








-
Tomato (Solanum lycopersicum L.), an important climacteric vegetable crop, is not only consumed globally as a fresh and healthy product, but also the model plant for the study of fleshy fruit development. In 2019, the world's cultivated crop area and yield of tomato were 5.03 M ha and 180.77 M t, respectively (FAOSTAT, 2021,
www.fao.org/faostat/en ). Nutrition and flavor of product are significantly correlated with consumer preferences. In terms of nutrients, tomatoes contain a variety of compounds, including: carotenoids (lycopene, β-carotene), phenolic (phenolic acids and flavonoids), vitamin (VC, VE), amino acids, minerals, etc., which endow fruit with anti-inflammatory, prominent antioxidant and anticancer activities[1]. Besides their functions as nutrients, carotenoids, polyphenols, and vitamins are also involved in response to both biotic and abiotic stresses in plants, participating in the process of stress defense that translate to crop yield stability. But the levels of these compounds vary significantly among commercial tomato varieties. For example, 'Tigerella' and 'Orange' presented the higher values of lycopene and phytoene, where the concentration reached 100 mg/100 g DW ~250 mg/100 g DW. While the content of these substances in 'Sunchocola' is only about 12 mg/100 g DW ~15 mg/100 g DW[2]. Nutrient deficiencies in tomato fruits may have adverse effects on human health, especially for populations in poor areas.During fruit ripening, the accumulation of the main biologically active substances are catalyzed by a series of enzymes encoded by structural genes, such as 1-DEOXY-D-XYLULOSE-5-PHOSPHATE SYNTHASE (DXS), ISOPENTENYL DIPHOSPHATE ISOMERASE (IDI), CHALCONE SYNTHASE (CHS), CHSCHALCONE ISOMERASE (CHI), PHOSPHOMANNOMUTASE (PMM), ASCORBATE OXIDASE (AO) and others[3−5]. Over the years, the synthetic pathways of these important compounds are constantly being supplemented with the evolution of molecular biotechnology[6,7]. The rate-limiting enzyme in the metabolic pathways has been identified, their expression can be triggered by developmental signals and highly regulated at multiple levels during the plant life cycle. As a complex and important biological process, fruit ripening is controlled by a complex regulatory network of interactions among multiple hormones, transcription factors (TF) and epigenetic modifications at the level of chromatin accessibility, transcription, translation and post-translational modifications. Although many of these factors have been identified and are also key molecular switches controlling compounds' biosynthesis in fruits, it remains largely unclear what upstream compounds might affect the transcriptional activation activity of them and what specific molecular regulatory mechanisms do they use to specifically regulate the expression of downstream genes.
The lack of flavor of fresh tomato fruits was the main factor affecting consumer consumption for decades. Flavor is the result of the interaction of many substances. The ratio of sugars and acids is the main determinant of tomato taste[4]. Volatile compounds derived from aromatic amino acids, fatty acids, and carotenoids are another important component of fruit flavor. Even though a number of compounds associated with flavor-imparting have been identified, flavor quality improvement is still a huge challenge that breeders face due to their high degree of complexity. On the one hand, the perception of flavor is not due to the additive effect of different compounds but their interaction, altering one or a few of them is unlikely to have a major effect on improving fruit flavor[8]. On the other hand, although multiple volatiles were identified in tomato fruit, the most important chemicals that have positive or negative effects on consumer preferences are still not fully understood. In order to effectively promote the improvement of the flavor profile of tomato fruits, we have summarized some important compounds that may play an important role in consumer preferences, and reported the key regulatory factors controlling their output.
-
Carotenoids are the main pigments in tomato fruits, conferring the fruits characteristic red coloration and antioxidant capacity[1]. They appear at the tomato breaker stage and accumulate in a type of plastid named chromoplast[9]. The major carotenoids found include: lycopene, lutein, phytoene and β-carotene. The concentration of carotenoids reported in tomatoes ranged from 2.2 to 297.9 mg/100 g DW, this difference is associated with tomato varieties[2]. In recent years, significant progress has been made in carotenoid metabolism study, the structural genes and enzymes that catalyze the core reactions of carotenoid biosynthesis and degradation have been identified in plants. As shown in Fig. 1, carotenoids are produced from the condensation of the isopentenyl diphosphate (IPP, isopentenyl diphosphate, C5) and dimethylallyl diphosphate (DMAPP), which are synthesized through the MEP (2-C-methyl-D-erythritol 4-phosphate) pathway[8]. In this pathway, DXS (1-Deoxy-D-xylulose-5-phosphate synthase) and DXR (1-deoxy-d-xylulose 5-phosphate reductoisomerase) are essential enzymes that determine the production rate of IPP. The expression of SlDXS is organ-specific, the highest levels of SlDXS1 transcripts were found in later stages of fruit ripening. Studies have shown the overexpression of the genes SlDXS1 and SlDXR could increase the carotenoids yields[10], whereas lack of DXS activity prevents formation of plastidial isoprenoids and strongly impairs plastid biogenesis[11]. Hydroxymethylbutenyl diphoshate reductase (HDR) is the second most important enzyme in the MEP pathway. Overexpression of HDR in Arabidopsis seedlings leads to increased accumulation of β-carotene and lutein[12], but there are few reports on the expression pattern of HDR in tomato fruit. When three IPP units were added to one DMAPP molecule, carotenoids begin to be synthesized in tomato plastids. The synthesis of 15-cis-phytoene, catalyzed by phytoene synthase, is a key control point in the formation of carotenoids, this process has often been the target of attempts to increase carotenoid accumulation in tomato fruit. Tomatoes have three PSY genes (PSY1, PSY2, PSY3), both PSY1 and PSY2 genes express in the fruits, but only the PSY1 mediate the fruit-specific carotenoid biosynthesis. Using VIGS, Fantini et al.[13], found PSY1-silenced fruits displayed a phenotype similar to yellow flesh and repressed carotenoid accumulation. Notably, the latest research shows that PSY2 (mediated leaf-like carotenogenesis) also remains functional in ripening tomato fruits and provides precursors for abscisic acid formation in fruits[12, 14]. Uncolored 15-cis-Phytoene then forms all-trans-lycopene (the predominant pigment in fruits) via catalyzation by the enzymes: phytoene desaturase (PDS), ζ-carotene desaturase (ZDS), 15-cis-ζ-carotene isomerase (ZISO) and Carotenoid isomerase (CRTISO). Inhibition of PDS by Norflurazon (a chemical inhibitor that can inhibit the expression of PDS) blocks carotenogenesis and bleaches photosynthetic tissue due to photooxidation of chloroplasts[15]. Interestingly, differential regulation of SlZDS and SlCRTISO was demonstrated in ripe peel and flesh, and their expression was higher in peel compared to flesh[16]. Hence, the peel of tomatoes should not be removed. Important branching pathways emerge in the ensuing metabolic process, which is a crucial step in generating carotenoid diversity. Furthermore, a greater variety of carotenoid isoforms have been shown to increase the antioxidant capacity of tomato fruit[17]. Recently, Mi et al.[18] expressed the LCYB-encoding gene of daffodil and Erwinia uredovora in tomatoes and demonstrated LCYB overexpression increased nutritional value conferred by the higher accumulation of β-carotene and β-ionone, and enhanced abiotic stress tolerance and fruit shelf life. This strategy provides an important direction for the development of tomato lines with high nutritional quality and outstanding environmental tolerance.
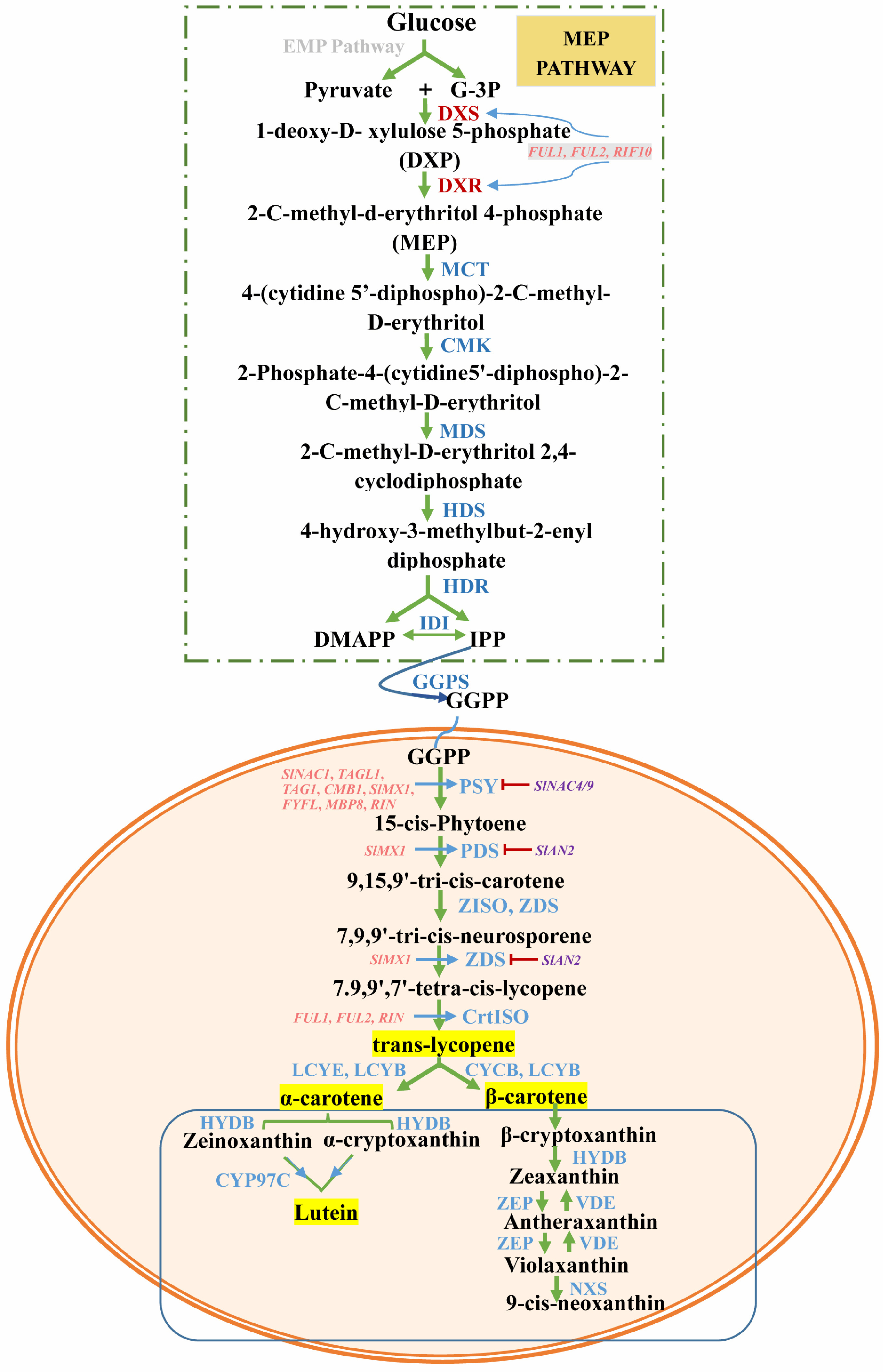
Figure 1.
The pathway of carotenoid biosynthesis and its transcriptional regulation. The pathway in the rectangle above represents the production of substrates for carotenoid synthesis through the MEP pathway. The pathway in the lower ellipsis represents the synthesis and decomposition of carotenoids in plastids. Enzymes are shown in blue. Blue arrows denote positive regulation, whereas red arrows denote negative regulation. DXS, 1-deoxy-D-xylulose-5-phosphate synthase; GGPP, geranylgeranyl pyrophosphate; DXR, 1-deoxy-d-xylulose 5-phosphate reductoisomerase; MCT, 2-C-methyl-d-erythritol 4-phosphate cytidylyltransferase; CMK, 4-(cytidine 5′-diphospho)-2-C-methyl-d-erythritol kinase; MDS, 2-C-methyl-d-erythritol 2,4-cyclodiphosphate synthase; HDS, (E)-4-hydroxy-3-methylbut-2-enyl diphosphate synthase; HDR, (E)-4-hydroxy-3-methylbut-2-enyl diphosphate reductase; IDI, isopentenyl diphosphate isomerase; GPPS, geranyl diphosphate synthase; PSY, phytoene synthase; PDS, phytoene desaturase; ZDS, ζ-carotene desaturase gene; ZISO, ζ-carotene isomerase; CrtISO, carotene isomerase; LCY, lycopene beta-cyclase; HYDB, β-carotene hydroxylase; CYP97C, cytochrome P450-type monooxygenase 97C; VDE, violaxanthin de-epoxidase; ZEP, zeaxanthin epoxidase; NXS, neoxanthin synthase.
The content and composition of carotenoids in fruit is not only related to the biosynthesis pathway, but also have relations with its degradation pathway. Carotenoids are important precursors of a number of bioactive compounds in plants, animals and fungi. They are cleaved either enzymatically, in general by carotenoid cleavage dioxygenases (CCD) and 9-cis-epoxycarotenoid dioxygenases (NCED) or non-enzymatically by reactions initiated by ROSs. In plants, CCDs can be divided into four groups CCD1s, CCD4s, CCD7s and CCD8s. Among them, due to the very relaxed double bond and substrate specificity of tomato SlCCD1A and SlCCD1B enzymes, they cleave carotenoids to form many tomato isoprenoid volatiles[19, 20]. Although carotenoid accumulation has been shown to be negatively associated with CCD expression, increasing the level of carotenoid by reducing carotenoid degradation is not considered an optimal approach due to carotenoid degradation being positively associated with formation of flavor and ABA[19].
Polyphenols
-
Tomatoes are important source of many polyphenolic compounds, including naringenin chalcone, chlorogenic acid, rutin, quercetin, hydroxybenzoic, caffeic acid, hydroxycinnamic, naringenin, and so on. And flavanols (including glycosides), anthocyanins, hydroxycinnamic acids and flavanones are the mainly polyphenols in tomato fruits. These compounds are involved in many important processes in plants, including pigmentation in fruits and vegetables, defense against biotic/abiotic stresses, as precursors for volatile production[21], and as metabolic markers (NC and its glycosides) for fruit ripening in tomatoes[22]. At present, Tohge et al.[23] have established an entire pan-species metabolic network of polyphenolic metabolism in tomatoes by utilizing natural variation. As shown in Fig. 2, the phenylpropanoid pathway provides a precursor for the production of flavonoids, and phenylalanine ammonialyase (PAL) is the first and key enzyme of this pathway. PAL proteins present high degrees of conservation, it can transfer the central flux of carbon from primary metabolism to the synthesis of countless phenols. PAL is encoded by a small gene family in plants with four members in Arabidopsis (PAL1–PAL4), and induces the expression of PAL1 and PAL2 which can increase the accumulation of flavonoids, anthocyanins, and sinapic acid[24]. While PALs play an important role in the accumulation of polyphenols, there are few reports about PAL genes analysis involved in tomato fruits. Cinnamate 4-hydroxylase (C4H, P450) is the second key enzyme in the phenylpropanoid pathway and is a major flux control for lignification. C4H is generally co-expressed spatially and temporally with other genes encoding phenylpropane pathway enzymes. According to previous reports, the expression of C4H can be caused by pathogens, inducers of defense responses, ultraviolet light, drought, and other environmental stresses, thereby imparting disease resistance to plants. However, for fresh fruits, high levels of lignin adversely affect consumer acceptance, and thereby reduce fresh market value. CYP73A24(class I C4H) is the only described full-length C4H sequence from tomato, its sense expressing resulted in considerable decrease in lignification and an increase in antioxidant flavonoids[25]. Following, the acetylation of coumaric acid is a central branch point in the phenols biosynthetic pathway. The action of chalcone synthase (CHS) represents the beginning of the specific flavonoid pathway. In general, normal tomato can produce anthocyanins in vegetative tissues, its fruit contains low levels of flavonoids and no anthocyanins[26]. Hence, increasing the level of flavonoids and anthocyanins in tomato fruits has been the focus of scientific attention. One of the major limiting factors in the flavonoid biosynthetic pathway is the lack of expression of the chalcone isomerase (CHI) gene in tomato fruit, which may be caused by a mutation in the promoter[27]. However, this limitation was broken with the discovery of the Aft gene that is more likely a regulatory gene or promoter region of a structural gene[28]. This gene is located in chromosome 10 and its presence in tomato leads to production of higher levels of the flavonols and anthocyanin pigment in tomato fruit. Similarly, concomitant ectopic expression of structural gene CHI and regulatory gene PAP1 (clone from onions) in tomatoes by gene stacking have been reported to also significantly increase rutin (130 times) and total anthocyanin (30 times) content[29]. Compared with the numerous studies on the biosynthesis pathways of polyphenols, the molecular and biochemical features associated with their degradation in plants have received little attention.
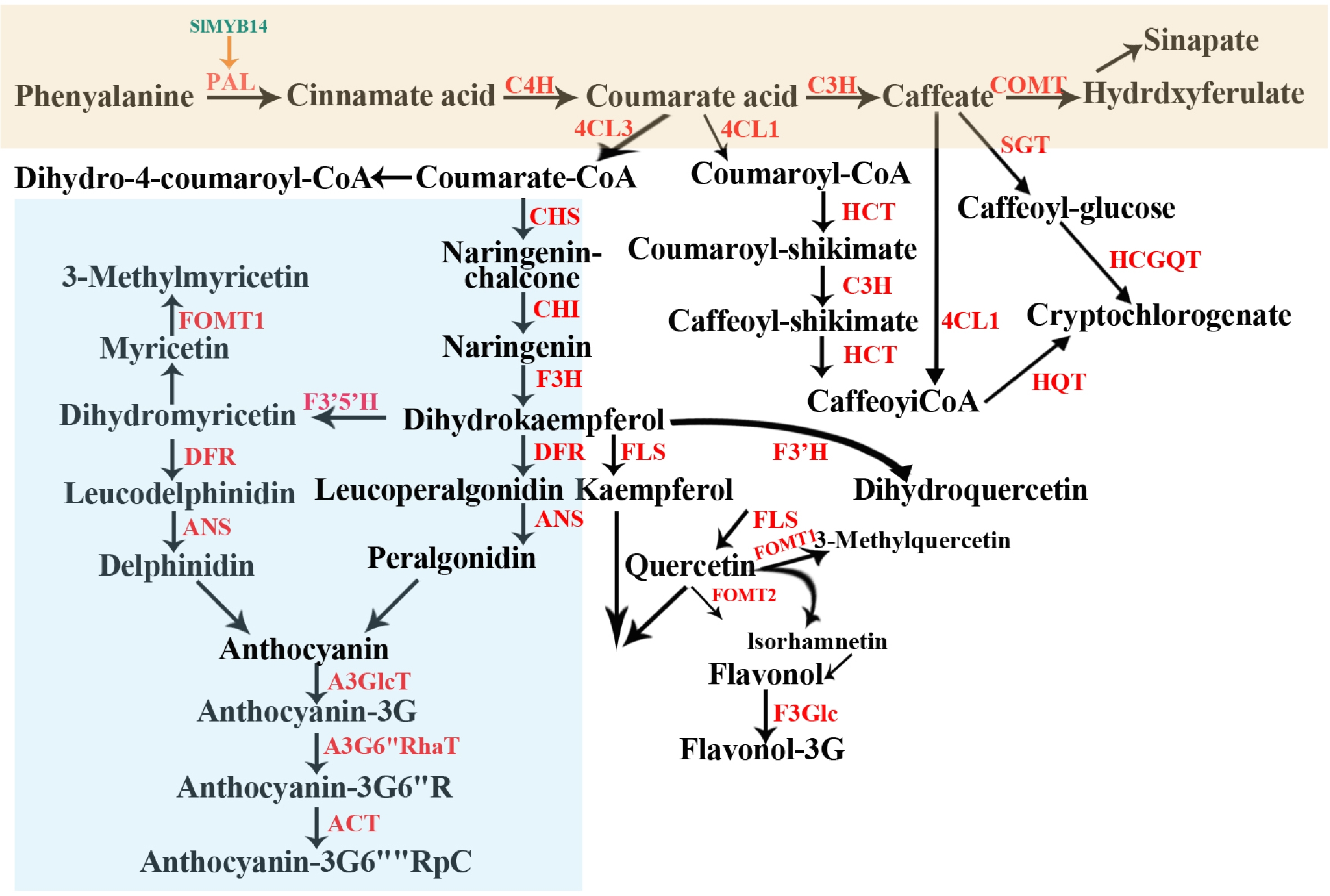
Figure 2.
Overview of the synthesis and metabolism framework of polyphenols in tomato. Enzymes are shown in red. Different colored rectangles represent the synthesis of different polyphenols. PAL, phenylalanine ammonia-lyase; C4H, cinnamate-4-hydroxylase; C3H, coumarate 3-hydroxylase; 4CL, 4-coumarate CoA ligase; COMT, catechol-Omethyl transferase; CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavanone 3-hydroxylase; DFR, dihydroflavonol reductase; ANS, anthocyanidin synthase; FOMT, flavonoid O-methyltransferase; F3'5'H, flavonoid-3,5-hydroxylase; A3GlcT, anthocyanin-3–O-glucosyltransferase; A3G6''RhaT, anthocyanin-3–O-glucoside-6′′–O-rhamnosyltransferase; ACT, Acylase; HCT, hydroxycinnamoyl-CoA shikimate/quinate hydroxycinnamoyltransferase; HCGQT, hydroxycinnamoyl glucose quinate hydroxycinnamoyl transferase; SGT, Solanidine: UDP-glucosyltransferase; HQT, hydroxycinnamoyl-Co A quinate hydroxycinamoyl transferase; F3'H, flavonoid 3-Hydroxylase; FLS, flavonol synthase.
Vitamin C
-
Vitamin C (ascorbic acid, VC) is well known for its powerful antioxidant activity, ROS scavenger capacity and its ability to enhance plant tolerance to stress. Regarding the synthesis pathway of plant VC, it can be roughly divided into the non-reversal and remedial pathway, and D-mannose/l-galactose (D-Man/L-Gal) pathway, galacturonic acid pathway, gulose pathway and myo-inositol pathway were successively proposed[30, 31]. Among which D-Man/L-Gal pathway is the predominant source of VC in plant[32]. In conventionally grown tomatoes, concentrations of VC ranging from 1.00 mg/100 g to 63.8 mg/100 g of fresh weight, are mainly affected by genetic, maturation factors, season and cultivation conditions. Seasonal variations in VC levels have been described in relation to light intensity variations and sugar content. Despite sugars as a substrate for vitamin C biosynthesis, its content is not the limiting factor for VC accumulation in fruit[33]. Indeed, environmental conditions, especially light, are important factors regulating the VC concentrations in fruit via increased gene expression/enzyme activities of the VC biosynthetic pathway. Light promoted VC biosynthesis through up-regulating the expression of structure genes encoding the GDP-mannose pyrophosphorylase (SlGME1, SlGME2), GDP-mannose-3',5'-epimerase (SlGME) and phosphodiesterase (SlGGP). Among them, GME (GDP-D-mannose epimerases), which catalyze the synthesis of GDP-D-mannose, is the most important light sensitive regulator in the D-Man/L-Gal pathway[34]. But overexpression of GME alone through transient in tobacco or kiwifruit resulted in little change in VC content, which may be because the genes' promoter is not induced and requires the presence of additional cis-acting elements for their function[35]. GDP-l-galactose phosphorylase (GGP) catalyzes the first ascorbate-specific step in the pathway and others have nominated GPP as the key control gene in ascorbate biosynthesis (Fig. 3). And overexpression GGP in tomato using a strong constitutive promoter (35S promoter) have been shown to increase the content of VC[36]. L-GalDH oxidizes L-galactose to L-galactono-1,4-lactone using NAD+ as an electron acceptor. Compared with other enzymes in the D-Man/L-Gal pathway, L-GalDH activity is relatively high, it can rapidly convert foreign aid L-galactose into ascorbic acid resulting in the accumulation of VC.
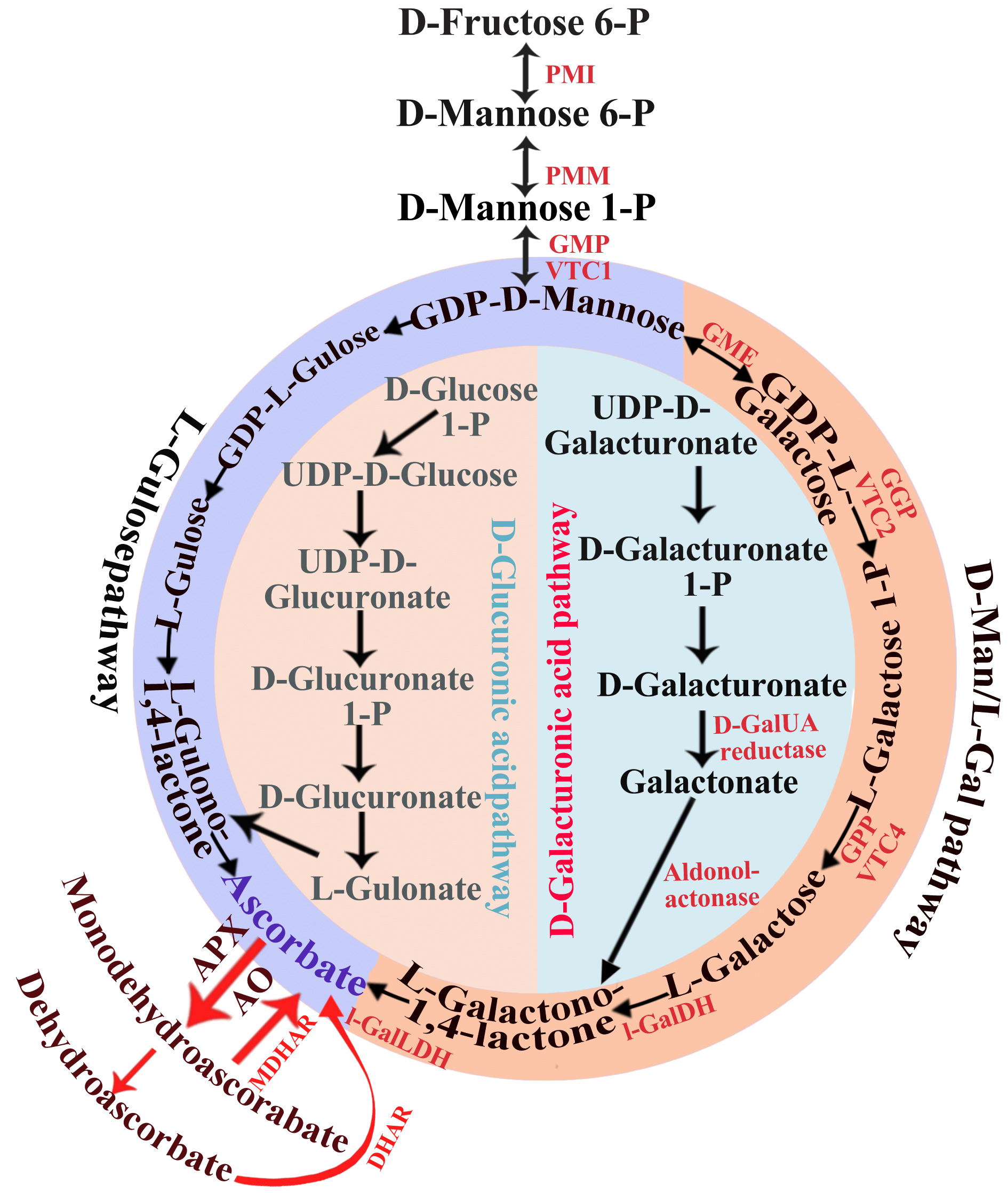
Figure 3.
Biosynthetic pathways of L-ascorbic acid in plants. PMI, mannose-6-phosphate isomerase; PMM, phosphomannomutase; GMP, GDP-mannose pyrophosphorylase (mannose-1-phosphate guanylyltransferase); GME, GDP-mannose-3′,5′-epimerase; GGP, GDP-L-galactose phosphorylase; L-GalDH, L-galactose dehydrogenase; L-GalLDH, L-galactono-1,4-lactone to produce ascorbate; AO, ascorbate oxidase; APX, ascorbate peroxidase; DHAR, dehydroascorbate reductase; MDHAR, monodehydroascorbate reductase; GPP, L-galactose-1-phosphate phosphatase.
The accumulation of VC in fruit is a balanced result of biosynthesis, oxidation, and recycling. Furthermore, many enzymatic/spontaneous oxidations and reductions effect the redox state of the ASA pool. ASA is converted to dehydroascorbate (MDHA) by ascorbate peroxidases (APXs), which contributes to cellular redox homeostasis, especially under stress conditions. DHA and MDHA can in turn be reduced back to ascorbate by dehydroascorbic acid (DHAR) and monodehydroascorbic acid (MDHAR), respectively[37]. And the expression of SlMDHAR was closely correlated with total ascorbate-ascorbate concentrations during ripening, higher levels of ascorbate could activate MDHAR activity[38].
Flavors
-
As a subjective phenotype, flavor integrates multiple sensory systems and is ultimately manifested at the brain level. Although the appearance and texture certainly affect the perceived taste, the underlying chemistry can be simplified as the process of the interaction between flavor substances in food and receptors for taste and smell in the human body[39]. Consumers generally believe that many modern commercial fruits have lost their unique flavor. With the pursuit of fruit yield and appearance quality in the past, this may have indirectly resulted in the loss of fruit flavor. In order to improve the flavor of fruits, researchers have made lots of effort. But the study of fruit flavors is difficult because it is a quantitatively inherited trait, and the metabolites contributing to flavor are greatly affected by environmental factors[40]. In tomato fruit, its flavor is the sum of sugars, acids (contributing to the taste) and many volatile chemicals (contributing to the smell). Improving the flavor of fruits is not just to increase the concentration of aroma substances, but also to explore the influence of the ratio of different compounds on the flavor of fruits.
Soluble sugars and organic acids
-
Sugars play an important role in the yield and quality of fruit. It can improve fruit sweetness and act as signal molecules and carbon energy sources. Generally, the sweetness of the fruit depends on the content and composition of soluble sugars that are mainly determined by genotype, ripening stage, and cultivation conditions. Fructose and glucose (hydrolysates of sucrose) were considered as the dominant soluble sugars in tomato fruits, which mainly accumulate during the red ripening stage[41]. There are three accepted ways of soluble sugar accumulation in fruits: (1) sucrose influx into the fruit via the vascular system, (2) sugar metabolism and the biosynthesis of starch as a transient storage form of carbohydrate, and (3) the breakdown of starch and intensive increases in the levels of hexose sugars[42]. Hence, promoting photosynthetic carbon fixation and sucrose hydrolysis in tomato plants may be two effective strategies to increase soluble sugar concentration in fruit. As shown in Fig. 4, the Calvin cycle is the primary pathway for photosynthetic carbon fixation in higher plants, which can provide precursors (triose phosphates) for the synthesis of sucrose. However, Ribulose-1,5-bisphosphate (RuBP) carboxylase/oxygenase (Rubisco) the key enzyme of this pathway, limit the capacity of carbon fixation due to its slow reaction rate and the properties cannot be discriminated well between CO2 and O2[43]. Some methods to improve Rubisco and to increase the rate of the regeneration phase have been reported, among which introducing carbon concentration mechanisms from Cyanobacterial to other higher plants is a very promising direction[43]. Moreover, due to the particular localization of Sedoheptulose-1,7-bisphosphatase (SBPase), regulating its activity to increase the photosynthetic carbon fixation of plants has also attracted extensive attention. Ding et al.[34] reported that overexpression of SBPase can significantly enhance accumulation of carbohydrates and confers tolerance to abiotic stress for tomatoes. Therefore, changing the activities of individual enzymes in the Calvin cycle may be a useful target for promoting photosynthetic carbon fixation in plants. As the end product of photosynthesis, sucrose needs to be transported from the source to heterotrophic sinks to the release of carbon and energy.
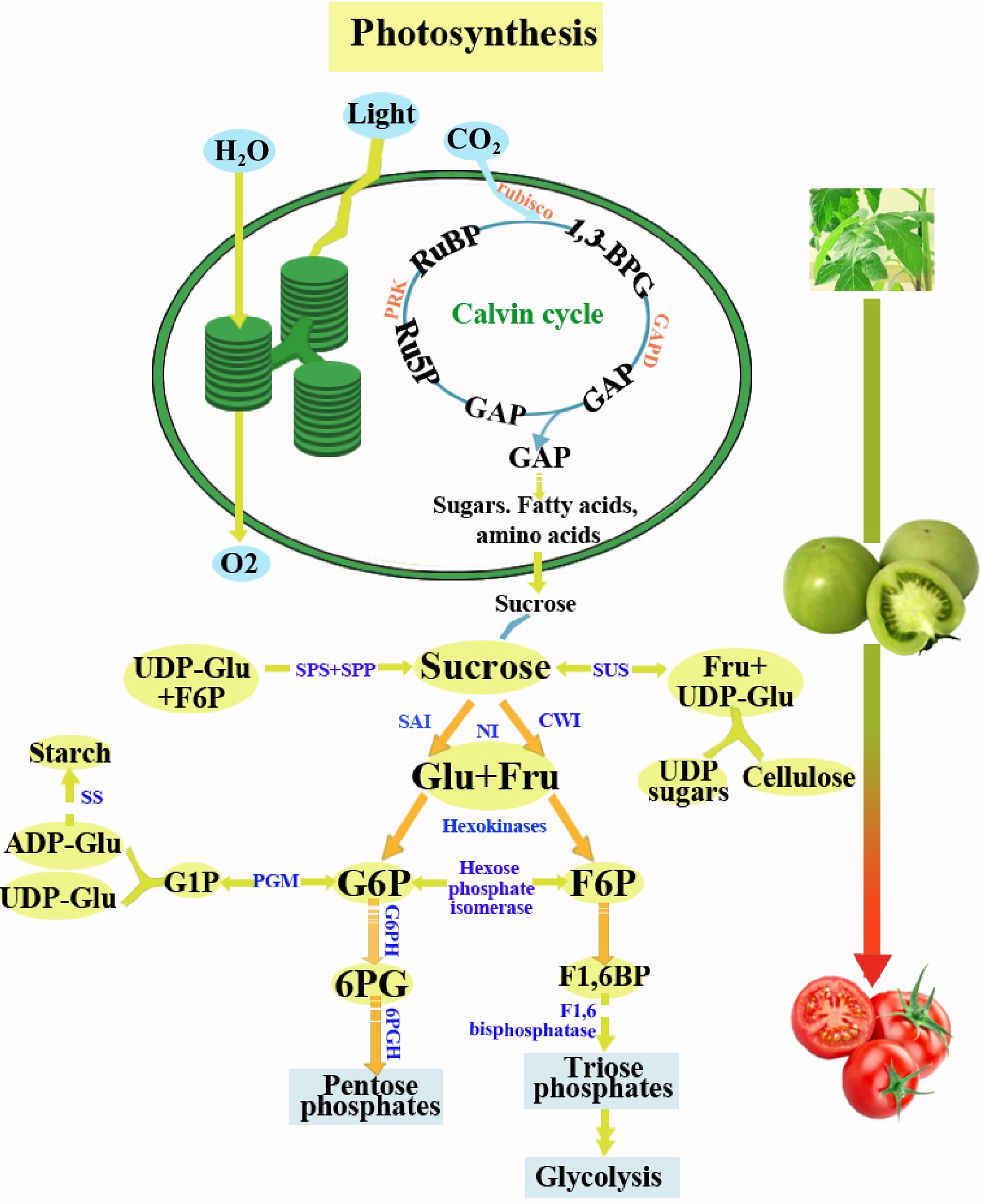
Figure 4.
Important intermediates in plant sugar metabolism. RuBP, ribulose-1,5-bisphosphate; 1,3-BPG, 1,3-bisphosphoglycerate; GAP, glyceraldehyde3-phosphate; Ru5P, Ribulose 5-phosphate; PRK, phosphoribulokinase; Glu, glucose; Fru, fructose; SAI, soluble acid invertase; NI, neutral invertase; CWI, cellwall invertase; G6P, glucose-6-phosphate; SUS, sucrose synthase; G1P, glucose-1-phosphate; F6P, fructose-6-phosphate; F1,6BP, fructose-1,6-bisphosphate; 6PG, 6-phosphogluconate; G6PH, glucose 6-phosphatedehydrogenase; 6PGH, 6-phosphogluconate dehydrogenase; PGM, Phosphoglucomutase; ADP-Glu, ADP-glucose; UDP-Glu, UDP-glucose; SS, Starch synthase; SPS, sucrose phosphate synthase; SPP, sucrose phosphate phosphatase;
During fruit ripening, the concentration of sucrose reduces and the content of glucose and fructose increase. This process has a strong correlation with the decrease in sucrose synthase (SUS), glucokinase and fructokinase activities and a high invertase activity. SUS and invertase are essential for carbon metabolism. SUS can transfer sucrose into fructose and UDP-glucose, and invertase can catalyze the hydrolysis of sucrose into glucose and fructose. Invertases are divided into acid invertases (cell wall invertase, CWI; soluble acid invertase, SAI) and neutral invertase (neutral invertase, NI)[44], and acid invertases were reported to be primary determinants of sucrose levels in tomato[45]. In addition to increasing the soluble sugar levels, the sweetness of the fruit can also be enhanced by enhancing the expression of sweet proteins. Brazzein, miraculin and monellin as the taste-modifying sweet proteins have been applied for developing transgenic plants to improve the sweetness and quality in tomatoes.
Organic acids represent the main carbon metabolism intermediates in plant cells, which give fruit a unique taste by changing the acidity. Their concentration in tomato fruits is highly dependent on the developmental stage and ripeness, and the degradation of them is regarded as an indication of fruit senescence[46]. Citric acid and malic acid are the most prevalent organic acids in tomato fruits and they have a significant contribution to flavor[47]. Organic acid synthesis begins with carboxylation of phosphoenolpyruvate (PEP) in the cytoplasm (Fig. 5), which is catalyzed by phosphoenolpyruvate carboxylase (PEPC). The products, oxaloacetate (OAA), can then be reduced to malate by cytosolic NAD-dependent malate dehydrogenase (NAD-cytMDH). The activity of NAD-cytMDH is generally related to malate synthesis in fruits. Overexpression of the apple MdcyMDH gene (encoding NAD-cytMDH) resulted in an increase in malate, fructose, and sucrose content, indicating its direct involvement in malate synthesis[48]. Synthetic malate and OAA can be converted into citrate in mitochondria or glyoxysome by the TCA cycle or glyoxylate cycle. Hence, this process affects the malate: citrate ratio of fruit cells. Malate oxidation by NAD-mtME however leads to the degradation of acidity. The activity of NAD-MDH and NAD-ME determine malate metabolism, both of which are regulated by the concentration of NADH and the pH[49]. For citric acid, it can be synthesized by citrate synthase (CS) through the TCA cycle in the mitochondrion and degraded by aconitase (ACO), γ-aminobutyric acid (GABA) shunt pathway and ATP-citrate lyase (ACL)[50, 51]. The activity of CS was reported to be positively correlated with citrate accumulation in watermelon, strawberry and citrus fruit, but transcriptomics and protein studies showed it is not responsible for the difference in citrate content between species[52]. In addition, Morgan et al.[53] used a biochemical analysis of a tomato introgression line and suggested that SlAco3b (encoding aconitase) may serve as a target for metabolic engineering to promote the accumulation of organic acids in tomato fruit. Overall, at present, it is difficult to determine the best way to engineer altered fruit carboxylic acid levels because a number of functional genes involved in the organic acid metabolism process are still unknown.
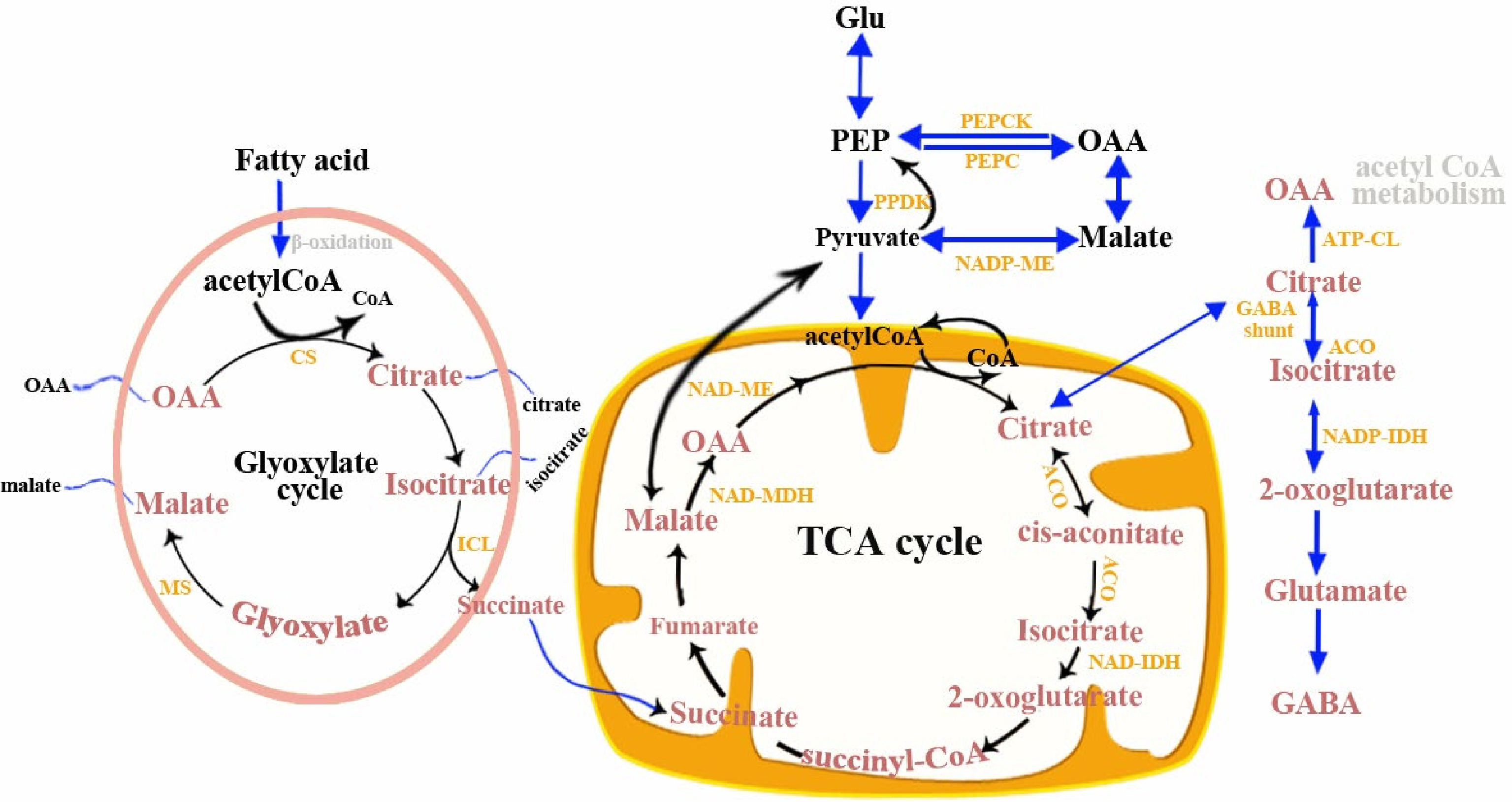
Figure 5.
Citric acid and malic acid metabolic pathways in fruit. TCA, tricarboxylic acid cycle; ACO, aconitase; ATP-CL, ATP-citrate lyase; CS, citrate synthase; ICL, isocitrate lyase; MS, malate synthase; NAD-MDH, NAD-malate; dehydrogenase; NAD-ME, NAD-malic enzyme; NAD-IDH, NAD-isocitrate dehydrogenase; NADP-ME, NADP-malic enzyme; OAA, oxaloacetate; NADP-IDH, NADP-isocitrate dehydrogenase; PDH, pyruvate dehydrogenase; PEPC, phosphoenolpyruvate carboxylase; PEPCK, phosphoenolpyruvate carboxykinase; PPDK, pyruvate orthophosphate dikinase.
Volatile compounds
-
The interaction between volatile compounds contributes to the aroma of the fruit. More than 400 volatile compounds have been identified in tomato fruits, but not all compounds contribute to the flavor of tomato. Only the compounds' odor activity with a value greater (OAV) than 1, are believed to be contributors to fruit flavor. About 30 volatile compounds were considered to contribute to tomato flavor, but their concentration varies widely among tomato varieties. However, the research of Tieman et al.[39] showed that not all these volatile compounds with OAV > 1 had a significant impact on consumer preferences. They associated volatile compounds with consumer preferences and pointed out that 16 volatiles were considered high-priority targets for genetic manipulation to improve tomato flavors. Based on the previous reports, we summarized some important volatile substances that affect tomato aroma and key genes that regulate their accumulation (Table 1), so as to enhance the aroma intensity in tomatoes through biotechnology.
Table 1. List of selected volatile compounds related to the flavor of tomatoes and the genes that regulate their accumulation.
Precursor Volatile compound Genes
(abbreviation)Full name Reference Fatty acid Hexanal TomloxC [72], [85] Trans-2-Heptenal 1-Penten-3-ol Hexanol Nonylaldehyde Heptaldehyde cis-3-Hexenol l-Penten-3-one Lsobutyl cyanide SlBCAT1 Tomato branched-chain amino acid aminotransferase [86] Trans-2-Pentenal SlscADH1 tomato short chain dehydrogenase-reductase [87] 3-Methylbutanol cis-3-hexenylacetate Hexylacetate Pentanol Isoleucine/Leucine 2/3-Methylbutanal SlscADH1 Tomato short chain dehydrogenase-reductase [87] Trans-2-Hexenal LeHPL Hydroperoxidelyase [88] Phenylalanine Methyl salicylate SlSAMT tomato salicylic acid methyl transferase [75] Phenylacetaldehyde LeAADC Aromatic amino acid decarboxylase [54] 2-Phenylethanol l-nitro-2-Phenylethane Benzylcyanide Benzaldehyde Methionine Eugenol Leucine 2-Isobutylthiazole lycopene β-Cyclocitral LeCCD4 Carotenoid cleavage dioxygenases [31] β-carotene β-ionone Carotenoid Pseudoionone LeCCD1B Carotenoid cleavage dioxygenases [31] ζ-caroteno Geranylacetone Carotenoid β-Damasccnonc Lycopene 6-Methyl-5-hepten-2-one Carotenoid Epoxy-B-ionone Geranial SlADH2 Tomato alcohol dehydrogenases [89] Neral Salicylaldehyde Trans-3-hexen-1-ol MiADH1 Mango alcohol dehydrogenase [90] Monoterpenoids Linalool SlGPPS, LeMTS1 Geranyl diphosphate synthases, linalool synthase [91] Most of the volatiles in fruits accumulate at the beginning of tomato ripening, and reach a peak at full maturity or shortly before[41]. Volatiles related to flavor are generally derived from essential nutrients, including fatty acids, amino acids, and carotenoids in plants[54]. Therefore, the factors that influence the accumulation of nutrients may also affect the aromatic volatiles in tomato fruits, such as some hormones (ethylene, abscisic acid, methyl jasmonate) and TFs (SNAC, MADS-box TFs, FUL, etc). At present, although some key genes have been confirmed, the regulation of volatile compounds in tomato fruits is still facing many challenges, mainly due to: (1) the complexity of the interaction between different volatiles, (2) the critical metabolic pathways for the synthesis of volatile compounds in tomato are not well understood. In the last decade, some genes involved in the biosynthesis of volatiles have been identified (Table 1). Fatty acid-derived volatiles are important contributors to consumer liking of tomato fruits[55]. And C5 and C6 volatiles (pentanol, 1-penten-3-ol, hexanal, cis-3-hexenol, etc.) are mainly synthesized through the lipoxygenase pathway, and the activity of lipoxygenase (encoded by TomloxC) is important for their biosynthesis[56]. The conversion of aldehydes to alcohols is also inseparable from the regulation of LeADH2. In addition, SpLIP1 (lipase gene) is the upstream regulatory gene of the lipoxygenase pathway, and can release fatty acids from the glycerol backbone. Its expression increased the concentration of fatty acid-derived volatiles[57]. For aromatic amino acid-derived volatiles, the first committed step catalyzed by the aromatic amino acid decarboxylase (AADS) is the limiting step for their synthesis[58]. Several of the important tomato aroma volatiles (2-phenylethanol, l-nitro-2-phenylethane, benzylcyanide) are derived from phenylalanine, they have fruity/floral properties that are considered to be a desirable flavor. And the expression of LePAR1 and LePAR2, which catalyze the conversion of 2-phenylacetaldehyde to 2-phenylethanol, significantly improved the concentration of 2-phenylethanol[59]. In particular, esters have a negative effect on tomato flavor, especially methyl salicylate, the concentrations of it should be decreased by suppressing the expression of SAMT (encoding O-methyltransferase). Carotenoids are not only important nutrients in tomatoes, but also an important source of fruit aroma. Carotenoid cleavage dioxygenase (CCD1A/B) linked the carotenoids to the many isoprenoid volatiles produced (β-ionone, neral, β-damasccnoncreleased etc) by cleaving the double bond in the carotenoid substrate[19, 60].
-
Increasing the accumulation of nutrients and flavors in tomato fruits are considered highly important in scientific study due to the impact on human health and market value. The accumulation of these important compounds (carotenoids, polyphenols, VC, flavors) is subjected to complicated regulation throughout the tomato life cycle in response to developmental and environmental signals. With the development of multi-omics technologies, the major flux-controlling step and rate-limiting enzymes involved in their metabolism pathway have been recognized. These steps and enzymes are usually regulated at many different levels by multiple regulatory factors. Therefore, understanding these key regulators and elucidating their molecular regulatory mechanisms can provide some insights for improving tomato fruit quality and accelerating the breeding process.
Carotenoids
-
As mentioned above, the steps catalyzed by DXS and PSY are the rate-limiting steps that determinate the rate of carotenoid biosynthesis. DXS mRNA accumulation in tomato is controlled by organ-specific and developmental signals, its expression can be regulated by transcription factor FRUITFULL (FUL), FUL1/2. FUL1/2, functions downstream of RIN and CNR in an ethylene-independent manner, which may not work at the onset of maturation, but alter the content of lycopene and its precursor cis-phytoene by regulating DXS expression in the later ripening process[61, 62]. However, whether FUL1/2 acts by directly regulating DXS expression, or by forming a complex with ripening factor RIN, still needs further study. Moreover, there is a crosstalk between the expression of DXS and PSY1, PSY1 activity positively mediates DXS mRNA levels[63]. Post-translational control is central to regulate MEP pathway activities, and DXS activity is tightly regulated at several post-translational levels. Recently, a post-translational regulation of PSY was reported in Arabidopsis. Clp protease (a major protease system in plastids) and ORANGE (OR) proteins were found to directly interact with the PSY, which maintain the homeostasis of PSY and positively regulate its protein abundance and enzymatic activity in the plastids[64]. Pulido et al.[65] found that the inactive forms of DXS can be specifically recognized by J20 (J-protein adapter) and delivered to Hsp70 chaperones for proper folding by interaction with ClB3, or degraded by the Clp protease via interaction with ClpC1.
The process catalyzed by PSY is the initial step in determining the rate of carotenoid production. PSY protein amounts and enzymatic activity are regulated by many genes (i.e. PIFs, HY5, RIN, WRKYs and MYB72). Meng et al.[66] silenced SlNAC1 (NAC transcription factors) by using an antisense system result in the higher transcription of SlPSY1 with higher carotenoid accumulation, whereas SlNAC4/9-silenced tomatoes have the opposite result[67]. Some members of the MADS-box transcription regulators (TAGL1, tomato agamous like 1gene; TAG1, tomato agamous 1 gene; CMB1, FYFL, MBP8) and RIN also effects the lycopene levels though direct regulation of PSY [68]. RNA-seq reveals MX1 protein can bind to the promoter of SlPSY1, SlPDS and SlZDS, overexpression of SlMX1 can enhance carotenoid content and drought resistance in tomatoes[69]. But the SlAN2 (from the R2R3-MYB family) is a negative regulator for the expression of SlPSY1, SlPDS and SlZDS, although it up-regulates the ethylene content. The activity of PSY is also regulated at several post-translational levels. Clp protease and OR were found to control the proteostasis of PSY, and further positively regulate its protein abundance and enzymatic activity in plastids. And the translation of PSY can be controlled by differential 5' UTR splicing in Arabidopsis. In addition, the expression of the PSY gene is tightly coordinated and controlled by feedback mechanisms, cis-carotenes and/or their cleavage products mediates negative feedback regulation. Therefore, for other enzymes involved in metabolic pathways, although not important rate-limiting enzymes, they are closely related to PSY gene expression and carotenoid diversity. Here, we also list some important factors that regulate their expression. SlcrtISO, expressed in all green tissues, was directly regulated by FUL1, FUL2 and RIN[70]. Agroinfiltration with Agrobacterium tumifaciens GV 3101 containing binary vector pRI 101(I) harboring CrtI gene and binary vector pRI 101(Y) harboring CrtY gene from bacterial sources could increase carotenoids in tomatoes[71]. The cyclization of lycopene is a central branch point in the carotenoid biosynthetic pathway, lycopene beta-cyclase (LCYB and LCYE) are important enzymes. The RNAi approach can be used to alternate the expression of SlLCYB and SlLCYE to control the ratio of β-carotene, lutein and other xanthopylls[66]. Furthermore, the presence of allelic variations could modify carotenoid profiles, such as t (enhanced prolycopene level) and Del (improved δ-carotene content) increase the diversity of lycopene isoforms that have the potential to increase the antioxidant properties of tomatoes[17].
Polyphenols
-
The accumulation of phenolics in tomato fruit is a complex trait controlled by various TFs and affected by the environment. t the transcription level, PAL is regulated by a variety of TFs, among which MYB proteins are one of the largest families of TFs in plants and involved in flavonoid biosynthesis. In tomatoes, SlMYB14 (R2R3-MYB protein) have been shown to recognize and bind to the promoter sequences of SlPAL and directly regulate the expression of SlPAL genes. Overexpression of SlMYB14 positively regulates the content of anthocyanins and flavonols[68]. And MYB TFs can interact with bZIP (basic leucine zipper) TFs to combine with ACGT elements in the promoters of flavonoid biosynthetic pathway genes thereby increasing flavanol production[72]. There are also other families of TFs detected in other plants (WRKY, KNOX, WDR, LIM) which could regulate the expression of PAL, but whether they directly target PAL in tomato fruits remains to be explored. Furthermore, PAL activity is controlled by post-translational regulation. It has been identified as a substrate for phosphorylation by a specific PAL kinase and a target protein for ubiquitination. And the disease resistance proteins, Pti4/5/6 with typical characteristics of TFs in tomatoes, were reported to not only regulate pathogenesis-related genes' expression, but also increase activities of PAL via mediate ubiquitin-proteasome pathway[73]. Anthocyanins as health-promoting compounds, are biosynthesized in a light-dependent manner in tomatoes. A variety of anthocyanins with special structures have been identified in tomatoes with the introgression of the Aft and atv genes that encode a functional anthocyanin activator (SlAN2-like) and nonfunctional version of the anthocyanin repressor SlMYBATV, respectively[74], but their content is still suboptimal. Therefore, improving the anthocyanin levels are the main objectives of many breeding programs. MYB-bHLH-WD repeat (MBW) complex were reported to directly regulate the synthesis of structural genes in the pathway of anthocyanin[68,72]. SlAN2 and SlAN2like as the MYB activator under light, participate in the formation of the MBW complex and induce anthocyanin accumulation. Vu & Lee[75] developed DNA markers of SlANT1 and SlMYBATV and found that tomato fruits with both dominant SlANT1 and recessive SLMYBATV alleles have high anthocyanin levels. And the expression of SlANT and some structural genes (CHS, DFR and ANS) can be activated by SlHY5. In addition, miRNAs are also regarded as tools to improve the quality of crops by mediating target mRNA degradation or by suppressing its translation. For example, sly-miR156d/e-5p, sly-miR160a, miR164a/b-5p, sly-miR168a/b-3p and so on have been found to participate in the regulation of the developmental program of tomatoes[7]. Some miRNAs: csn-miR5251, csn-miR477, csn-miR3444 and csn-miR156 have been reported to target genes of C4H, PAL, DFR and CHI in tea, and negatively regulate flavonoid metabolism[76]. But not much is known about their specific roles in the regulation of the vegetative phase of development of tomatoes. Hence, the anthocyanin biosynthesis pathway could be achieved through regulating tissue-specific expression of miRNA and master transcriptional regulators.
Vitamin C
-
At present, there are few reports of TFs regulating the genes involved in the D-Man/L-Gal pathway. In Arabidopsis, two TFs were found to function in the D-mannose/L-galactose pathway, AMR1: a negative regulator and AtERF98: a positive regulator[77,78]. There are two positive TFs in tomatoes, these are SlHZ24 and SlbHLH59[79]. The overexpression of TFs has little effect on VC content. However, controlling GGP translation would be a fast-response switch to control the VC level. The increasing experimental evidence supports GGP as a rate-limiting step of the D-Man/L-Gal pathway. GGP expression patterns are correlated with regulation of their promoters. Li et al.[80] compared the promoter structures of the gene encoding GGP in Actinidia, and suggested that the ArGGP promoter sequence contains both activation and repression regulatory elements. They constructed deletions of different regions of the GGP promoter to increase the transcription level of GPP, thereby increasing the concentration of VC. Previous reports indicated that overexpression of individual genes, VTC2, markedly increased VC levels in tomato[81]. In turn, VC participates in the regulation of GGP expression at the post-transcriptional level through feedback inhibition. An upstream open reading frame (uORF) is found in the rather long 5′-UTR region of GGP, its preferable translation led to competitive suppression of translation in the downstream region, while the disruption of its structure caused the cancellation of GGP feedback regulation and the increase of VC levels in Arabidopsis[82]. Therefore, GGP is now a fascinating target for improving VC accumulation in plants.
Flavors
-
Recently, in order to increase the content of soluble sugars in tomato, some genes that regulate invertase activity in tomato have been identified and characterized (Table 2). INH1, as a regulatory protein, inhibit the activity of CWIN at the post-translational level, and suppressing INH1genes expression could elevate fruit hexose levels[83]. CIF (belonging to the PMEI-RP family) as a vacuolar invertase inhibitor mediate sucrose metabolism and influences fruit ripening[84]. SlCIF1 and SlCIF2 were identified in tomatoes, and their silencing increased CWIN activity. However, their overexpression maybe promotes the accumulation of sugar. By analyzing the global expression profile of transgenic-SlCIF1 tomatoes using microarray, SlCIF1 can activate the expression of SlHSP17.7, which regulates expression of genes involved in sucrose metabolism and increases the fructose content in fruit. Furthermore, RIN not only affects the accumulation of flavonoids, but also directly regulates the transcription level of metabolic genes (VIN, hexokinase, fructokinase) by interacting with the CArG box element in the promoter[84]. Furthermore, SUT is an essential membrane protein for the accumulation of soluble sugars in fruits. There are three SUT genes (LeSUT1, LeSUT2, and LeSUT4) identified in the phloem of tomato, they are involved in the long-distance transportation of sucrose. The antisense inhibition of LeSUT2 significantly reduces soluble sugars (glucose, fructose, and sucrose) and results in a reduction of fruit yield[61]. In addition, expression of heterologous genes, such as SnRK1 (from Malus hupehensis Rehd.), GhDWF4 (from upland cotton), MdSWEET17 (isolated from apple) in tomatoes is also an effective strategy to increase sugar content in fruit[85−87]. Metabolism or membrane transport of H+ appear to correlate with the accumulation of organic acids. In the neutral or slightly alkaline cytoplasmic environment, malic acid and citric acid exist in the form of dianion malate and trianion citrate, respectively, and in acidic environments, these anions are protonated and pumped into the vacuole by V-ATPase and V-PPase. WRKYs, important TFs in higher plants, was reported to be able bind to the W-box of the VHA-B1 promoter and activate its expression. The activated VHA can regulate H+ transport, which can further synthesize organic acid molecules and improve the level of organic acids[88]. But, what molecular mechanism of VHA regulates the accumulation of organic acids still needs to be further explored. Recently, lots of pH-associated genes (named PH genes) have been found to be associated with fruit acidity and pH increase. A novel PH-like gene, CrMYB73 (encoding R2R3 MYB TFs in 'Ponkan' fruit), has been identified and used to improve the accumulation of citric acid in tobacco leaves[89]. Also, there are other TFs exhibiting strong correlation with organic acids levels during fruit ripening/postharvest fruit ripening/senescence, such as WRKY, AP2/ERF, NAC, bHLH35, bHLH113. Further experiments are needed to demonstrate their function and molecular regulation mechanism in tomato fruit.
Table 2. Overview of the genes that affect the accumulation of soluble sugar and organic acids in tomatoes.
Categories Genes
(abbreviation)Full name Function Reference Soluble sugar SlCIF1/2 Tomato cell wall invertase inhibitor gene It can inhibit the activity of CWIN at the post-translational level [60] INH1 Invertase inhibitor gene It can inhibit the activity of CWIN at the post-translational level SlHSP17.7 Tomato heat shock proteins encoding gene It can increase the accumulation of sugar [48] SlVPE1/2/3/4/5 Tomato vacuolar processing enzyme encoding gene It can promote acid invertases synthesis and improve hexose level [76] SlPEPCK Tomato phosphoenolpyruvate carboxykinase encoding gene It can regulate gluconeogenesis and enchance high sugar/acid ratio [77] LeLIN Fruit apoplastic invertase encoding gene Increase the activity of invertases [78] SlARF4 Abaxial encoding genes Inhibite the activity of AGPase and decreasing starch levels [79] TFT1, TFT10 14-3-3 protein encoding gene Down-regulated sucrose phosphate synthase activity [80] SlHXK1 Hexokinase encoding gene It can inhibit the accumulate of starch in tomato leaves [81] LeHT1/2/3 Tomato hexose transporter encoding gene Encode the hexose transporters and concentrate hexoses in storage parenchyma cells [82] SlSUT1 Tomato sucrose transporter encoding gene Phloem loader of sucrose in Solanaceae / RIN Ripening inhibitor Regulate sugar metabolism [61] SlMBP22 MADS-box subfamily, Bsister (Bs) genes It positively modulates the levels of starch and soluble sugar in tomato leaves [81] SlVIF Vacuolar invertase inhibitor gene It can inhibit the activity of VIN at the post-translational level [61] Organic acids SlPEPC2 Allosteric enzyme phosphoenolpyruvate carboxylase encoding gene Promote the decomposition of malic acid [83] TRXL1 Thioredoxin-like 1 encoding gene Activate NADP-MDH and increases malate [8] IDH3 Isocitrate dehydrogenases encoding gene Promote citric acid degradation [66] AST2 Aspartate aminotransferase encoding gene LeACS2 Ethylene biosynthesis gene Enhance the metabolization of malic and citric acid HMGCR1 3-hydroxy-3-methylglutaryl-CoA reductase encoding gene Inhibit the accumulation of critic acid MDH Malate dehydrogenase encoding gene Increase the content of malic acid PDC3 phosducin-like 3 encoding gene Decrease the content of malic acid SlAREB1 ABA-response element binding factor Mediate ABA signal and increase citric acid and malic acid level [84] AP2/ERF APETALA2/Ethylene Response Factor Regulate the organic acid levels [66] SlWRKYs Ethylene-responsive genes SNAC Stress-responsive NAC transcription factors The regulation of fruit flavor is a complex system with multiple interactions. At present, the complete metabolic network of volatile compounds has not yet been established and the related genes are likely to be quite large, which makes improving fruit flavor a big challenge. According to previous research, volatile compounds can be synthesized from primary metabolites (fatty acids, amino acids, carbohydrate) via various metabolomic pathways (Table 1). The oxylipin pathway was reported as one of the most important biosynthetic pathways, related to the generation of alcohols, aldehydes, and esters in many fruits[90]. As mentioned above, volatile compounds synthesis was strongly correlated with the transcription profile of TomloxC. RIN and MADS-box TFs (such as FUL1/2) can directly bind to the promoters of TomloxC to promote its expression. In addition, apocarotenoid volatiles are also present in tomato fruit, which can be generated by the action of SlCCD1A and SlCCD1B[30]. Because volatile compounds are mainly formed in late fruit ripening, the expression of the CCD gene may be regulated by transcription factors associated with fruit ripening. At present, the research on flavor substances mainly focuses on the identification of functional genes related to the formation of volatile compounds, and there are still few reports on the transcription factors that interact with them.
-
Fruit represents an excellent source of biologically active molecules important for the human diet, such as carbohydrates, vitamins and antioxidant compounds. Carotenoids, phenolics, vitamin C and flavors as indispensable metabolites are widely studied in plants due to their importance to industry, human health, and plant development. Over the years, enhancing the nutritional value of tomatoes has attracted increasing interest, not only to promote scientific research progress, but also to help solve the problem of insufficient human intake of nutrients. However, the difference in nutrients and flavors is a distinctive characteristic of cultivars within a species, regulated at multiple levels by environmental factors, metabolic regulation, and breeding practices. In order to steadily increase the content of nutrients and flavor substances in tomato fruit, some molecular techniques are used to optimize the breeding process, such as overexpression of RNAi, CRISPR-CAS9. Using these biotechnological tools for regulating important metabolic pathways requires an important prerequisite: a comprehensive metabolic regulatory network to be established. Hence, it is necessary to understand the metabolic pathways of these compounds and the interrelationships between different metabolic pathways, because the change of one pathway will have unexpected effects on one or more related pathways. The genes and enzymes involved in carotenoids, phenolics and vitamin C biosynthetic pathways have been identified and validated with the development of multi-omics technology. The expression levels of some rate-limiting enzymes in metabolic pathways are significantly regulated by some transcription factors, such as SlNAC1, FUL, C4H, SlMXI and so on. It is evident that regulation of promoting carotenoid accumulation in tomato holds enormous promise for satisfying both agricultural needs and scientific interests. Efforts should be made to facilitate the manipulation and improvement of agronomical and economical quality in tomato crops by explaining the precise mechanism of different carotenoid accumulation in each transgenic line and the upstream regulatory genes of some important TFs in the future. At present, much of our understanding of key active substances in metabolic pathways is from analyzing the transcriptional profiles of mutant plants. However, the polyphenol metabolism pathway has not been supplemented due to the redundancy of regulatory genes and failure to obtain mutants. Fortunately, an entire pan-species metabolic network of polyphenolic metabolism in tomatoes has been established by utilizing natural variation. The rate-limiting steps/enzymes (PAL, CHS, CHI) are gradually being understood, which serve as important targets for genetic engineering to increase polyphenol levels in fruit. Wild genotype is thought to have strong antioxidant activity, so identifying more potential functional genes and important regulatory networks from it is of great significance for improving tomato quality. Regarding flavors in tomato fruits, although the metabolic network of these compounds are still a long way from being understood due to the complexity of their interactions, the high demand for tastier tomato has triggered increasing interest in regulating of the accumulation of flavor compounds. The taste of tomatoes can be improved by adjusting the sugar and acid content, and the ratio between them can be adjusted by regulating the gene expression of SlCIF, SlVPE1, SlPEPC, IDH3 and so on. But our knowledge of regulation of the pathways is still rudimentary and the optimal concentrations for volatiles are not yet defined. Once these optimal concentrations are determined, they should be achievable by transgene or introgression of specific alleles.
Overall, the promotion of the accumulation of important compounds is mainly through the regulation of their biosynthetic and degradation pathways, so control of the activity of rate-limiting enzymes in the pathways is the key to controlling the flux of compounds. TFs, as key regulators of target genes, have been extensively studied. With the rapid expansion of the molecular toolbox, the increasing number of TFs will be recognized in the future, but the regulatory mechanisms behind them has not be well understood and needs to be further explored. Moreover, the regulation of target genes is not limited to the study of TFs, and some small RNAs are gradually entering people's field of vision. As RNA molecules affect all steps of gene expression and regulation, miRNA (sly-miR156a-c, sly-miR156d/e-5p, sly-miR396a/b-5p, sly-miR171a and so on) can change the tomato characteristics of floral abscission, fruit shape, cell number, etc, and can also interact with lncRNA to mitigate different stress conditions as well as being involved in tomato fruit ripening and the ethylene pathway. There are however few reports on the regulation of ncRNA on the accumulation of nutrients and flavor substances in tomato fruits.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lu Y, Zhu H. 2022. The regulation of nutrient and flavor metabolism in tomato fruit. Vegetable Research 2:5 doi: 10.48130/VR-2022-0005 

The regulation of nutrient and flavor metabolism in tomato fruit
- Received: 19 October 2021
- Accepted: 03 March 2022
- Published online: 29 April 2022
Abstract: The accumulation of nutrients and flavors are important factors that determine the consumption of tomato fruit. Carotenoids, polyphenol and vitamin C belong to antioxidant molecules and have been proven to regulate several biochemical processes and prevent many chronic diseases. In recent years, significant progress has been made in our understanding of the biosynthetic pathway and regulatory networks of these compounds in tomatoes. Numerous genes have been determined which promote the accumulation of these nutrients, such as SlNAC1, SlCCD, SlHY5. Meanwhile, since the mechanisms that control nutrient metabolism during fruit ripening are systematic and sophisticated, there are still many unknown factors that affect their content, which need to be further explored. In addition, not only are nutritional properties important, but flavor profiles play a core role in the market for tomato production. However, it is generally accepted by consumers that many tomato varieties have declined nutrient content and little flavor with people's pursuit of fruit yield and size. Hence, improving their levels are the main objectives of many breeding programs. In order to effectively enhance the nutritional and flavor quality of tomato fruits, we reviewed the critical steps in metabolic pathways of these important nutrients and flavor compounds, summarized key regulators for controlling the accumulation of these compounds, and hope to offer valuable candidate genes for the production of high-quality tomato fruits.
-
Key words:
- Tomatoes /
- Metabolisms /
- Accumulations /
- Flavor /
- Nutrition