










-
The widespread consumption of tea, which is derived from the tender shoots of Camellia sinensis and related species, can be attributed to its captivating flavors[1]. Furthermore, tea is rich in various biologically active metabolites, such as flavonoids, theanine, and caffeine, which are known to confer a range of health benefits[2−4]. Among these metabolites, caffeine (1,3,7-trimethylxanthine) is the dominant purine alkaloid in most teas and is globally recognized for its ability to stimulate the central nervous system[5]. In addition to caffeine, theobromine (3,7-dimethylxanthine) and theacrine (1,3,7,9-tetramethyluric acid) are also typical purine alkaloids found in tea plants[6,7]. The content of these purine alkaloids varies considerably among different tea accessions, with most containing caffeine at concentrations of 25.0−45.0 mg/g and theobromine at 2.4−4.3 mg/g[5,8].
The excessive consumption of caffeine can have adverse effects, such as insomnia, migraines, and changes in intraocular pressure[9,10]. This has led to a growing demand in the market for naturally low-caffeine or decaffeinated tea products. In response to this demand, researchers have turned their attention to identifying unique natural variations within tea plant species that accumulate low levels of caffeine or none at all[11]. One such species, Camellia ptilophylla, possesses a natural variant that accumulates theobromine only[12]. This variant represents an efficient and unique raw material for the development of low-caffeine tea products through breeding programs.
The biosynthesis pathway of caffeine has been extensively studied[13,14]. Caffeine is synthesized from xanthosine[15] in three methylation steps, by methyltransferase (XMT), 7-methylxanthine methyltransferase (MXMT) and tea caffeine synthase (TCS), and one nucleoside removal step[16,17]. TCS1, which catalyzes 1-N and 3-N methylation of 7-methylxanthine, is essential to catalyze conversion of theobromine to caffeine. Prior studies have revealed that TCS1 in most tea cultivars exhibits higher enzymatic activity and expression levels compared to C. ptilophylla, and furthermore, the TCS1 enzyme in C. ptilophylla solely possesses 3-N methyltransferase activity[18−20]. Thus, the low-caffeine levels in C. ptilophylla could be related to the reduced capacity of TCS1. Recent studies have documented that transcription factors (TFs) also play a critical role in caffeine biosynthesis[21−24], such as CsMYB184 and CsNAC7[16,25]. Li et al. have proven that CsMYB184 was significantly downregulated in C. ptilophylla compared to other tea cultivars[16]. Therefore, the mechanism underlying caffeine deficiency in C. ptilophylla still remains elusive and requires further investigation.
Catechins, which confer bitter and astringent tastes, are also important indicators of tea quality[26,27]. EGCG is the primary catechin monomer found in C. sinensis, with an average content of 94.6 ± 12.6 mg/g, whereas GCG is typically found in much smaller amounts (0.9 ± 2.2 mg/g)[28,29]. However, in C. ptilophylla, GCG is the main catechin and can reach 93.85 ± 0.96 mg/g[30]. Despite the extensive literature on tea catechins[31], there remains a knowledge gap regarding the molecular biology of different catechin monomers that accumulate in C. ptilophylla.
In this study, the content of purine alkaloids in a C. ptilophylla population (32 plants) were quantified by HPLC. Two C. ptilophylla plants, one was caffeine-free containing only theobromine (TB), and the other contained both theobromine and caffeine (CAF) were used to analyze content of catechins and theanine. RNA-seq was used to investigate the different mechanisms of purine alkaloid and catechins accumulation in TB and CAF.
-
Samples containing one bud and two leaves each were collected from 32 individual C. ptilophylla plants in June 2022 in the Conghua tea garden (Guangzhou, Guangdong province, China), and the samples were stored at −80 °C for chemical composition analysis and RNA extraction.
Chemicals
-
Caffeine, theobromine, theacrine and theophylline standards were bought from Beijing Weiye Research Institute of Metrology and Technology (Beijing, China). Magnesium oxide was purchased from FUCHEN Chemical Reagents LTD (Tianjin, China).
Theanine, catechin (C), gallocatechin (GC), epicatechin (EC), epigallocatechin (EGC), catechin gallate (CG), epicatechin gallate (ECG), epigallocatechin gallate (EGCG) and gallocatechin gallate (GCG) standards were purchased from Shanghai Yuanye Bio-Technology Co., Ltd. (Shanghai, China).
Acetonitrile and methanol were obtained from BCR International Trading, Inc. (Florida, USA). Formic acid was purchased from Shanghai EN Chemical Technology Co., Ltd. (Shanghai, China)
Quantification of purine alkaloids
-
Freeze-dried powder (0.1 g) was used to extract four kinds of purine alkaloids in 30 mL ultrapure water containing 1.5% magnesium oxide (w/v) by ultrasonic vibration at 100 °C for 30 min and then filtered through a 0.22 mm Millipore membrane. High-performance liquid chromatography (HPLC) was used to determine the purine alkaloids content according to a previous method with minimal changes[32]. Briefly, 10 μL filtered extract was injected into an XSelect HSS C18 SB column (4.6 × 250 mm, 5 mm, Waters Technologies, Milford, MA, USA) at a flow rate of 0.9 mL/min, with the column maintained at a constant temperature of 35 ± 1 °C. The detection of caffeine, theobromine, theophylline and theacrine was performed on a Waters Alliance E2695 equipped with a 2489 UV/Vis detector (Waters Technologies) at a wavelength of 280 nm. The compounds were isocratically eluted with 30% methanol/70% ultrapure water (v/v).
Determination of theanine and catechins
-
Theanine was quantified according to a previous method[2]. C, GC, EC, EGC, CG, GCG, ECG and EGCG content were detected with the instruments mentioned above at 280 nm. In brief, 0.2 g fine freeze-dried tea powders were subjected to extraction with 8 mL 70% methanol (v/v). One mL of the resulting supernatant was then filtered through a 0.22 mm Millipore membrane before being injected into an XSelect HSS C18 SB column (4.6 × 250 mm, 5 mm, Waters Technologies). Thereafter, catechin monomers were separated via gradient elution chromatography using a mobile phase consisting of 0.1% aqueous formic acid (v/v; A) and 100% acetonitrile (B). For the first 5 min, the mobile phase was 8% B; then from 5−14 min, B was increased from 8%−25%; finally, B was decreased from 25%−8% from 14−30 min.
RNA extraction and sequencing
-
Total RNA was isolated and purified from each sample using an R4151B-HiPure Plant RNA Kit B (Magen Biotechnology, Guangdong, China). The RNA concentration was quantified with a NanoDrop2000 spectrophotometer (Thermo Fisher Scientific, MA, USA). cDNA libraries were constructed according to the method in a previous study[15]. Samples were designated into two groups, TB (TB-1, TB-2, TB-3) and CAF (CAF-1, CAF-2, CAF-3). Raw reads in FASTQ format were first processed through in-house Perl scripts[33]. Paired-end clean reads were aligned to the reference genome obtained from the TPIA database (
http://tpdb.shengxin.ren/ ) using Hisat2[34]. Finally, the reads mapped to each gene were counted by featureCounts[35].Functional annotation and analysis of differentially expressed genes
-
For the functional annotation of the proteins, the gene sequences of samples were compared against the Kyoto Encyclopedia of Genes and Genomes (KEGG), and the Gene Ontology (GO) database. GO and KEGG enrichment analyses of differentially expressed gene sets were implemented using the KOBAS package and topG (
www.bioconductor.org/packages/release/bioc/html/topGO.html ), respectively[36]. Differential expression analysis was performed using EdgeR[37]. False discovery rate control was implemented by adjusting the obtained P-values via the Benjamini and Hochberg's approach. Genes fulfilling the criteria of |log2 (Fold Change)| > 1 and p value < 0.05 were considered differentially expressed. In addition, differentially expressed candidate transcription factor (TF) genes were predicted with iTAK for plants[38].qRT-PCR
-
Premier 3 was used to design the primers for qRT-PCR, and the sequence is given in Supplemental Table S1. qRT-PCR was conducted by a BioRad CFX384TM Real-Time System (Bio-Rad, Hercules, CA, USA) according to our previously published research[2]. CsActin1 was used as the internal reference, and the relative expression levels were calculated based on the 2−ΔΔCᴛ method[39].
Statistical analysis
-
All data are presented as the mean ± SD (n = 3). The double coordinate figure was produced with GraphPad Prism 9 software and correlation analysis was made in Origin 2022b R1. Heat maps were constructed with TBtools[40]. IBM SPSS Statistics 25 was used to run a Duncan's multiple-range test at p value < 0.05 to reveal significant differences. Corrplot package in R (version 4.1.0) was used to determine the correlation coefficient between two genes, and the Co-expression network was visualized by Cytoscape (version 3.9.1).
-
Concentrations of the main purine alkaloids in a C. ptilophylla population comprising 32 single plants were quantified (Fig. 1a). In 26 of the single plants, theobromine was the sole alkaloid, and its concentration ranged from 27.33−61.36 mg/g. The other six C. ptilophylla plants exhibited high levels of caffeine (37.43−54.86 mg/g) and low levels of theobromine (0.58−7.46 mg/g). As for theacrine and theophylline, they were not detected in the population (Fig. 1a).
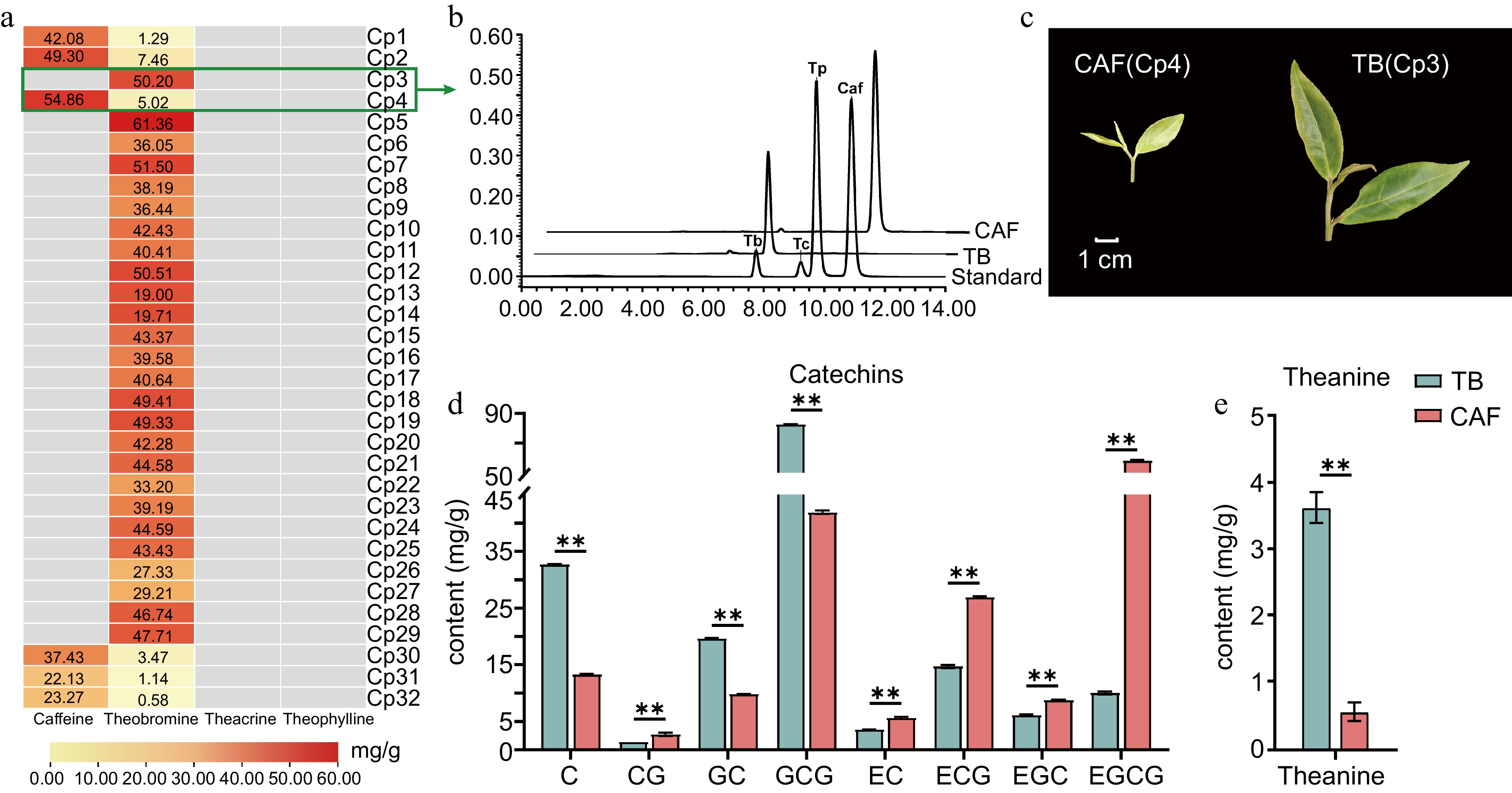
Figure 1.
The main secondary metabolites of Camellia ptilophylla. (a) Content of purine alkaloids in the tender leaves of 32 C. ptilophylla, where gray rectangles indicate the compound was not detected; (b) tender leaves of CAF and TB; (c) HPLC chromatogram of purine alkaloids (caffeine, theobromine, theacrine and theophylline) in TB and CAF; concentrations of (d) catechins and (e) theanine in TB and CAF. Catechin (C), gallocatechin (GC), epicatechin (EC), epigallocatechin (EGC), catechin gallate (CG), epicatechin gallate (ECG), gallocatechin gallate (GCG), and epigallocatechin gallate (EGCG). ** Significant difference between samples at the 0.01 level (P < 0.01) as determined by Student's t-test.
To understand the differential pathways for accumulation of purine alkaloids, two C. ptilophylla plants with contrasting purine alkaloid contents (i.e., one of the theobromine-only plants (Cp3, TB) and one of the theobromine and caffeine plants (Cp4, CAF) were subjected to further analysis. The two had significantly different morphologies, especially in the size of the bud and first two leaves, which were significantly larger in TB than in CAF (Fig. 1c). The levels of the major flavor substances, theanine and catechins, varied considerably between TB and CAF. For example, theanine content was significantly higher in TB (3.60 mg/g) in comparison to CAF (0.55 mg/g; Fig. 1e). Notably, the catechins contents were greatly different between TB and CAF (Fig. 1d). GCG was the dominant catechin monomer in TB, with a content of 82.76 mg/g, while in CAF it was only 41.9 mg/g. In contrast, EGCG was the dominant catechin monomer in CAF, at 58.36 mg/g, while it was 10.05 mg/g in TB (Fig. 1d).
RNA-Seq and reference genome alignment
-
To investigate the mechanism of differential accumulation of purine alkaloids, TB and CAF were subjected to RNA-seq analysis. A substantial amount of high-quality RNA sequencing data was generated, amounting to 44.85 Gb of clean reads and an average of 7.47 Gb clean reads per sample. The base-calling accuracy of the sequencing data were assessed by Q20 (base-calling error probability of 99%) and Q30 (base-calling error probability of 99.90%) values, which were both above 90.00% (Supplemental Table S2). Moreover, the Pearson correlation coefficients between the three replicates were all above 0.99, indicating a high level of consistency and reproducibility among the samples (Supplemental Fig. S1). The proportion of mapped reads, ranging from 85.54% to 86.33%, further indicated the high quality of the RNA-Seq data. These results collectively suggested that the RNA-Seq data in this study met the requirements for subsequent analyses.
Analysis of differentially expressed genes (DEGs) and functional enrichment
-
Analysis of the transcriptome data revealed 6,125 DEGs between TB and CAF, where DEGs differed by at least 2-fold, according to the transcript per kilobase million (TPM) mapped reads model. Among them, 2,514 were upregulated and 3,611 were downregulated (Fig. 2a). KEGG enrichment predicted that 127 metabolic pathways were associated with the annotated DEGs, including carbon metabolism, phenylpropanoid biosynthesis, photosynthesis, and glutathione metabolism (Supplemental Table S3). Particularly noteworthy was the enrichment of a considerable number of DEGs in the phenylpropanoid and flavonoid biosynthesis pathways, which are responsible for the biosynthesis of catechins. Purine metabolism, which was our primary focus, was also enriched as expected (Fig. 2c). In light of these findings, we focused on purine metabolism, phenylpropanoid biosynthesis, and flavonoid biosynthesis to delve into the underlying mechanisms responsible for the observed differences in the caffeine, theobromine, and catechins content at the transcript level. In addition, biological process enrichment of GO was shown in Supplemental Fig. S2. To validate the RNA-Seq data, 14 genes were chosen for RT-qPCR analysis, and the RT-qPCR analysis results were largely consistent with the RNA-seq results (R2 = 0.8012; Fig. 2b), indicating the reliability of the RNA-seq data.
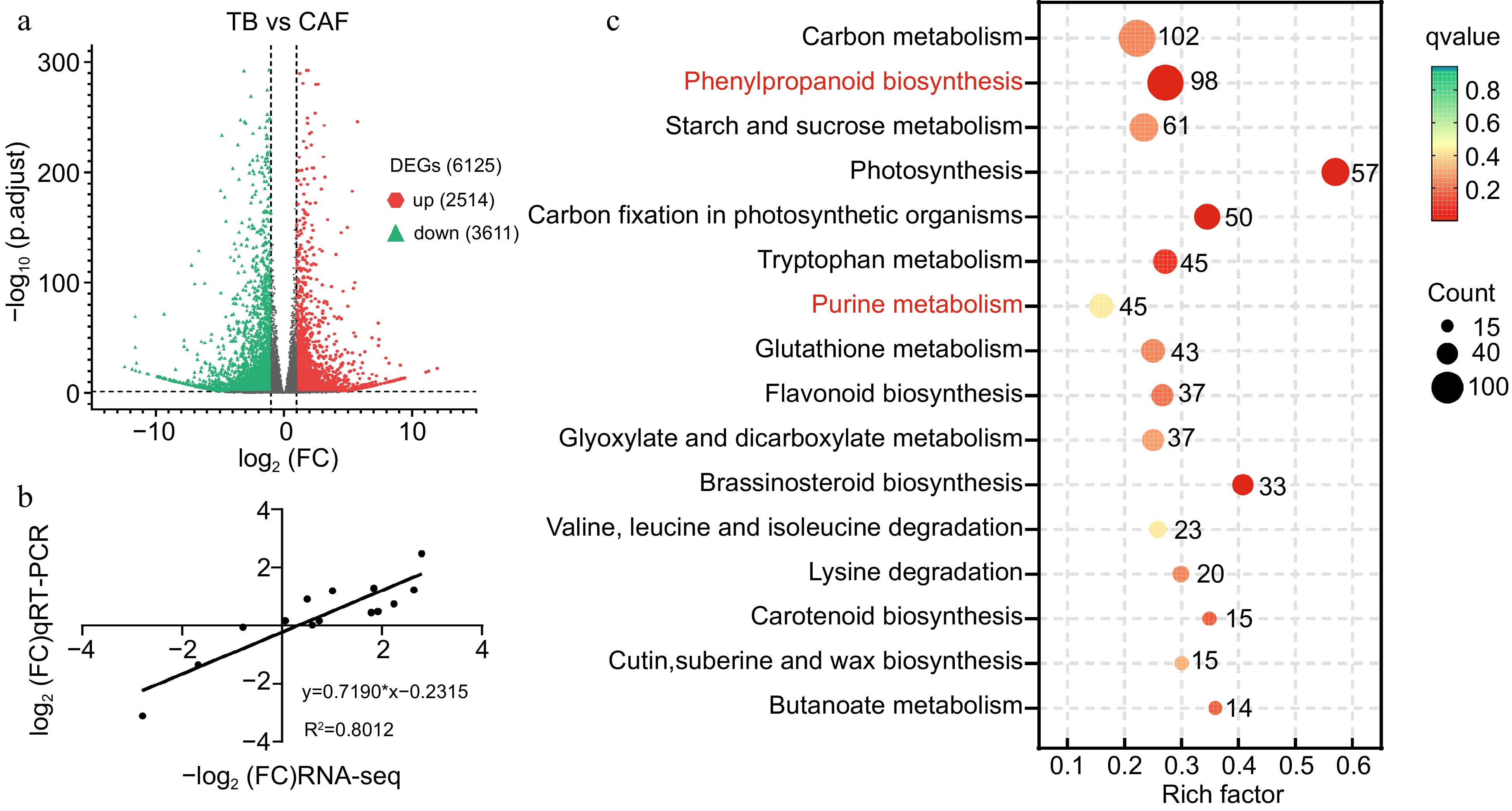
Figure 2.
(a) Volcano map showing differentially expressed genes (DEGs) between TB and CAF plants; (b) scatterplot of RNA-seq transcript abundance fold change values RNA-Seq (on the x-axis) plotted against RT-PCR expression values (on the y-axis). Actin (TEA019484) was used as the internal reference gene; and (c) KEGG pathways for the DEGs between TB and CAF. Red indicates the two pathways that were subsequently analyzed.
DEGs in purine alkaloid metabolism pathway
-
Purine alkaloid metabolism includes both synthesis and degradation pathways (Fig. 3a & b). Xanthosine, the main precursor of caffeine and theobromine, can be synthesized in five different pathways, including the 1) S-adenosylmethionine (SAM) cycle, 2) adenosine monophosphate (AMP) pathway, 3) guanine monophosphate (GMP) pathway, 4) de novo route, and 5) nucleic acids degradation (NAD) route (Fig. 3a). Heatmaps of the genes involved in purine alkaloid metabolism were generated, including xanthosine synthesis genes (Fig. 3c), methyltransferase genes (Fig. 3d), and degradation genes (Fig. 3e). The expression data is available in Supplemental Table S4. SAM synthetases (SAMS-1, SAMS-3 and SAMS-5), S-adenosyl-L-homocysteine hydrolases (SAHH-1 and SAHH-2), adenosine kinases (ADK-1 and ADK-5) and IMP dehydrogenase-2 (IMPDH-2) in xanthosine biosynthesis pathway were all significantly upregulated in TB.
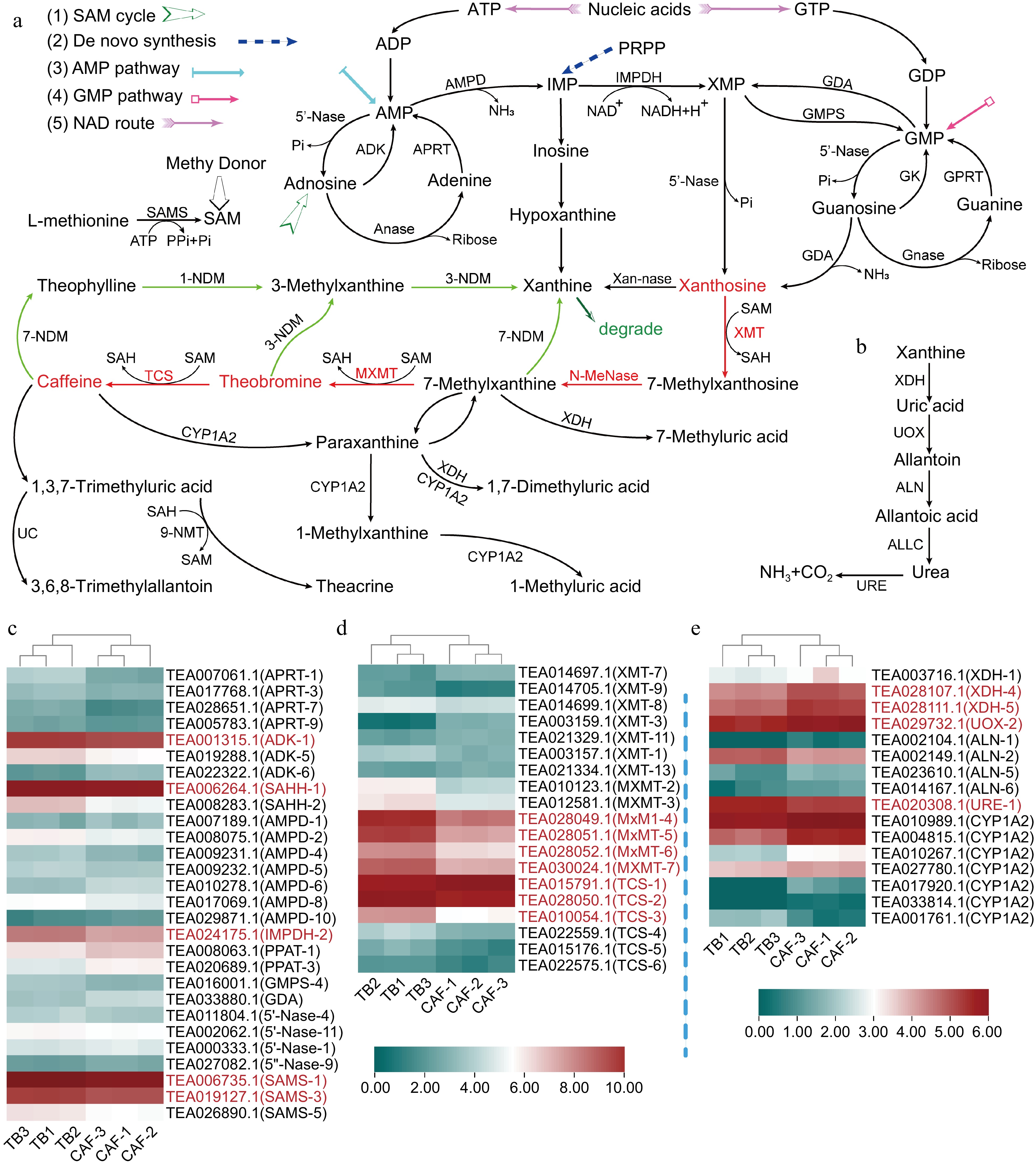
Figure 3.
Differential expression of genes involved in purine alkaloid biosynthesis and degradation; (a) xanthosine synthesis pathways: 1) SAM cycle, 2) de novo synthesis, 3) AMP pathway, 4) GMP pathway, and 5) the nucleic acid degradation (NAD) pathway, where red lines represent the main caffeine synthesis pathway, starting from xanthosine, and green lines represent the main pathway for caffeine degradation; (b) the xanthine degradation pathway; and heatmaps of the (c) genes in xanthosine biosynthesis; (d) methyltransferase genes involved in transforming xanthosine to caffeine; and (e) genes involved in xanthine degradation. Abbreviations: AMP deaminase (AMPD), 5′-nucleotidase (5′-Nase), IMP dehydrogenase (IMPDH), S-adenosyl-L-methionine synthase (SAMS), S-adenosyl-L-homocysteine hydrolase (SAHH), adenosine kinase (ADK), adenosine nucleosidase (Anase), adenine phosphoribosyl transferase (APRT), GMP synthase (GMPS); guanosine deaminase (GDA), xanthosine methyltransferase/7-methylxanthosine synthase (XMT), N-methyl nucleosidase (N-MeNase), methylxanthine methyltransferase/theobromine synthase (MXMT), 1,3-dimethylxanthine methyltransferase/tea caffeine synthase (TCS), 5-phosphoribosyl-1-pyrophosphate (PRPP), N-demethylase (NDM), cytochrome P450 family 1 subfamily A polypeptide 2 (CYP1A2), xanthine dehydrogenase (XDH), urate oxidase (UOX), allantoinase (ALN), allantoicase (ALLC), urease (URE), and 5-phosphoribosyl-1-pyrophosphate (PRPP) amidotransferase (PPAT). Genes written in red had significant differences in gene expression.
Xanthosine to theobromine and caffeine involves one nucleoside transfer, plus two and three methylations, respectively. The methyltransferase genes involved, MXMT-4, MXMT-5, and MXMT-7, were significantly upregulated in TB, where and MXMT-5 exhibited over four-fold higher expression in TB than in CAF (Fig. 3d). TCS-1, which catalyzes the last methylation to produce caffeine, was downregulated in TB as anticipated. However, TCS-2 and TCS-3 were upregulated in TB, which indicated that these genes may have roles in theobromine biosynthesis.
Another aspect affecting alkaloid content is the degradation pathway (Fig. 3b). In general, degradation of purine alkaloids was less active in TB than in CAF. Two xanthine dehydrogenase genes (XDH-4, XDH-5), and urate oxidase-2 (UOX-2) were downregulated in TB, while allantoinase-2 (ALN-2) and urease-1 (URE-1) increased. In addition, six out of seven cytochrome P450 family 1 subfamily A polypeptide 2 genes (CYP1A2), which sequentially demethylate caffeine and the resulting metabolites, were downregulated in TB, possibly because there was no caffeine to degrade in TB.
Expression level of catechin biosynthesis genes
-
The biosynthesis pathway of catechins, encompassing phenylpropanoid and flavonoid biosynthesis, is shown in Fig. 4a. In the phenylpropanoid biosynthesis pathway, phenylalanine ammonia-lyase (PAL) and cinnamate 4-hydroxylase (C4H) were upregulated in TB, while chalcone synthase (CHS) and cinnamate 3-hydroxylase (C3H) exhibited no noticeable differences in expression (Fig. 4b).
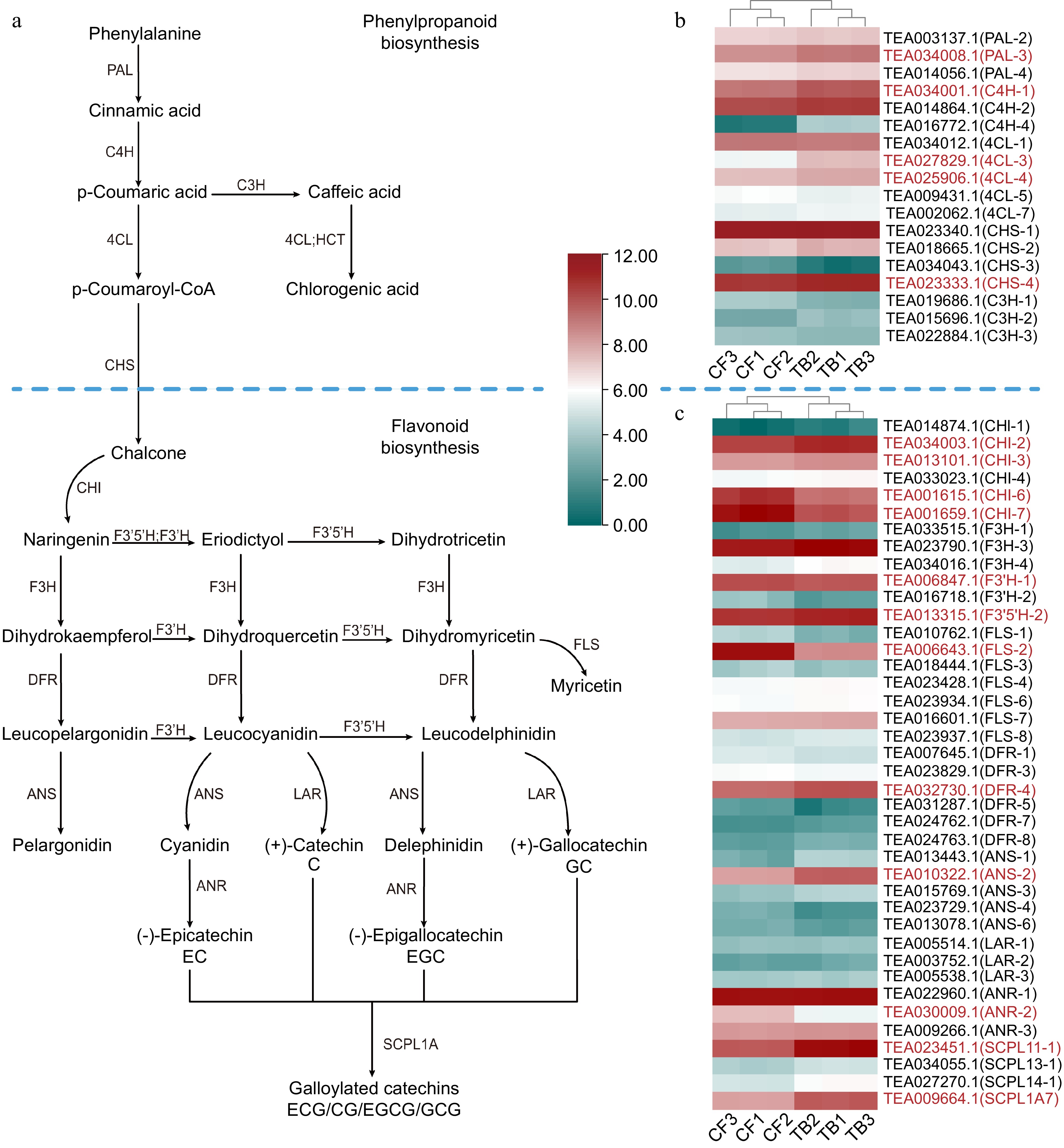
Figure 4.
(a) Phenylpropanoid, flavonoid, and catechin biosynthesis pathways and heatmaps of the expression level of (b) phenylpropanoid biosynthesis genes and (c) flavonoid biosynthesis genes. Phenylalanine ammonia-lyase (PAL), 4-coumarate-CoA ligase (4CL), cinnamate 4-hydroxylase (C4H), chalcone synthase (CHS), cinnamate 3-hydroxylase (C3H), chalcone isomerase (CHI), flavanone 3′-hydroxylase (F3′H), flavanone 3-hydroxylase (F3H), flavonoid 3′5′-hydroxylase (F3′5′H), flavonol synthase (FLS), dihydroflavonol reductase (DFR), leucoanthocyanidin reductase (LAR), anthocyanidin synthase (ANS), and serine carboxypeptidase-like acyltransferases (SCPL). Red represents a significant difference in gene expression.
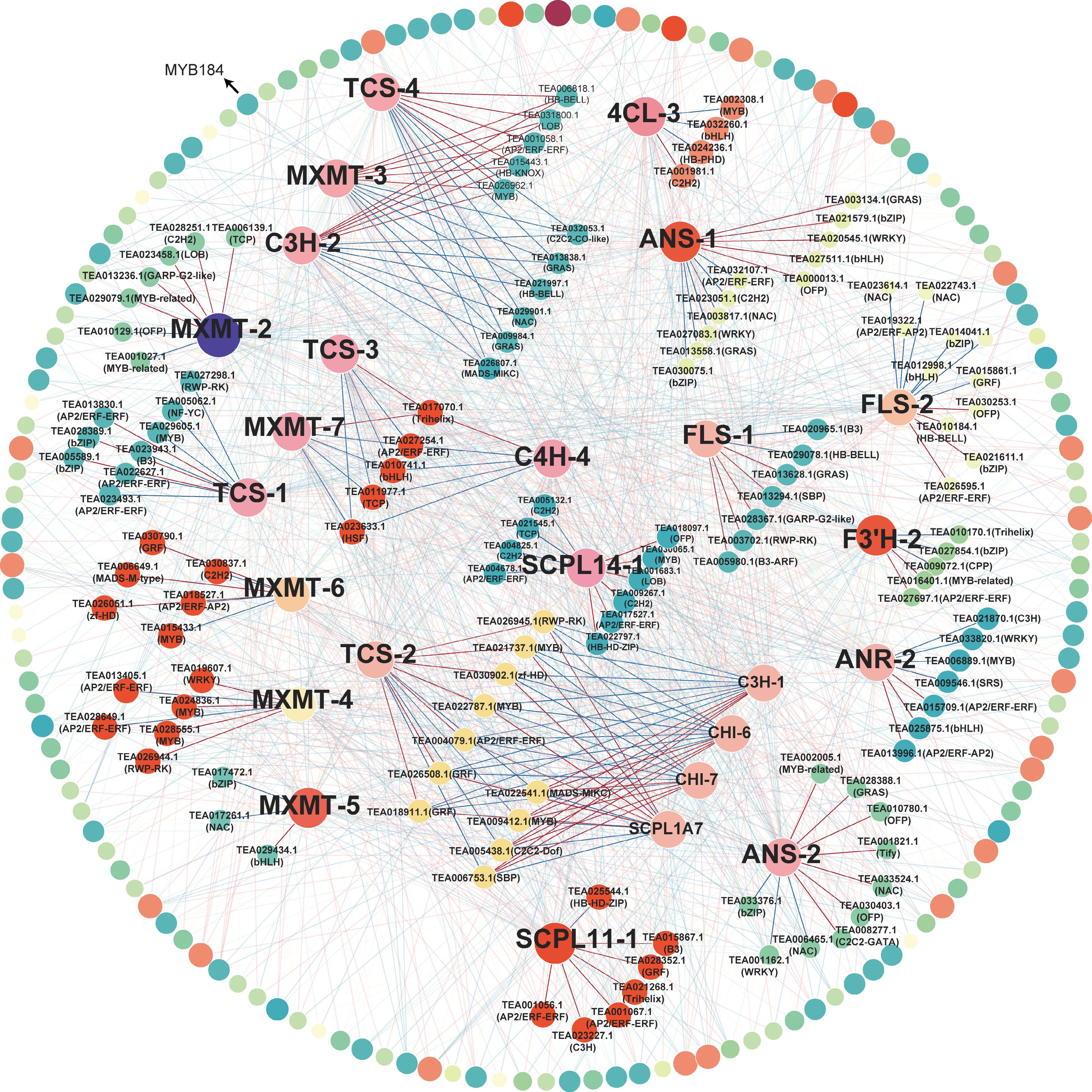
Figure 5.
Coexpression network between TFs and purine alkaloid and catechin biosynthesis genes. Pairs with P < 0.05 and | Spearman's rank correlation coefficient | > 0.8 were included in the analysis. Red lines and blue lines represent positive and negative correlations, respectively. The names of TFs with a high degree of co-expression are given. The size and color of nodes depend on the degree of co-expression. The outer ring with grey lines represents TFs with low correlation.
Regarding the flavonoid pathway, two chalcone isomerase genes (CHI-6, CHI-7), flavonoid 3′-hydroxylase-1 (F3′H-1) and anthocyanidin reductase-2 (ANR-2) were significantly downregulated in TB. Conversely, dihydroflavonol reductase-4 (DFR-4), anthocyanidin synthase-2 (ANS-2), flavonoid 3-hydroxylase (F3H) and flavonoid 3′5′-hydroxylase (F3′5′H-2) were upregulated in TB, which was consistent with the contents of C, GC and GCG, while leucoanthocyanidin reductases (LARs) didn't have significant differences between samples. Glucosyltransferases and serine carboxypeptidase-like acyltransferases 1 subfamily A (SCPL1A), a kind of epigallocatechin acyltransferase, participates in the synthesis of ECG and EGCG[41,42]. Among the four upregulated SCPL1A genes (SCPL11-1, SCPL13-1, SCPL14-1, SCPL11-1) in TB, SCPL11-1 showed the highest expression, with a 3-fold increase in TB, which might be the primary reason for the accumulation of GCG in TB. The expression data is available in Supplemental Table S5.
Co-expression network
-
A total of 327 transcription factors (TFs) were identified by RNA-seq, with the AP2/ERF-ERF family being the most prominent (48 TFs, 14.68%), followed by the MYB (33 TFs, 10.09%) and NAC families (22 TFs, 6.73%) (Supplemental Table S6). To explore the potential regulatory mechanisms underlying differential gene expression in the purine alkaloid and catechin biosynthesis pathways, co-expression networks were constructed (Fig. 5). The key gene of theobromine synthesis MXMT-4[15] was significantly correlated with two AP2/ERF-ERFs (TEA013405.1, TEA028649.1) and two MYBs (TEA028555.1 and TEA024836.1). Interestingly, our studies showed that there was no positive correlation between the expression level of CsMYB184 and CsTCS1. In addition, B3 (TEA015867.1), GRF (TEA028352.1) and AP2/ERF-ERF (TEA001056.1) were significantly related to CsSCPL11-1 expression, indicating that those TFs might regulate GCG biosynthesis.
Data availability statement
-
The raw sequencing data in the study are publicly available in the NCBI BioProject: PRJNA972319.
-
C. ptilophylla was thought to contain theobromine only instead of caffeine, but the current evidence does not support this view[16]. The random amplified polymorphic DNA (RAPD) technique was used to analyze C. ptilophylla that differed in purine base composition, and it revealed that C. ptilophylla containing only theobromine was more closely related to other C. ptilophylla plants containing both theobromine and caffeine but was more distantly related to cultivated species of tea plants[43]. In our studies, C. ptilophylla showed intra-species diversity, i.e., a small number of single plants contained caffeine, which was consist with the results of a recent published study[30]. In addition, we found that GCG was much more abundant than EGCG in TB, while EGCG was the major catechin in CAF, which means that not only purine alkaloids differ among C. ptilophylla populations, but that catechin compositions also have intraspecies diversity in this species. Whether there is any connection between the high GCG content and defective caffeine biosynthesis needs to be further confirmed. Our research has provided new insights into the metabolites of C. ptilophylla population.
RNA sequencing was used to discover the differential accumulation of purine alkaloids and catechins between TB and CAF. In agreement with previous studies[16,44−47], we found that TCS1 expression was downregulated in TB compared to CAF (Fig. 3d), which might be the cause of defective caffeine biosynthesis in TB. Interestingly, TCS-2, TCS-3 and xanthosine biosynthesis genes (SAMS-1, SAHH-1, and ADK-1) were upregulated in TB, which might be related to the high content of theobromine in TB. As expected, the expression level of CYP1A2s, which catalyze the demethylation of caffeine, were all downregulated in TB. In addition, XDH-4, XDH-5, and UOX-2, which participate in the degradation of purine alkaloids, were less active in TB (Fig. 3e). CsMYB184, a crucial transcription factor responsible for regulating caffeine biosynthesis in tea plants, has a proven link to TCS1, where lower transcription of this TF coincides with reduced TCS1 expression in C. ptilophylla[16]. Different from previous studies, our results showed that the expression level of CsMYB184 in TB was significantly higher than that in CAF, and there was no positive correlation to the expression level of CsMYB184 and TCS1. Based on the co-expression network, two AP2/ERF-ERFs (TEA013405.1, TEA028649.1) and two MYBs (TEA028555.1 and TEA024836.1) appeared to be highly related to MXMT-4, a key gene regulating the biosynthesis of theobromine, which implied that those TFs might regulate theobromine biosynthesis.
Regarding catechin biosynthesis, F3'H-1 and F3'5'H-2 were significantly upregulated in C. ptilophylla, which was consistent with the elevated GCG contents. Furthermore, CsSCPL11-1, thought to be related to the biosynthesis of EGCG[26,41,42], was significantly upregulated in TB, which was consistent with the observed GCG content, but was contrary to the amount of EGCG in TB. Meanwhile, GC is the precursor to synthesize GCG[28]. The significant increase of GC content in TB might be the reason for the high amount of GCG. These results suggest that the upregulation of F3'H, F3'5'H and SCPLs may played an important role in the synthesis of GCG in C. ptilophylla. Additionally, the co-expression network showed that transcription factors B3 (TEA015867.1), GRF (TEA028352.1) and AP2/ERF-ERF (TEA001056.1) were significantly related to CsSCPL11-1.
-
C. ptilophylla is typically thought of as a naturally decaffeinated tea. However, there is intraspecies variation. In the population studied here, ~19% of the plants contained high levels of caffeine (CAF), while the majority were caffeine-free (TB). Xanthosine synthesis pathways were more active in TB than that in CAF, especially in the SAM cycle and AMP pathway, and the high theobromine content in TB may have been caused by significantly high expression of both TCS and MXMT and a lower level of alkaloid degradation compared to CAF. Additionally, CsMYB184 (TEA029017.1) appeared to be involved in the biosynthesis of both caffeine and theobromine, which requires further verification. Furthermore, an increase in GCG content was observed in TB, which corresponded to the upregulated expression of F3'H, F3'5'H and SCPL, suggesting that these genes may play a crucial role in the accumulation of GC and GCG. This study lays out fundamental metabolite accumulation patterns in a C. ptilophylla population and provides possible regulation mechanisms of purine alkaloid and catechin biosynthesis. The understanding of purine alkaloid and catechins metabolism will facilitate the breeding of low-caffeine or high GCG tea plant cultivars for meeting diverse demands of consumers.
This work was supported by the Natural Science Foundation of Guangdong Province (2023A1515010786, 2022A1515111141).
-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Pearson correlation coefficient between these three replicates.
- Supplemental Fig. S2 Pathway in BP enrichment of GO.
- Supplemental Table S1 List of qPCR primers used in this study.
- Supplemental Table S2 Data output and quality statistics.
- Supplemental Table S3 KEGG pathway and gene involved.
- Supplemental Table S4 genes involved in purine alkaloid metabolims.
- Supplemental Table S5 genes involved in polyphenol metabolism.
- Supplemental Table S6 genes annotated as TF by iTAK.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ying C, Chen J, Chen J, Zheng P, Zhou C, et al. 2023. Differential accumulation mechanisms of purine alkaloids and catechins in Camellia ptilophylla, a natural theobromine-rich tea. Beverage Plant Research 3:15 doi: 10.48130/BPR-2023-0015 

Differential accumulation mechanisms of purine alkaloids and catechins in Camellia ptilophylla, a natural theobromine-rich tea
- Received: 25 April 2023
- Revised: 28 May 2023
- Accepted: 30 May 2023
- Published online: 27 June 2023
Abstract: Tea is consumed worldwide due to its charming flavor and the refreshing effects conferred by caffeine. Caffeine however has undesirable side effects, such as sleep disturbance. Camellia ptilophylla is known for its low caffeine content, and the biosynthesis of purine alkaloids in this species has become a hot topic. In this study, the accumulation of purine alkaloids in a natural C. ptilophylla population (32 plants) was analyzed, and the results showed that 81.25% of this population were caffeine-free, containing only theobromine (TB), while six plants contained both theobromine and caffeine (CAF). RNA-seq analysis of two C. ptilophylla plants with contrasting purine alkaloid contents (TB and CAF) revealed that xanthosine synthesis genes of the SAM cycle and AMP pathway were significantly related to the differential accumulation of purine alkaloids between TB and CAF. The high theobromine content in TB was attributed to the significantly higher expression of TCS-2, TCS-3 and MXMTs and downregulation of the xanthosine degradation pathway in comparison to CAF. Additionally, CsMYB184 was significantly upregulated in TB, opposing the expression pattern of TCS1, but consistent with that of other TCSs and MXMTs. Furthermore, the upregulated expression of catechin biosynthesis genes, F3'H, F3'5'H and SCPLs in TB corresponded to a higher gallocatechin gallate (GCG) content. Overall, these findings provide new insights into the accumulation of theobromine and GCG, which may facilitate the development of tea plant cultivars with low-caffeine or high GCG to meet the diverse demands of consumers.
-
Key words:
- Camellia ptilophylla /
- Theobromine /
- Gallocatechin gallate