied or optimized to promote plant transformation. These include CUP-SHAPED COTYLEDON (CUC) genes[46], ENHANCER OF SHOOT REGENERATION (ESR)[47], PLETHORA[48], WIND-INDUCED DEDIFFERENTIATIONs (WINDs)[49], ARABIDOPSIS RESPONSE REGULATOR (ARR)[50], and WUS-related homeobox (WOX) genes[51], ABAINSENSITIVE 3 (ABI3)[52], FUSCA3 (FUS3)[53], AGAMOUS LIKE (AGL15)[54,55], LEAFY COTYLEDON LIKE (LIL)[56], SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE1 (SERK1)[57], and RWP-RK DOMAIN-CONTAINING 4 (RKD4)/GROUNDED (GRD)[58]. These genes need to be tested individually, in combination, or with WUS, BBM and/or GRF-GIF to evaluate their potential effects on the transformation of ornamental plants.
-
Traditional plant transformation systems typically include tissue culture/regeneration, molecular cloning of constructs, construct delivery, and efficient selection of target events. However, regeneration can often be an insurmountable obstacle in the transformation of recalcitrant plants, including many ornamentals. The elucidation and application of WUS/BBM and GRF-GIF genes have considerable promise for overcoming the barrier. In some cases, the need for tissue culture can be avoided entirely using an Agro-injection approach. Further development of these approaches will enable the broad application of advanced breeding biotechnologies for ornamental crops.
The work was financially supported by the United States Department of Agriculture (USDA) - Agriculture Research Service (ARS) Base funds to the Duan laboratory, and the USDA Floriculture and Nursery Research Initiative (FNRI) grant # 8020-21000-071-23S and the USDA National Institute of Food and Agriculture (NIFA) Hatch project 02685 to the Liu laboratory. The authors thank the anonymous reviewers for their constructive comments and suggestions.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Duan H, Maren NA, Ranney TG, Liu W. 2022. New opportunities for using WUS/BBM and GRF-GIF genes to enhance genetic transformation of ornamental plants. Ornamental Plant Research 2:4 doi: 10.48130/OPR-2022-0004 

New opportunities for using WUS/BBM and GRF-GIF genes to enhance genetic transformation of ornamental plants
- Received: 04 December 2021
- Accepted: 23 January 2022
- Published online: 24 February 2022
Abstract: Broad application of plant transformation remains challenging because the efficiency of plant regeneration and regeneration-based transformation in many plant species is extremely low. Many species and genotypes are not responsive to traditional hormone-based regeneration systems. This regeneration recalcitrance hampers the application of many technologies such as micropropagation, transgenic breeding, and gene editing in various plant species, including ornamental flowers, shrubs, and trees. Various developmental genes have long been studied for their ability to improve plant meristematic induction and regeneration. Lately, it was demonstrated that the combined and refined expression of morphogenic regulator genes WUSCHEL and BABY BOOM could alleviate their pleiotropic effects and permit transformation in recalcitrant monocots. Moreover, ectopic expression of plant growth-regulating factors (GRFs) alone or in combination with GRF-interacting factors (GIFs) improved the regeneration and transformation of dicot and monocot species. Fine-tuning the expression of these genes provides new opportunities to improve transformation efficiencies and facilitate the application of new breeding technologies in ornamental plants.
-
Key words:
- Genetic transformation /
- WUS /
- BBM /
- GRF-GIF /
- Ornamental plants