









-
Anthocyanins, defined as plant pigments, are derivatives of flavone and confer red, blue and purple colors to many plant organs such as flowers, leaves, stems, fruits, and roots. The accumulation of anthocyanins in flowers is helpful for pollination and propagation by attracting pollinators like birds and insects[1,2]. Moreover, anthocyanins provide protection against abiotic and biotic stresses by acting as scavengers of reactive oxygen species and also beneficial to human health[3−6].
The anthocyanin biosynthetic pathway is well-illustrated in many plants[7]. Anthocyanins are biosynthesized from a condensation of malonyl-CoA and 4-coumaroyl CoA and there are seven enzymes involved in the biosynthetic pathway, including the upstream enzymes chalcone synthase (CHS), chalcone flavanone isomerase (CHI) and flavanone 3-hydroxylase (F3H) and the downstream enzymes flavonoid 3'-hydroxylase (F3'H), dihydroflavonol 4-reductase (DFR), leucoanthocyanidin dioxygenase (LDOX), and UDP-glucose:flavonoid 3-O-glucosyltransferase (UFGT). Anthocyanins, biosynthesized in endoplasmic reticulum (ER), are needed to be transported into the vacuole. Glutathione S-transferase (GST) is one of anthocyanin transporters[8]. The acidic environment of the vacuole enhances the stability of anthocyanins.
Many studies have reported that anthocyanin biosynthesis is tightly regulated by a ternary complex of MYB-bHLH-WD40 (MBW) transcription factors. There is slight difference in the regulation mechanism between monocots and dicots. In monocots, genes in the anthocyanin biosynthesis pathway are regulated by a MBW complex. In dicots, regulations of upstream genes (CHS, CHI, and F3H) are conducted by R2R3-MYB without co-regulators, but the downstream genes (F3′H, DFR, ANS/LDOX, and UFGT) are regulated by a MBW complex[9]. In the MBW complex, MYB and, to some extent, bHLH are the main components controlling genes that are activated[10].
Peach (Prunus persica) is an economically important fruit crop. Anthocyanin pigmentation is important for the economic value of peach. Several peach MYB genes are demonstrated to regulate the anthocyanin biosynthesis in different tissues. MYB10.1 and MYB10.3, with bHLH3, are the likely regulators of anthocyanin biosynthesis in peach fruit[2]. PpMYB10.1 also is a regulator of anthocyanin biosynthesis in red-skinned peach[11]. PpMYB108 was suggested to be associated with anthocyanin biosynthesis and red color of peach flower by targeting the DFR gene[12]. Peace, a MYB-like transcription factor, is involved in peach flower anthocyanin biosynthesis[13]. PpMYB10.4 represents a novel branch of anthocyanin-activating MYB genes and activates leaf red coloration in peach[14].
Besides the transcription factors (TFs) in the MBW complex, TFs and regulatory genes such as SPX[15], MiR156[16], MdLNC499[17], PIF3[18], COP1[19], HY5[20], WRKY[21], ERF[22,23]; MADS[24], NAC[25] and JASMONATE ZIM domain (JAZ) genes[26] have also been reported to affect anthocyanin synthesis. NACs are identified by a NAC domain at the N-terminus and comprise a plant-specific transcription factor family. Its members regulate many biological processes, such as shoot apical meristem formation[27], fruit ripening[28,29], wood formation[30], leaf senescence[31], and cell elongation[32]. The NAC domain is responsible for the nuclear localization and DNA binding activity[33]. Members of the NAC family are reported to regulate anthocyanin biosynthesis in several plants, including apple[34], litchi[35], and peach[25]. In addition, the formation of NAC homodimers or heterodimers is an important way to play its biologic function. NAC transcription factor BL, a key gene for blood flesh of peach, interacts with PpNAC1 and activates the PpMYB10.1 transcription[25].
In this study, PpNAC25 was cloned from peach, which is a homologous gene of PpNAC1 and highly expressed in peach fruit. The tissue-specific expression profile of PpNAC25 was analysis. Then, PpNAC25 was overexpressed in poplar and the pigment phenotype of poplar shoot was analyzed. Protein interaction of PpNAC25 and PpNAC1 was determined.
-
Peach cultivar 'QiuMiHong (QMH)' was used in this study and is kept at the fruit tree germplasm repository of Henan Agricultural University (Henan Province, China). Shoot tips, stem, young and old leaves, flower bud, bloom flowers and fruit flesh were collected from 'QMH'. All samples were prepared during April to August of 2021. Shoot tips and stem were sampled from sylleptic branches. All collected samples were frozen in liquid nitrogen immediately and then stored at −80 °C.
Phylogenetic tree
-
Peach NAC amino acid sequences were downloaded from plant transcription factor database (
http://planttfdb.gao-lab.org ). The sequences of NAC involved in fruit ripening, anthocyanin biosynthesis and thickening of the cell wall were downloaded from the genome database of Phytozome (https://phytozome-next.jgi.doe.gov ). Multiple sequence alignments were performed with clustal X software (default parameters). A neighbor-joining tree was constructed using software MEGE version 7.0. The bootstrap analysis was performed based on 1,000 replicates.Gene expression analysis
-
Total RNA was extracted with RNA-easy isolation reagent (Vazyme). First-strand cDNA was synthesized using a PrimeScript first strand cDNA synthesis kit transcriptase (Vazyme). The RT-qPCR was performed using ABI PRISM 7500 FAST sequence detection system. Three biological replicates were performed for each individual experiment. The qRT-PCR reaction procedure was carried out according to Cheng et al.[36]. Melt curve was detected to guarantee primer quality for qRT-PCR assay. The peach gene PpGAPDH (Prupe.8G132000) was used as an internal control[36]. Supplemental Table S1 shows the primers used for RT-qPCR.
Transformation of poplar
-
The tissue culture seedling of Populus × euramericana cv. 'Nanlin895' is kindly provided by Laigeng Li. The coding sequence of PpNAC25 was amplified by PCR using specific primers in Supplemental Table S1. Then, the coding sequence was cloned into pSAK277 vector to generate 35S-PpNAC25 construct. The construct was delivered into 'Nanlin895' by the protocol described by Li et al.[37]. The construct of 35S-GFP was used as the control. All transgenic lines were confirmed by detecting transgene expression. Three confirmed transgenic lines were planted in a nutrient soil substrate. Transgenic lines were multiplied through micro-cutting propagation for biological repeats and cultured at a culture room (light intensity: 150 µmol m−2 s−1; 25 ± 1 °C; photoperiod: 16 h/8 h of light/dark).
Measurement of anthocyanin
-
The methanol-HCl method was used to extract total anthocyanin from the shoot tips of poplar. Approximately 1 g of the samples were incubated in 5 mL of 1% (v/v) methanol-HCl overnight in the dark. The absorbance of each extract was measured at 530, 620, and 650 nm with a spectrophotometer. The anthocyanin concentration was measured and calculated as described[38].
Yeast one hybrid (Y1H)
-
The full-length coding sequences of PpNAC25 were cloned into indicated restriction enzyme sites of the pB42AD vector. The promoter of PpMYB10.1 and PpMYB10.2 were cloned into indicated restriction enzyme sites of the pLacZi vector. Yeast one-hybrid analysis was carried out according to a previous report[39].
Dual-luciferase assay
-
The dual-luciferase reporter transactivation assay was performed as previously reported[40]. The PpNAC25 coding sequence and promoter region (2 kb upstream of the ATG) of PpMYB10.1 and PpMYB10.2 were cloned into effector and reporter vectors. Overnight-cultured A. tumefaciens GV3101 cells (Tolo Biotech, China) harboring the constructed plasmids were resuspended (OD600 = 0.5) in infiltration buffer (10 mM MES pH5.8, 10 mM MgCl2, and 100 μM acetosyringone) for 3 h in the dark, then infiltrate into 4-week-old N. benthamiana leaves. Luciferase activity of six transformed tobacco leaves from two independent tobacco plants was measured using a dual-luciferase reporter assay system (Tecan SPARK, Austria). Four biological repeats were carried out.
Yeast two hybrid (Y2H)
-
The sequence with the activation domain removed of PpNAC25 and full-length coding sequences of PpNAC1 were cloned into indicated restriction enzyme sites of the pGBKT7 vector. The full-length coding sequences of PpNAC25 and PpNAC1 were cloned into the indicated restriction enzyme sites of the pGADT7 vector. Yeast two-hybrid assays were performed as the Matchmarker GAL4-based two-hybrid system (Clontech). Constructs were cotransformed into the yeast Y2H.The yeasts were grown on selection medium supplemented with DDO (SD base/-Leu/-Trp) and then transferred onto selection medium supplemented with QDO(SD base/-Leu/-Trp/-His/-Ade), with or without 5-bromo-4-chloro-3-indolyl-α-d-galactopyranoside (X-α-Gal) to determine the interactions of the PpNAC25 with PpNAC1.
Bimolecular fluorescence complementation (BiFC) assays
-
BiFC assays were performed as described previously[41]. Full-length PpNAC25 and PpNAC1 cDNAs were cloned into the pBiFC vectors, which contained either N-terminal or C-terminal half of YFP. The PpNAC25-Ecn and PpNAC1-Enn vectors were then transferred into the Agrobacterium tumefaciens strain GV3101 (pSoup). The A. tumefaciens cells containing pNC-BiFC-Ecn/Enn infiltrated 4-week-old Nicotiana benthamiana leaves.
-
Members of the NAC family are reported to regulate anthocyanin biosynthesis in many plants[25, 28, 31,32]. In this study, a fruit-highly expressed NAC (Prupe.4G040900) gene was cloned from the flesh of peach 'QMH' and showed a highest sequence similarity with AtNAC25 after blasting against the Arabidopsis genome sequence database. Then, this NAC gene was named as PpNAC25. PpNAC25 was located in Chr.4 and contained three exons and two introns. PpNAC25 had a conserved NAC domain distributed in the first and second exon (Fig. 1a). A phylogenetic tree was constructed using all peach NACs and some NACs from different plants (Supplemental Fig. S1). Figure 1b showed a branch of the phylogenetic tree which contained three groups of NAC family members involved in the regulation of fruit ripening, anthocyanin biosynthesis and the deposition of secondary cell wall. Our results showed that PpNAC25 had a conserved NAC domain and was clustered with ripening-related NACs.
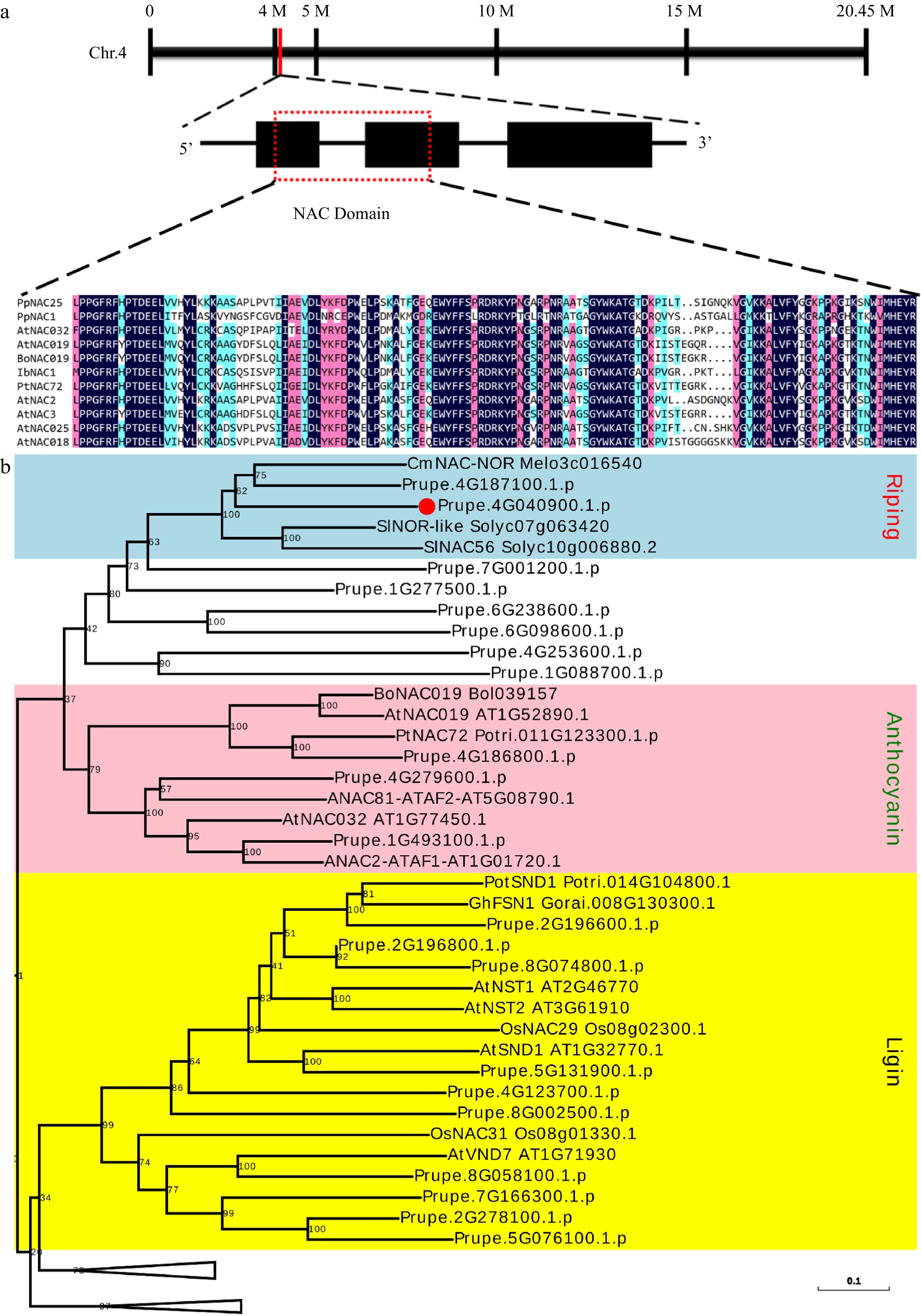
Figure 1.
Gene location and sequence analysis of PpNAC25. (a) Gene location, structure and multiple sequence alignment of NAC domain. (b) A branch of the phylogenetic tree is constructed using NACs in peach and NACs involved in anthocyanin, second cell wall, and ripening from different plants. PpNAC25 was highlighted in bold. The intact phylogenetic tree can be found in Supplemental Fig. S1.
Subcellular location and transcriptional activation activity of PpNAC25
-
The subcellular location of PpNAC25 protein was analyzed in tobacco leaf. The vectors of 35S::GFP and 35S::PpNAC25::GFP were constructed and transiently expressed in tobacco leaves. The green fluorescence induced by 35S::GFP was detected in the nucleus and cell membrane, while the fluorescence of 35S::PpNAC25::GFP was only distributed in the nucleus (Fig. 2a). This result suggested that PpNAC25 proteins were located in the nucleus.
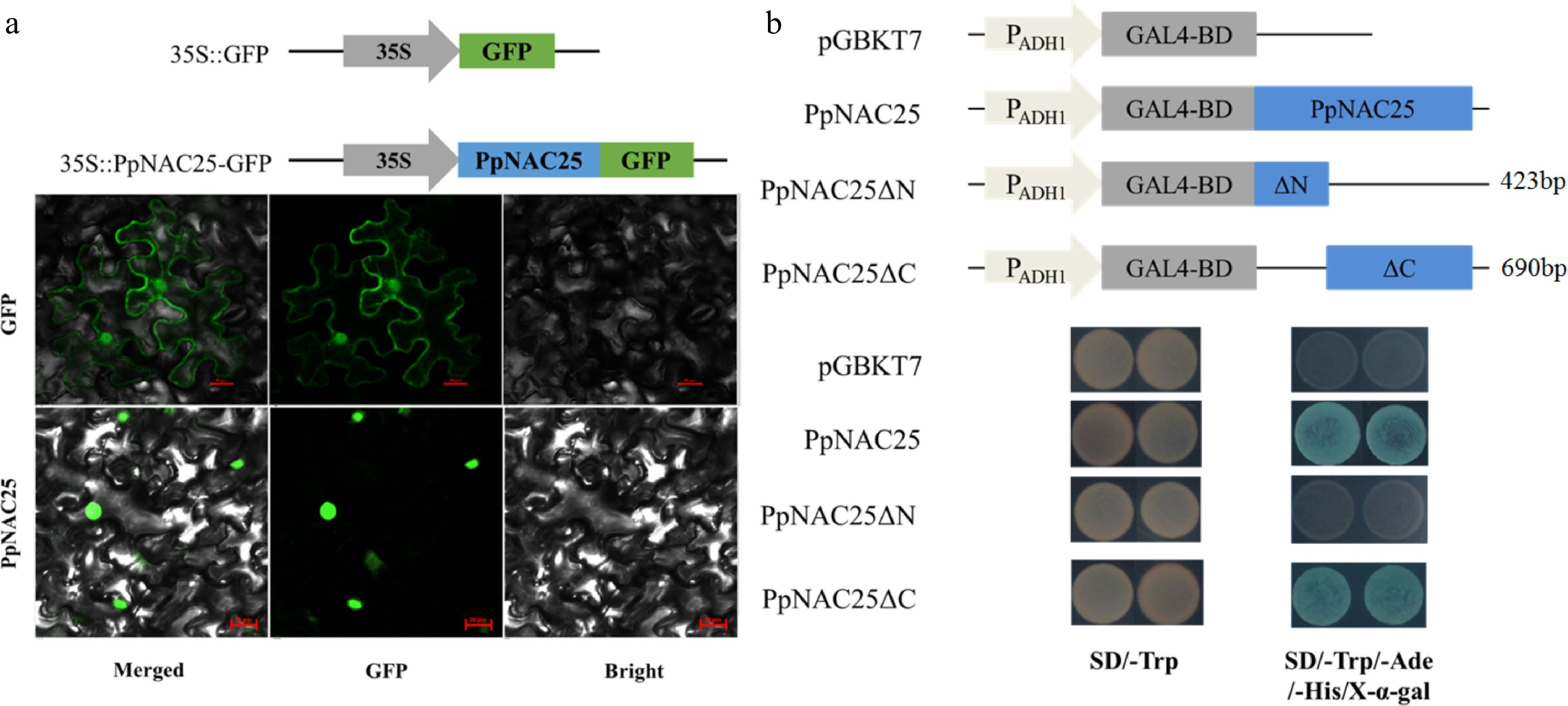
Figure 2.
(a) Subcellular location and (b) transcriptional activation activity of PpNAC25.
The transcriptional activation activity of PpNAC25 protein was analyzed in yeast cells. The vectors of pGBKT7 and pGBKT7-PpNAC25 were transformed into the yeast cell (Fig. 2b). The transfromants containing pGBKT7 and pGBKT7-PpNAC25 grew normally on the media of SD/-Trp, but only the yeast containing pGBKT7-PpNAC25 grew normally and turn to blue on the media of SD/-Trp/-His/-Ade/X-α-gal (Fig. 2b). The results demonstrated that PpNAC25 had a transactivation activity in yeast.
To further confirm the transactivation activity of PpNAC25, two more vectors pGBKT7-PpNAC25ΔN and pGBKT7-PpNAC25ΔC were constructed (Fig. 2b). PpNAC25ΔN represent the 423 bp N-terminal sequences and PpNAC25ΔC represent the 690 bp C-terminal sequences. The transformants containing pGBKT7-PpNAC25ΔN and pGBKT7-PpNAC25ΔC grew normally on the media of SD/-Trp, but only the latter grew normally and turned blue on the media of SD/-Trp/-His/-Ade/X-α-gal (Fig. 2b). Our data confirmed that the activation region of PpNAC25 was located at the C-terminus.
Transcription profile of PpNAC25 in different tissues
-
Seven tissues (shoot tips, stems, young leaves, old leaves, flower bud, bloom flowers, and fruit) were selected to analyze the expression of PpNAC25 (Fig. 3). Among these tissues, two fruit samples were collected at different developmental stages. The result of qPCR showed that PpNAC25 expressed in all detected tissues, but its transcript level in fruit was higher than that in other tissues. Among the remaining six tissues, PpNAC25 was relatively more highly expressed in leaves and flowers than in shoot tips and stem.
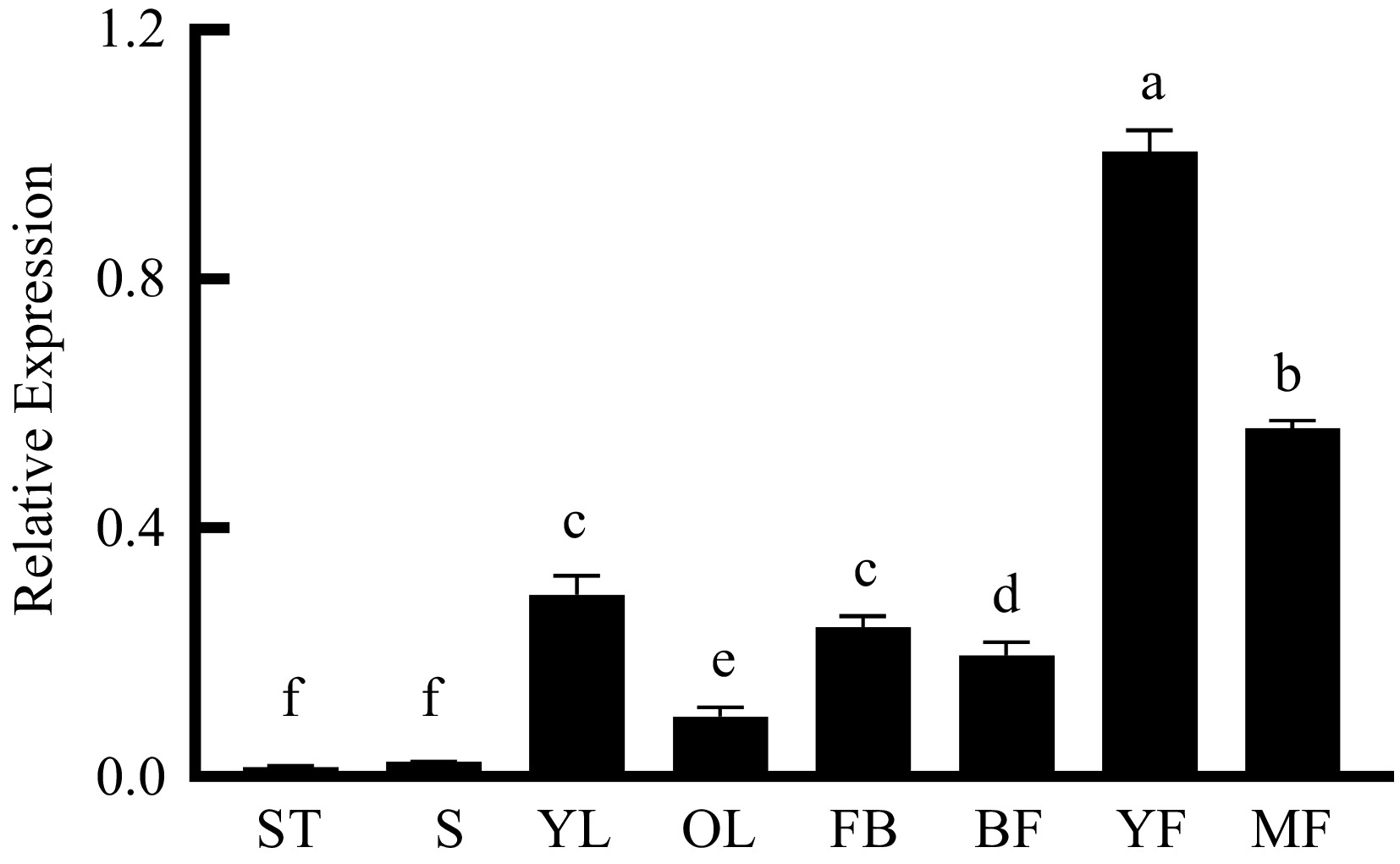
Figure 3.
Tissue specific expressions of PpNAC25. Expression of PpNAC25 in shoot tips (ST), stems (S), young leaves (YL), old leaves (OL), flower buds (FB), bloom flowers (BF), young fruit (YF) and mature fruit (MF).
Overexpression of PpNAC25 in poplars
-
To analyze the gene function of PpNAC25, 35S::PpNAC25 was delivered into poplar and three transgenic lines (#1, #9 and #17) were obtained (Fig. 4d). 35S-GFP was used as control. Our results showed that all three lines were redder in shoot tips than the control (Fig. 4a & b). Analysis of anthocyanin content showed that the transgenic lines were significantly higher than the control (Fig. 4c). Then, we analyzed the transcription level of anthocyanin biosynthesis genes in poplar. Three downstream genes (DFR, ANS and UFGT) of anthocyanin biosynthesis showed a significantly higher transcript level in transgenic lines than in the control (Fig. 4e−g). Mover, compared with control, GST, involved in anthocyanin transportation, was also highly expressed in transgenic lines (Fig. 4h). All these results demonstrated that overexpression of PpNAC25 promotes the anthocyanin accumulation by upregulating the genes involved in anthocyanin biosynthesis and transportation in shoot tips of poplar.
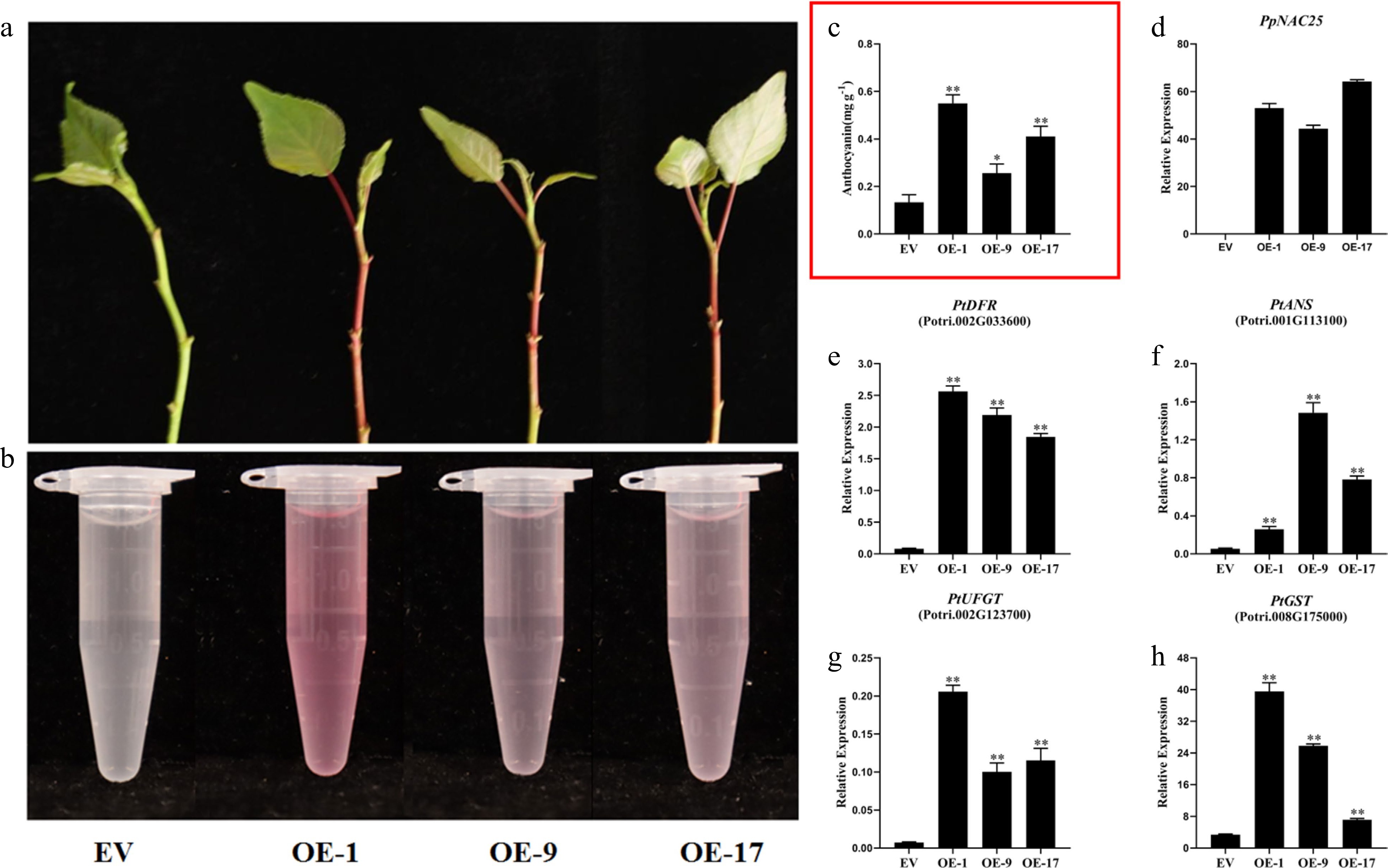
Figure 4.
Effect of PpNAC25 on the anthocyanin biosynthesis of poplar. (a), (b) the pigment of shoot tips from GFP and three transgenic lines. (c) The anthocyanin content of GFP and three transgenic lines. (d) The transcript levels of PpNAC25 in GFP and three transgenic lines. Anthocyanin biosynthetic gene (e) PtDFR, (f) PtANS, (g) PtUFGT and (h) PtGST in GFP and three transgenic lines. Error bars represent ± SD of three replicates (* P < 0.05, ** P < 0.01, *** P < 0.001).
Analyzing regulation of PpNAC25 on PpMYB10.1 and PpMYB10.2
-
To clarify the regulation mechanism of PpNAC25 on anthocyanin biosynthesis, the target gene of PpNAC25 was analyzed in peach. PpMYB10.1 and PpMYB10.2 are two important transcription factors positively regulating anthocyanin biosynthesis of peach fruit[11, 25, 42,43]. Therefore, 2 kb promoter sequences of PpMYB10.1 and PpMYB10.2 were cloned and used to analyze the DNA binding ability of PpNAC25. In Y1H assay, the transformants containing pB42AD-PpNAC25/ pLacZi-PpMYB10.1 or pB42AD-PpNAC25/ pLacZi-PpMYB10.2 grew normally and turn blue on the media of SD/-TU+/X-α-gal. This result suggested that PpNAC25 could bind to the 2 kb promoters of PpMYB10.1 and PpMYB10.2, respectively (Fig. 5a). Meanwhile, the Luc/Ren assay showed that PpNAC25 could significantly enhance the transcriptional activity of the promoters of PpMYB10.1 and PpMYB10.2, respectively (Fig. 5b). These results suggested that PpNAC25 can activate the transcription of PpMYB10.1 and PpMYB10.2 by likely binding to its promoters.
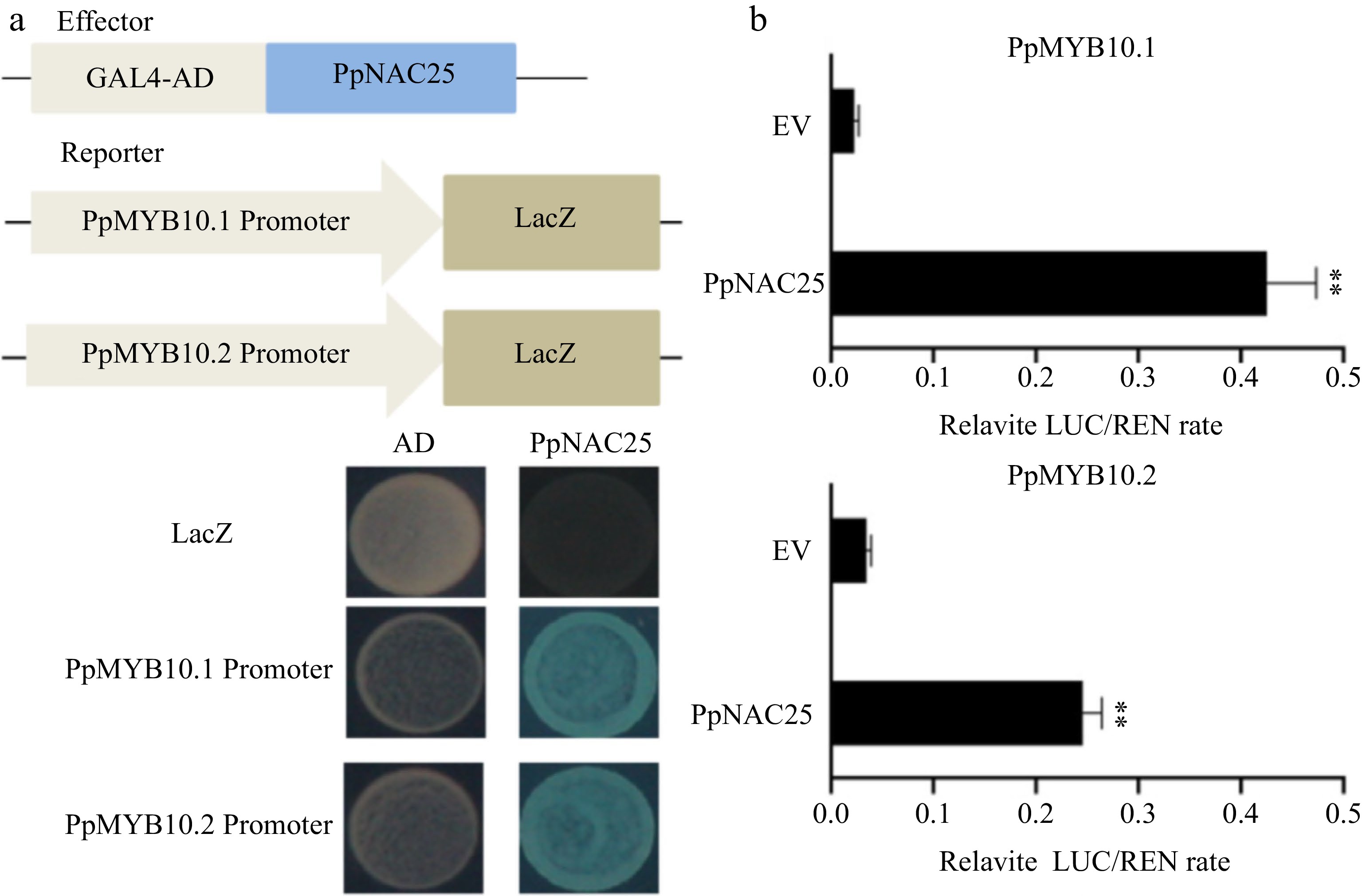
Figure 5.
PpMYB10.1 and PpMYB10.2 are targeted genes of PpNAC25. (a) The binding ability of PpNAC25 to the promoters of PpMYB10.1 and PpMYB10.2 using yeast one-hybrid assays. (b) Determination of PpMYB10.1 and PpMYB10.2 promoter activity induced by PpNAC25 using luciferase (LUC)/Renilla Luciferase (REN) assay. Asterisks indicate statistically significant differences (** P < 0.01, Student's t test, n ≥ 3).
Protein interaction of PpNAC25 and PpNAC1
-
Dimerization of NAC transcription factors are reported to play an important role in regulation of plant development. PpNAC1 (Prupe.4G187100) showed a closed relationship with PpNAC25 in sequence similarity (Fig. 1b) and positively regulates the anthocyanin accumulation of peach fruit[25]. In this study, protein interaction of PpNAC25 and PpNAC1 was analyzed using Y2H assay. The coding sequences of PpNAC25 and PpNAC1 were inserted into pGBKT7 and pGADT7. In Y2H assay, yeast containing PpNAC25 and PpNAC1 grew normally and turned blue in SD/-THLA+X-α-gal (Fig. 6a). Moreover, BiFC assay showed that the green fluorescence was only detected in tobacco leaves co-transformed with PpNAC25-ECN and PpNAC1-ENN (Fig. 6b). These results suggested that PpNAC25 interacted with PpNAC1.
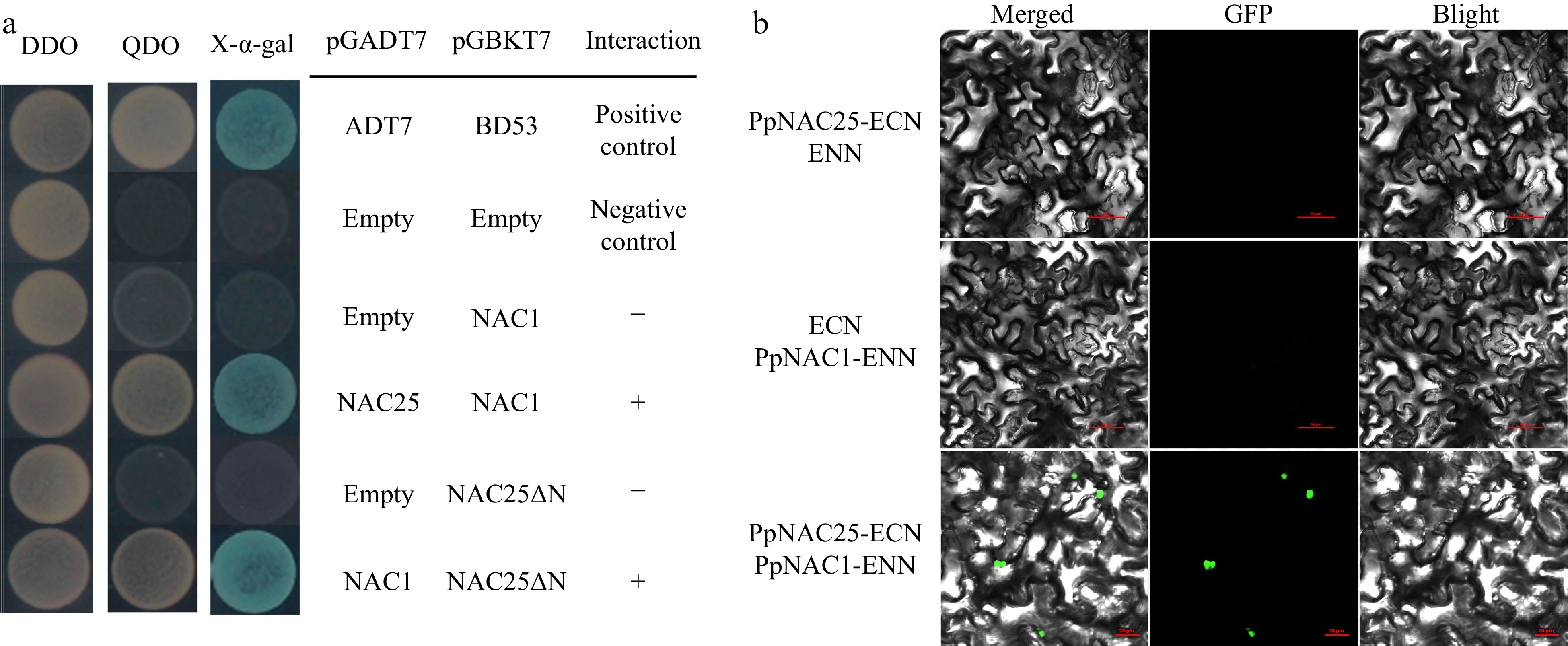
Figure 6.
Protein interactions of PpNAC25 and PpNAC1 were analyzed using (a) Y2H and (b) BiFC assay. DDO = SD/−TL. QDO = SD/−THLA.
-
In this study, we reported a NAC transcription factor PpNAC25, which showed a closed relationship with tomato NOR (no-ripening) (a well-known ripening-related gene) and positively regulated peach anthocyanin biosynthesis. NOR and its orthologous genes is showed to promote fruit ripening in several plants[44−47]. However, it is not well documented that NOR and its orthologous genes play their role in the regulation of anthocyanin biosynthesis. It is common that anthocyanin is rapidly biosynthesized during fruit ripening. This means that NOR may be involved in anthocyanin biosynthesis of fruit. In this study, the phylogenetic tree showed that PpNAC25 has a close relationship with NOR from tomato and was highly expressed in peach fruit. Meanwhile, overexpression of PpNAC25 in poplar upregulated the transcript level of genes involved in anthocyanin biosynthesis and transport and enhanced the anthocyanin accumulation in shoots. In addition, PpNAC1 is another homologous gene of tomato NOR and positively regulates anthocyanin biosynthesis of peach fruit[25]. All these results suggested that the NOR homologue gene PpNAC25 positively regulates the anthocyanin biosynthesis in peach.
The regulation of anthocyanin biosynthesis by NAC transcription factor is well studied in many plants. MdNAC42 increases anthocyanin accumulation in red-fleshed apple by interacting with MdMYB10[48]. MdNAC52 promotes the biosynthesis of anthocyanin and proanthocyanidin by activating MdMYB9 and MdMYB11[34]. LcNAC13 interacts with LcR1MYB1 and could bind to the promoter of anthocyanin biosynthesis genes during litchi fruit ripening[35]. In this study, PpMYB10.1 and PpMYB10.2, anthocyanin biosynthesis-promoting genes in peach fruit, were suggested as the targeted genes of PpNAC25. PpNAC1, a homologue gene of PpNAC25 in peach, enhances anthocyanin biosynthesis by directly activating the transcription of PpMYB10.1 in peach fruit[25]. These results suggested PpNAC25 increases anthocyanin biosynthesis probably through activating the transcription of PpMYB10.1 and PpMYB10.2 in peach.
It is well-known that only some peach cultivar fruits flesh is red to varying degrees at mature/ripening stage. However, NOR homologue gene PpNAC1, not only in red flesh cultivars 'DHP' but also in white flesh cultivar 'BF', shows a gradual increase in transcript level during the fruit development[25]. In our study, PpNAC25 was also expressed highly in peach fruit of 'QMH', a white flesh cultivar. These suggested that PpNAC1 and PpNAC25 are necessary but not sufficient for anthocyanin biosynthesis in peach flesh. There are two explanations for this phenomenon. Firstly, NOR homologue genes PpNAC1 and -25 should cooperate with other regulator genes and then activate the anthocyanin biosynthesis in peach flesh. BL is another NAC gene and specially expressed in red flesh cultivar 'DHP', which could interact with PpNAC1 and then promote anthocyanin biosynthesis[25]. In this study, our results demonstrated a protein interaction between PpNAC1 and PpNAC25. This suggested that a complex of NAC protein regulates anthocyanin biosynthesis in peach fruit. Secondly, the function of PpNAC1 and −25 may be repressed by anthocyanin repressors. It is common for plants to modulate pigmentation through anthocyanin repressors[49]. Until now, the reported repressors come from a wide variety of transcription factor families, including MYB, SPL, HD-ZIP, LBD, NAC, bZIP, and ARF[50−58]. In addition, miR828 and miR858 also play the role of repressors in anthocyanin biosynthesis by targeting the anthocyanin activator AtMYB113[59].
The two explanations are to some extent supported by two kinds of peach cultivars with blood flesh trait. Peach 'DHP' and 'WYX' show the blood flesh trait which is controlled by a single dominant locus at the top of LG5, while the blood-flesh trait of peach 'Harrow Blood' is controlled by a single recessive locus, designated bf (blood-flesh), which is mapped to the top of linkage group (LG) 4[24,60−64]. Therefore, with the exception of the positive regulator BL gene in 'DHP', it could be believed that anthocyanin negative regulator may play an important role in the anthocyanin biosynthesis of peach flesh. In our future study, we think that identifying the anthocyanin repressors is an important way to illustrate the phenotype of blood flesh in peach.
-
Anthocyanins are responsible for the red color of peach fruit which is one of the most important fruit quality traits attracting consumers. In this study, a NAC transcription factor, PpNAC25, showed a closed relationship with the well-known ripening-related gene NOR (no-ripening) and was highly expressed in peach fruit. Overexpressing PpNAC25 in poplar enhanced anthocyanin biosynthesis. PpNAC25 can target the anthocyanin-activator PpMYB10.1 and PpMYB10.2 and improve anthocyanin biosynthesis. Moreover, PpNAC25 interacted with PpNAC1, a homologous gene of NOR and also an anthocyanin-promoter. Our research provides evidence that PpNAC25 is a positive regulator of anthocyanin biosynthesis in peach fruit.
The work was conducted at the Henan Provincial Key Laboratory of Fruit and Cucurbit Biology and supported by the Joint Funds of the National Natural Science Foundation of China (U1804114), the Henan Province Outstanding Foreign Scholar Program (GZS2020007), and the Modern Agricultural Industry Technology of Henan Province (S2014-11-G02).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 List for primer used.
- Supplemental Fig. S1 The phylogenetic tree with all peach NACs and some NACs from different plants.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Geng M, Shao Y, Zhang M, Zheng X, Tan B, et al. 2022. Overexpression of peach NAC25 promotes anthocyanin biosynthesis in poplar shoots. Fruit Research 2:21 doi: 10.48130/FruRes-2022-0021 

Overexpression of peach NAC25 promotes anthocyanin biosynthesis in poplar shoots
- Received: 22 September 2022
- Accepted: 23 November 2022
- Published online: 28 December 2022
Abstract: Anthocyanins, a categories of metabolites derived from flavonoid biosynthesis pathway, are responsible for the red color of peach. Anthocyanin metabolism is modulated by a complicated regulatory network. In this study, our results demonstrated that PpNAC25 was a component of anthocyanin regulatory network in peach. PpNAC25 showed a closed relationship with the well-known ripening-related gene NOR (no-ripening) and was highly expressed in peach fruit. Overexpression of PpNAC25 in poplar resulted in a redder shoot tips compared with EV control. PpNAC25 overexpression upregulated the anthocyanin biosynthetic and transportation genes in transcript levels and then increased anthocyanin contents. In Y1H and Luc/Ren assay, PpNAC25 bound to the promoter of the anthocyanin-activator PpMYB10.1 and PpMYB10.2 and activated its transcript. Moreover, PpNAC25 formed a heterodimer with PpNAC1, an anthocyanin-regulator. Our researches provide evidence that PpNAC25 may be a positive regulator of anthocyanin biosynthesis in peach fruit.
-
Key words:
- Peach /
- Anthocyanin /
- NAC transcription factor /
- MYB 10.1.