













-
Production and care of ornamental plants usually receive less attention relative to other horticultural crops and therefore may suffer more from suboptimal growth conditions. It is vital for us to understand the development of ornamental plants at the biochemical level in response to environmental cues, which will lead to a healthier cultivation systems.
Growth of plants is inseparable from the supply of nitrogen (N)[1]. Nitrogen plays key roles in the formation of cells, tissues and organs as the primary constituent of proteins, nucleic acids, nucleoside phosphates, phospholipids, and more importantly, secondary metabolites[2]. Both inorganic and organic forms of N exist in plants and soils with ammonium and nitrate as the most important inorganic N for growth and development. Generally, ornamental plants prefer nitrate over ammonium[3]. Recently, several studies suggest that with the rise of atmospheric CO2 concentrations, nitrate reduction in C3 plants such as Arabidopsis and wheat is inhibited whereas ammonium utilization is unaffected[4]. This is problematic as this reduction in nitrate assimilation can lead to the decrease of N use efficiency and the accumulation of ammonium. And the latter will cause ammonium toxicity in plants via acidic stress[5] and impair shoot growth significantly which also affects the production of ornamental plants. Therefore, different forms of inorganic N should be assessed independently in horticultural crops including ornamental plants.
Plant roots can take up both ammonium and nitrate from soil[6]. The absorption of inorganic N is an active process that is controlled by plasma membrane-localized transporters in the root[7]. There are two nitrate uptake transporter families in higher plants, namely low-affinity transporter (LATS) NRT1/PTR and high-affinity transporter (HATS) NRT2[8,9], while ammonium uptake is mediated by the high affinity transporters of the AMT/MEP/Rh (AMT) subfamily[7]. In cells, the reduction of nitrate to ammonium requires two consecutive reactions. Firstly,
${\text {NO}_3^-} $ Generally, the assimilation of ammonium is far more energy efficient compared to nitrate assimilation[16]. However, nitrate is more available for plants due to its higher mobility[17]. Also, soil aeration status and soil pH value have effects on the availability of inorganic N. In well aerated agricultural soils, nitrate is the principal inorganic N source, whereas in undisturbed soils, ammonium is dominant. Plants grown in acidic soils take up more nitrate, while in soils with neutral pH values, they prefer taking up more ammonium. Meanwhile, different plant species will preferentially select specific N sources, thereby forming different N utilization and distribution mechanisms, which are mainly related to the long-term adaptation of plants to N environments.
The floral organ is one of the main nitrogen pools and its formation requires the regulation of multiple metabolic pathways. However, roles of N metabolism in flower development and function fulfillment are still poorly understood. Amino acid metabolism, a part of plant N metabolism, is of great significance to the development and function of floral organs[18]. In addition, amino acids are also important precursors for the synthesis of secondary metabolites, such as phenolics, alkaloids and other nitrogen-containing secondary substances. Studies have shown that the expression of the gene encoding 5-enolpyruvylshikimate-3-phosphate (EPSP) synthase in Petunia was associated with petal specificity, and the expression level was significantly increased during flower opening[19]. EPSP synthase is also a key enzyme in the shikimate pathway and is essential for the production of aromatic compounds that provide substances such as phenylpropanoid and indoleacetic acid for petal coloration and development[19,20]. Studies have shown that inhibition of EPSP synthase leads to a decrease in the level of aromatic amino acids, which in turn inhibits protein synthesis[21,22]. So far, besides studies related to amino acid transporters in various floral tissues[23,24], only a few studies have focused on amino acid metabolism in floral tissue[25]. The de novo synthesis of proline has been reported in floral tissues such as pollen grains[18]. Efforts should be made to dissect different forms of inorganic N on floral nitrogen metabolism, which will lead to a better understanding of the metabolic features in floral tissues.
In this study, we treated Petunia plants with three forms of inorganic N at the same concentration of total N including1 mM NH4NO3 (the control), 2 mM NH4Cl (NH4-N) and 2 mM NaNO3 (NO3-N) from the vegetative to the reproductive stage. Plant growth, 15N utilization and distribution in different tissues, petal color traits and petal amino acid profiles were thoroughly analyzed to compare effects of different inorganic N on floral development and establish unique roles of ammonium in floral tissues. Results are discussed in the context of establishing roles of different forms of inorganic N in the formation of ornamental traits including flower development and petal pigmentation.
-
Petunia × hybrida cv 'Mitchell Diploid' (MD) plants were used in this study. After approaching their 6−8 leaf stage grown in culture medium (Pindstrup Sphagnum peat moss: perlite: vermiculite = 3:1:1) under 16/8-h day/night cycles, temperature of 22/18 °C, light intensity of 150 μmol m−2 s−1, and 50% relative humidity, MD plants were then hydroponically cultured with the same and regular amount of inorganic N for one month but in three different forms, 1 mM NH4NO3 as the control (NH4NO3), 2 mM NH4Cl alone (NH4-N) and 2 mM NaNO3 alone (NO3-N). This experimental set-up had been tested to make sure that after one month of treatment, MD plants could reach their reproductive stage and produce flowers for sampling. The total N content at 2 mM was chosen because it was the regular amount of N used in nutrient solution. Other than inorganic N, the nutrient solution was prepared as the following formula: 0.67 mM NaH2PO4, 0.60 mM MgC12·6H2O, 0.30mM K2SO4, and 0.30 mM CaCl2·2H2O. The pH value of nutrient solution was adjusted to 5.8. During the treatment, nutrient solutions were replaced every 10 d and clean water was supplemented as needed.
Measurement of Petunia developmental phenotypes
-
After 30 d of hydroponic treatment with different forms of inorganic N, the plant height, branch number, flower number, leaf area, corolla area, and the fresh and dry weight of both above- and under-ground tissues in MD plants were measured. Plant height, the natural height of the main stem, was analyzed with a tape measure. Stem thickness was the width of the main stem near the root of the plant, measured with a vernier caliper. Considering that the Petunia plant was short, the number of branches longer than 5 cm was counted. Petals and leaves were randomly collected from plants and measured for their area via Image J software (
https://imagej.nih.gov/ij ). Tissue weights were determined using ME820 balance. A total of 3−4 biological replicates were analyzed for each parameter.15N isotope labeling and related analysis
-
In the first and second weeks of hydroponic treatment, 0.3% 15N-urea (Shanghai Research Institute, Shanghai, China), 15N natural abundance is 10.16%; Natural 15N abundance is 0.365%) solution was brushed on the same leaves. At the end of treatment, we collected roots, stems, leaves and flowers (three biological replicates) and dried them in ovens at 60 °C for 2 d to obtain the dry weight via a ME820 balance. The samples were ground with a mortar into fine powder and sieved with a 100-mesh sieve. Total N content in various tissues was measured via a Kjeldahl nitrogen determination apparatus. 15N abundance in a specific tissue (%) was determined by DELTA V Advantage isotope mass spectrometer (Thermo Fisher, MAT-251). The calculations of 15N absorption, 15N utilization rate and 15N distribution rate in each tissue were carried out according to the following equations:
Ndff (%) = (15N abundance in a specific tissue − Natural 15N abundance) / (15N abundance in 15N urea − Natural 15N abundance) × 100;
15N absorption (mg plant−1) = Ndff × Total N in a specific tissue;
15N utilization rate (%) = (Ndff × Total N in a specific tissue) / Total content of 15N applied to plants × 100;
15N distribution rate (%) = (15N absorption of a specific tissue / Total 15N absorption of 4 tissues) × 100.
In the equations, Ndff (%) is the percentage of N derived from 15N urea; total content of 15N applied to plants was 0.4893 g according to the amount of 15N urea used inthe label experiment.
Determination of amino acid content in Petunia petals
-
Petal tissues were collected at the early, middle and mature stages during the development of MD flowers and quenched with liquid nitrogen. Petal tissues (~100 mg in FW) were ground into a fine powder in liquid nitrogen and then extracted with 1.5 ml methanol/chloroform/water (4 °C). Norvaline was added to extracts as the internal standard. The extract was filtered with a 0.45 μm cellulose acetate filter and by aspiration with N2. Amino acid content was determined according to the LC-MS/MS method[26] with slight modifications. After re-dissolution with ddH2O, samples were injected into a Thermo Scientific TSQ Quantum Access MAX Triple Quadrupole Mass Spectrometer coupled with a Thermo Fisher Hypersil GOLDTM C18 column (product# 25002-102130).
Determination of Petunia flower color
-
CIELab coordinates including perceptual lightness (L*), redness/greenness (a*), yellowness/blueness (b*), chroma (C*), and hue angle (h°) of Petunia petals were measured for each treatment via a portable colorimeter (CR-10). Four biological replicates were analyzed.
Data analysis
-
Data were recorded and analyzed in Excel, and statistical analyses were performed using Data Processing System (DPS) Software[27]. Duncan's Multiple Range Test was performed in DPS software to determine the statistical difference with P value less than 0.05 between treatments. Clustering analysis, correlation analysis and principal component analysis were performed in R Studio Version 1.1.456 (
www.rstudio.com ) with R version 3.5.2 using gplots and pheatmap (https://cran.r-project.org/web/packages/pheatmap/ ) packages, and factoextra (http://www.cran.r-project.org/web/packages/factoextra/ ) packages, respectively. -
In this study, MD plants were treated hydroponically with the same level of inorganic N for one month but in three different forms, 1 mM NH4NO3 as the control (NH4NO3), 2 mM NH4Cl alone (NH4-N) and 2 mM NaNO3 alone (NO3-N). The concentration of total N for each treatment was set up at 2 mM as this is the regular level of N used in nutrition solutions. After one month of treatment, MD plants approached their reproductive stages and could produce a couple of flowers (Fig. 1a). We measured the total content of N in above- and underground tissues and confirmed that Petunia plants treated with NH4NO3, NH4-N and NO3-N had very similar content of total N, which were 4.0 ± 0.8, 4.0 ± 0.5 and 4.4 ± 0.2 g plant−1, respectively (Fig. 1a). Relative to the NH4NO3, NH4-N treatment only enhanced the dry weight (DW) of above-ground tissues whereas NO3-N treatment significantly increased the fresh weight (FW) and DW of above- and under-ground tissues (Fig. 1b, c). NH4-N and NO3-N treatments did not affect plant height (Fig. 1d) but increased the number of branches by 36.8% and 26.3%, respectively (Fig. 1e). NH4-N and NO3-N also significantly increased the number of flowers by 45.7% and 32.6%, respectively (Fig. 1f). However, NH4-N and NO3-N treatments greatly reduced the area of leaf and corolla (Fig. 1g, j) with NH4-N had more negative effects on leaf area than NO3-N treatment.
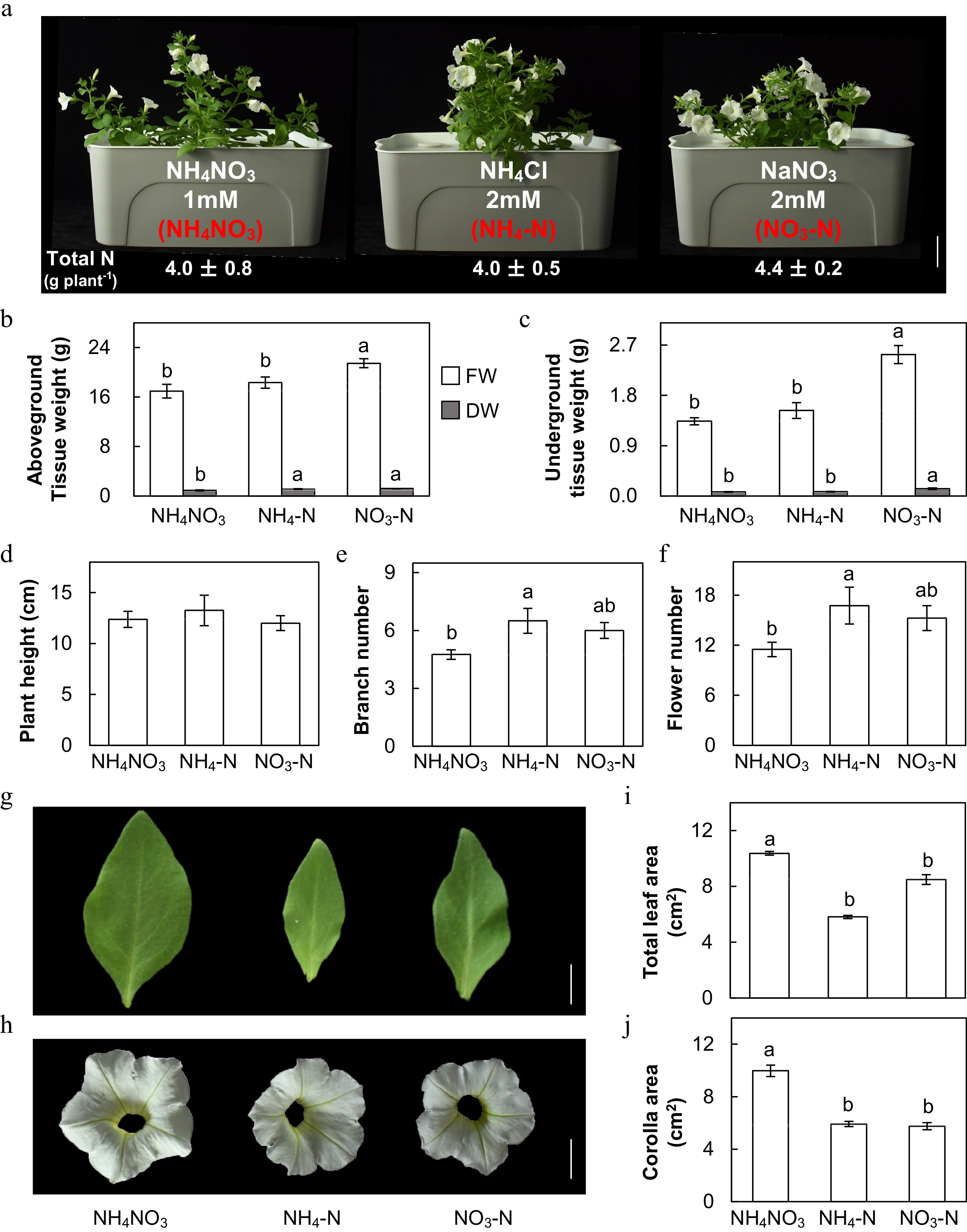
Figure 1.
Effects of ammonium-nitrate (NH4NO3), ammonium (NH4-N), and nitrate (NO3-N) on Petunia plants. (a) Petunia plants (MD) after treatment (scale bar = 5 cm). (b), (c) Fresh weight (FW) and dry weight (DW) of above- and under-ground tissues (SE, n = 4). (d)−(f) Plant height, number of branches and number of flowers measured for each treatment (SE, n = 4). (g), (h) Close-up view of the representative, fully expanded leaves and petals of fully-opened flowers of each treatment (scale bar = 1 cm). (i), (j) Total leaf area and corolla area measured for each treatment (SE, n = 4). Different letters above the bars indicate significant difference between treatments using one-way ANOVA test in SPSS software at P < 0.05.
15N uptake and distribution in Petunia differed with treatments at the tissue-level
-
In order to determine the effects of different N forms on uptake and distribution of N in Petunia plants, we performed a labeling experiment with 15N-urea. At first, N content in each tissue was measured and the results indicated that leaves had the highest content of N relative to roots, stems and flowers. Different forms of N did not affect N conent on the level of individual tissues (Fig. 2a). The absorption, ulitization rate and distribution rate of 15N in each tissue were then calculated based on formulas described in the Materials and Methods (Fig. 2b−d). Realtive to NH4NO3 treatment, NO3-N did not have effects on the tissue-level 15N absorption, utilization and distribution. In both NH4NO3 and NO3-N treatments, leaf-absorption of 15N was the highest among all tissues and roots, stems and flowers absorbed and utilized similar amounts of 15N. However, for the treatment of NH4-N, flower-absorption of 15N was significantly higher than that of roots. Therefore, in NH4-N, both 15N utilization and 15N distribution rate in flowers were significantly higher than those in roots. More importantly, among the three treatments, NH4-N had the highest 15N distribution rate in flowers.
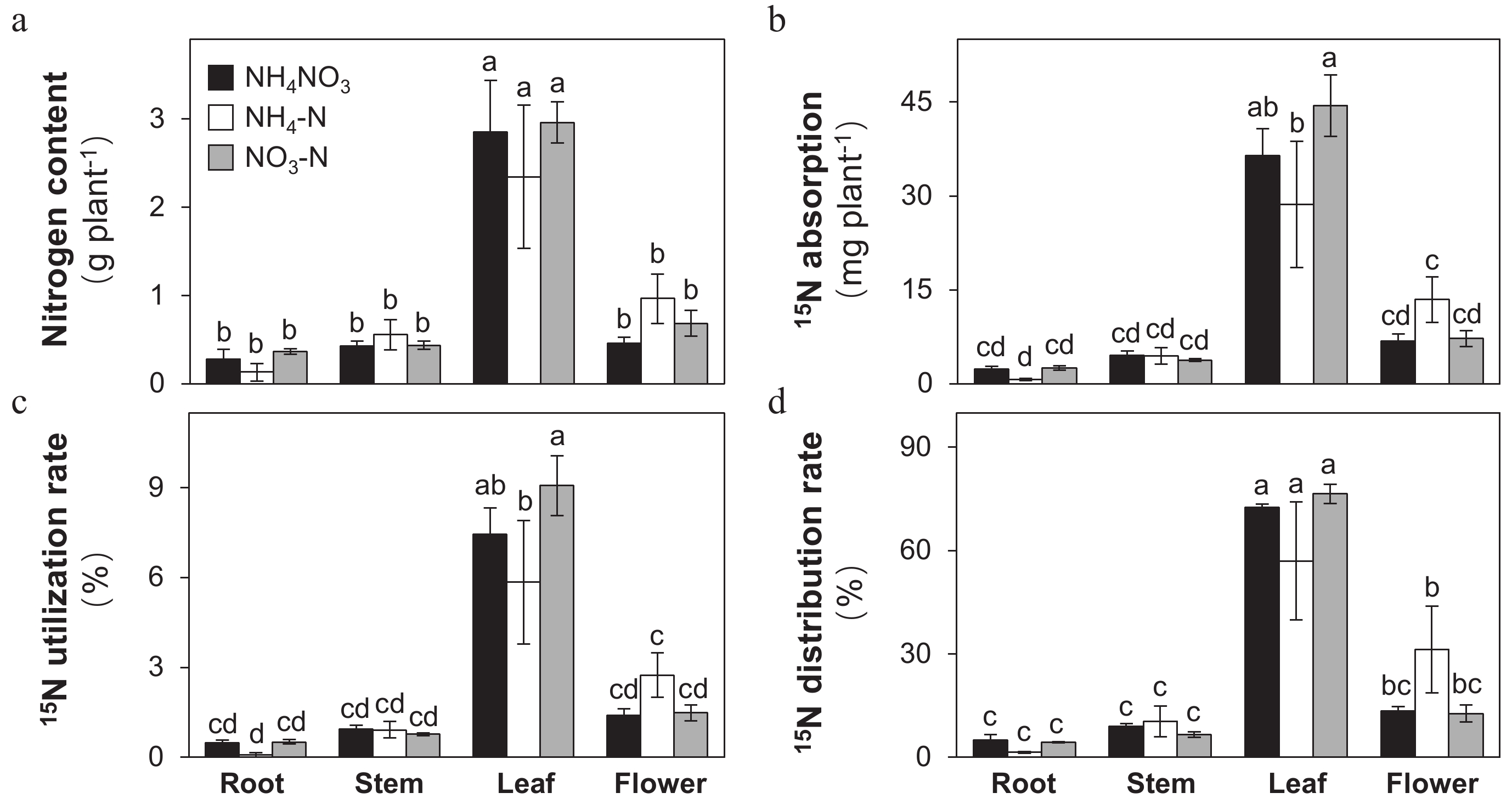
Figure 2.
Effects of ammonium-nitrate (NH4NO3), ammonium (NH4-N), and nitrate (NO3-N) on the tissue-level distribution of N within Petunia plants. (a) Nitrogen content , (b) 15N absorption content, (c) 15N utilization rate, and (d) 15N distribution rate in roots, stems, leaves and flowers measured for each treatment (SE, n = 3). Different letters above the bars in each tissue indicate significant difference between treatments using one-way ANOVA test in SPSS software at P < 0.05.
Different forms of inorganic N influence petal color traits in Petunia
-
Considering that NH4-N dramatically changed 15N distribution into floral tissues compared to other forms of N, we measured the color traits of Petunia plants for each treatment via a portable colorimeter. Changes in CIELab coordinates including perceptual lightness (L*), redness/greenness (a*), yellowness/blueness (b*), chroma (C*), and hue angle (h°) of Petunia petals are presented in Fig. 3. Relative to NH4NO3 treatment, both NH4-N and NO3-N treatment decreased L* and h° and increased b* and C* (Fig. 3a). However, there was no significant difference between NH4-N and NO3-N treatments in terms of their effects on color traits of white petals. The principal component analysis (PCA) of color traits further confirmed our findings (Fig. 3b).
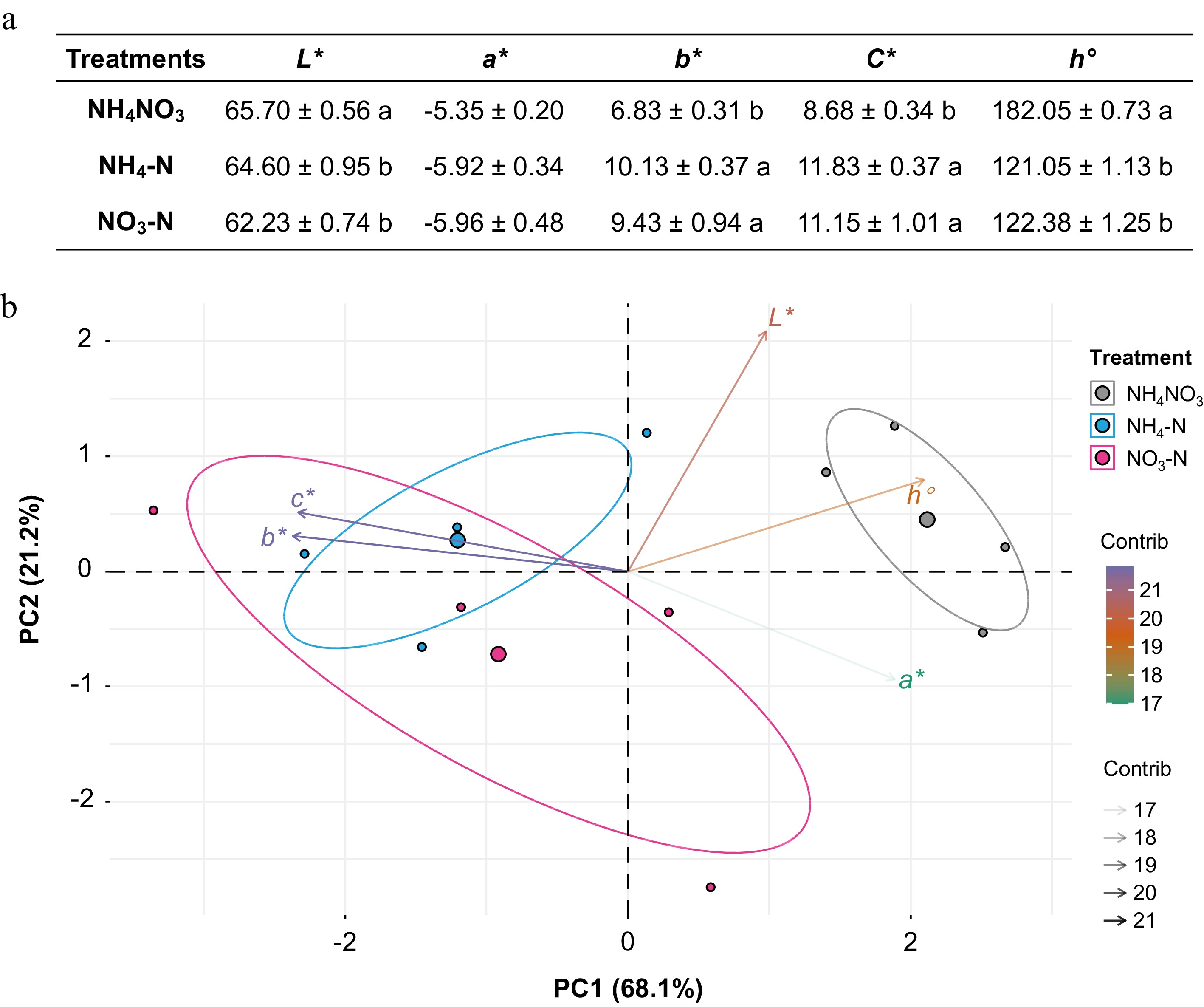
Figure 3.
Effects of ammonium-nitrate (NH4NO3), ammonium (NH4-N), and nitrate (NO3-N) on petal color traits. (a) CIELab coordinates including perceptual lightness (L*), redness/greenness (a*), yellowness/blueness (b*), chroma (C*), and hue angle (h°) of Petunia petals measured for each treatment via a portable colorimeter (SE, n = 4). Colors (green to purple) and transparency of arrows toward each parameter indicate their contributions to the principal components. Different letters above the bars in each column indicate significant difference between treatments using one-way ANOVA test in SPSS software at P < 0.05. (b) The principal component analysis (PCA) of color traits in three treatments.
Different forms of inorganic N changed the petal amino acid profile in Petunia
-
Most pigments in Petunia petals (except yellow) are derived from flavonoid and anthocyanins and amino acids were their direct precursors. We then analyzed the effects of different N forms on the petal amino acid profile in Petunia plants at three developmental stages (Fig. 4a). Via LC-MS/MS, we had identified a total of 14 amino acids in petals including four glutamate (Glu) family amino acids, five aspartate (Asp) family amino acids, three aromatic amino acids and serine and leucine (Supplemental Table S1). We first compared effects of different N forms on total content of glutamate family, aspartate family and aromatic amino acids (Fig. 4b−d). Under treatment of NH4NO3, amino acid profiles displayed clear patterns of changes with the development of flowers. Content of Glu family and aromatic amino acids increased from S1 to S3 stage whereas Asp family decreased in NH4NO3 treated petals. NO3-N treated plants lost this developmental pattern with similar content of amino acids accumulated in petals throughout the three stages. NH4-N treated plants accumulated slightly more amino acids that NO3-N treated plants. More importantly, Glu family amino acids in NH4-N treated plants showed similar developmental patterns to those treated with NH4NO3, suggesting that NH4-N treatment has more positive effects on the petal amino acid profile than NO3-N treatment.
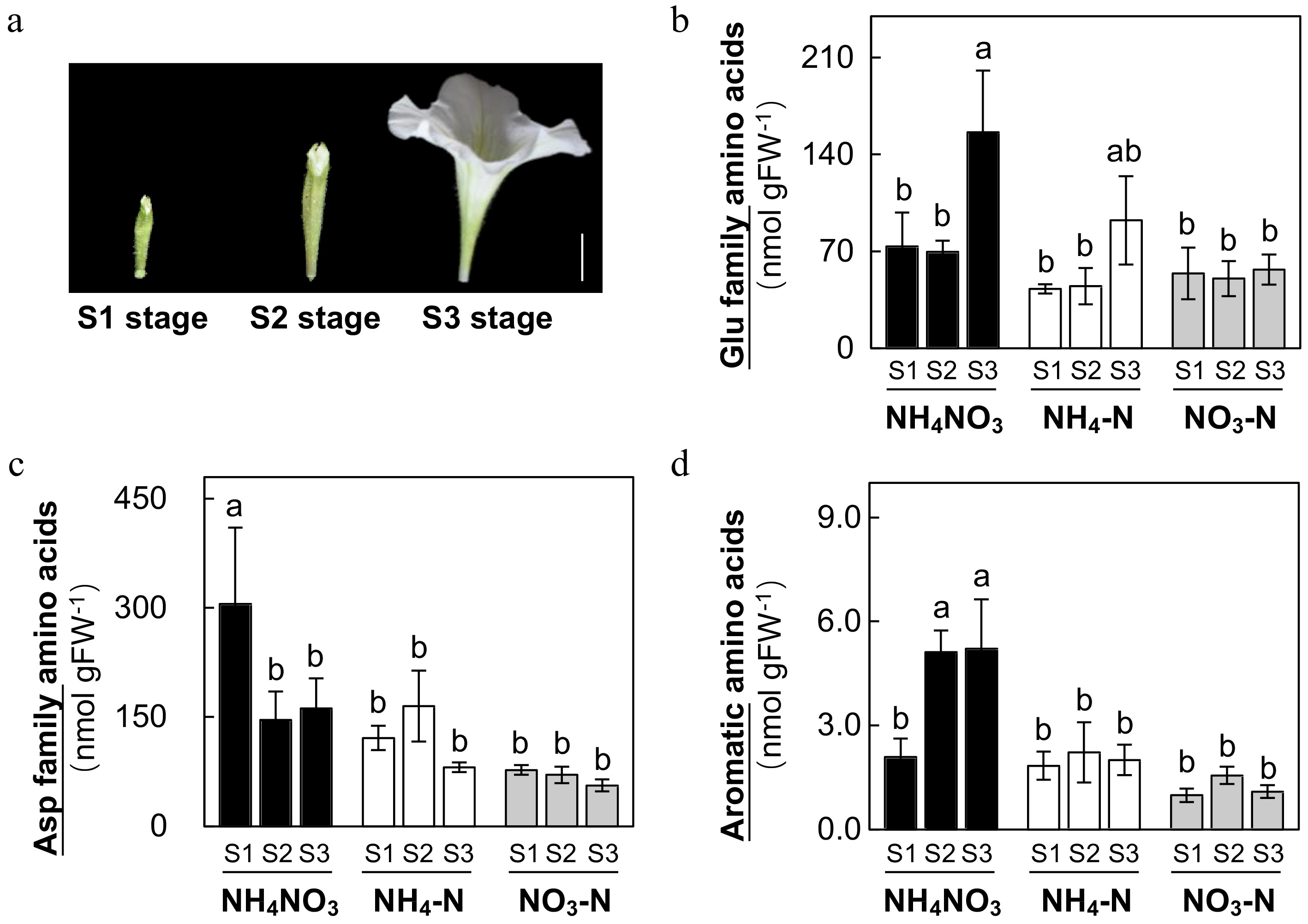
Figure 4.
Effects of ammonium-nitrate (NH4NO3), ammonium (NH4-N), and nitrate (NO3-N) on petal amino acid profiles in Petunia plants. Petal tissues were collected at S1, S2 and S3 stages indicated in (a) (scale bar = 1 cm) and total concentrations of glutamate family amino acids, aspartate amino acids and aromatic amino acids (b)−(d) (SE, n = 4). Different letters above the bars indicate significant difference between treatments using one-way ANOVA test in SPSS software at P < 0.05. Concentrations (nmol g FW−1) of individual amino acids are reported Supplemental Table S1.
We then presented changes in individual amino acid content in detail (Fig. 5). As mentioned above, Glu family amino acids including Glu, glutamine (Gln), arginine (Arg) and proline (Pro) showed very similar trends in changes of their content in both NH4NO3 and NH4-N treatments but not in NO3-N treatment (Fig. 5a). Different forms of N did not affect the developmental changes of the content of threonine (Thr), isoleucine (Ile) and lysine (Lys) in NH4-N and NO3-N treatments relative to NH4NO3 (Fig. 5b). However, content of two major Asp family amino acids, Asp and asparagine (Asn), were significantly influenced by treatments of NH4-N and NO3-N mainly at the S1 stage of flower development. Aromatic amino acids displayed significant changes under various N treatments. Content of tyrosine (Tyr), tryptophan (Trp) and phenylalanine (Phe) in NH4NO3 treated petals increased with flower development except at the S3 stage, content of Phe decreased (Fig. 5c). NH4-N and NO3-N treatments did not change content of Tyr, Trp and Phe significantly from the S1 to the S3 stage. To be noted, NH4-N petals did have slightly higher content of Tyr than those treated with NO3-N. Content of serine (Ser) did not significantly change with various treatments and content of leucine (Leu) in NO3-N was lower than those in the other two treatments.
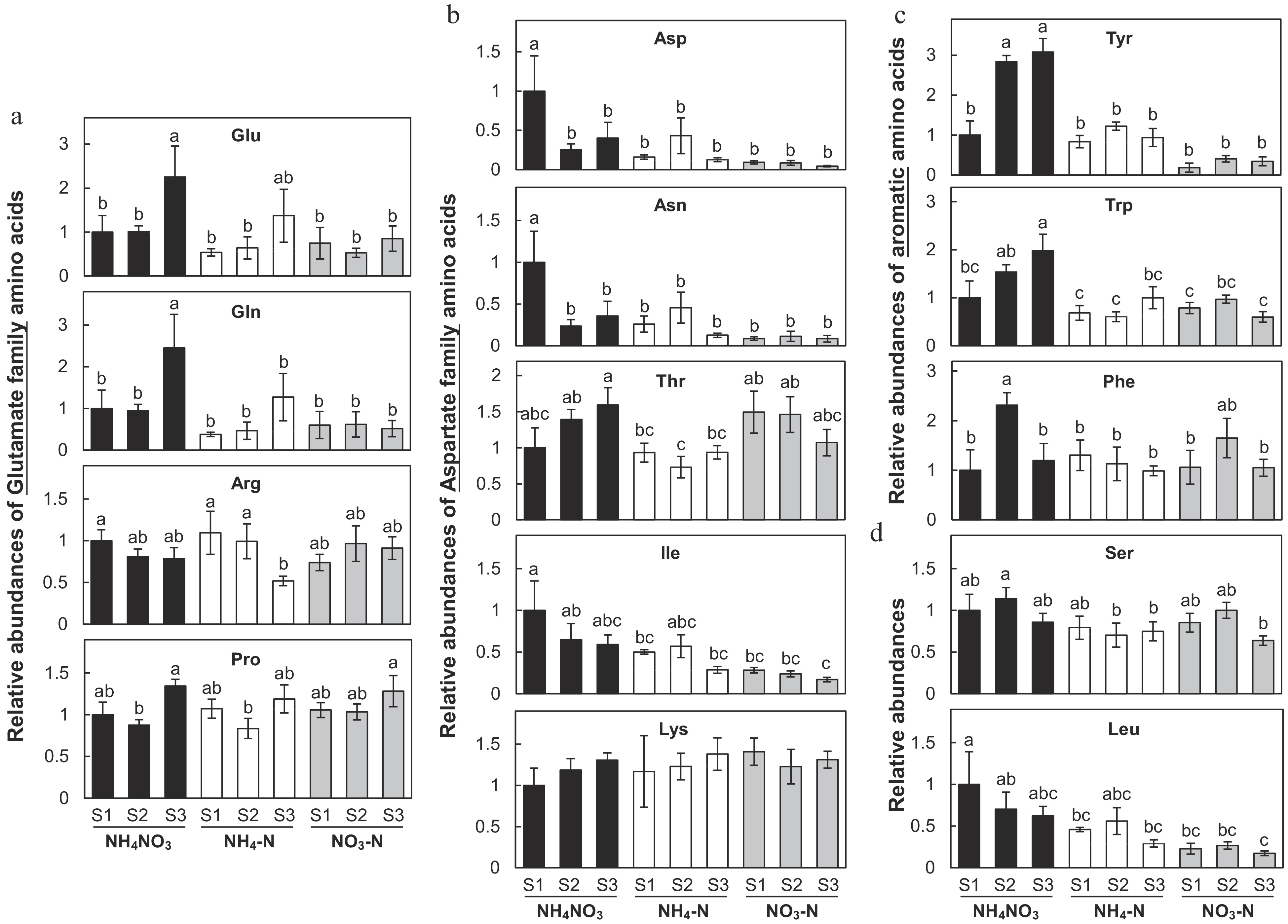
Figure 5.
Effects of ammonium-nitrate (NH4NO3), ammonium (NH4-N), and nitrate (NO3-N) on relative abundance of amino acids in Petunia petals. (a) Relative abundance of glutamate family amino acids, (b) aspartate family amino acids, (c) aromatic amino acids, and (d) Ser as well as Leu were displayed in column plots (SE, n = 4). Different letters above the bars indicate significant difference between treatments using one-way ANOVA test in SPSS software at P < 0.05. Concentrations (nmol g FW−1) of individual amino acids are reported in Supplemental Table S1.
Clustering analysis and principal component analysis of petal amino acid profiles highlighted differences between treatments
-
We carried out clustering analysis and PCA with amino acid data to better dissect the effects of different N forms on the petal amino acid profile (Fig. 6). First, via clustering analysis, amino acids could be categorized into two groups. Group one included Asp, Asn, Ile, Leu and Arg whose content were the highest at the S1 stage with NH4NO3 treatment. Group two including Phe, Ser, Thr, Tyr, Trp, Glu, Gln, Pro and Lys, on the other hand, increased in their content at the S2 and S3 stages with NH4NO3 treatment. We then could clearly observe that both NH4-N and NO3-N treatment significantly influenced the petal amino acid profile relative to that with NH4NO3 treatment (Fig. 6a). PCA with amino acid data provided more information (Fig. 6b). Except S2, the petal amino acid profile in NH4-N was much closer to that in NH4NO3 at both the S1 and S3 stages. Taken together, different forms of N could greatly affect the amino acid profile in Petunia petals. However, relative to NO3-N treatment, total content of amino acids and changes in content of individual amino acids in NH4-N treated petals were closer to those with NH4NO3 treatment.
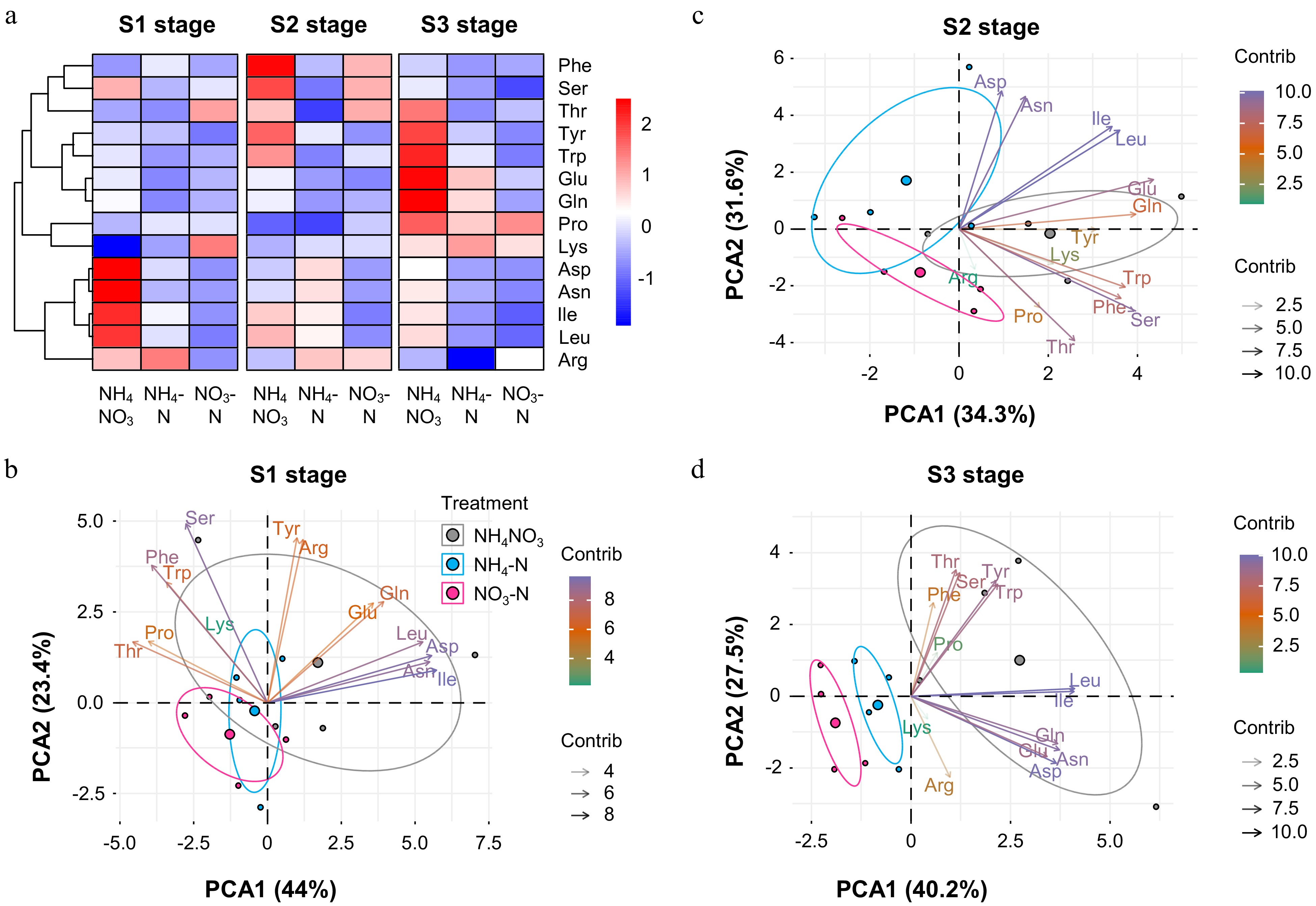
Figure 6.
Further analysis of amino acid data. Heatmap representation of concentrations of 14 amino acids at (a) S1−S3 stages and (b), (c), (d) PCA of amino acids at each developmental stage. Colors (green to purple) and transparency of arrows toward each parameter indicate their contributions to the principal components.
Amino acid profiles were correlated with petal color traits in response to different forms of N
-
Correlation analysis with petal color traits and amino acid profile data indicated that changes in the content of some amino acids were connected to petal color traits (Fig. 7). Thr, Tyr and Trp was positively correlated with the hue angle (h°) of Petunia petals. Thr and Leu were negatively correlated with the yellowness/blueness (b*) and chroma (C*) whereas Tyr was negatively correlated with b*. These data suggested that when the petal amino acid profile was modified by different forms of N, the color traits such as color and brightness of petals could change accordingly.
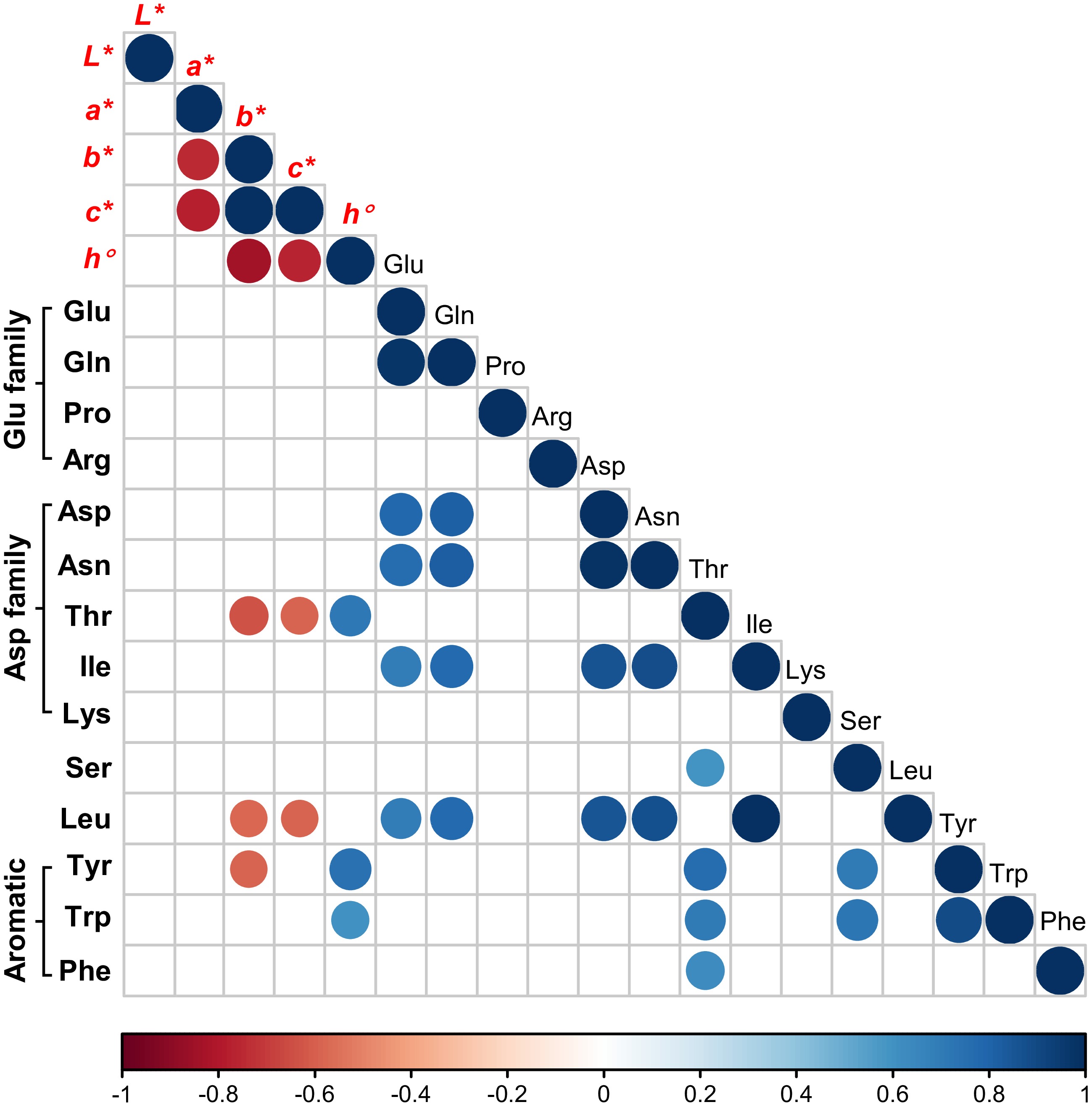
Figure 7.
Correlation analysis of L*, a*, b*, c*, h° and 14 amino acids. Data were processed via R programming. The intensity of blue and red color as well as the size of each dot are proportional to the calculated correlation coefficient (r).
Considering we collected these data from a white-flower cultivar 'Mitchell Diploid', we purchased four commercially available cultivars of Petunia with four flower colors (Fig. 8a, b) and treated them in the same way to further analyze various forms of N on Petunia petals with colors. Petal color traits including L*, a*, b*, C* and h° of white-, pink-, dark pink- and red-flower Petunia plants treated with NH4NO3, NH4-N, and NO3-N were analyzed and presented in Supplemental Table S2. Interestingly, regardless of petal colors, relative to NH4NO3 treatment, both NH4-N and NO3-N treatment could influence L*. For pink-flower cultivar, NH4-N and NO3-N treatments slightly decreased a*, b* and C*. And for dark pink- and red-flower cultivars, NH4-N and NO3-N treatments slightly increased a*, b* and C* (Supplemental Fig. S1). PCA of petal color traits for each cultivar are shown in Fig. 8c−f and revealed more information about the different treatments. For the white-flower cultivar, similar to data we collected from 'Mitchell Diploid', NH4-N and NO3-N could be clearly separated from NH4NO3 with a*, b* and C* contributing the most to separation (Fig. 8c). But NH4-N and NO3-N could not be separated from each other. For the pink-flower cultivar, NH4-N could be partially separated from NO3-N in PC2 with b* and h° contributed the most (Fig. 8d). And for the dark pink-flower cultivar, NH4-N could be completely separated from NO3-N in PC2 with a* and C* contributed the most (Fig. 8e). For the red-flower cultivar, NH4-N could be partially separated from NO3-N in PC2 with b* and h° contributed the most. To be noted, NO3-N could be completely separated from NH4NO3 in PC1 (Fig. 8f). Taken together, different forms of N could impact petal color traits. And for petals with colors, NH4-N and NO3-N could affect the redness/greenness and yellowness/blueness but the lightness of petals contributed less to differences between treatments.
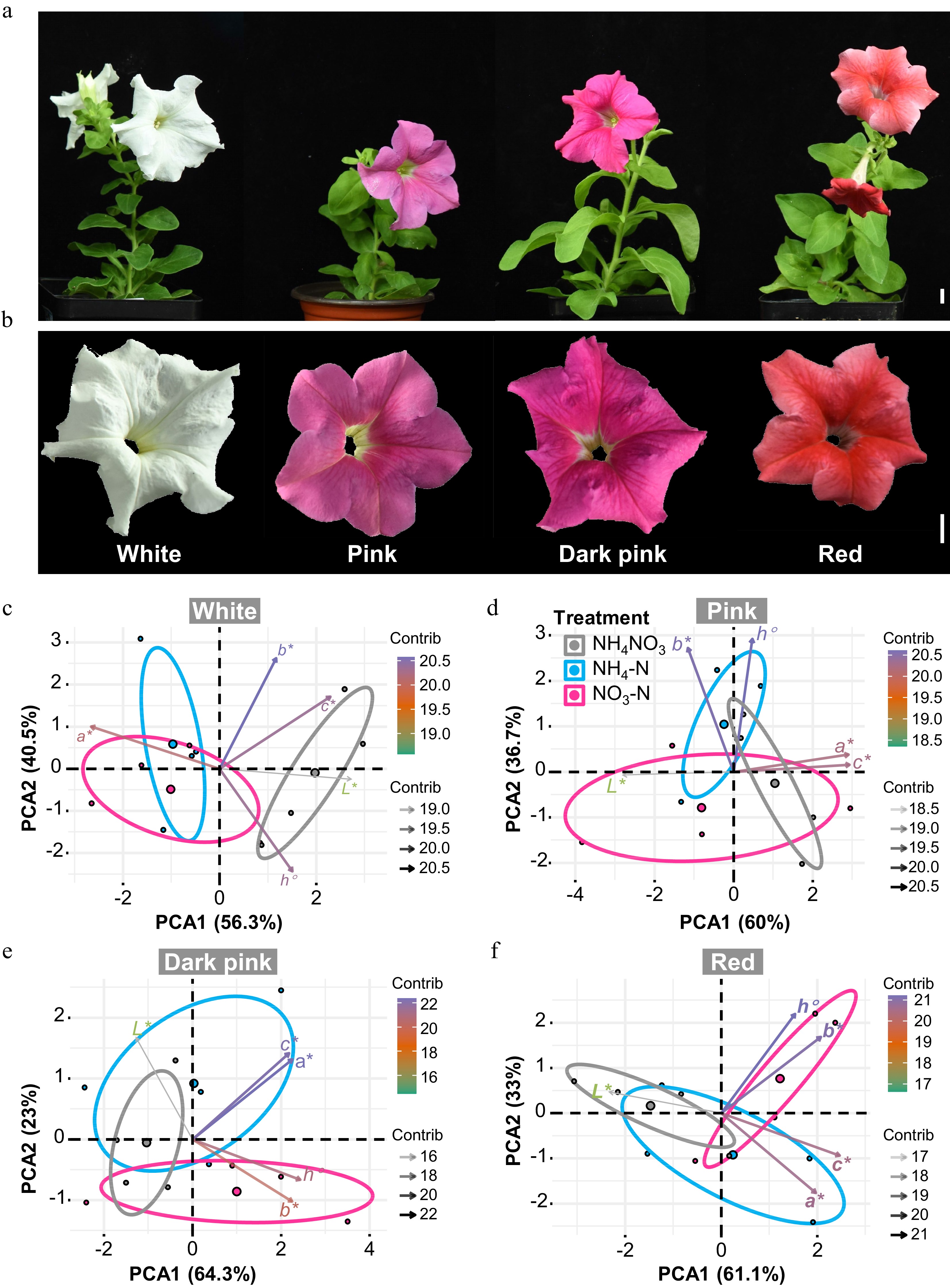
Figure 8.
Relations between amino acids and petal color traits of Petunia cultivars with different colors. (a), (b) Plant and petal appearance of four Petunia cultivars (Scale bar = 1 cm). (c)−(f) PCA of L*, a*, b*, C* and h° in each cultivar. Colors (green to purple) and transparency of arrows toward each parameter indicate their contributions to the principal components.
-
Ornamental traits such as size of flowers and color of petals are fundamentally determined by the metabolic status on the whole-plant level. Extensively performed agricultural practice and continuously increased concentration of CO2 in the atmosphere have both aggravated N conditions in soil. This study has been designed to specifically investigate the effects of ammonium or nitrate as the sole source of inorganic N on the developmental and chemical features of floral tissues in Petunia.
According to the total N content on the whole-plant level after each treatment (Fig. 1a), we think that the Petunia x hybrida cv 'Mitchell Diploid' plants we used in this study do not have preferences to a specific form of inorganic N. To be noted, given the same amount of inorganic N (2 mM), plants treated with NO3-N alone accumulated more biomass. Previous studies[28] pointed out that responses of Petunia plants to different forms of inorganic N depended on the concentrations of N applied to plants and the medium used for plant growth. Ammonium is more effective than nitrate on the growth promotion in rice[29] and maize[30]. Complex effects of ammonium and nitrate on plant growth, CO2 assimilation and activities of key enzymes involved in N metabolism have also been reported in blueberry, raspberry and strawberry[31]. Similarly, different ratios of nitrate to ammonium applied to sweet pepper at different developmental stages bring various effects on flowering, fruit set and yield[32]. It is then clear to us that effects of different forms of inorganic N on ornamental plants should be discussed case by case[17].
We found that in Petunia plants, NH4-N and NO3-N treatment could both enhance the number of branches and flowers at the cost of leaf and corolla area. Plants can generally benefit from the direct application of NH4-N at an appropriate level since the assimilation of ammonium is far more energetically efficient relative to nitrate assimilation. Meanwhile, nitrate is more mobile which can compensate this limitation. Therefore, applications of NH4-N and NO3-N alone could be superior to NH4NO3 treatment. N can shape the architecture of plants. Nitrate regulates shoot branching on the molecular level to control the outgrowth of axillary buds[33]. Nitrate can also control the transition from the vegetative to reproductive phase and therefore, is involved in flowering time regulation[34]. On the other hand, ammonium application usually leads to accumulation of proteins which is generally beneficial to crop growth. However, there are clear differences between the two treatments. Studies on tobacco have also shown that ammonium nitrogen treatment results in a reduction in leaf area[35]. 15N labeling experiments confirmed that only NH4-N changed the distribution of 15N between various tissues with floral tissues absorbing and utilizing more 15N. Meanwhile, relatively smaller amounts of 15N was distributed into the roots and leaves in NH4-N treated plants. Taken together, when ammonium is used as the sole source of inorganic N, Petunia plants can manage to import more N into floral organs and produce more flowers with smaller size. More importantly, both NH4-N and NO3-N treatment can modify the amino acid profile on the whole-plant level. And as the basic compounds for plant development and signaling molecules[36], amino acids may play direct roles in the formation of plant architecture via regulating the meristem tissues[37], which requires further study.
Here we need to clarify that in most cases, ammonium (NH3 or
${\text {NH}_4^+} $ For amino acids in petals, there are few studies available. Roles of amino acids in flower development and senescence have been briefly reported in Eustoma grandiflorum[42]. Although most local amino acids in floral tissues are imported from roots and leaves, it has been recently proposed that some amino acids can be synthesized in flowers[18]. For example, de novo synthesis of Asn may exist in flowers due to the fact that asparagine synthetase1 (ASN1) is expressed abundantly in Arabidopsis flowers[43]. The biological roles of Asn in floral tissues can be important. On one hand, Asn carries more N than other amino acids and serves as a good N sink[44]. On the other hand, hydrolysis of Asn can release ammonium which in turn can be used to synthesize Gln[45,46]. More importantly, amino acids provide precursors and energy for petal color and scent. We think the metabolic roles of amino acids in regulating ornamental traits should be much more emphasized in this field. Accumulation of free amino acids is usually considered a sign for ammonium stress[47]. The fact that total content of 14 amino acids in NH4-N petals were not higher than those in NH4NO3 indicates that the NH4-N in our study was not a stress treatment. Given that the assimilation of inorganic N into amino acids primarily occur in roots, the 15N label data and petal amino acid profiles collected in this study indicated that Petunia plants treated with NH4-N alone loaded relatively more amino acids such as Glu, Gln, Asp and Asn to their flowers (Fig. 5a, b). We also found that changes in petal amino acid profiles were correlated with changes in their color traits (Fig. 7), which should be further investigated at the molecular level in ornamental plants.
CONCLUSIONS
-
In summary, we have analyzed effects of ammonium and nitrate alone on plant growth, the tissue-level N utilization and distribution, petal color traits and petal amino acid profiles in Petunia plants. We have confirmed that NH4-N can increase 15N distribution to floral tissues, modify the amino acid profiles in petals which in turn bring changes to petal colors. Mechanisms of governing flower development by different forms of N or other environmental cues require further investigation and will definitely lead to a better understanding of the metabolic regulation of ornamental traits.
We gratefully acknowledge the support of National Key Research and Development Program of China (2018YFD1000405 to F.M.). We thank College of Horticulture Science and Engineering at Shandong Agricultural University for supporting the Experimental Core Facilities where we performed mass spectrometry and phenotypic analysis. We thank State Key Laboratory of Crop Biology at Shandong Agricultural University for providing lab space and plant growth chambers. This work was supported by a grant from National Key Research and Development Program of China (2018YFD1000405) to Fangfang Ma.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Concentrations of 14 free amino acids in Petunia petals at three developmental stages measured via GC-MS for each treatment (SE, n=4). Different letters in each row indicate significant difference between treatments using one-way ANOVA test in SPSS software at P < 0.05.
- Supplemental Table S2 CIELab coordinates including perceptual lightness (L*), redness/greenness (a*), yellowness/blueness (b*), chroma (C*), and hue angle (h°) in petals of four petunia cultivars measured for each treatment via a portable colorimeter. Detailed data of four biological replicates for each treatment were presented here.
- Supplemental Fig. S1 Effects of ammonium-nitrate (NH4NO3), ammonium (NH4-N), and nitrate (NO3-N) on petal color traits in four commercially available petunia cultivars. Changes in CIELab coordinates including L*, a*, b*, C*, and h° in white- (a), pink- (b), dark pink- (c) and red-flower cultivar (d) were plotted in column figures (SE, n=4; scale bar = 1cm). Different letters above the bars in each column indicate significant difference between treatments using one-way ANOVA test in SPSS software at P < 0.05.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Guo H, Wang G, Zhang P, Wang G, Bao Z, et al. 2022. Ammonium and nitrate impact petal color traits and amino acid profiles differently in Petunia. Ornamental Plant Research 2:14 doi: 10.48130/OPR-2022-0014 

Ammonium and nitrate impact petal color traits and amino acid profiles differently in Petunia
- Received: 05 August 2022
- Accepted: 06 September 2022
- Published online: 22 September 2022
Abstract: Nitrogen (N) is vital for ornamental plants to fulfill fundamental cellular functions. Effects of inorganic N, including nitrate and ammonium, on ornamental traits at the biochemical level are less reported. Given that the continuously increased concentration of CO2 in the atmosphere has greater negative effects on nitrate reduction than ammonium utilization in C3 plants, it is vital for us to dissect various effects of different inorganic N forms on flower development, which will lead to a better understanding of the metabolic regulation of ornamental traits. In this study, Petunia × hybrida cv 'Mitchell Diploid' (MD) plants at 6−8-leaf stage were treated hydroponically with the same and regular amount of inorganic N for one month but in three different forms, 1 mM NH4NO3 as the control (NH4NO3), 2 mM NH4Cl alone (NH4-N) and 2 mM NaNO3 alone (NO3-N). Results indicated that relative to the control, effects of NH4-N and NO3-N treatment alone on plant growth, N utilization and distribution, petal color traits and petal amino acid profiles were different by varying degrees. We have confirmed that NH4-N could specifically increase 15N distribution to floral tissues, change the amino acid profiles in petals which in turn bring changes to petal colors. Results with four commercially available Petunia cultivars with different flower colors also validated our findings. This study has been designed to specifically investigate effects of ammonium or nitrate as the sole source of inorganic N on the developmental and chemical features of floral tissues in Petunia.
-
Key words:
- ammonium /
- nitrate /
- petal /
- color traits /
- amino caid profiles /
- 15N label