









-
Shoot branching determines the architecture and yield of crops. In non-heading Chinese cabbage (NHCC), it is known as tillering, a special mode of shoot branching.
Lateral shoot development consists of two stages: the formation and outgrowth of axillary buds. Axillary buds are derived from axillary meristems (AMs) which are located in leaf axils. Leaf axil meristematic cells undergo three phases to form axillary buds: the maintenance and activation phase are controlled by SHOOTMERISTEMLESS (STM) and the emergence phase is characterized by the establishment of the WUSCHEL (WUS) - CLAVATA3 (CLV3) feedback loop[1]. LATERAL SUPPRESSOR (LAS) and CUP-SHAPED COTYLEDON 2 (CUC2) are putative regulatory hubs based on the abundance of transcription factors that bind to their promoters[1]. Arabidopsis las mutants are not able to form lateral shoots during vegetative development[2]. Lateral suppressor (Ls) and MONOCULM 1 (MOC1) are orthologs of LAS in tomato (Solanum lycopersicum) and rice (Oryza sativa). They encode GRAS family transcription factors and have a similar effect on shoot regulation[3,4].
Axillary buds remain dormant or develop into branches after forming. Axillary buds dormancy or activation are regulated by multiple factors including genes, environment and phytohormones. In rice, TEOSINTE BRANCHED1 (OsTB1)/FINE CULM 1 (FC1) represses axillary buds outgrowth but does not affect axillary buds formation[5]. In other words, OsTB1/FC1 works at the second stage of lateral shoot development. BRANCHED1 (BRC1), the ortholog of OsTB1, engages in multiple regulatory pathways and acts as an integrator of branching signals within axillary buds[6]. BRC1 encodes a TCP (teosinte branched1, cycloidea, proliferating cell factors) transcription factor which contains a basic helix-loop-helix (bHLH)-type DNA binding motif named TCP domain and a R domain rich in polar residues (arginine, lysine and glutamic acid)[7]. BRC1 prevents axillary bud outgrowth and its mutation leads to rosette branches increasing in Arabidopsis[6]. The function of TB1/BRC1 seems conserved to regulate lateral branches outgrowth in monocot[5,8,9] and dicot[6,10−12] plants.
In non-heading Chinese cabbage, BcBRC1 (named BcBRC1b here) can respond to the treatment of GR24 and 6-Benzylaminopurine (6-BA) which are synthetic analogs of strigolactone (SL) and cytokinin respectively[13]. It suggests BcBRC1b might be also located in the downstream of SL and cytokinin and be involved in tillering regulation. However, the function of its paralog, BcBRC1a, remains unknown. In this study, we cloned BcBRC1a from non-heading Chinese cabbage and preliminarily analyzed its function.
-
In NHCC, TCP transcription factor family contained 37 members with a TCP domain (Supplemental Table S1). To identify genes from NHCC most similar to BRC1, a neighbor-joining (NJ) phylogenetic tree based on the full-length protein sequences of B. campestris and A. thaliana TCP family was constructed. The TCP family could be divided into two subfamilies, class I (PCF) and class II. The basic domain of class I has a four-amino-acid deletion compared with Class II[14]. Class II contained two clades, CYC/tb1 and CIN. BcBRC1a (BraC03g038980/BcTCP17) and BcBRC1b (BraC01g038590/BcTCP2) in CYC/tb1 clade were closest to BRC1 (AtTCP18) (Fig. 1a).
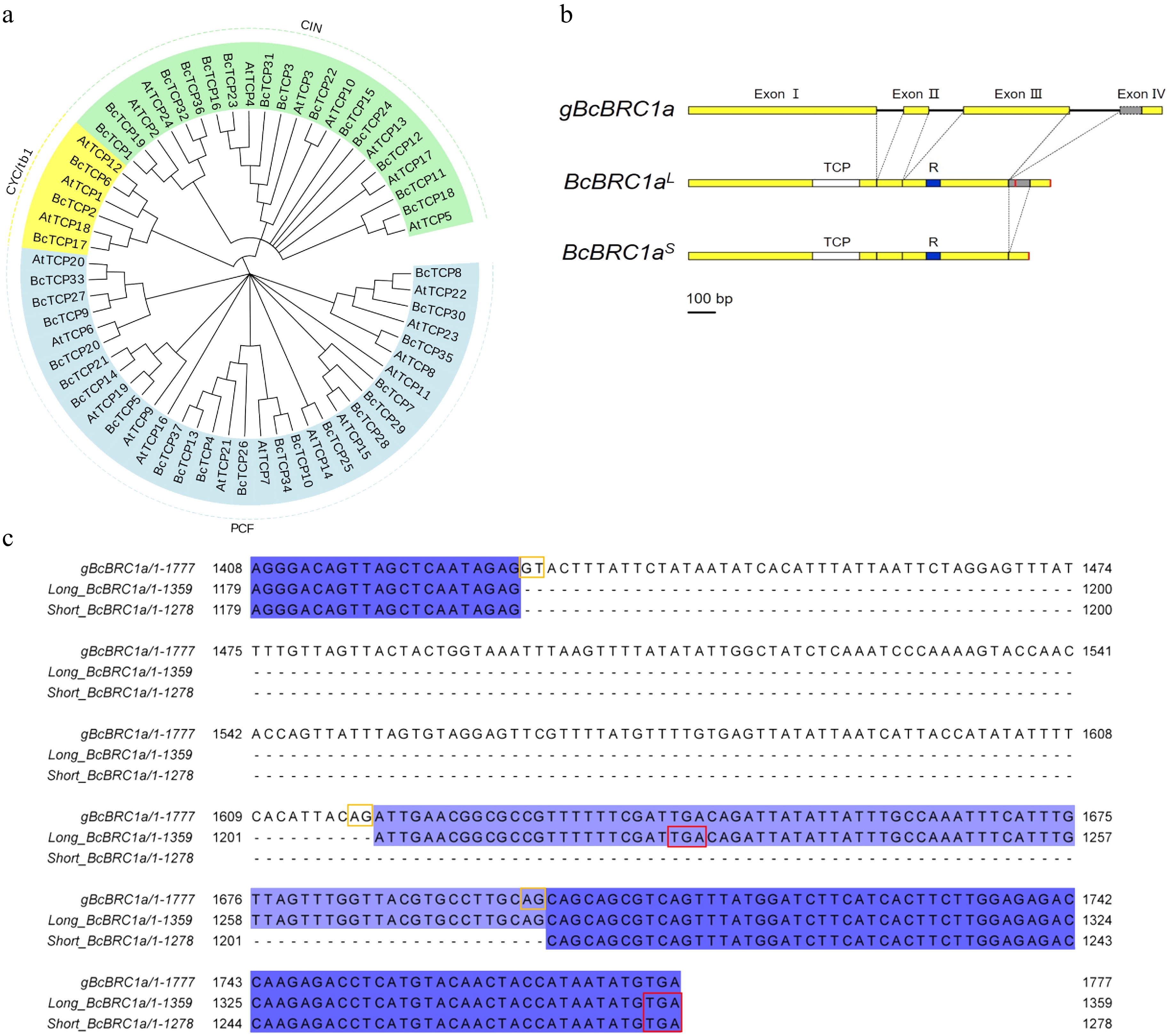
Figure 1.
TCP family analysis and BcBRC1a gene structure and partial sequence. (a) Phylogenetic tree showing relationships between TCP family transcription factors in B. campestris and A. thaliana. Blue, yellow and green backgrounds represent PCF, CYC/tb1 and CIN clades respectively. AtTCP18 is BRC1, BcTCP17 is BcBRC1a and BcTCP2 is BcBRC1b. (b) BcBRC1a gene structure. Yellow boxes represent exons and black lines represent introns. Grey box is alternative splicing region. TCP and R domain are filled with white and blue, respectively. The red color indicates termination codon. Scale bar = 100 bp. (c) BcBRC1a 3’ end sequence. Termination codons are bounded by red squares and alternative splicing sites are bounded by yellow squares.
To investigate the structure and function of BcBRC1a, we cloned it and found that BcBRC1a had two transcripts from alternative splice conforming to the GT-AG rule[15], a long one named BcBRC1aL and a short one named BcBRC1aS which had an alternate acceptor site (AA) in Intron III (Fig. 1b, c).
Interestingly, the extra segment of BcBRC1aL exon Ⅳ had a termination codon TGA which led to early termination of BcBRC1aL translation. BcBRC1aL had a 1227-bp open reading frame (ORF), while BcBRC1aS had a 1278-bp ORF. They could encode two proteins with identical TCP domain that is the main domain and R domain but different C-terminal domains (Fig. 1b). BcBRC1aS was the ancestral isoform (Supplemental Fig. S1).
Expression pattern of BcBRC1a
-
Non-heading Chinese cabbage cultivar 'Maertou' initiated tillering at the early vegetative stage, while 'Suzhouqing' began tillering at the late vegetative stage close to the reproductive stage[13]. To reveal expression pattern of BcBRC1a clearly, total RNA of non-heading Chinese cabbage cultivars 'Suzhouqing' and 'Maertou' from four tissues at 'Maertou' tillering stage (Supplemental Fig. S2) was extracted for qRT-PCR. The results showed that transcript level of BcBRC1a in 'Maertou' was higher than 'Suzhouqing' generally during the tillering period of 'Maertou' (Fig. 2a). Both in 'Maertou' and 'Suzhouqing', BcBRC1a was expressed mainly in leaf axil (LA) and secondly in shoot tip (ST), which suggested that BcBRC1a worked in the leaf axil primarily (Fig. 2a).
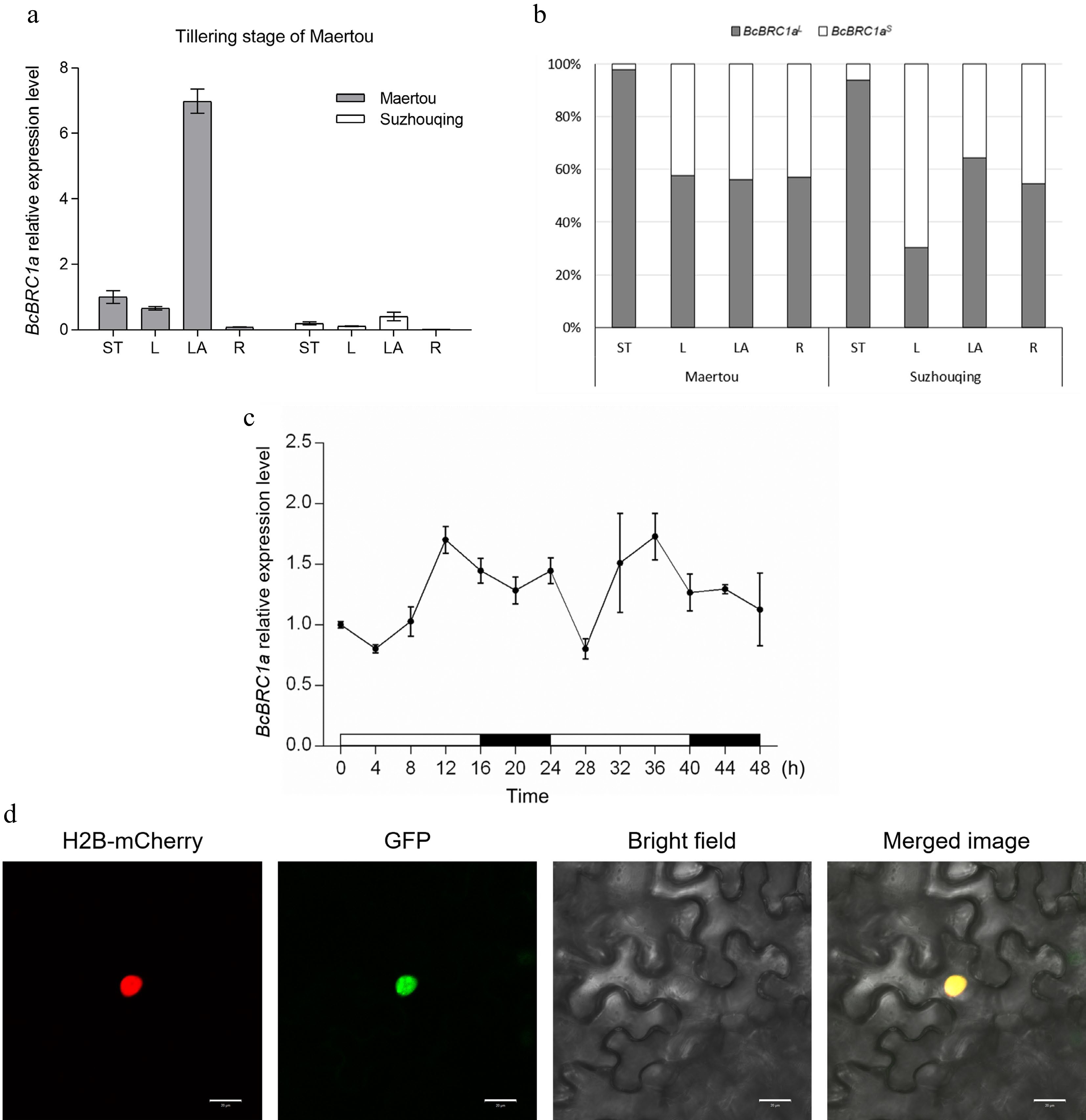
Figure 2.
Expression pattern of BcBRC1a in 'Maertou' and 'Suzhouqing' at the 'Maertou' tillering stage. (a) BcBRC1a relative expression level within four tissues. ST, shoot tip; L, leaf; LA, leaf axil; R, root. Error bars represent SE. (b) Proportion of BcBRC1aL and BcBRC1aS expression within four tissues. (c) Circadian expression of BcBRC1a. White bars represent daylight and black bars represent darkness. Error bars represent SE. (d) Subcellular location of BcBRC1aL. Scale bar = 20 μm.
In terms of two transcripts of BcBRC1a, BcBRC1aL played a major role in shoot tip (ST) and was expressed at a higher level than BcBRC1aS in almost all tissues except for leaf (L) of 'Suzhouqing' (Fig. 2b). So BcBRC1aL was chosen as a candidate for further study.
Previous studies showed that BRC1/TB1 expression had a daily oscillation like other TCP transcription factors[16,17]. So we detected BcBRC1a expression levels every four hours under long-day conditions (16 h light and 8 h dark) during two continuous photoperiods. We found that BcBRC1a was expressed rhythmically. BcBRC1a expression peaked at 12 h and 36 h and was lowest at 4 h and 28 h (Fig. 2c). Evening elements (EE) motif is one of the binding targets of CIRCADIAN CLOCK ASSOCIATED1 (CCA1) which is a key clock regulator and can prevent tillering in rice by positively regulating SL pathway gene expression including D14 and OsTB1[17]. Interestingly, EE motif of which sequence is (AA)AATATCT was also found in the promoter region of BcBRC1a (Supplemental Fig. S3).
BcBRC1aL is located in the nucleus
-
To analyze BcBRC1aL subcellular location, a 35S: BcBRC1L (CDS)-GFP recombinant plasmid was constructed and BcBRC1aL was transiently expressed in tobacco epidermal cell through the Agrobacterium-mediated method. Signals of GFP and mCherry (fused with a nuclear marker) merged in the nucleus (Fig. 2d), which indicated that BcBRC1aL was a nuclear location protein.
Ectopic expression of BcBRC1aL
inhibits the growth of Arabidopsis -
To analyse the function of BcBRC1a, 35S: BcBRC1aL (cDNA)-GFP was transformed into Arabidopsis (Supplemental Fig. S4). As shown in Fig. 3a, the crown diameter of BcBRC1aL overexpression (OE) lines were much smaller than WT during the vegetative period. OE14 growth was almost stagnant and BcBRC1a mRNA level of OE14 was significantly higher than other lines (Supplemental Fig. S5). During the reproductive period, primary rosette-leaf (R1) branch numbers of OE14 and OE17 decreased significantly (Fig. 3b, c). The phenotypes of BcBRC1aL transgenic lines illustrated that ectopic expression of BcBRC1aL at least inhibited the growth of Arabidopsis and probably inhibited its shoot branching.
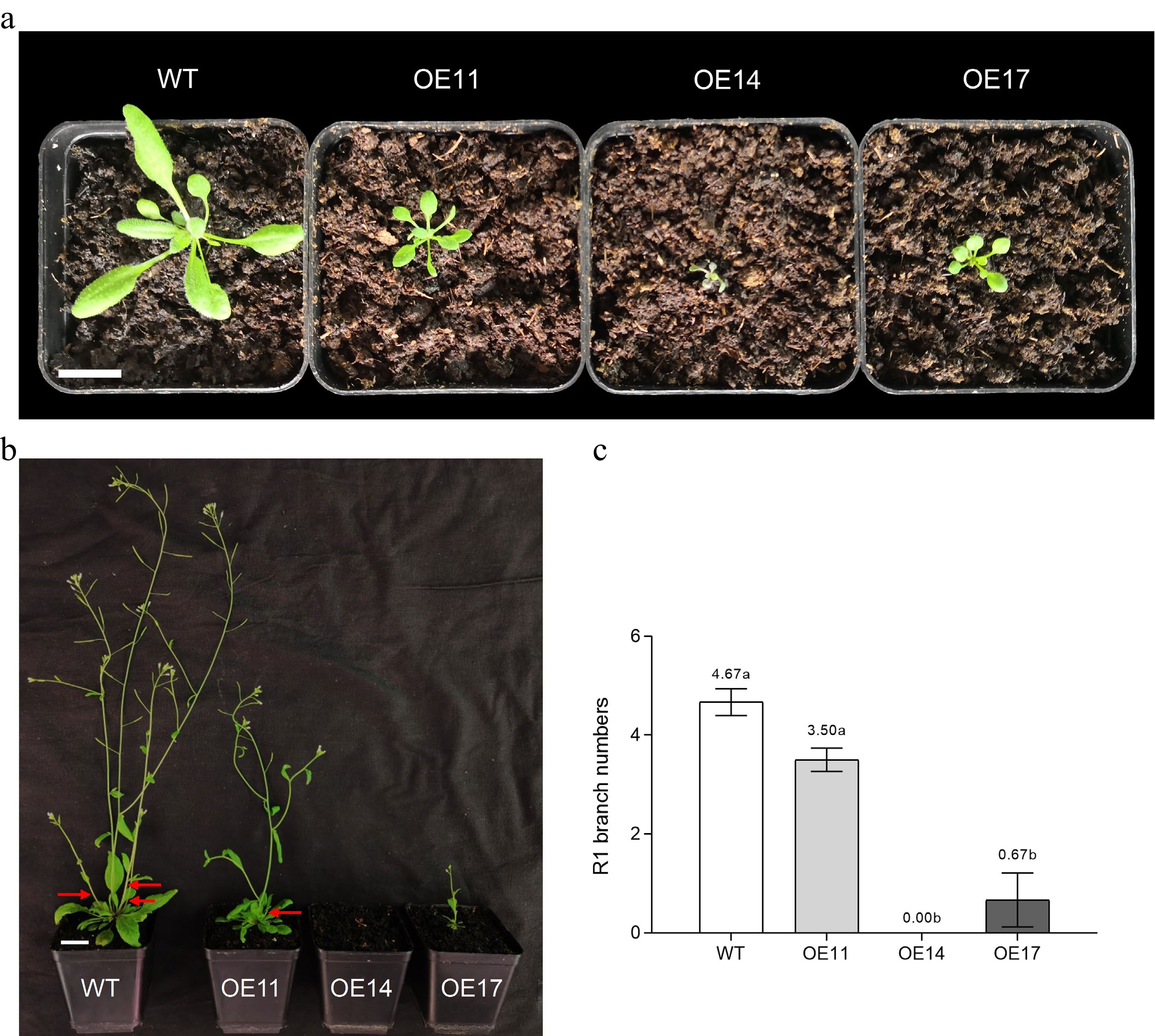
Figure 3.
Overexpression of BcBRC1aL in Arabidopsis (T2 lines). (a) Phenotype during vegetative period. Scale bar = 2 cm. (b) Phenotype during reproductive period. Red arrows indicate primary rosette-leaf (R1) branches. Scale bar = 2 cm. (c) R1 branch numbers of Arabidopsis transgenic lines. Three biological replicates, α = 0.05.
Silence of BcBRC1a enhances tillering of 'Maertou'
-
To reveal the function of BcBRC1a more reliably, VIGS experiment was conducted in 'Maertou' (Fig. 4a). In pTY-BRC1-1/2 plants, BcBRC1a mRNA level reduced by 88.6% and 79.7%, respectively, which meant that the experiment was effective (Fig. 4b). In addition, tiller numbers of pTY-BRC1-1/2 plants significantly increased compared with pTY plants (Fig. 4c). The result showed that BcBRC1a could prevent the outgrowth of tillering.
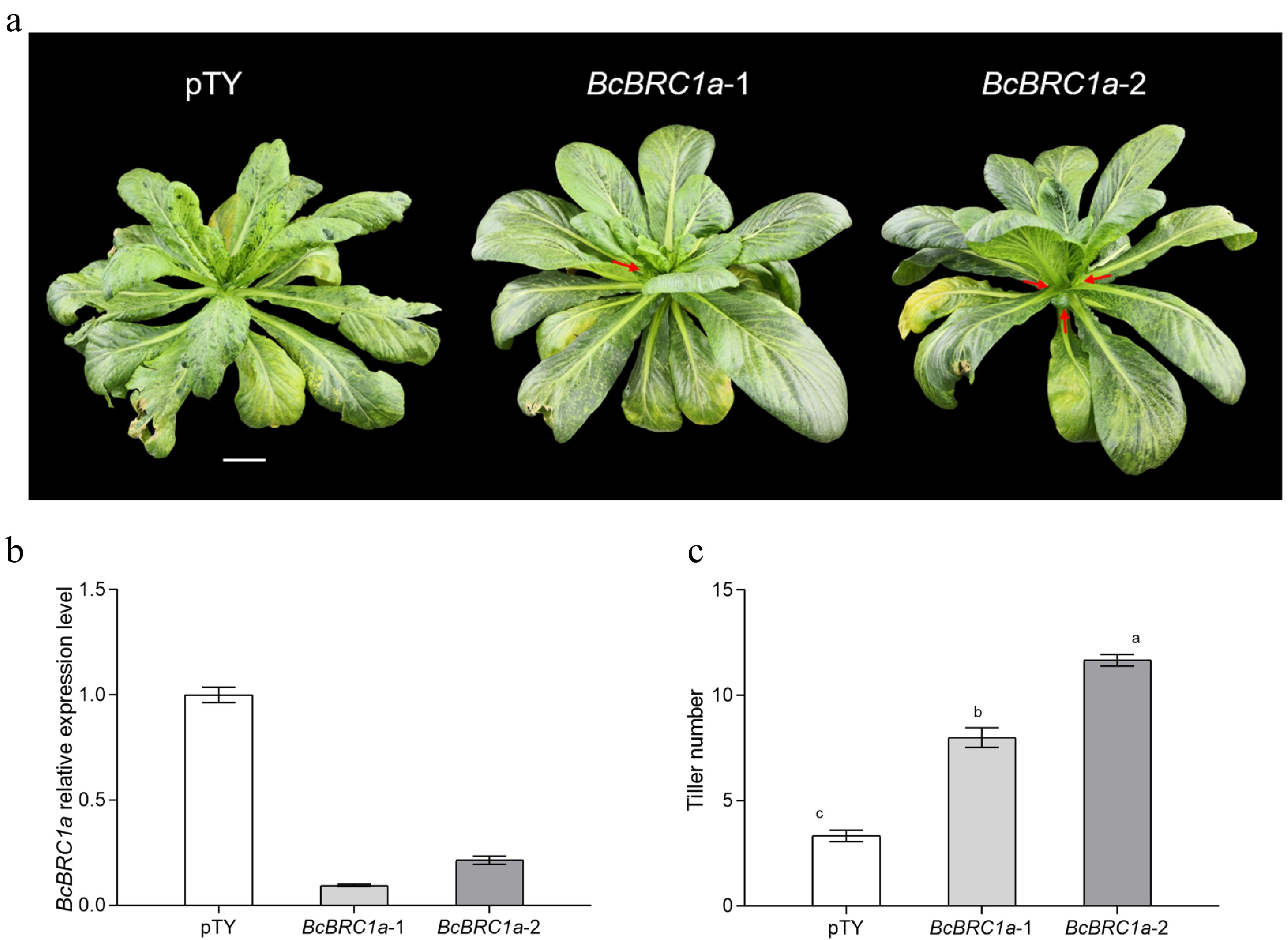
Figure 4.
BcBRC1a silencing in 'Maertou'. (a) Phenotype of ‘Maertou’ after silencing BcBRC1a. Scale bar = 2 cm. Red arrows indicate tillers. BcBRC1a relative expression level (b) and tiller numers (c) of BcBRC1a silencing plants. Three biological replicates, α = 0.05.
BcBRC1a responds to decapitation
-
Dormant axillary buds can be activated by decapitation due to the breakdown of apical dominance.
Although tillering buds of 'Maertou' would elongate constantly under suitable conditions, their elongation was accelerated after removing the stem tips (Fig. 5a), while axillary buds of 'Suzhouqing' were stimulated from dormancy to activation (Fig. 5b). Tiller numbers and length of decapitated 'Maertou' and 'Suzhouqing' were significantly larger and longer than intact ones (Fig. 5c, d). However, decapitation could not stimulate the nodes which could not have had axillary buds to generate tillers. In other words, decapitation only had an effect on axillary buds outgrowth but not on formation.
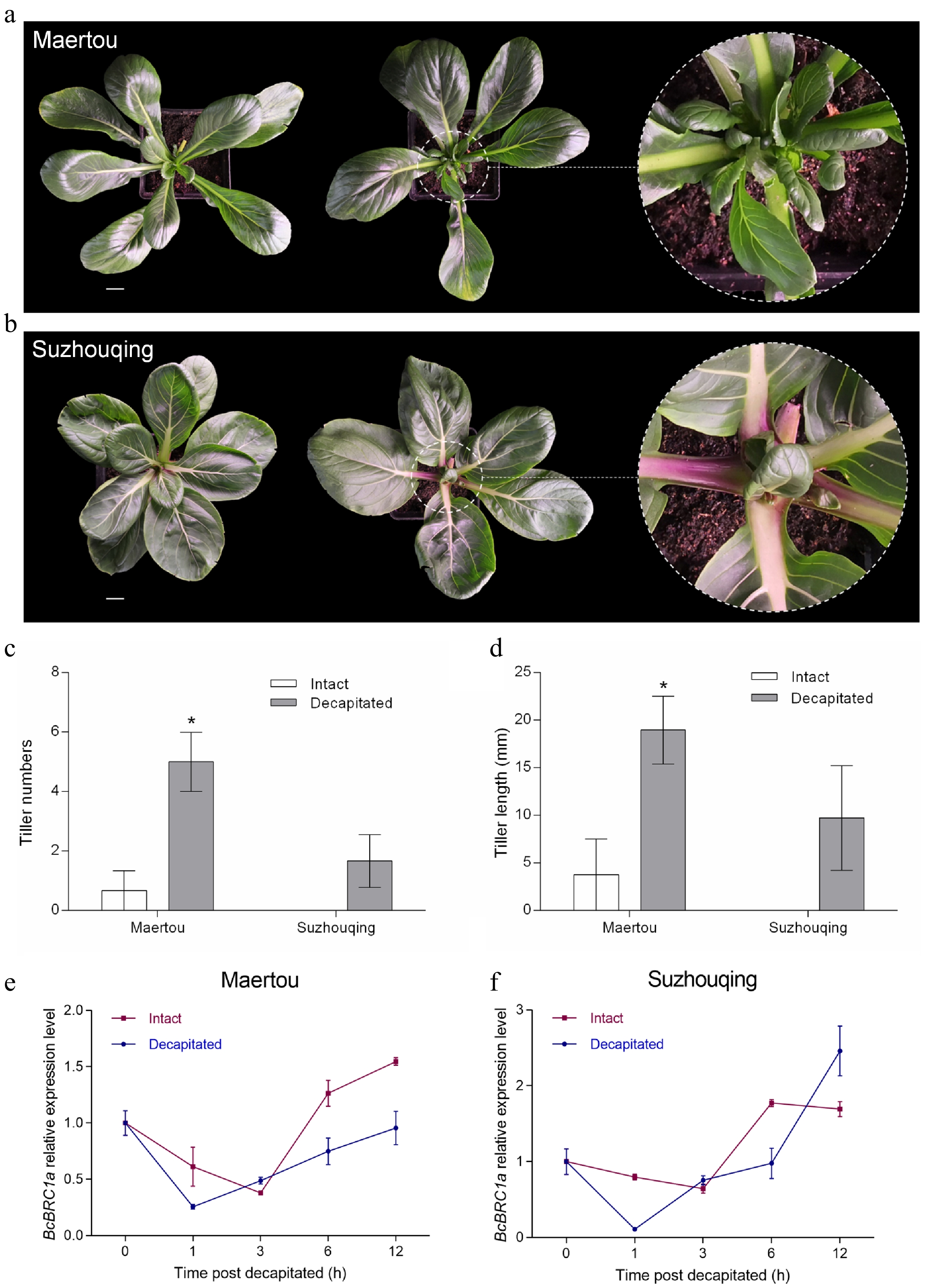
Figure 5.
Decapitated treatment for NHCC. ‘Maertou’ (a) and ‘Suzhouqing’ (b) tillering phenotype after 10 d of decapitation. Scale bars = 2 cm. Left, intact plant; right, decapitated plant. Tiller numbers (c) and length (d) of ‘Maertou’ and ‘Suzhouqing’ after 7 d of decapitation. Error bars represent SE (the same below). BcBRC1a expression in ‘Maertou’ (e) and ‘Suzhouqing’ (f) responds to decapitation. * represents P < 0.05.
To investigate whether decapitation would influence BcBRC1a expression, we conducted qRT-PCR to detect BcBRC1a relative expression level. In the control (intact) group, BcBRC1a transcript abundance decreased within 3 h and then increased presenting a similar trend in two cultivars (Fig. 5e, f). BcBRC1a was immediately down-regulated within 1 h after decapitation compared with the control in both 'Maertou' and 'Suzhouqing' and the down-regulation was stronger in 'Suzhouqing' than in 'Maertou' (Fig. 5e, f). After 1 h, BcBRC1a expression had a rebound in decapitated groups but the overall level was lower than intact groups (Fig. 5e, f). Therefore, decapitation could result in down-regulation of BcBRC1a.
Together, these results suggested that BcBRC1a responds to decapitation and the increase of tillers caused by decapitation in NHCC might be associated with the reduction of BcBRC1a expression.
-
Branching/tillering is an important agronomic trait. In many crops, multiple branches/tillers are usually regarded as an adverse trait for crop yields. However, in NHCC which is harvested mainly for leaves, multiple branches/tillers contribute to the increase in yield. The function of TB1/BRC1 has been studied in many species, which is the hub regulator of shoot branching and related to the dormancy of axillary buds.
In this study, we identified two orthologs of BRC1 in NHCC and cloned BcBRC1a. BcBRC1a was active in leaf axils at vegetative stage of 'Maertou' (Fig. 2a), which is consistent with previous studies[5,6,10]. The circadian clock plays an important role in regulating the growth and development of plants. CCA1 is a key clock regulator and the overexpression and repression of OsCCA1 reduce and enhance tiller numbers in rice[18]. BcBRC1a was expressed rhythmically (Fig. 2c) probably due to OsTB1/BRC1 acting as the target of CCA1[17,19,20]. Therefore, in NHCC, BcBRC1 expression may also be regulated by CCA1 in a circadian-dependent manner like in rice. It is worth investigating how circadian clock is related to tillering in NHCC and verifying whether CCA1 binding to the promoter of BcBRC1a in the following experiments.
Alternative splicing gives proteins much more diversity than genes[21]. There was an alternate acceptor site in Intron III of BcBRC1a, which made BcBRC1a generate two transcripts and proteins (Fig. 1b, c). The alternative splicing phenomenon of BRC1 was also detected in tomato (S. lycopersicum) and potato (Solanum tuberosum)[10,11]. Splicing of intron I causes a frameshift, which leads to short SlBRC1a and StBRC1a have a divergent C-termination (H domain) from long SlBRC1a and StBRC1a. StBRC1aS can affect StBRC1aL subcellular localization and reduce the transcriptional activity of StBRC1aL[11]. As for BcBRC1a, whether alternative splicing makes difference in its function still needs further study.
The ectopic expression of BcBRC1aL promoted by 35S in Arabidopsis made the growth of transgenic lines seriously inhibited (Fig. 3a), which was similar with what was found in Arabidopsis and potato[11,16]. During the reproductive period, branch numbers of OE14 and OE17 were significantly smaller than WT (Fig. 3b, c). Besides, silencing BcBRC1a increased tiller numbers of 'Maertou' (Fig. 4a, c). The results indicated that BcBRC1a could inhibit tillering in NHCC.
It is worth noting that BcBRC1a transcript abundance was higher in 'Maertou' (with tillers) compared with 'Suzhouqing' (without tillers) (Fig. 2a), which seems inconsistent with its inhibiting effect. This may be interpreted as there being a BRC1-independent way to regulate tillering in NHCC like pea and Arabidopsis[16,22−24]. On the other hand, BRC1/TB1 only controls the outgrowth of axillary buds[5,6]. As for axillary buds formation, it is regulated by many other genes. It is likely that another tillering regulatory factor plays a decisive role in 'Suzhouqing'.
Sugars play a central role in early outgrowth of axillary buds and act prior to phytohormones[25]. In this study, it was found that BcBRC1a expression was down-regulated within 1 h after decapitation, which suggests that BcBRC1a may prevent sink strength of axillary buds. Previous research shows that BcBRC1b responds to exogenous GR24 and 6-BA[13]. It may have a synergistic effect with BcBRC1a because the dimerization of TCP transcription factors can affect their DNA binding activity[26−28]. But this assumption still needs experimental proof.
-
In non-heading Chinese cabbage, BcBRC1a has two transcripts, BcBRC1aL and BcBRC1aS. The proteins they encode have identical TCP domains and R domains but different C-termination domains. BcBRC1a expression is more active in leaf axil than shoot tip, leaf and root and is controlled by circadian rhythms. BcBRC1a is a negative regulatory factor for tillering in NHCC. Besides, BcBRC1a can respond to decapitation treatment. Our findings are a supplement for BRC1-like gene function in diversified species and lay a foundation for revealing internal tillering regulatory networks in non-heading Chinese cabbage.
-
The non-heading Chinese cabbage 'Maertou' and 'Suzhouqing' cultivars were used. The seeds were grown in plastic pots containing soil mixture in a plant growth chamber (16 h light and 8 h dark at 27 °C). Tobacco (Nicotiana benthamiana) and Arabidopsis wild type (ecotype Col-0) plants were grown in the plant growth chamber (16 h light at 24 °C and 8 h dark at 18 °C).
Sequence alignment and phylogenetic analysis
-
The sequences of B. campestris were acquired from NHCC001 genome[29]. The Hidden Markov Model profile (HMM) and BLASTP were used to search for B. campestris TCP family proteins taking O. sativa (
http://rice.plantbiology.msu.edu/index.shtml ) and A. thaliana (www.arabidopsis.org/index.jsp ) as references[14,30,31].The protein sequences of B. campestris and A. thaliana TCP family were aligned with MUSCLE in MEGA 7.0 using the default options[32,33]. The phylogenetic tree was constructed using the Neighbor-Joining (NJ) method[34]. We set 1000 bootstrap replications, p-distance model and complete gaps deletion and other options retained the default settings. The tree was polished with Evolview v2[35].
Cloning of BcBRC1
-
Total RNA of 'Maertou' was extracted with an RNA easy Mini Kit (Tiangen, Beijing, China) and the first strand of cDNA were synthesized via reverse transcription using a PrimeScript™ II 1st Strand cDNA Synthesis Kit (Takara, Dalian, China). The full-length BcBRC1a was amplified with PrimeSTAR® Max DNA Polymerase (Takara, Dalian, China) and BcBRC1a-F/R primers (Supplemental Table S2). The PCR product was inserted into the pEASY-Blunt Cloning vector (TransGen Biotech, Beijing, China) and sequenced by TSINGKE Biological Technology (Nanjing, China).
Vector construction for subcellular localization and Arabidopsis transformation
-
The pRI101-GFP vector was cut with NdeⅠ and BamHⅠ restriction enzymes and the linearized pRI101-GFP vector was obtained. The coding sequence (CDS) region and full-length cDNA of BcBRC1aL was connected with linearized pRI101-GFP vector using In-Fusion® HD Cloning Kit (Takara, Dalian, China) to construct a complete 35S:BcBRC1aL (CDS/cDNA)-GFP vector.
Subcellular localization
-
35S:BcBRC1aL (CDS)-GFP recombination plasmid was introduced into Agrobacterium GV3101 strain (Tolo Biotech, Shanghai, China). Agrobacteria were cultured overnight until OD600 reached 0.8 and injected into tobacco leaves. After 24 h dark and 24-36 h light culture, the leaves were cut into pieces to detect fluorescence under a confocal laser scanning microscope (Zeiss, LSM780, Jena, Germany).
Arabidopsis transgenic plants
-
35S:BcBRC1aL (cDNA)-GFP recombination plasmid was introduced into Agrobacterium GV3101 strain (Tolo Biotech, Shanghai, China). A. thaliana plants were transformed with Agrobacterium using the floral dip method[36]. Seeds of transgenic lines were screened on 1/2 Murashige and Skoog (MS) medium with 50 mg/L kanamycin and 16 mg/L timentin in the illuminatuin incubator (16 h 75 μmol·m−2·s−1 light and 8h dark at 25 °C) and the green seedlings were transferred into pots containing soil mixture in the plant growth chamber after 14 d. The phenotype of WT and T2 lines were analyzed.
Silencing of BcBRC1 via the virus-induced gene silencing (VIGS) technique
-
The pTY vector we used for VIGS is derived from Turnip yellow mosaic virus (TYMV). Two 40-bp specific DNA fragments were designed in the conserved coding region of BcBRC1aS and BcBRC1aL, and then they were reversely complemented to obtain two 80-bp palindrome sequences (5'-CTAGATGTCGCCAAAGAGTTGTTCGGCTTACAAGACATGCGCATGTCTTGTAAGCCGAACAACTCTTTGGCGACATCTAG-3' and 5'-CTAAAGGGACAAGAGATCGTAGGATGAGGCTCTCGCTAGATCTAGCGAGAGCCTCATCCTACGATCTCTTGTCCCTTTAG-3', BcBRC1a-1 and BcBRC1a-2) and form hairpin structures. The synthesis of hairpin structures and construction of pTY-BcBRC1a-1 and pTY-BcBRC1a-2 vectors were done by the GeneScript company (Nanjing, China).
The pTY and pTY-BcBRC1a-1/2 plasmids were imported into 'Maertou' using the method of Yu et al.[37].
Decapitation for NHCC
-
One month after germination, half of 'Maertou' and 'Suzhouqing' seedlings had their stem tips removed for phenotyping and quantitative real-time PCR and the other half were kept intact as a comparison. The tiller (length > 5 mm) numbers and length were counted and measured on the 7th day after decapitation. Three biological replicates were set. The t-test method was adopted to analyze the data.
Quantitative real-time PCR
-
Total RNA was extracted from NHCC and Arabidopsis with an RNA easy Mini Kit (Tiangen, Beijing, China) and reverse transcription was conducted with Hifair® III 1st Strand cDNA Synthesis SuperMix for qPCR (gDNA digester plus) (YEASEN, Shanghai, China). SYBR Green method was used for qRT-PCR with Hieff® qPCR SYBR Green Master Mix (High Rox Plus) (YEASEN, Shanghai, China). The following pairs of primers were used (Supplemental Table S2): RT-BcBRC1a-F/R, RT-BcBRC1aL-F/R, RT-BcActin-F/R, RT-AtActin-F/R. Three biological and technical replicates were applied. The results were calculated as the 2−ΔΔCᴛ method[38]. Ratio(BcBRC1aL) was the expression level of BcBRC1aL relative to BcBRC1a. Ratio(BcBRC1aS) was calculated by substracting Ratio(BcBRC1aL) from one.
${\rm{ Ratio}}(BcBRC1a^{L}) = \frac{\text{E(}{{BcBRC1a}}^{{L}}\text{)}}{\text{E}{(}{BcBRC1a}{)}}\times{100{\text{%}}} $ $ {\rm{Ratio}}(BcBRC1a^{S}) = 1 - {\rm{Ratio}}(BcBRC1a^{L}) $ This work was supported by the National Natural Science Foundation of China (32172562), and National vegetable industry technology system (CARS-23-A-16), and A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Information of TCP transcription factor family of NHCC.
- Supplemental Table S2 Primer Sequences.
- Supplemental Fig. S1 Alignment of C-termination of AtBRC1, BcBRC1aS and BcBRC1aL.
- Supplemental Fig. S2 Phenotype of ‘Maertou’ and ‘Suzhouqing’. Left, ‘Maertou’; right, ‘Suzhouqing’. Bar = 2 cm. The red arrows indicate tillers.
- Supplemental Fig. S3 Promoter analysis of BcBRC1a. The fragment is taken from 1988 bp before ATG. Evening elements (EE) motif is highlighted with yellow.
- Supplemental Fig. S4 PCR validation of Arabidopsis transgenic lines. M, marker; BC, blank control; PC, positive control.
- Supplemental Fig. S5 BcBRC1a relative expression level in Arabidopsis transgenic lines.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Long Y, Zhao T, Xu L, Zhang W, Huang F, et al. 2022. BcBRC1a is a negative regulator for tillering in non-heading Chinese cabbage. Vegetable Research 2:11 doi: 10.48130/VR-2022-0011 

BcBRC1a is a negative regulator for tillering in non-heading Chinese cabbage
- Received: 04 May 2022
- Accepted: 15 August 2022
- Published online: 31 August 2022
Abstract: Shoot branching is a decisive factor for crop yield. Molecular mechanism for regulating shoot branching (tillering) needs to be determined. Plenty of previous studies have illustrated that BRANCHED1 (BRC1) is a key integrator of shoot branching regulating signals. However, BcBRC1 function in non-heading Chinese cabbage (Brassica campestris ssp. chinensis) (NHCC) remains unknown. Here, we defined two BRC1 orthologs, BcBRC1a and BcBRC1b, from NHCC and focused on the BcBRC1a gene to describe its alternative splicing characteristic and structure. BcBRC1a was expressed rhythmically and mainly in leaf axils at the 'Maertou' cultivar tillering stage. BcBRC1aL encoded a nuclear location protein. Its ectopic expression caused Arabidopsis growth inhibition and silencing BcBRC1a led to increased tiller numbers in 'Maertou'. Removing the shoot tips of NHCC caused axillary buds to be released from apical dominance and BcBRC1a expression down-regulation. Our research determined that BcBRC1a acts as a negative regulator for tillering in non-heading Chinese cabbage and sets the foundation for further studies.
-
Key words:
- BcBRC1a /
- Tillering /
- Non-heading Chinese cabbage /
- Alternative splicing /
- Decapitation