










-
Plants produce large amounts of natural products with specialized functions, which are crucial to their development, defense responses and the interactions with the ecological environment[1]. Among these products, terpenes constitute an important and diverse category and play an important role in primary and secondary metabolism of plants. In primary metabolism, terpenoids such as gibberellin, cytokinin, auxin and carotenoid are widely involved in plant growth and development[2]. Defense against biological and abiotic stress is the most important function of terpenoids, which are highly diverse and species-specific as secondary metabolites[3]. In human society, terpenes are widely utilized in flavor, perfume, medicine and ecological pesticide[4−6]. Although terpenes have considerable economic value, it is still a major challenge to improve their utilization efficiency, especially for wild resources. Monoterpenes, as C10 terpenes, typically colorless, lipophilic, volatile, and antibacterial, are the source of the unique fragrance and flavor of essential oils[7,8]. Monoterpenes consist of isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP), which are isomeric 5-carbon structural units, derived from the mevalonate (MVA) pathway in the cytosol/peroxisome and the methyl erythritol phosphate (MEP) pathway in the plastid[9]. Then, one IPP and one DMAPP are catalyzed to form geranyl diphosphate (GPP). The last step is to convert GPP into a monoterpene parent scaffold via the monoterpene synthase.
The MEP pathway is mainly controlled by 1-deoxy-d-xylose 5-phosphate synthase (DXS) and 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR), which has been known as the rate-limiting enzyme of the MEP pathway in plants[10,11]. DXR is an enzyme that catalyzes 1-Deoxy-D-xylulose-5-phosphate to 2-C-Methyl-D-erythritol-4-phosphate, the second step of the MEP pathway[12]. It requires the participation of cofactors NADPH, Mn2+, Co2+, or Mg2+ to convert DXP into an important precursor MEP for terpenoid synthesis[13]. Since the first DXR was cloned from Escherichia coli, the gene has been found in several other species and studied for its function in terpenoid synthesis[14]. There is transport peptide-guided DXR localization in plastids at the N-terminal, and it has a conservative Cyr-Ser–(Ala/Met/Val/Thr) motif; there is a proline enrichment area P (P/Q) PAWPG (R/T); two DXR functional binding motifs (LPADSEHSAI, NKGLEVIEAHY), and two highly conservative NADPH binding motifs (GSTGS [I/V] GT and LAAGSN [V/I] T)[15,16]. Overexpression of key rate-limiting genes in the terpene biosynthesis pathway is an effective way to improve plant terpenoid yield. Interestingly, DXR function in terpene biosynthesis seems to be species-specific. Overexpression of the DXR effectively promotes monoterpene synthesis of peppermint essential oil, terpene indole alkaloids in Catharanthus roseus, artemisinin in Artemisia annua, and taxene in transgenic Arabidopsis thaliana[17−21]. Importantly, DXR expression is significantly correlated with the cumulative distribution of monoterpenes[22−24]. In contrast, the expression level of DXR does not affect the production of terpenes in Solanum lycopersicum, Oryza sativa, or Lavandula angustifolia[11,25,26].
Litsea cubeba, belonging to the Lauraceae family, is primary woody oil tree in China[7]. L. cubeba fruits are rich in essential oil (3%−4%), which is widely used in essence, spices, medicine, cosmetics, industrial raw materials, and other fields[4,27−29]. For example, L. cubeba essential oil (LcEO) plays an important role in the synthesis of vitamin E/K/a and saponin ketone, which acts as a superior food-flavoring agent[30]. Meanwhile, LcEO has broad-spectrum antibacterial activities and contributes to improving the treatment of coronary heart disease and other cardiovascular diseases[31,32]. The components of LcEO include terpenes, higher alcohols, and organic acids, depending on the maturity of organs[4,33]. Geranial, neral, pinene, linalool, and other monoterpenes make up about 96–98% of the LcEO[7]. In our previous study, LcGPPS.SSU1 and LcTPS42 were demonstrated as the structure genes involved in the monoterpene biosynthesis pathway, and transcription factor including ERF and MYC2, effectively improving the production of monoterpenes[7,34−37].
The terpene biosynthesis pathway has been characterized by multi-omics analysis of L. cubeba. However, to date, few strategies have been reported to improve the yield of LcEO by regulating the key genes encoding MEP pathways. In this study, we firstly investigated the full-length cDNA and the promoter sequence of LcDXR. The expression patterns were analyzed in different tissues and different developmental stages in fruits. Instantaneous overexpression of LcDXR in L. cubeba was also found to increase monoterpene content. In addition, the enhanced monoterpene biosynthesis capacity was tested by overexpressing LcDXR in tobacco. The results indicated that LcDXR played a key role in regulating the formation of monoterpenes.
-
Different tissues of L. cubeba, including flowers, leaves, stems, roots, and fruits at different development stages (60, 90, 120, and 150 d after flowering, DAF), were collected from a farm in Hangzhou City, Zhejiang Province, China (30°27′94′′ N, 119°58′43′′ E), then quickly frozen, and stored at −80 °C. For Agrobacterium-mediated transformation, Nicotiana benthamiana was grown in a growth chamber at 26 °C, with a photoperiod of 16 h/8 h.
LcDXR cloning and sequence analysis
-
The complete sequence of LcDXR was identified based on the L. cubeba genome and the DXR conserved domain (PF08436) using HMMER (
www.hmmer.org ). The full-length amino acid sequences of LcDXR and DXR from other plants were aligned using ClustalW software (www.ebi.ac.uk/Tools/msa/clustalo ). Maximum-likelihood phylogenetic tree was constructed based on CIPRES (www.phylo.org ) with the JTT model using 1,000 bootstrap replicates.RNA extraction and quantitative real-time PCR (qRT-PCR)
-
To analyze the expression profiles of LcDXR, the total RNA of different tissues was extracted and reversed to the first strand cDNA synthetase as previously described[34]. For qRT-PCR, ubiquitin-conjugating (UBC) gene was used as a reference gene[38]. The comparative cycle threshold (Ct) method was used to analyze the expression level of LcDXR in different tissues and stages. The primer sequences are listed in Supplemental Table S1. Three biological replications were made for each gene.
Expression of recombinant LcDXR in Escherichia coli and enzymatic assays
-
To determine the enzyme activity of LcDXR, the full-length CDS of LcDXR was inserted into pET28a (+) vector. The recombinant plasmid was transferred to E. coli ER2566 (Weidi, China), and the recombinant protein was induced and purified as described previously. The recombinant protein was detected by SDS-PAGE and Western blot analysis.
For in vitro enzyme activity assays, 20 μg DXR purified protein was incubated with 50 mM Tris-HCl, 5 mM MgCl2, 1.23 mM NADPH, and 0.25M DXP at 30 °C for 15 min and the time course was measured by monitoring the formation of the generated NADPH in a 340 nm spectrophotometer[14]. DMSO was added to replace the DXP in the control reaction. Three technological replications were carried out.
Transient overexpression of LcDXR in L. cubeba
-
To investigate the function of LcDXR in monoterpene biosynthesis, the transient overexpression of L. cubeba was constructed as in a previous study[34]. Two-month-old tissue culture seedlings of L. cubeba were generated for transient overexpression. The completed CDS of LcDXR was cloned into the overexpression vector pCambia1300S. A. tumefaciens LBA4404 strains containing empty vector or pCambia1300S-LcDXR were cultured and then infiltrated by vacuum into sterility seedling leaves under the same growth condition. The seedlings were then placed on MS medium and cultured at 25–27 °C with a photoperiod of 16 h light/8 h dark for 40–72 h. Leaves were collected for GC-MS and qRT-PCR analysis according to a previous study[7]. The target compounds were identified by comparing their mass spectra with those in the NIST08 library.
Overexpression of LcDXR in tobacco
-
To further confirm the function of LcDXR, pCambia1300S-LcDXR was transformed into Agrobacterium EHA105 strains for tobacco transformation[39]. After selection with Hygromycin B and PCR amplification using LcDXR specific primer, ten positive LcDXR transformation lines were obtained. qRT-PCR was performed to investigate the expression levels of LcDXR as described above, and actin was used as the reference gene[36]. The effect of introducing LcDXR on monoterpene production was determined by GC-MS analysis.
-
The complete LcDXR open reading frame was 1,413 bp with a 5' noncoding region of 34 bp and a 3' noncoding region of 53 bp, encoding a protein containing 470 amino acid residues. LcDXR showed high sequence identity (> 85.56%) with previously reported DXRs. In the phylogenetic tree of DXRs, LcDXR grouped near to DXRs from Cinnamomum kanehirae and C. camphora, which all belong to the Lauaceae family (Fig. 1a). To further investigate sequence variants among DXRs, we conducted multiple sequence alignment of DXR amino acid sequences and found that LcDXR has the conserved domain of LPADSEHSAI and NKGLEVIEAHY for binding DXP and two NADPH binding motifs, GSTGSIGT and LAAGSNVT (Fig. 1b).
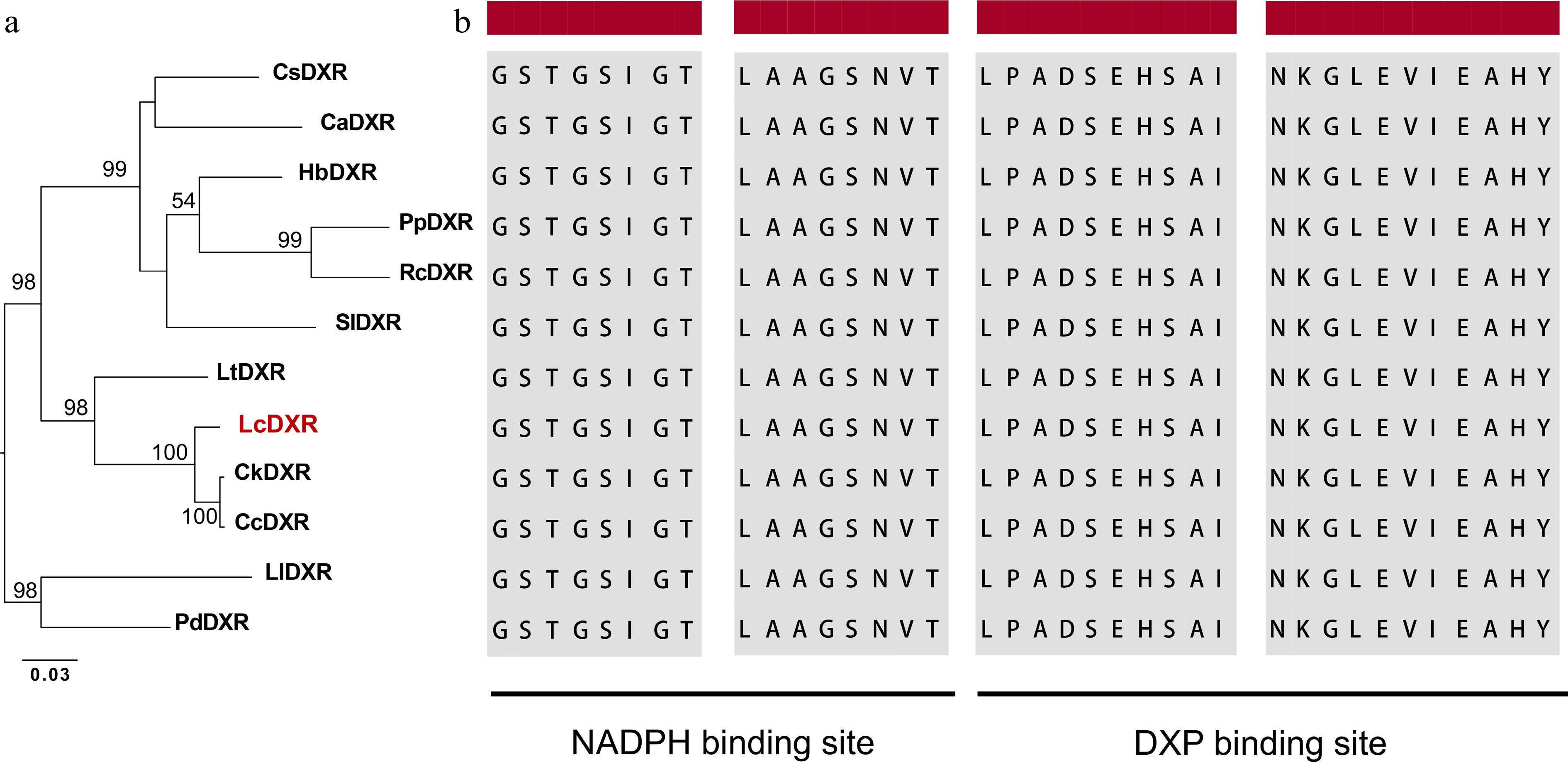
Figure 1.
Sequence analysis of the LcDXR. (a) Maximum-likelihood tree of DXR protein sequences, including LcDXR and other DXRs. The following protein sequences were used for the analysis: CsDXR (XP_028055701), CaDXR (XP_027123133), HbDXR (XP_021669849), PpDXR (XP_007210092), RcDXR (XP_024173969), SlDXR (AAK96063), LtDXR (UOO00992), CkDXR (RWR93981), CcDXR (AOW69227), LlDXR (AHJ57307), PdDXR (XP_008787962). (b) Sequence alignment of DXRs. The black background indicates the NADPH binding domain and the DXP binding domain.
Tissue-specific expression patterns of LcDXR in L. cubeba
-
It has been reported that the production of essential oil increases with the development of fruits in L. cubeba[40,41]. To study the expression pattern of LcDXR, qRT-PCR was performed on the roots, stems, leaves, flowers, and different development stages of fruits of L. cubeba. As the results shown in Fig. 2a, LcDXR was widely expressed in all tissues and highly expressed during fruit ripening. Interestingly, expression peaks were detected at 60 and 120 DAF of fruits, and the biosynthesis of essential oil reached its peak at the same time[40]. These results indicated that LcDXR played an important role in primary and secondary metabolism in all tissues of L. cubeba. It produced a prominent effect in the accumulation of monoterpenes during the early and middle fruit developmental stages.
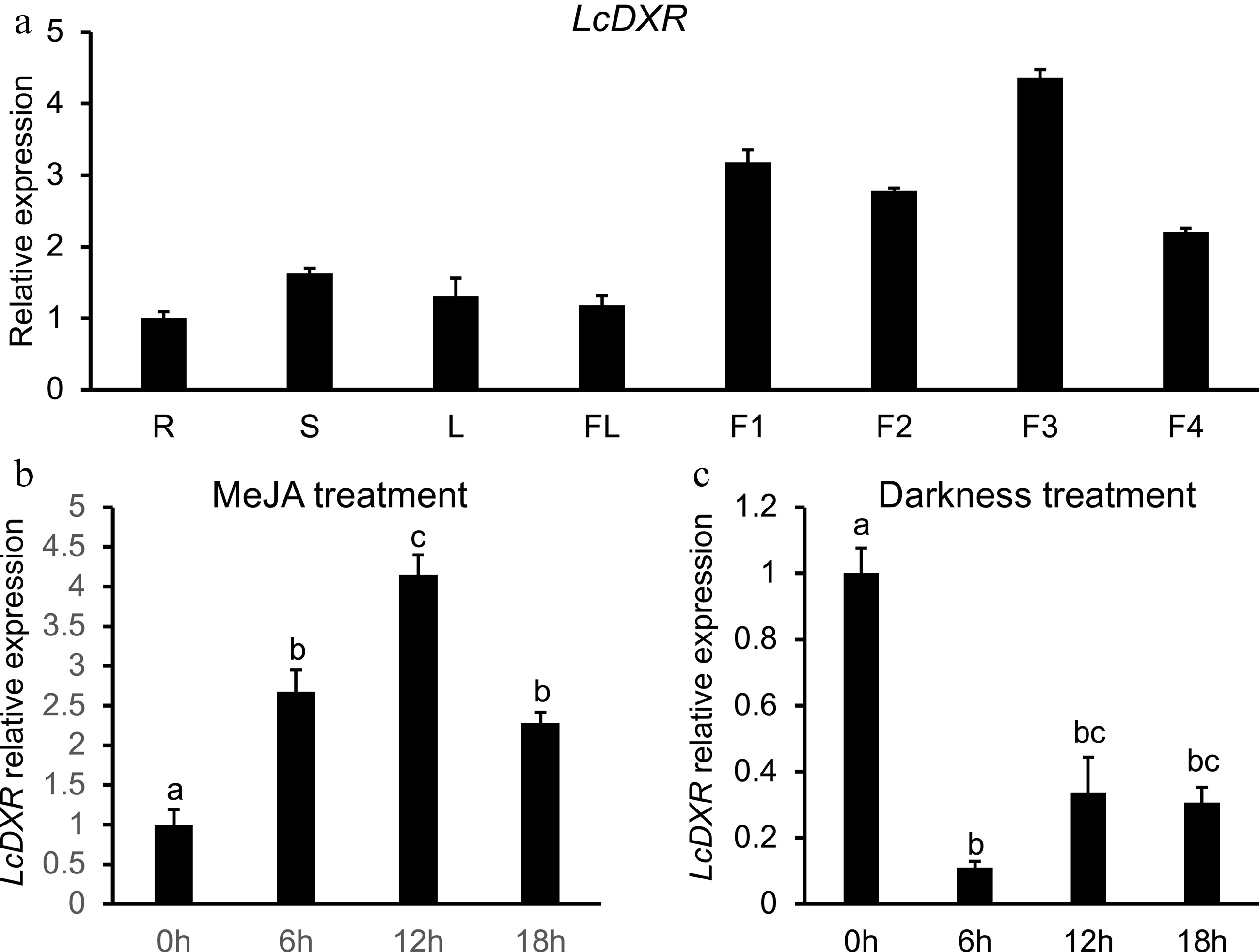
Figure 2.
Tissue specific expression patterns of LcDXR. (a) The transcript levels of LcDXR in different tissues of L. cubeba. R: roots; S: stems; L: leaves; FL: flower; F1, fruits at 60 DAF; F2, fruits at 90 DAF; F3, fruits at 120 DAF; F4, fruits at 150 DAF. (b) The expression levels of LcDXR were induced by MeJA in 60 DAF fruits. (c) The expression levels of LcDXR were repressed by darkness in 60 DAF fruits. Error bars represent ± SD from three biological repeats. The different letters above the bars represent the significant difference (P < 0.01).
In addition, the promoter region (−2,000 bp) of LcDXR was amplified and many cis-elements were identified, such as MYB, MYC, G-box, ABRE and other motif; ABA, SA, and MeJA signal transmission were noted (Supplemental Fig. S1). In our previous study, the biosynthesis of terpenes was found to respond to darkness and MeJA treatment[34,37]. Corresponding to the peak of monoterpene biosynthesis in L. cubeba at 12 h after MeJA treatment, LcDXR reached the peak of expression at the same time, which increased 4-fold compared with 0 h (Fig. 2b). In fact, the inhibition of darkness on terpene biosynthesis is involved in energy metabolism, light and other factors. Meanwhile, reduced expression of LcDXR was found in L. cubeba grown under the dark conditions for 48 h (Fig. 2c). These results confirmed that LcDXR was induced by MeJA treatment and repressed by darkness.
Enzyme activity of LcDXR
-
To determine the enzymatic activity of LcDXR, recombinant pET28a plasmid (~63 kDa) with the CDS of LcDXR was expressed in E. coli. According to SDS-PAGE and western blot analysis, the purified LcDXR recombinant protein had a molecular weight similar to that expected (Fig. 3a, b). The catalytic reaction of DXR requires the conversion of NADPH into NADP+. NADPH has a robust ultraviolet absorption peak at A340 and NADP+ does not, while the light absorption value of NADPH at A340 decreases during the catalytic reaction[12]. Accordingly, when LcDXR was added to the enzyme reaction system, NADPH was consumed at A340 rapidly decreased, and the reaction rate slowed in the later stage (Fig. 3c). These results suggested that LcDXR had the catalytic activity.
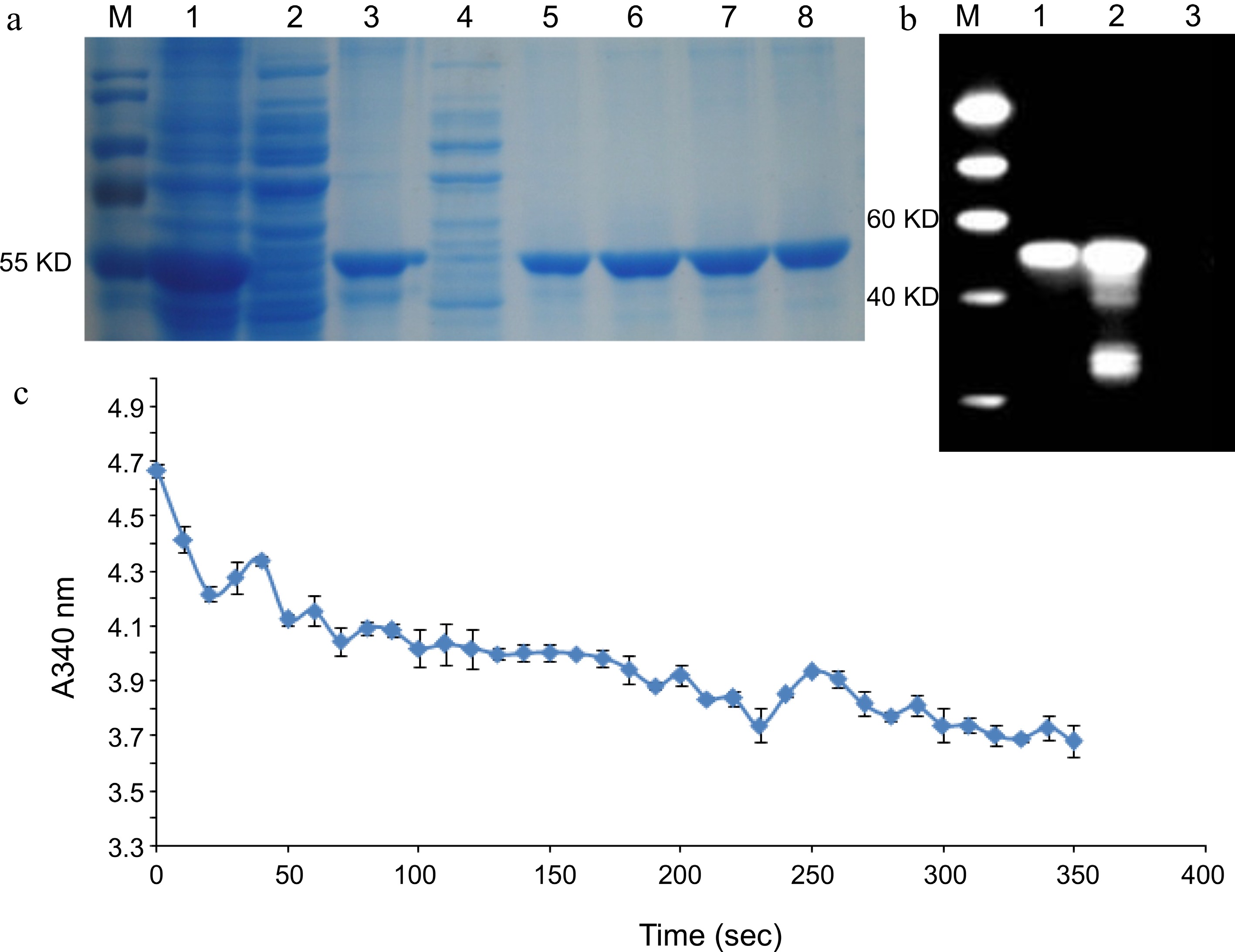
Figure 3.
Enzyme activity of LcDXR. (a) SDS-PAGE analysis of recombination protein with His tag. M: Protein Marker, 1: Insoluble protein before purified, 2: Cell lysate supernatant, 3: Transmission fluid, 4: Washing fluid, 5−8: Elution fluid with protein. (b) Western blot analysis of recombinant LcDXR protein. M: Protein Marker, 1: The inclusion protein, 2: Total protein induced in pET–LcDXR2/Arctic Express, 3: Washing fluid. (c) Enzyme activity assay of recombinant LcDXR. NADPH has a robust ultraviolet absorption peak at A340 and NADP+ does not, while the light absorption value of NADPH at A340 decreases during the catalytic reaction. Data represent the mean ± SEs of three biological replicates.
Transient overexpression of LcDXR increases the terpenes biosynthesis in L. cubeba
-
A rapid and efficient transient overexpression system provides a method to detect the gene function in L. cubeba without stable transformation system. To investigate whether the overexpression of LcDXR contributed to the monoterpene accumulation, transient overexpression analysis was performed in L. cubeba leaves and three independent transgenic lines were conducted. Among the LcDXR overexpression lines, the expression levels of LcDXR were significantly increased by 9.6-, 7.1-, and 7.8-fold compared to controls (Fig. 4a). The introduction of LcDXR accelerated the synthesis of monoterpenes, showing an increase of up to 4.0-fold compared with the control (Fig. 4b). It is noteworthy that the LcDXR overexpression activated the expression of rate-limiting genes involved in the terpene biosynthesis pathway, including LcDXS[7], LcGPPS.SSU1[37], and LcTPS42[7], which were increased 1.5- to 4.0-fold in overexpressed leaves (Fig. 4c). However, compared to controls, a smaller change seen in 3-hydroxy-3-methylglutaryl CoA synthetase (LcHMGS) was investigated, which involved the MVA pathway in the cytoplasm[2]. In summary, the transient overexpression of LcDXR provided preliminary evidence that LcDXR is the key gene for monoterpene biosynthesis of L. cubeba.
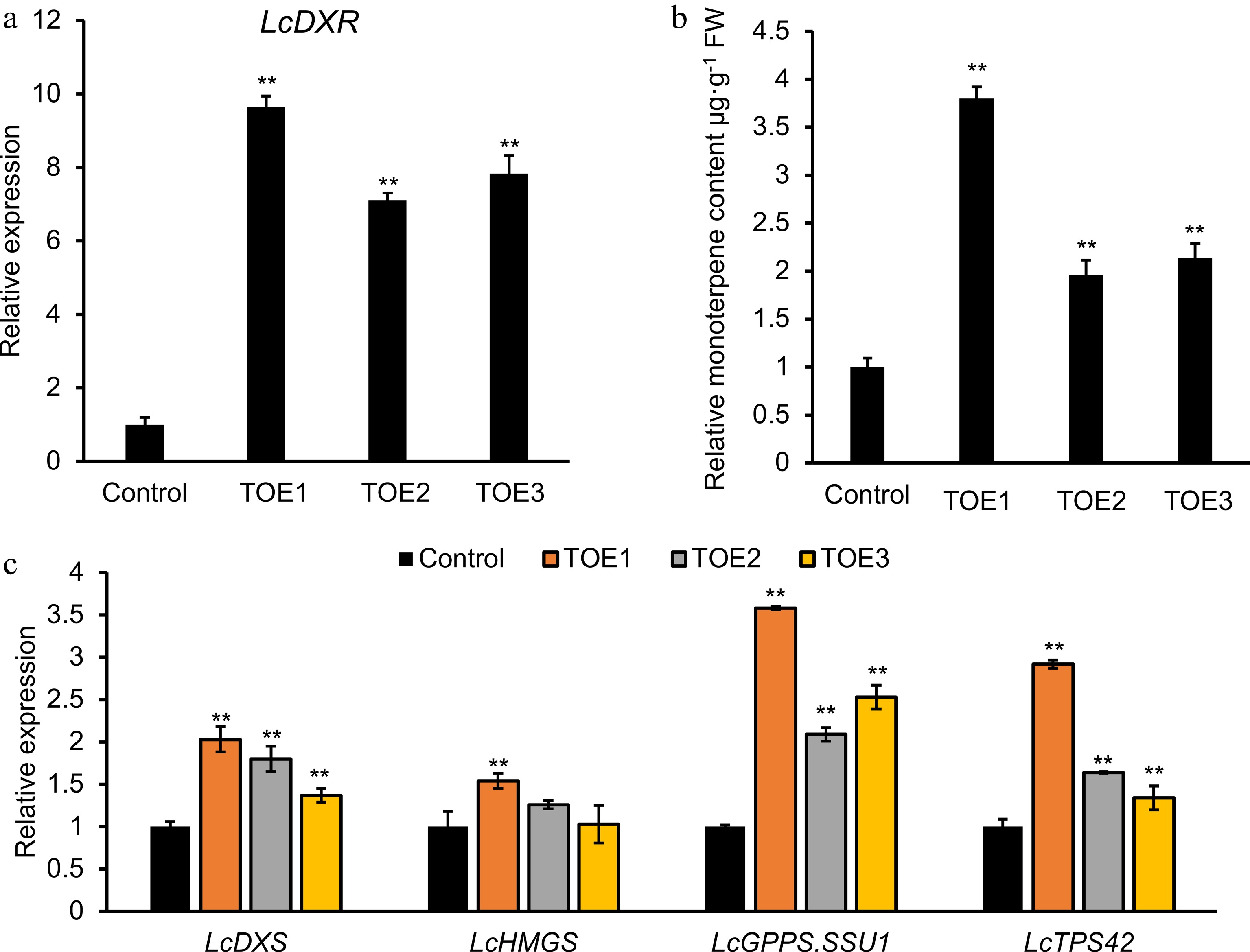
Figure 4.
Transient overexpression of LcDXR in L. cubeba leaves. Leaves were infiltrated with LcDXR under the control of the Cauliflower mosaic virus 35S promoter, and three independent transient overexpression lines (TOE) 1-3 were conducted and grown for 2 d, respectively. (a) Relative expression level of LcDXR in transient overexpressed L. cubeba. (b) The content of monoterpenes in LcDXR transient overexpressed L. cubeba. (c) Relative expression level of key genes involved in MVA and MEP pathway in LcDXR transient overexpressed L. cubeba. Data represent the mean ± SEs of three biological replicates. **, P < 0.01.
Overexpression of LcDXR promotes monoterpene biosynthesis in tobacco
-
To further understand the function of LcDXR, we constructed transformation tobacco lines carrying pCambia1300S-LcDXR vector. Ten independent transgenic lines were identified through hygromycin B selection and PCR amplification. The transcriptional levels of LcDXR in stable overexpression lines (SOE) 1, SOE5, and SOE7 were the highest and thus were further used to evaluate monoterpene content and the expression of key genes at the 2-month stage (Fig. 5a). As the result shown in Fig. 5b, LcDXR transgenic lines had no obvious morphological changes. The content of monoterpenes collected from leaves was positively correlated with the transcriptional level in different transgenic lines. Compared to the control, the content of limonene, α-pinene, eucalyptol, linalool, terpineol and camphor were significantly increased by LcDXR overexpressed. In the leaves of SOE1, SOE5 and SOE7 transgenic tobacco line, limonene was increased from 0.2 μg·g−1 towards 1.9 μg·g−1 (Fig. 5c). Linalool, terpineol and camphor, which were not detected in the control, showed the content of 0.8 to 0.5 μg·g−1 in pCambia1300S-LcDXR lines. This result suggested that although tobacco contained many kinds of terpene synthases, the lack of substrates restrict the diversity and yield of monoterpenes.
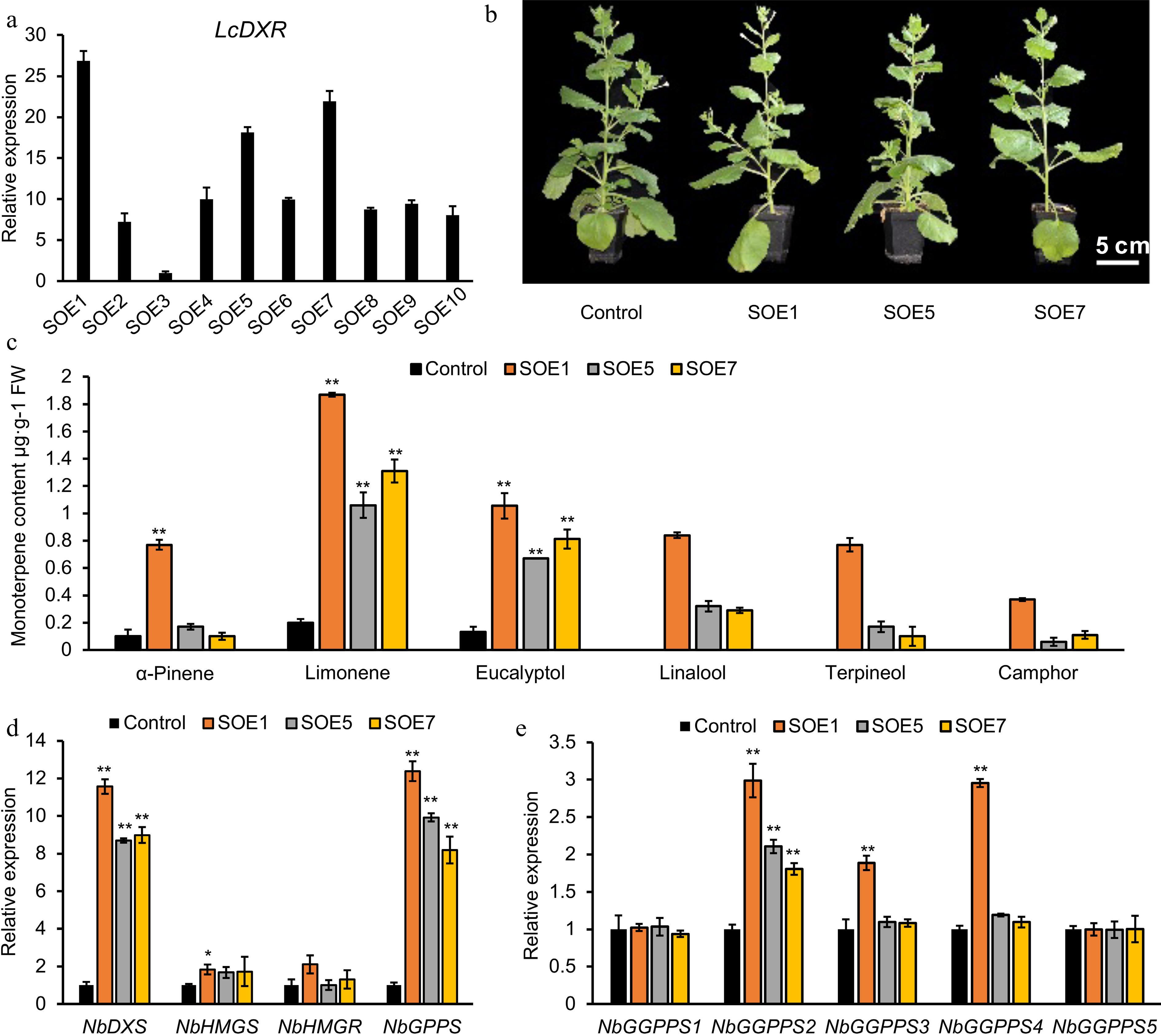
Figure 5.
Monoterpene production was increased by overexpression of LcDXR in transgenic tobacco. (a) Relative expression of LcDXR in ten independent stable overexpression lines (SOE), respectively. (b) The content of monoterpenes in LcDXR transgenic tobacco. (c) Different compounds of monoterpenes in transgenic tobacco. (d) Relative expression levels of key genes involved in MVA and MEP pathway. Data represent the mean ± SEs of three biological replicates. *, P < 0.05; **, P < 0.01.
Furthermore, the expressional level of rate-limiting genes in the MEP pathway encoding DXS was upregulated approximately 11-fold in transgenic leaves compared to controls (Fig. 5d). Similarly, as the key gene of the direct precursor for the synthesis of monoterpene, NbGPPS was upregulated 12-fold. On the contrary, the expression levels of NbHMGS were slightly affected by LcDXR through the crosstalk mechanism, and those of NbHMGR unchanged. As with monoterpenes, diterpenes are derived from the MEP pathway and located in the chloroplast. Thus, the expression of GGPPSs, the direct precursor synthase gene of diterpenoids, were also analyzed. There are five GGPPS genes in tobacco, among which only NbGGPPS2 significantly increased approximtely 3-fold in all three SOE lines; NbGGPPS3 and NbGGPPS4 were only increased in SOE1 (Fig. 5e). In summary, introducing LcDXR activated the metabolic flux of the monoterpene biosynthesis pathway.
-
According to previous studies, precursor supply is a limiting factor for monoterpene biosynthesis through the plastid MEP pathway[14,20]. During the process of monoterpene precursor formation in plants, DXR, which catalyzes the conversion of DXP into MEP, is a key rate-limiting enzyme of the MEP pathway[14]. IPP and DMAPP synthesized by the MEP pathway mainly contribute to monoterpene biosynthesis. LcEO, containing more than 90% monoterpenoids, has important economic value in commercial and industrial fields. Therefore, L. cubeba may be a potentially model plant for studying monoterpene biosynthesis[7]. Breaking through the bottleneck of the MEP pathway has been one of the most central challenges for metabolic engineering of monoterpene with the extensive commercial development of plants essential oil[10]. Although genes involved in the terpenoid synthesis pathway have been identified in L. cubeba, functional analysis to determine the rate-limiting genes is still absent.
Numerous previous studies conducted expression profiling of the DXR gene in different plant species and tissues, such as petals and stamens of R. rugosa and Cymbopogon winterianus, roots of Salvia miltiorrhiza, and leaves of peppermint, which have reported a higher level of expression in the main site of isoprenoid biosynthesis[11,12,14−17,20−26,42]. In C. camphora, CcDXR showed the highest expression in flowers, subsequently followed by leaves and stems, respectively[43]. In L. cubeba fruits, the transcript accumulation of LcDXR showed a direct correlation with monoterpene biosynthesis (Fig. 2). In our study, LcDXR was expressed in all tissues and up-regulated during fruit development, suggesting that LcDXR played a key role in the precursor formation of monoterpenes in fruits, which served as factories for essential oil production, similar to what has been reported in P. ginseng and C. roseus[44,45]. Notably, the highest expression of LcDXR was observed at 60 DAF and 120 DAF when the essential oil entered the rapid production phase (Fig. 2). Previous studies have reported that genes related to MEP pathway are highly expressed in photosynthetic tissues[7]. In fact, the MEP pathway is located in the chloroplast and responsible for phytohormones, chlorophyll, which is closely related to photosynthesis.
Key enzymes catalyze the synthesis of terpenoid precursors and the formation of various intermediates in the pathway of terpenoid biosynthesis. Thus, regulating the expression of these key enzyme genes to increase the content of downstream target products is an important way for the regulation of terpenoid biosynthesis[46]. Increasing the expression of key genes in the metabolic pathway may also affect the metabolic flux of the pathway through feedback regulation or crosstalk mechanism[47]. Overexpression of rate-limiting genes involved in the terpene precursor biosynthesis pathway is an effective strategy to improve terpene production, which has been conducted in several plants including C. roseus, W. somnifera, A. thaliana, Cucumis melo, Solanum lycopersicum, and Lilium[6,10,20,22,44]. Importantly, DXP is the precursor of not only MEP but also thiamine and pyridoxine[20]. As a branch point of carbon flow in the MEP biosynthesis pathway, DXR is a key rate-limiting enzyme in that pathway and an effective target for terpenoid synthesis regulation in plants[48]. However, the regulation of the DXR gene in plants is species-specific. In particular, in peppermint (Mentha × Piperita), an improvement of 44% in oil yield compared to WT was detected in DXR transgenic lines[21]. In contrast, overexpression of DXR had no effect on lavender[25]. Carotenoid accumulation during tomato fruit ripening had no obvious relationship with DXR expression[11]. In addition,
SlDXR did not influence the content of IPP or DMAPP in Synechococcus leopoliensis[49]. In our study, the concentration of monoterpenes was significantly enhanced in L. cubeba leaves overexpressing LcDXR compared to the control (Fig. 4). In a previous study, high levels of ectopic expression of LiDXS and LiDXR induced sclareol biosynthesis without altering growth characteristics and ecological habits[50]. It is generally considered that DXS catalyzes the rate-limiting step of the MEP pathway; nevertheless, DXS-overexpression-based metabolic engineering strategies are not sufficient to overcome the limitations of utilizing the MEP pathway for high-yield terpene production in tobacco[47]. In this study, overexpressing LcDXR in tobacco was effective in the activating of genes involved in terpene biosynthesis and leaf monoterpene production (Fig. 5). In fact, although tobacco has the potential to synthesize various monoterpenes, the lack of corresponding substrates limits the monoterpene synthesis in tobacco. Therefore, it is an effective way to improve the terpene content of tobacco by introducing the key genes of highly efficient synthetic substrates through metabolic engineering. It is worth noting that the MEP pathway is not only a precursor pathway for monoterpenes, but also for diterpenoid biosynthesis. Diterpenoids such as chlorophyll and carotenoids are crucial for plant growth and development. Overexpression of designed NtGGPPS1 in ntggpps1 mutants increases photosynthetic efficiency and biomass in tobacco leaves[51]. However, NbGGPPS1 was not affected in LcDXR-OE lines while GGPPS2 was significantly increased. In fact, the phenotype was not changed in LcDXR-OE lines. In carrots (Daucus carota), DXS limits the production of chlorophyll in roots and leaves, while DXR appears to have a small regulatory effect[52]. Obviously, the regulatory effect of MEP pathway on diterpenoids such as chlorophyll, and the synergistic relationship between photosynthesis and terpene synthesis need further research. The monoterpene synthesis pathway is strong in plasticity through feedback and crosstalk mechanisms[53−56]. Previous studies have shown that the increase of downstream products can up-regulate the expression level of upstream genes involved in precursor synthesis pathway through a feedback regulation, to accelerate the metabolic flux of MEP and MVA pathways[57,58] DXS, as the first gene of the MEP pathway, is the key target for downstream product feedback regulation of the MEP pathway. In our study, overexpression of DXR both promoted the expression of downstream genes by improving the synthesis of precursors and activated the expression of DXS genes through the feedback regulation, which together contributed to the increase in monoterpene synthesis Figs 4 & 5). It is worth noting that the key genes of the MVA pathway, HMGR and HMGS, had smaller change in transgenic L. cubeba and tobacco. This result indicated that there was a weaker metabolic flux between the MEP and MVA pathways in LcDXR overexpressed lines. The increased metabolic flux in the MEP pathway regulated by LcDXR overexpression was not detected in the MVA pathway. In fact, the MEP or MVA pathways tend to preferentially synthesize certain metabolites in different plants[36]. Monoterpene, as the main terpenoid (accounting for up to 98% of all terpenoids) in L. cubeba, is located in the chloroplast and derived from MEP pathway. In addition, we hypothesize that the contribution of LcDXR to monoterpene products is universal, regarding its identical functions to monoterpene biosynthesis in L. cubeba and tobacco.
-
In conclusion, we functionally characterized LcDXR, the key gene of the MEP pathway, which played an important role in the biosynthesis of monoterpenes in L. cubeba. Expression analysis suggested that it is the crucial gene to control the production of essential oil monoterpenes in fruits. Enzyme activity assay showed that LcDXR catalyzed DXP with NADPH as a cofactor. Overexpression of LcDXR significantly increased the contents of monoterpenes in L. cubeba and tobacco. Our results indicated that LcDXR could be employed as a universal target gene for metabolism regulation to improve the content of monoterpenes in plant tissues.
This work was supported by the Key Scientific and Technological Grant of Zhejiang for Breeding New Agricultural Varieties (2021C02070-3) and the National Natural Science Foundation of China (32101561).
-
Yicun Chen is the Editorial Board member of Forestry Research. She was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and her research groups.
- Supplemental Fig. S1 Promoter sequence of LcDXR.
- Supplemental Table S1 Primer list.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhao Y, Liu Y, Chen Y, Gao M, Wu L, et al. 2023. Overexpression of 1-deoxy-D-xylulose-5-phosphate reductoisomerase enhances the monoterpene content in Litsea cubeba. Forestry Research 3:11 doi: 10.48130/FR-2023-0011 

Overexpression of 1-deoxy-D-xylulose-5-phosphate reductoisomerase enhances the monoterpene content in Litsea cubeba
- Received: 07 February 2023
- Accepted: 18 April 2023
- Published online: 24 April 2023
Abstract: Monoterpenes are important components of plant essential oils and have long been used as raw materials for spices and food flavorings. Litsea cubeba is an economically aromatic plant species, the fruits of which produce essential oil with monoterpenes as the dominant components. As a branch point of carbon flow in the methyl erythritol phosphate (MEP) biosynthesis pathway, 1-deoxy-D-xylo-5-phosphate reductoisomerase (DXR) is a key rate-limiting enzyme that catalyzes the MEP pathway’s second committed step. Therefore, DXR has become an effective regulation target to improve the biosynthesis of plant monoterpenes. In this study, we identified a DXR from L. cubeba, which was highly expressed in fruits, induced by MeJA and repressed by darkness. An enzyme assay showed that recombination LcDXR protein catalyzed DXP with NADPH as the cofactor. Transient overexpression of LcDXR significantly increased the content of monoterpenes in L. cubeba. Furthermore, LcDXR-overexpressing tobaccos were conducted and showed almost 5.9-fold increase in monoterpenes production, including limonene, α-pinene, eucalyptol, linalool, terpineol and camphor. Overexpression of LcDXR activated the metabolic flux of monoterpene biosynthesis through crosstalk and feedback mechanism. In addition, LcDXR-overexpressing tobaccos had no effect on phenotype of transgenic tobaccos. Our results demonstrate that LcDXR is a critical regulator of the monoterpene production in L. cubeba and other plants.
-
Key words:
- Litsea cubeba /
- Monoterpene /
- DXP reductoisomerase /
- MEP pathway