








-
China is recognized as the most diverse country for bamboo, with 43 genera and 728 species[1]. Southwest China, particularly Yunnan Province is the richest area of bamboo diversity in the country, accounting for 50% of all bamboo species diversity[2]. Yunnan has three types of bamboo forests viz. cold-temperate, temperate, and tropical bamboo forests[3]. Since 2017, many studies of bambusicolous fungi have been conducted in Yunnan[2,4−10]. However, studies on bambusicolous fungi in this region remain limited compared to other regions such as Hong Kong and Taiwan[2,11]. Among these studies, Xylariales has been relatively under-researched in Yunnan (comprising 14% of known species), compared to Pleosporales (39% of known species). Nonetheless, Sordariomycetes has emerged as the largest group of Ascomycota found on bamboo[2].
The genus Oxydothis was introduced by Penzig & Saccardo[12] to initially accommodate three species (viz. O. grisea, O. maculosa, and O. nigricans) within the Amphisphaeriaceae, with O. grisea designated as the type species. The taxonomic placement of Oxydothis has been subject to extensive historical discussion[13−21]. A comprehensive study of Oxydothis was carried out by Konta et al.[20] who introduced Oxydothidaceae to accommodate the genus Oxydothis within Xylariales. Species of Oxydothis are characterized by solitary or aggregated ascomata that form in large groups, appearing as darkened, raised regions or dots on the host surface, and cylindrical asci with a J+ (rarely J-) subapical apparatus. The ascospores are filiform to fusiform, hyaline, 1-septate, and have pointed or blunt ends. In some species, ascospores produce appressoria upon germination[20,22]. The asexual morph of Oxydothis has been linked to Selenosporella species by Samuels & Rossman[23], although the sexual-asexual connection between Oxydothis and Selenosporella remains unproven. Furthermore, Samuels & Rossman[23] reported that the asexual morph of O. selenosporellae sporulated in vitro, producing black stromatic masses with perithecia developing throughout the colony. This morph displays macronematous, mononematous conidiophores that are (1–)2–3 septate, unbranched or branched, brown to olivaceous, with finely denticulate conidiogenous cells. A minute refractive scar remains after the conical dehiscence, and the conidia are arcuate, hyaline, and unicellular.
Oxydothis is a common genus mainly found on monocotyledons, such as bamboo (Poaceae), palms (Arecaceae), and Pandanus (Pandanaceae) where it exists mainly as a saprobe[18−20,22,24−29]. A few species have been reported as phytopathogens[30] or as endophytes[31]. Hyde et al.[22] suggested that the Oxydothis species may serve as biological control agents against plant pathogens. Species in this genus with appressorium formation are involved in protein and amino acid degradation, as well as secondary metabolite synthesis i.e. melanin biosynthesis[32]. Neoxylaria, Oxydothis, and Xylaria species collected from palms produce appressoria indicating they may have the ability to produce secondary metabolites especially when they live as endophytes[24,32]. Ninety-one epithets are currently listed under the Oxydothis in Index Fungorum (
https://indexfungorum.org/Names/Names.asp ; accessed on 11 November 2024), though more than half of the described species lack molecular data to clarify their phylogenetic relationships. Among those that do have sequence data, most species are represented only by ITS, LSU, and SSU sequences. Recently, Zhang et al.[29] introduced five new Oxydothis species (O. caryotae, O. foliata, O. palmae, O. pyriforme, and O. sinensis) and improved the phylogenetic resolution of the genus using multigene phylogeny based on a concatenated dataset of ITS, LSU, SSU, tef1-α, and rpb2 sequences. Unfortunately, only a few protein coding genes (i.e. rpb2 and tef1-α) are currently available for Oxydothis species in GenBank, resulting in incomplete classification of the phylogenetic relationships within the genus.During ongoing studies of Xylariomycetidae in Southwest China[33−37], numerous new species have been reported. As part of this continuing effort, the new species Oxydothis ailaoshanensis was discovered in the Ailaoshan rain forest of Yunnan Province, China on dead bamboo culms. This is the second report of an Oxydothis species associated with a bamboo host in China. The new species is described based on its morphological characteristics and supported by multi-gene phylogenetic analyses.
-
Dead bamboo culms were collected from the Ailaoshan subtropical evergreen broad-leaved forest (24.536944° N, 101.019444° E, 2,500 m), in Yunnan Province, China during the rainy season (April 2023). Specimens were processed according to the methods outlined by Senanayake et al.[38]. Observation and photography of the morphological features followed the procedure described in Dissanayake et al.[37]. Melzer's reagent was used to examine the apical apparatus of the asci. Measurements were taken using the Tarosoft (R) Image Framework software and photo plates representing the morphology were edited with Adobe Photoshop CS6 software (Adobe Systems, USA). Type specimens (holotype and isotype) were deposited in the Herbarium of Cryptogams, Kunming Institute of Botany, Chinese Academy of Sciences (KUN-HKAS), Kunming, China. The new taxon was registered in MycoBank (
www.mycobank.org ) and Facesoffungi[39].DNA extraction, PCR amplification, and sequencing
-
Fungal DNA was extracted from 15–20 fresh fruiting bodies of the fungus as described in Wanasinghe et al.[40] using Forensic DNA Kit (Omega®, Norcross, GA, USA). LSU, SSU, ITS, and tef1-α gene were amplified with primer pairs LR0R/LR5[41], NS1/NS4[42], ITS5/ITS4[42], and EF1-983F/EF1-2218R[43] respectively. The thermal cycling program was followed by Konta et al.[20] and Hu et al.[26]. The amplified PCR fragments were sent to a commercial sequencing provider (BGI, Ltd Shenzhen, PR China). All the sequences generated in this study were deposited in GenBank (Table 1).
Table 1. Names, strain numbers, and GenBank accession numbers of the taxa used in the phylogenetic analyses.
Species name Strain no. GenBank accession no. ITS LSU SSU tef1-α Oxydothis ailaoshanensis HKAS 130464T PQ635200 PQ587528 PQ587530 PQ584438 O. ailaoshanensis HKAS 130465IST PQ635201 PQ587529 PQ587531 PQ584439 O. calamicola MFLUCC 14-1165T NA KY206761 KY206767 NA O. caryotae HKAS 115712T PP592449 PP621075 PP639207 PP761002 O. chinensis ZHKUCC 22-0134T OR164912 OR164957 NA NA O. coperniciae CMUB 40043T PP278359 PP278849 PP278850 NA O. cyrtostachicola MRC0007T DQ660334 DQ660337 NA NA O. daemonoropsicola MRC 0005 DQ660335 DQ660338 NA NA O. dehongensis ZHKU 23-0986T PP580831 PP002130 PP002127 PP001172 O. dehongensis ZHKU 23-0987 PP580832 PP002131 PP002128 PP001173 O. dehongensis ZHKU 23-0988 NA PP002132 PP002129 PP001174 O. foliata MFLU 24-0165T PP592450 PP621076 PP639208 PP761003 O. fortunei GMBC0315T NR_187011 NG_228961 NA NA O. fortunei GMBC0389 ON510944 ON510945 NA NA O. frondicola HKUCC 3173/Mt14 AF009803 AY083835 AY083818 NA O. garethjonesii MFLUCC 15-0287T KY206773 KY206762 KY206768 KY206777 O. hohnelli HKUCC 3854 NA DQ810227 DQ810259 NA O. inaequalis MRC0004T DQ660336 DQ660339 NA NA O. metroxylonicola MFLUCC 15-0281T KY206774 KY206763 KY206769 NA O. metroxylonis MFLUCC 15-0283T KY206775 KY206764 KY206770 KY206779 O. narathiwatensis MFLUCC 24-0085T PP824654 PP824658 PP824659 NA O. palmae HKAS 115711T PP592451 PP621077 PP639209 PP761004 O. palmicola MFLUCC 15-0806T KY206776 KY206765 KY206771 NA O. phoenicis MFLUCC 18-0269T MK088065 MK088061 MK088063 MK087667 O. phoenicis MFLUCC 18-0270IS MK088066 MK088062 MK088064 MK087668 O. pyriforme HKAS 115710T PP592452 PP621078 PP639210 PP761005 O. rhapidicola MFLUCC 14-0616T NA KY206766 KY206772 NA O. sinensis GZCC21-0240T PP592453 PP621079 PP639211 PP761006 Oxydothis sp. JHGB17 3A MH268015 NA NA NA Oxydothis sp. IFO 32218 NA DQ810225 DQ810261 NA Oxydothis sp. E04B-2 PP592454 PP621080 PP639212 NA O. yunnanensis GZUCC 0127T ON176681 ON176684 NA NA Vialaea mangiferae MFLUCC 12-0808T KF724974 KF724975 NA NA V. minutella BRIP 56959 KC181926 KC181924 NA NA Superscripts 'T' and 'IST' represent the type and isotype strains. Newly generated sequences are indicated in bold. 'NA' sequences are unavailable. Phylogenetic analysis
-
Newly generated sequences were subjected to BLAST search in the NCBI GenBank database and sequences of closely related taxa were downloaded. Phylogenetic analysis was performed using ITS, LSU, SSU, and tef1-α sequences (Table 1). Multiple alignments, including both consensus sequences and reference sequences, were generated using MAFFT v. 7[44] and manually refined using BioEdit v. 7.0.5.2[45]. The individual datasets were combined into a concatenated dataset and further refined with BioEdit. Combined and individual datasets were subjected to maximum likelihood (ML) and Bayesian inference (BI) analyses. The best-fit substitution models were evaluated using MrModeltest v. 2.3[46] with the Akaike Information Criterion (AIC) as the selection criteria executed in PAUP v. 4.0b10[47]. ML and BI analyses were performed on the CIPRES Science Gateway platform[48]. For ML analyses, RAxML-HPC2 on XSEDE v. 8.2.10[49,50] was used, applying the GTR + I + G model with 1000 bootstrap repetitions. The BI analysis was executed with MrBayes on XSEDE v.3.2.7a[51−53] under the GTR + I + G, with one million generations and sampling every 100 generations. The analysis stopped automatically once the standard deviation of split frequencies fell below 0.01, with a burn-in fraction of 0.25. Phylogenetic results were considered significant if ML bootstrap values (MLB) ≥ 60% and Bayesian posterior (BYPP) ≥ 0.95, which were displayed above each node in the resulting tree. The phylogram was visualized using the FigTree v1.4.0 program[54], and final reorganization was done in Microsoft PowerPoint (2019).
-
The combined ITS, LSU, SSU, and tef1-α matrix comprised 34 strains, including Vialaea mangiferae (MFLUCC 12-0808) and V. minutella (BRIP 56959) as outgroup taxa. The concatenated alignment consisted of 3,421 characters (ITS: 1–680 bp, LSU: 681–1,510 bp, SSU: 1,511–2,512 bp, tef1-α: 2,513–3,421 bp), including gaps. The species-level relationships within Oxydothis in both ML and BI trees were similar in topology. The best-scoring RAxML tree was selected to represent the taxa relationship with a final likelihood value of –15,291.651757 (Fig. 1). The phylogenetic tree obtained from ML analysis in Fig. 1 showed a topology consistent with previous work[28,29]. The matrix had 990 distinct alignment patterns with 33.64% of characters being undetermined or gaps. The proportion of invariable sites I = 0.453933, the gamma distribution shape parameter alpha = 0.576006 and the Tree-Length = 1.338729. The Bayesian analysis ran for one million generations, with the average standard deviation of split frequencies reaching below 0.01 (0.009728). This analysis generated 1,922 trees, from which 721 were sampled after discarding the 25% as burn-in. The alignment contained a total of 995 unique site patterns.
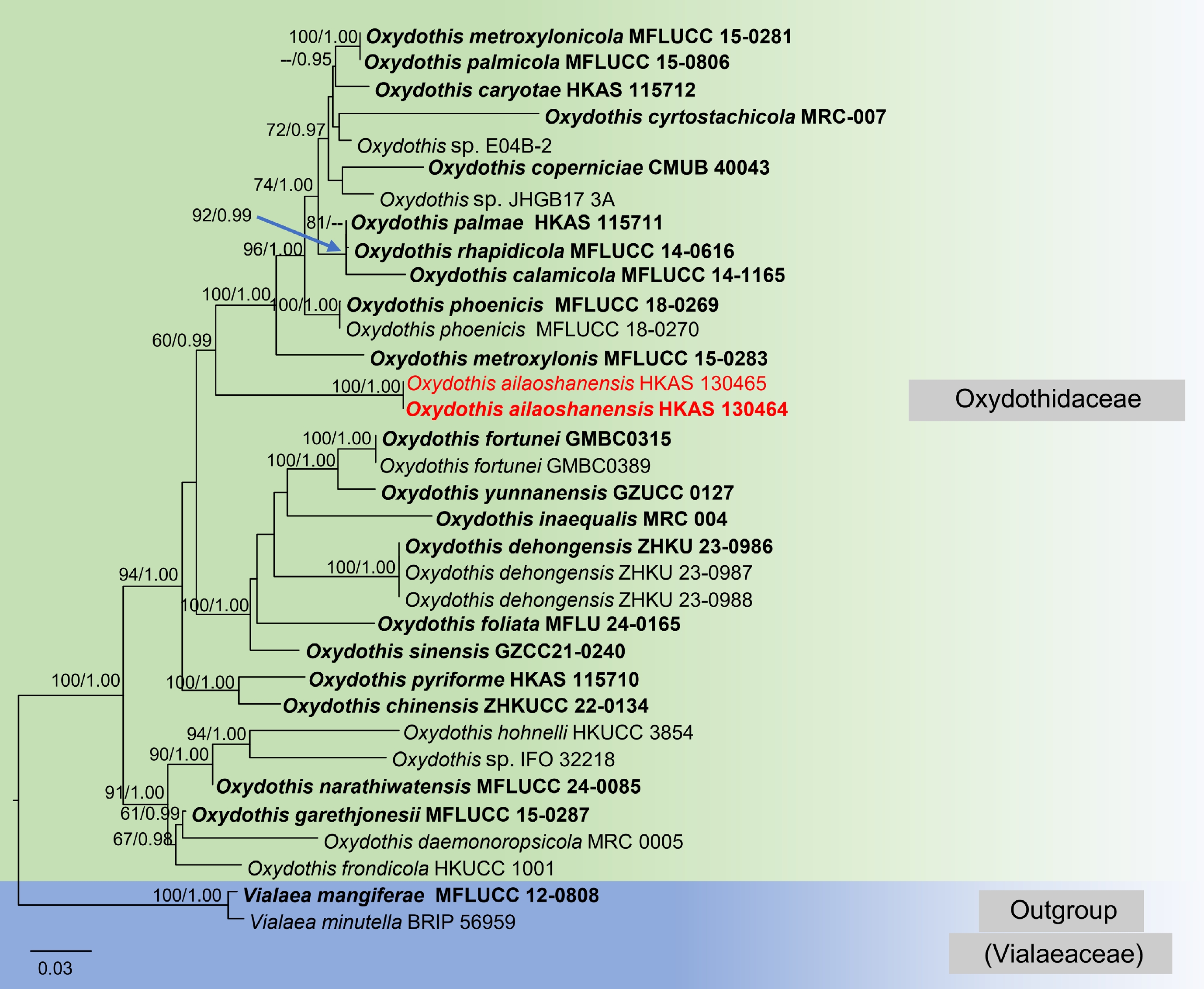
Figure 1.
RAxML tree based on a combined dataset of analyzed ITS, LSU, SSU, and tef1-α sequences. Bootstrap support values for ML equal to or greater than 60%, Bayesian posterior probabilities (BYPP) equal to or greater than 0.95 are shown as MLB/BYPP above the nodes. The new isolate is in red; ex-type strains are in bold. The scale bar represents the expected number of nucleotide substitutions per site.
Based on the results of multigene phylogeny, two strains of the new collection (Oxydothis ailaoshanensis sp. nov.) formed a robust subclade within the monophyletic clade of Oxydothis (60% MLB, 0.99 BYPP, Fig. 1). Oxydothis ailaoshanensis (HKAS 130464 and HKAS 130465) formed an independent lineage that is basal to O. calamicola (MFLUCC 14-1165), O. coperniciae (CMUB 40043), O. caryota (HKAS 115712), O. cyrtostachicola (MRC-007), O. metroxylonicola (MFLUCC 15-0281), O. metroxylonis (MFLUCC 15-0283), O. palmicola (MFLUCC 15-0806), O. palmae (HKAS 115711), O. phoenicis (MFLUCC 18-0269, MFLUCC 18-0270), and O. rhapidicola (MFLUCC 14-0616).
Taxonomy
-
Oxydothis ailaoshanensis L.S. Dissan., Phookamsak & Wanas. sp. nov. (Fig. 2)
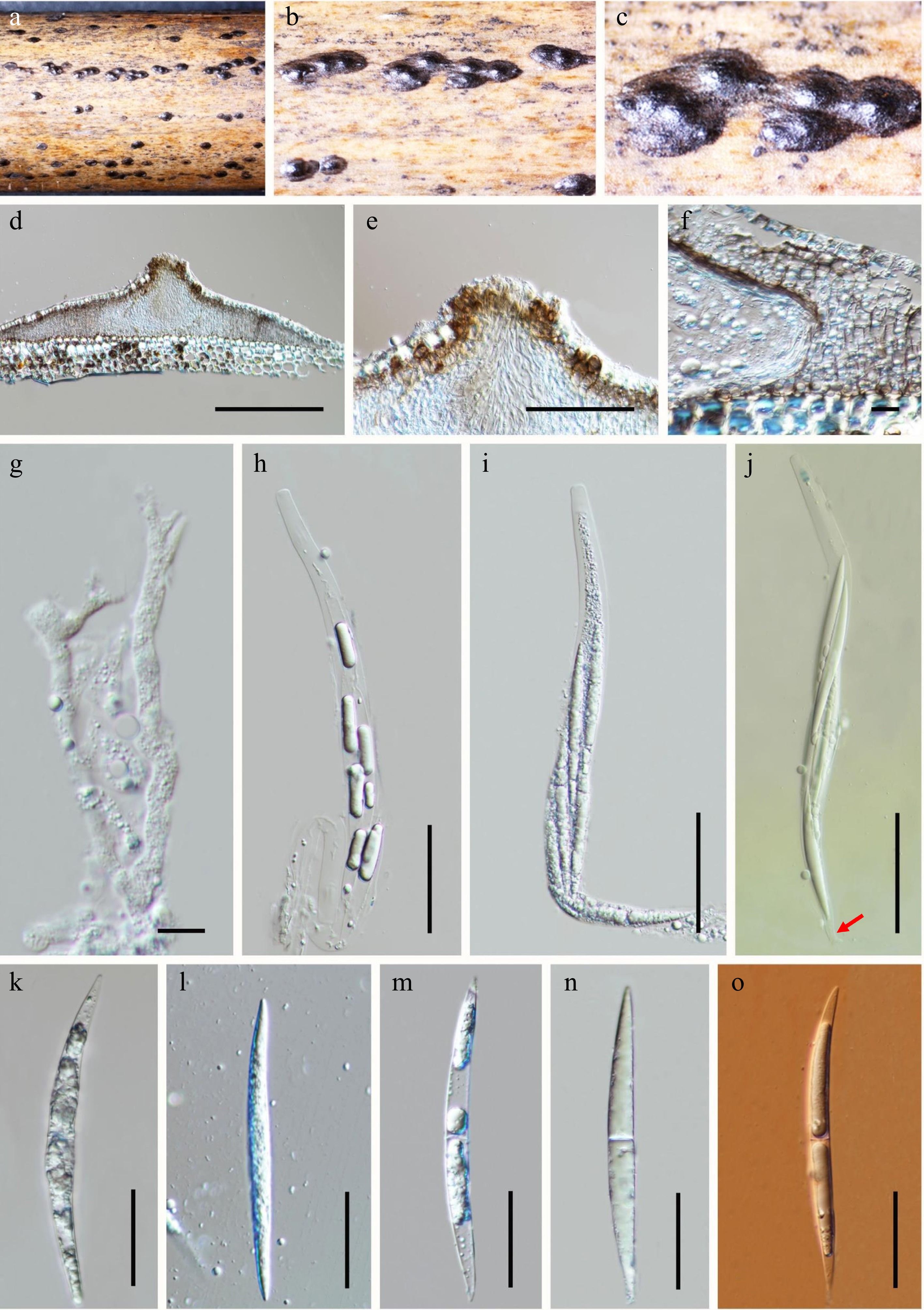
Figure 2.
Oxydothis ailaoshanensis (HKAS 130464, holotype). (a) Substrate. (b), (c) Ascomata on the host surface. (d) Section of an ascoma. (e) Close up of ostiole. (f) Peridium. (g) Paraphyses. (h)–(j) Asci (j = Asci in Melzer's reagent showing J+, apical apparatus, arrow showing short pedicel). (k)–(o) Ascospores. (o = Ascospore in Congo Red). Scale bars: (c) 200 μm, (e) 50 μm, (f, g) 10 μm, (h–j) 50 μm, (k–o) 20 μm.
MycoBank: MB856009; Facesoffungi number: FoF 14917
Etymology − The specific epithet is derived from the locality, Ailaoshan, where the holotype was collected
Holotype − HKAS 130464
Saprobic on dead bamboo culms (Poaceae). Sexual morph: Ascomata 450–550 μm high × 130–150 μm diameter (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Material examined – CHINA, Yunnan Province, Ailaoshan Forest Mountain (24.536944° N, 101.019444° E, 2,500 m), on dead culms of bamboo, 7 April 2023, L.S. Dissanayake, ALF23-10 (HKAS 130464, holotype), ibid., ALF23-10A (HKAS 130465, isotype).
Note: The multigene phylogenetic analyses indicate that Oxydothis ailaoshanensis is closely related to O. metroxylonis (MFLUCC 15-0283), with 60% MLB, 0.99 BYPP statistical support (Fig. 1). The nucleotide difference between O. ailaoshanensis and O. metroxylonis in ITS, LSU, SSU, and tef1-α are 124/555 bp (22.3%), 37/826 bp (4.5%), 13/981 bp (1.3%), and 95/906 bp (10.5%) respectively. Oxydothis ailaoshanensis differs from O. metroxylonis in having smaller, immersed ascomata (450–550 × 130–150 μm) that become raised and superficial with the long axis horizontal to the host surface, shorter asci (100–120 × 10–15 μm) that are mostly straight, and longer ascospores (55–75 × 3–5 μm) that are elongated and fusiform. In contrast, O. metroxylonis has larger ascomata (716–1,580 μm diam), with an axis oblique or perpendicular to the host surface, longer asci (165–181 × 9–15μm) with a cylindrical-clavate shape, and shorter ascospores (47–57 × 4–6 μm) with central curve[20]. Morphologically, O. ailaoshanensis is similar to O. bambusicola sharing characteristics such as solitary or aggregated ascomata with a central papilla, 8-spored, unitunicate, cylindrical asci with a J+, apical apparatus and elongated fusiform, 1-septate, hyaline ascospores[18]. Both species were isolated from bamboo hosts[18]. However, O. ailaoshanensis can be distinguished from O. bambusicola by its larger ascomata (450–550 × 130–150 μm vs 130–375 × 90–160 μm), shorter asci (100–120 × 10–15 μm vs 240 × 23 μm), and shorter ascospores (55–75 × 3–5 μm vs 93 × 7 μm) which taper gradually from the center to the pointed ends. In contrast, O. bambusicola ascospores are gradually tapering to the rounded apices and are covered with small amounts of mucilage. Based on the phylogenetic evidence and morphological differences, we describe our new collection as a distinct species, O. ailaoshanensis.
-
This study listed 89 accepted Oxydothis species in Table 2 with their hosts. Among them nearly all host species for Oxydothis belong to Arecaceae, except Oxydothis aequalis, O. bambusicola, and O. miscanthicola, recorded from Poaceae[18,31,55]. Notably, O. aequalis and O. bambusicola were recorded on bamboo hosts in the Philippines and the Hong Kong region in China respectively. In the current study, O. ailaoshanensis is introduced also from a bamboo host in China marking a new record for Poaceae hosts in Yunnan Province. Other Oxydothis species, i.e. Oxydothis caryotae, O. chinensis, O. fortunei, O. palmae, O. pyriforme, and O. sinensis were introduced from an Arecaceae host in China, Guangdong Province and Guizhou Province[26,27,29]. Based on the findings in Table 2, Oxydothis appears to exhibit host specificity primarily within Arecaceae and Poaceae with a distribution across both temperate and tropical regions.
Table 2. Host occurrences and distribution of all known Oxydothis species.
Species name Host Family Country Ref. Oxydothis acutata Orania spp. Arecaceae Philippines [31] O. aequalis Bamboo, Calamus sp. Arecaceae, Poaceae Australia, Brunei, Malaysia, Philippines [31,57] O. alexandrarum Archontophoenix alexandrae Arecaceae Australia [31,58] O. angustispora Licuala ramsayi Arecaceae Australia, Brunei, Thailand [57−59] O. asiatica Calamus flabellatus, Daemonorops sparsiflorus, Licuala sp. Arecaceae Australia, Brunei, China (Hong Kong) [57,60,61] O. asymmetrica Calamus conirostris Arecaceae Brunei [57] O. atypica Licuala longicalycata Arecaceae Thailand [62] O. atypica Licuala longicalycata Arecaceae Thailand [59] O. australiensis Archontophoenix sp. Arecaceae Australia [31] O. bambusicola Indocalamus sp. Poaceae China (Hong Kong) [18] O. batuapoiensis Daemonorops oxycarpa Arecaceae Brunei [57] O. belalongensis Licuala sp. Arecaceae Brunei [57] O. bruneiensis Calamus sp., Licuala sp. Arecaceae Brunei [57] O. calami Calamus sp., Salacca wallichiana Arecaceae Australia, Burma, China (Hong Kong), Indonesia, Myanmar, Philippines [31,57,60,61,63] O. calamicola Calamus sp. Arecaceae Thailand [20] O. caryotae Caryota sp. Arecaceae China (Guangdong Province) [29] O. chinensis Pandanus sp. Arecaceae China (Guangdong Province) [27] O. circularis Myrsine sp. Primulaceae Brazil [31] O. coperniciae Copernicia alba Arecaceae Thailand [28] O. cyrtospora Licuala ramsayi Arecaceae Australia [57] O. cyrtostachicola Cyrtostachys renda Arecaceae Thailand [19] O. daemonoropis Daemonorops sp. Arecaceae Philippines [31] O. daemonoropsicola Daemonorops margaritae Arecaceae Australia, China (Hong Kong), Malaysia, Thailand [57,58,60,61] O. dispariapicis Daemonorops oxycarpa Arecaceae Brunei [57] O. elaeicola Calamus sp., Elaeis sp., Livistona sp., Pandanus sp. Arecaceae Brazil, China (Hong Kong, Taiwan), Democratic Republic of the Congo, Honduras, Nigeria, Sierra Leone, Tanzania [31,60,64−67] O. elaeidis Elaeis sp. Arecaceae China (Taiwan), Democratic Republic of the Congo, Tanzania, Zaire [31,60,65,66] O. extensa Licuala ramsayi Arecaceae Australia [57] O. foliata Licuala sp. Arecaceae Thailand [29] O. fortunei Trachycarpus fortunei Arecaceae China (Guizhou Province) [26] O. froehlichii Calamus radicalis Arecaceae Australia [31] O. froehlichiae Calamus sp., Linospadix sp. Arecaceae Australia [31] O. frondicola Licuala sp., Archontophoenix sp. Arecaceae Australia, Malaysia, Thailand [31,58,59] O. garethjonesii Eleais sp. Arecaceae Thailand [20] O. gigantea Palm Arecaceae Australia, Indonesia [31,58] O. grisea Arenga sp., Calamus sp., Heliconia sp., Licuala sp., Ptychosperma sp. Arecaceae China (Taiwan), Malaysia, Indonesia, Venezuela [31,59,66,68] O. hoehnelii Arenga sp., Calamus sp., Licuala sp. Arecaceae Philippines [31,59−61] O. hongkongensis Daemonorops sp., Calamus sp. Arecaceae Australia, China (Hong Kong) [57,60,61] O. ianei Trachycarpus sp. Arecaceae China (Hubei Province), UK [58] O. inaequalis Wallichia siamensis Arecaceae Thailand [19] O. insignis Eugenia sp. Myrtaceae Brazil [31] O. licualae Archontophoenix sp., Calamus sp., Jessenia sp., Licuala sp. Arecaceae Australia, China (Hong Kong), Ecuador, Malaysia, Philippines, Thailand [31,57−60] O. licualicola Licuala sp. Arecaceae Brunei, Myanmar [57,63] O. linospadicis Linospadix microcarya Arecaceae Australia [30,31] O. livistonae Licuala sp., Livistona sp. Arecaceae Brunei, Philippines, Thailand [31,57,59,69] O. livistonica Calamus sp., Licuala sp., Livistona sp. Arecaceae China (Hong Kong), Japan, Thailand [31,57,59−61] O. livistonicola Licuala sp. Arecaceae Australia [57] O. luteaspora Calamus sp. Arecaceae Australia [31] O. maculosa Palm Arecaceae Indonesia [31,70] O. magnicolla Calamus sp., Licuala sp. Arecaceae Brunei [56] O. manokwariensis Calamus sp., Daemonorops sp. Arecaceae China (Hong Kong), Indonesia [31,57,60] O. maquilingiana Daemonorops sp. Arecaceae Philippines [31] O. mauritiae Mauritia flexuosa Arecaceae Ecuador [57] O. megalospora Calamus sp. Arecaceae Brunei [57] O. metroxylonis Metroxylon sagu Arecaceae Thailand [20] O. miscanthicola Miscanthus floridulus Poaceae China (Hong Kong) [54] O. narathiwatensis Eleiodoxa conferta Arecaceae Thailand [56] O. nigra Archontophoenix sp., Licuala sp., Livistona sp. Arecaceae Australia, China (Hong Kong), Malaysia [31,58,60,61] O. nigricans Ptychosperma sp. Arecaceae Indonesia [12] O. nonamyloidea Livistona sp. Arecaceae Indonesia [31] O. nonspecifica Calamus sp., Licuala sp. Arecaceae Australia, Brunei [57] O. nontincta Licuala sp. Arecaceae Brunei [57] O. nypae Nypa fruticans Arecaceae Brunei [31] O. nypicola Nypa fruticans Arecaceae Brunei [31] O. obducens Calamus sp., Linospadix microcarya Arecaceae Australia, China (Hong Kong) [31,57,60,61] O. oedema Mauritia flexuosa Arecaceae Brunei, China (Hong Kong), Guiana, Malaysia, Papua New Guinea, Seychelles [31,57,58,60,61] O. opaca Rhopalostylis sp., Ripogonum sp. Arecaceae New Zealand [31,73] O. oraniopsidis Calamus sp., Laccospadix sp., Licuala sp., Oraniopsis sp. Arecaceae Australia, India, Thailand [31,57,59] O. palmae Licuala sp. Arecaceae China (Guangdong Province) [29] O. palmicola Eleais guineensis Arecaceae Thailand [20] O. pandani Pandanus sp. Arecaceae China (Hong Kong), French Polynesia, Tubuai, United States [31,60,61] O. pandanicola Livistona chinensis, Pandanus sp., Pritchardia sp. Arecaceae Indonesia, Philippines, United States [31,57,60,61] O. parasitica Licuala ramsayi Arecaceae Australia [30,31,57] O. parvula Calamus sp., Orania sp., Phoenix sp. Arecaceae China (Hong Kong), Philippines [31,57,60,61] O. perangusta Licuala sp. Arecaceae Brunei [57] O. phoenicis Phoenix paludosa Arecaceae Thailand [22] O. poliothea Palm Arecaceae Venezuela [31] O. pusillispora Licuala sp. Arecaceae Brunei [57] O. pyriforme Licuala sp. Arecaceae China (Guangdong Province) [29] O. ragae Palm Arecaceae Indonesia [31] O. ragae Arenga sp. Arecaceae China (Hong Kong), Irian Jaya [31,60,61] O. rattanica Calamus sp., Daemonorops sp., Eleiodoxa sp. Arecaceae Brunei, China (Hong Kong), Thailand [57,60] O. rattanicola Calamus sp., Daemonorops sp. Arecaceae Australia, China (Hong Kong) [57] O. rhapidicola Rhapis excelsa Arecaceae Thailand [20] O. rhopalostylidis Rhopalostylis sapida Arecaceae New Zealand [23] O. rimicolla Calamus pogonacanthus Arecaceae Brunei [57] O. rubella Calamus sp. Arecaceae Australia [31,57,70] O. sabalensis Sabal sp., Serenoa sp. Arecaceae USA [71,31] O. saltuensis Archontophoenix sp., Cocos nucifera, Licuala sp., Livistona sp. Arecaceae Australia, Brunei, Indonesia, Papua New Guinea, Seychelles, Sri Lanka [31,57,58] O. selenosporellae Rhopalostylis sapida Arecaceae New Zealand [23,31,72,73] O. sinensis Livistona chinensis Arecaceae China (Guangdong Province) [29] O. tayabensis Calamus sp. Arecaceae Philippines [31] O. uniseriata Calamus radicalis Arecaceae Australia [57] O. wallichianensis Wallichia siamensis Arecaceae Thailand [19] Despite the substantial diversity within this genus, only 23 species currently have sequence data available in GenBank. Some species lack informative genetic markers such as ITS for Oxydothis calamicola, O. hohnelli, and O. rhapidicola, or LSU and SSU sequences data for O. chinensis, O. cyrtostachicola, O. daemonoropsicola, O. fortunei, O. inaequalis, and O. yunnanensis. Previous studies on Oxydothis taxonomy relied on ITS, LSU, and SSU sequences[20,22,26,28,56]. However, recent advancements in phylogenetic studies such as that by Zhang et al.[29], introduced five new Oxydothis species using a multigene dataset including ITS, LSU, SSU, tef1-α, and rpb2. Only nine species incorporate tef1-α in their datasets, further supporting the phylogenetic placement of Oxydothis species in Oxydothidaceae. In the present study, we contribute to this approach by providing a combined ITS, LSU, SSU, and tef1-α phylogeny for introducing O. ailaoshanensis. The use of multiple gene markers in phylogenetic analyses has enhanced our ability to resolve species-level relationships within Oxydothis.
While ITS, LSU, and SSU have previously been used as the primary markers for differentiating species, the addition of protein-coding genes such as tef1-α and rpb2 has improved resolution by increasing phylogenetic signal and reducing ambiguities in species placement[29]. For Oxydothis ailaoshanensis, the multigene analysis including tef1-α, coupled with morphological data such as the unique ascomata and spore characteristics (Fig. 2), has successfully differentiated it from close relatives such as O. metroxylonis. Nevertheless, some Oxydothis species still lack sequences for key informative gene regions, limiting comprehensive phylogenetic analyses. Future studies should focus on obtaining missing sequence data, especially for protein-coding genes, across more species in this genus. This could further clarify phylogenetic relationships and species boundaries within Oxydothis.
An ITS BLAST search of the sequences linked Oxydothis to some leaf-litter-based ascomycetes (i.e., AF502894, AF502896, AF502740) and various uncultured fungal strains (i.e., KT328718, GU174316, AM999626, KC222801). However, these strains lack associated morphological data, which limits deeper insights into their morpho-phylogenetic relationships. This absence of morphological connections emphasizes the need for comprehensive morphological and molecular characterization of these strains. The present findings suggest that Oxydothis diversity remains underexplored in this region, with potentially numerous species still awaiting discovery.
Rungtiwa Phookamsak sincerely acknowledges the Introducing Talents Start-up Fund of Kunming Institute of Botany, Chinese Academy of Sciences, Yunnan Revitalization Talent Support Program 'Young Talent' Project (Grant No. YNWR-QNBJ-2020-120), Yunnan Revitalization Talent Support Program "High-end Foreign Expert" Project and the Independent Research Department of Economic Plants and Biotechnology, Yunnan Key Laboratory for Wild Plant Resources, Kunming Institute of Botany, Chinese Academy of Sciences (Grant No. Y537731261). Jianchu Xu thanks the Yunnan Department of Sciences and Technology of China (Grant No. 202302AE090023, 202303AP140001). Dhanushka N. Wanasinghe is funded by the Distinguished Scientist Fellowship Program (DSFP), King Saud University, Kingdom of Saudi Arabia.
-
The authors confirm contribution to the paper as follows: conceptualization, formal analysis, writing – original draft: Dissanayake LS; data curation: Dissanayake LS, Wanasinghe DN; methodology: Dissanayake LS, Phookamsak R; resources, project administration: Wanasinghe DN, Xu J; supervision: Xu J; writing – review and editing: Wanasinghe DN, Phookamsak R. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated for this study can be found in the NCBI, GenBank and MycoBank.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Dissanayake LS, Phookamsak R, Xu J, Wanasinghe DN. 2024. Oxydothis ailaoshanensis sp. nov. (Oxydothidaceae, Xylariales) from dead bamboo culms in Yunnan Province, China. Studies in Fungi 9: e016 doi: 10.48130/sif-0024-0016 

Oxydothis ailaoshanensis sp. nov. (Oxydothidaceae, Xylariales) from dead bamboo culms in Yunnan Province, China
- Received: 12 November 2024
- Revised: 05 December 2024
- Accepted: 05 December 2024
- Published online: 20 December 2024
Abstract: During the investigation of ascomycetes on Poaceae in Yunnan Province, China, a fungus was collected from dead bamboo culms in a terrestrial habitat in the Ailaoshan subtropical evergreen broad-leaved forest. Based on morphological characterization and phylogenetic analyses, this fungus was introduced as Oxydothis ailaoshanensis sp. nov. Maximum likelihood and Bayesian inference analyses of a concatenated dataset of internal transcribed spacer (ITS), large subunits (LSU) of the nuclear ribosomal RNA gene, small subunit (SSU) of the nuclear ribosomal RNA gene, and translation elongation factor 1-alpha (tef1-α) sequences were performed to clarify the phylogenetic affinities of the new species. Phylogenetically, Oxydothis ailaoshanensis forms an independent lineage, basal to O. metroxylonis. Oxydothis ailaoshanensis differs from O. metroxylonis in having smaller, immersed ascomata that become raised and superficial with the long axis horizontal to the host surface, shorter asci that are mostly straight, and longer ascospores that are elongated and fusiform. The new species was justified based on morphological traits and multigene phylogenetic analyses in comparison with closely related species. A detailed description, micrograph, and a phylogenetic tree of the new species are provided.
-
Key words:
- 1 new species /
- Ailaoshan forest /
- Bambusicolous fungi /
- Molecular phylogeny /
- Sordariomycetes /
- Taxonomy