










-
Teosinte Branched 1/Cycloidea/PCF (TCP) proteins are a conserved and widespread family of plant-specific transcription factors[1]. Different plant species have evolved multiple ways to precisely regulate the downstream target genes of the TCPs, and expression of the TCPs themselves is also tightly regulated. TCPs have important regulatory roles in the development of roots, stems, leaves, flowers, and fruits; they also participate in plant secondary metabolism and responses to biotic and abiotic stress[2−4].
The TCP gene family in plants is named after its first identified members — i.e., Teosinte Branched 1 (TB1) in maize, Cycloidea (CYC) in snapdragon, and Proliferating Cell Factor 1 (PCF1) and PCF2 in rice[2,5,6]. TCP TFs contain a typical basic helix-loop-helix (bHLH) structure and can be divided into three subgroups (PCF, CIN, and CYC/TB1) according to the characteristics of this conserved TCP domain[6]. Many studies have shown that PCF family members can regulate cell proliferation and growth, leaf and flower development, and other processes[7−9]. However, compared with the CIN subgroup, there have been far fewer studies on the PCFs. TCP family members mainly regulate aspects of plant growth and development in the meristem, including vascular tissue development[10−12], seed germination, and embryonic growth[13]. TCPs can also influence leaf shape and size through direct regulation of leaf development-related genes, miRNAs, and plant hormones[5,14,15]. A number of TCPs can also enhance plant stress response by regulating cell osmotic pressure, changing cell permeability, reducing harmful substances, and regulating hormone responses. For example, heterologous expression of PeTCP10 in Arabidopsis thaliana increased antioxidant enzyme activities and relative water content while reducing electrolyte leakage and Na+ accumulation, thus improving overall stress resistance[16]. Similar results were also demonstrated in rice[17] and white birch[18], and overexpression of maize ZmTCP42 in Arabidopsis significantly increased the sensitivity of seed germination to ABA and promoted drought resistance[19]. According to the RNA-seq analysis, AsTCP17 responded considerably to drought. AsTCP17-OE Arabidopsis increased antioxidant enzyme activity and lowered reactive oxygen species under drought treatment[20]. Drought stress boosted the survival rate and ABA content of GbTCP5-OE Arabidopsis while decreasing the water loss rate and malondialdehyde (MDA) content of the leaves. GbTCP5 in cotton was shown to be a positive regulator of drought stress[21]. Different from the above results, ZmTCP14 in maize was found to play a negative feedback role in regulating the response of plants to drought stress. The results showed that ZmTCP14 gene was regulating the accumulation of ROS in plants to reduce drought resistance under drought stress[20,22]. MdTCP46 overexpression in Arabidopsis reduced ABA sensitivity and drought stress resistance. MdTCP46 significantly decreased MdABI5 transcription activity, therefore negatively modulating the ABA signal and drought response mediated by MdABI5[23]. After low temperature treatment in Chrysanthemum nankingense, it was found that CnTCP2/4/14 responded quickly to cold stress. Over-expression assays in Arabidopsis showed the negative regulatory factor of cold acclimation by down-regulating the content of cold-induced genes such as AtCBF1/2/3, AtCOR15A and AtKIN1, and increasing MDA[24]. Cold treatment increased the expression of BcTCP3/12/21/22 in Pak-choi by more than 30 times[25], and it shows that BcTCP3/12/21/22 plays an important role in low temperature response. In general, TCP family transcription factors play an essential and multifaceted function in plant drought and low temperature stress. Abscisic acid (ABA) is an endogenous plant hormone that regulates many aspects of plant growth and development, including seed germination, seedling growth, reproduction, and aging, as well as plant responses to various abiotic stresses. Many studies have shown that TCP transcription factors play a key role in ABA signal transduction and regulation of the drought response[23,26,27]. AtTCP14 interacts with DNA Binding with One Finger 6 (DOF6) transcription factors, inhibits expression of the ABA biosynthesis gene ABA Deficient 1 (ABA1) and other ABA response-related genes, and plays a regulatory role in seed germination via the ABA pathway[28]. Overexpression of AtTCP4 increased the sensitivity of Arabidopsis to ABA and inhibited the promotion of leaf growth by endogenous ABA, and exogenous ABA could eliminate this inhibition[29].
Methyl jasmonate (MeJA) is a phytohormone that is synthesized in response to environmental stimuli and has important roles in regulating plant growth and development, hormone signal transduction, and responses to biotic and abiotic stress[30,31]. Jasmonate-Zim Domain (JAZ) proteins are essential regulators of plant responses to the jasmonic acid (JA) signaling pathway, and studies have shown that JAZ and MYB transcription factors interact to regulate plant growth and development[32]. MYB transcription factors have roles in biotic and abiotic stress responses, and cis-acting elements such as MBSI, MBSII, MBSIG, MYBCORE, AC-box, and E-box in the promoters of downstream target genes interact with MYB transcription factors to participate in plant stress signal transduction[33]. Given the role of TCPs in regulating the ABA signaling pathway, there is a strong possibility that these transcription factors may also participate in MeJA signaling. However, this potential interaction remains to be explored.
Tea is one of the top three non-alcoholic beverages worldwide, making the tea plant an important global cash crop. Tea plant growth, development, and stress response have important effects on yield and quality during horticultural production. In recent years, both cold waves and hot, dry conditions have frequently affected tea plant growth and thus tea output, causing major losses to farmers and local economies. A better understanding of the molecular basis of stress responses in tea is therefore needed to inform management practices and breeding efforts.
Previous studies have shown that CsTCPs have roles in tea shoot germination[34], stem tip elongation[35], disease response[36], leaf development, and hormone responses[37]. Although many studies have examined the functions of TCP transcription factors in model crops, little is known about the roles and regulation of TCP transcription factors in tea plants. Here, we selected two candidates (CsTCP5 and CsTCP18) from among 37 previously identified TCPs in tea plants[38] and introduced them into Arabidopsis for functional characterization. CsTCP5 was selected because its orthologs AtTCP14/AtTCP15 have regulatory roles in leaf development, stem elongation[39], auxin signal transduction[7], seed germination[13], cell proliferation[40], and other processes. The ortholog of CsTCP18 , AtTCP20, is involved in cell proliferation and division[8,41], leaf development, and JA signal transduction[42]. CsTCP5 and CsTCP18 were cloned from tea plants, overexpressed in Arabidopsis, and functionally characterized to better understand their roles in tea plant regulatory pathways.
-
To generate Arabidopsis lines overexpressing CsTCP5 and CsTCP18, the coding sequences (CDSs) of CsTCP5 and CsTCP18 were individually cloned into the pK7FWG 2.0 vector and incorporated into Agrobacterium tumefaciens strain GV3101. These vectors were transformed into Col-0 Arabidopsis plants by Agrobacterium-mediated transformation using the floral dip method[43,44]. The pK7FWG 2.0 empty vector was also transformed into Col-0 Arabidopsis (WT) to serve as a control (EV). The primer pairs used are listed in Supplemental Table S1.
One-year-old tea seedlings of similar size and consistent growth were chosen for treatment and sampling as described in Supplemental Table S2. Spray treatments were applied until the plant leaves were completely wet. Each sample consisted of one bud and two leaves, and there were three biological replicates of each treatment.
Arabidopsis seedlings from the T3 generation were grown on a 1/2 Murashige and Skoog (MS) plate for 10 days and then transplanted into growth substrate (vermiculite : nutrient soil : perlite = 9:3:1). Processing and sampling began after the plants had grown normally for 14 days (Supplemental Table S3).
Arabidopsis, Nicotiana benthamiana, and approximately 1-year-old tea plants of C. sinensis 'Longjing43' were planted in a light incubator at the Tea Science Research Institute, College of Horticulture, Nanjing Agriculture University, Jiangsu Province, China. The incubator maintained a consistent temperature of 22 °C and a 14-h/10-h (day/night) photoperiod.
Subcellular localization assays
-
CsTCP5 and CsTCP18 were inserted into the plant expression vector pK7FWG 2.0 with the EGFP tag to construct 35S::CsTCP5-EGFP and 35S::CsTCP18-EGFP. The 35S::pK7FWG 2.0-EGFP, 35S::CsTCP5-EGFP, and 35S::CsTCP18-EGFP expression vectors were separately co-transformed with the nuclear marker P19 into tobacco (N. benthamiana) leaf cells via A. tumefaciens for transient expression. Localization of the fluorescent proteins was observed with an ultra-high-resolution confocal microscope (ZEISS LSM 800, Jena, Germany).
Transcriptional activation activity
-
CsTCP5 and CsTCP18 were inserted into the pGBKT7 vector via BamH I single-enzyme digestion. The ligation products and unloaded pGBKT7 were co-transformed into yeast Y2H competent cells, and colony growth was observed on synthetic defined (SD) media. Trp-deficient single colonies on SD/-Trp plates were picked and diluted to the same number of spots on plates containing SD/-His/-Ade medium coated with X-α-gal. Plates were placed in an inverted position and incubated for approximately 3 d.
Y2H and BiFC assays for protein interactions
-
For Y2H assays of protein interactions, CsMYB46, CsMYB105, CsJAZ3, CsJAZ10 and CsJAZ11 were ligated into the pGADT7 vector by BamH I single-enzyme digestion. Combinations of these ligation products, unloaded pGADT7, and pGBKT7-CsTCP5/CsTCP18 were co-transformed into yeast Y2H competent cells, and colony growth was observed.
For BiFC assays of protein interactions, CsTCP5, CsTCP18, CsMYB46, CsMYB105, CsJAZ3, CsJAZ10, and CsJAZ11 were inserted into pCAMBIA1300-C-YFPC and pCAMBIA1300-C-YFPN vectors using BamH I single-enzyme digestion. Combinations of the resulting plasmids were transformed into Agrobacterium tumefaciens strain GV3101, and subsequent experimental steps were performed as described above for subcellular localization assays. The primer sequences are provided in Supplemental Table S4.
Phenotypic observations of transgenic Arabidopsis
-
For observation of germination rates, 50 sterilized T3 Arabidopsis seeds were placed onto solid media with various concentrations of ABA and MeJA: ½ MS, ½ MS + 10 μM ABA, ½ MS + 20 μM ABA, ½ MS + 50 μM ABA, ½ MS + 5 μM MeJA, and ½ MS + 20 μM MeJA. Each treatment was replicated three times.
For observation of root lengths, seedlings with root lengths of about 1 cm were transferred to different media: ½ MS, ½ MS + 10 μM ABA, ½ MS + 20 μM ABA, ½ MS + 5 μM MeJA, and ½ MS + 20 μM MeJA. Each treatment was replicated three times using three seedlings per strain.
For observation of survival rate, T3 Arabidopsis seedlings were grown on ½ MS plates for 10 d, then transferred to growth substrate (vermiculite : nutrient soil : perlite = 9:3:1) and grown under normal conditions for 14 d. They received 3 L of water during this period. Water was then withheld for 14 d, and the number of surviving seedlings was counted.
To observe the number of flowering plants, T3 Arabidopsis seedlings were grown on ½ MS plates for 10 d, then transferred to growth substrate (vermiculite : nutrient soil : perlite = 9:3:1) and grown under normal conditions for 14 d (3 L of water in total). Water was then withheld for 7 d, and the number of flowering plants was counted.
Biochemical assays of transgenic Arabidopsis
-
Activities of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) and contents of malondialdehyde (MDA) and proline were measured using commercial kits (Nanjing Jiancheng Institute of Bioengineering, Nanjing, China).
Quantitative real time-PCR
-
Quantitative PCR was performed on a Roche Light Cycler 480 II system using SYBR Green dye (Takara, Dalian, China) with the following program: 95 °C for 30 s; 40 cycles of 95 °C for 10 s and 60 °C for 30 s. The β-actin gene from C. sinensis was used as an internal control[45], and there were three biological replicates and three technical replicates of each sample type. The primer pairs used are listed in Supplemental Table S5. Relative gene expression was quantified using the 2−ΔΔCᴛ method[46].
Statistical analysis
-
The experimental data were sorted and analyzed using Microsoft Excel 2019 software (Microsoft Corp., Albuquerque, USA). Significant differences between the experimental treatments were determined by one-way ANOVA and Duncan's multiple range test performed in IBM SPSS Statistics 26.0 (IBM Corporation, New York, USA). All data are presented as mean ± standard deviation (SD) of three independent replicates, and different letters indicate significant differences. Graphs were constructed using GraphPad 8.0.1 (GraphPad Software, San Diego, USA).
-
We first cloned CsTCP5 and CsTCP18 from C. sinensis using their CDS sequences from the TPIA tea genome database (
http://tpia.teaplant.org/index.html ) (Supplemental Fig. S1 & S2). In our previous study, in silico subcellular localization predictions suggested that CsTCP5 was localized outside the nucleus and CsTCP18 inside the nucleus[38]. Here, we performed subcellular localization assays to confirm these predicted localizations in living cells of N. benthamiana. The fluorescent signal of 35S::pK7FWG 2.0-EGFP was distributed throughout the nucleus and cytoplasm, whereas the fluorescent signals of 35S::CsTCP5-EGFP and 35S::CsTCP18-EGFP were observed only in the nucleus (Fig. 1), indicating that both CsTCP5 and CsTCP18 were localized in the nucleus, in contrast to earlier predictions.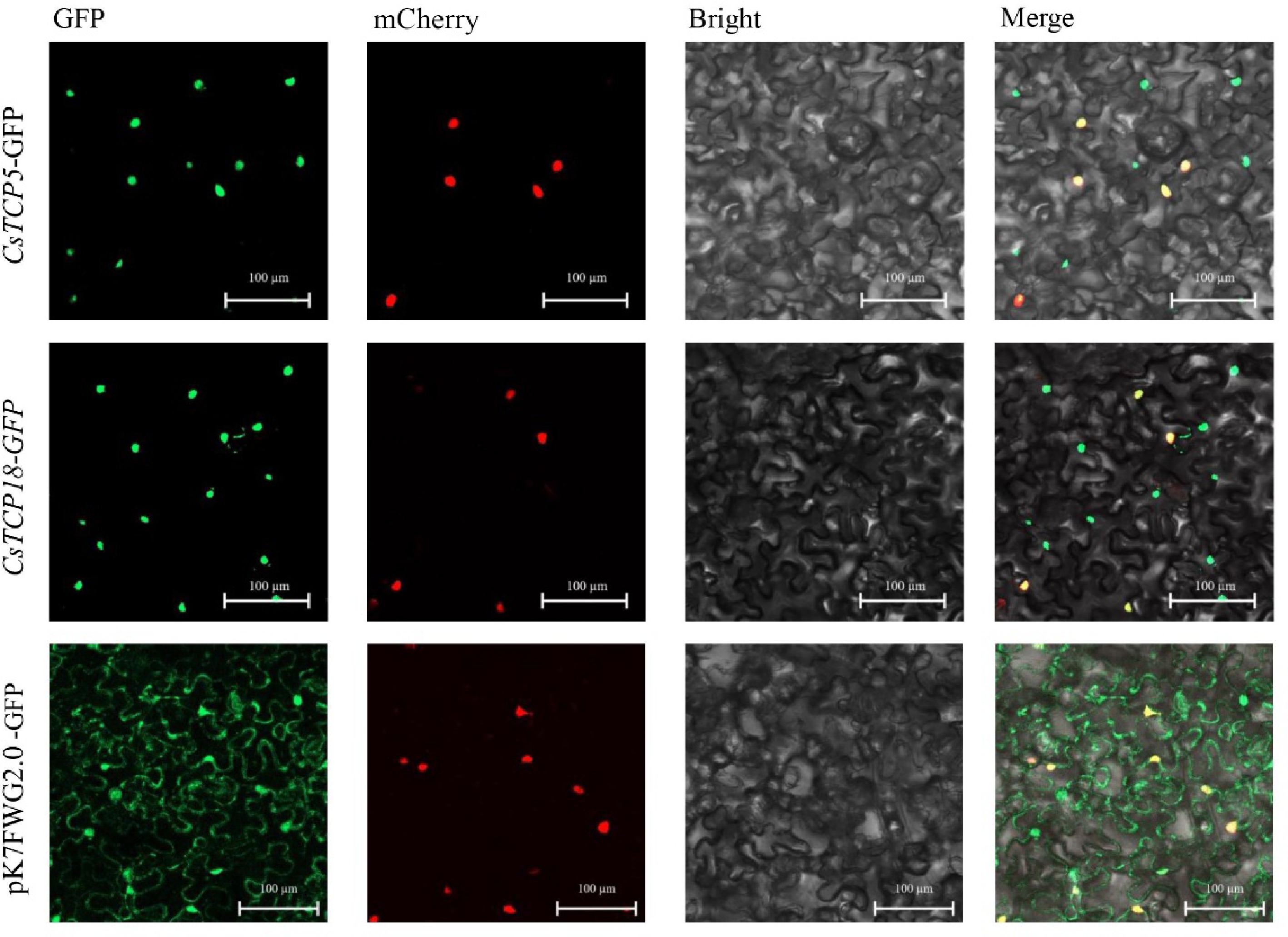
Figure 1.
Subcellular localization of CsTCP5 and CsTCP18 proteins in N. benthamiana leaf cells. GFP: green fluorescence channel. mCherry: red fluorescence channel. Bright: bright field channel. Merge: superimposed channel. Images are representative of at least three independent experiments. Bar = 100 µm.
Expression of CsTCP5 and CsTCP18 under hormone and stress treatments
-
To determine whether CsTCP5 and CsTCP18 are induced by different hormones and stresses, we performed RT-qPCR on one-year-old 'Longjing 43' cuttings that had been exposed to MeJA, ABA, low temperature, drought, and combinations of these treatments (Supplemental Table S2). CsTCP5 and CsTCP18 were influenced by different hormones and stresses, and their expression patterns were distinct (Fig. 2a, b). Transcript levels of CsTCP5 were significantly reduced by MeJA treatment, whereas those of CsTCP18 were not. By contrast, levels of CsTCP5 increased significantly and those of CsTCP18 decreased significantly after ABA treatment. Low temperature (4 °C) reduced the transcript levels of both genes, and this reduction was greater for CsTCP18. Drought had no significant effect on CsTCP5 expression but significantly reduced that of CsTC18. The combination of MeJA and low temperature (MLT) reduced the expression of CsTCP5 more than either treatment alone and reduced the expression of CsTCP18 more than MeJA alone. By contrast, the combination of ABA with drought (AD) had opposite effects on CsTCP5 and CsTC18. CsTCP5 expression was significantly reduced compared with ABA alone, whereas CsTCP18 expression was strongly and significantly increased compared with ABA or drought alone. Thus, ABA, MeJA, low temperature, and drought treatments significantly altered the expression of CsTCP5 and CsTCP18, although their specific effects differed between the two genes.
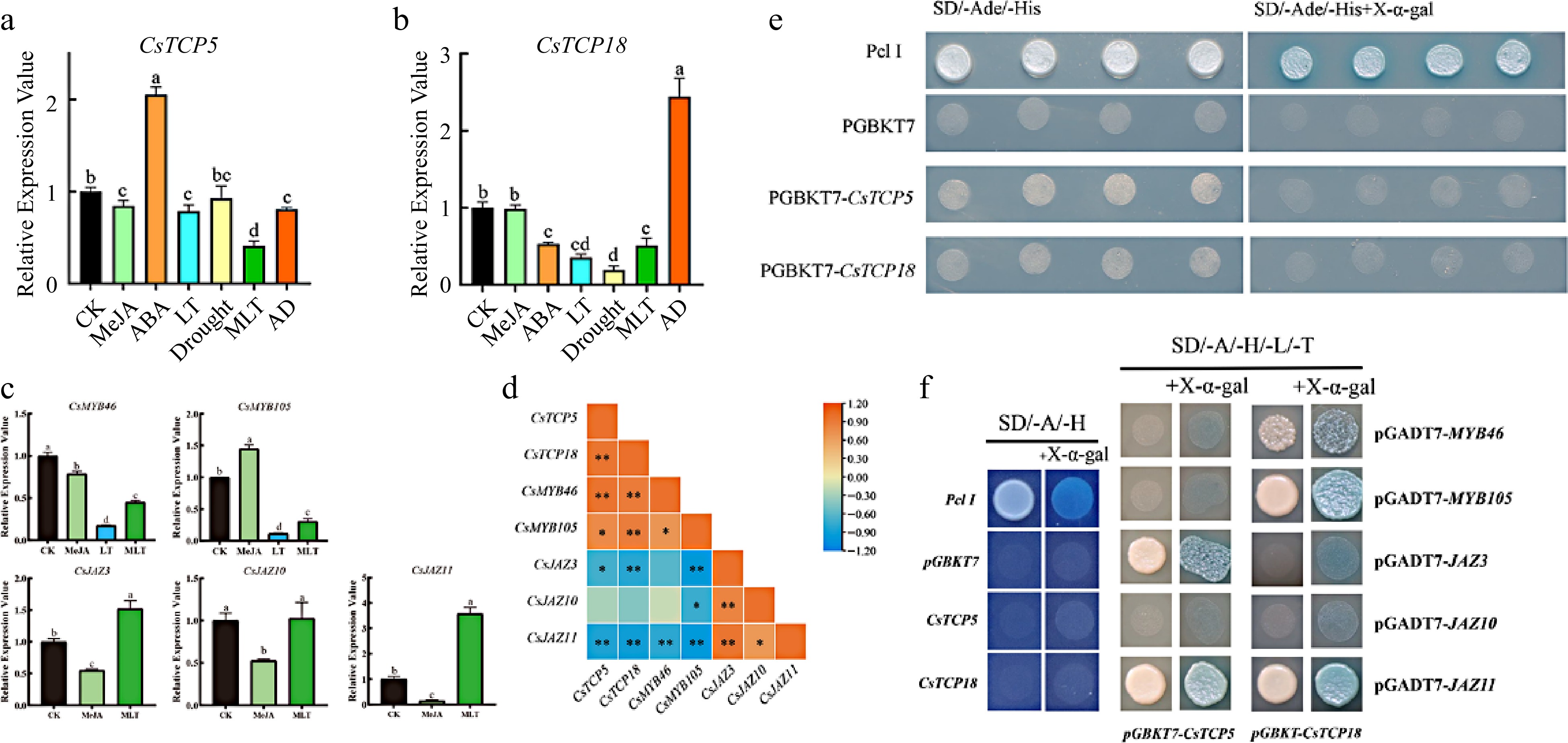
Figure 2.
Relative expression of CsTCP5, CsTCP18, CsMYBs, and CsJAZs in response to different treatments and Y2H assays. (a) Relative expression of CsTCP5. (b) Relative expression of CsTCP18. (c) Relative expression of CsMYBs and CsJAZs under MeJA, LT and/or MLT treatments. (d) Correlation analysis of CsTCP5/18, CsMYB46/105, and CsJAZ3/10/11 expression. CK: control group. MeJA: methyl jasmonate. ABA: abscisic acid. LT: low temperature. MLT: MeJA and low temperature. AD: ABA and drought. In (a)–(c), different letters indicate significant differences (p < 0.05). Error bars represent the SE of three replicates. In (d), asterisks indicate significant differences between values (* p < 0.05, ** p < 0.01). (e) Y2H transcriptional activation assay of CsTCP5 and CsTCP18. SD/-Ade/-His, SD medium lacking adenine and histidine. SD/-Ade/-His+X-α-gal, SD medium lacking adenine (Ade) and histidine (His) and containing 10 μM X-α-gal. Pcl I, positive control. PGBKT7, negative control. (f) Y2H assays reveal interactions of CsTCP5 and CsTCP18 with CsMYBs and CsJAZs in yeast cells. SD/-A/-H, SD medium lacking adenine and histidine. SD/-A/-H/-L/-T, SD medium lacking adenine, histidine, leucine, and tryptophan. +X-α-gal, 10 μM X-α-gal. Pcl I, positive control. PGBKT7, negative control.
We next examined the expression of CsMYB46/CsMYB105 and CsJAZ3/CsJAZ10/CsJAZ11 under a subset of hormone and stress treatments (Fig. 2c). All except CsMYB105 showed significantly reduced expression in response to MeJA, and the two MYBs showed significantly reduced expression in response to LT. Effects of the combined MLT treatment differed: expression of the two MYB genes was significantly lower under MLT conditions than under control conditions, whereas expression of the three JAZ genes was significantly greater or unchanged.
Correlation analysis revealed significant correlations between the expression of CsTCP18 and that of CsMYB46, CsMYB105, CsJAZ3, and CsJAZ11. The correlations were positive for the MYB genes and negative for the JAZ genes (Fig. 2d). Likewise, expression of CsTCP5 was significantly correlated with that of CsMYB46, CsMYB105, CsJAZ11, and CsJAZ3, with positive correlations for the MYBs and negative correlations for the JAZs. Expression of CsTCP5 and CsTCP18 was also positively correlated.
Assessment of CsTCP5 and CsTCP18 transcriptional activation activity
-
We performed Y2H assays to determine whether CsTCP5 and CsTCP18 had transcriptional activation activity (Fig. 2e). Yeast transformed with the positive control Pcl I grew well on SD/-His/-Ade deficient medium, as did yeast transformed with CsTCP5 or CsTCP18. By contrast, yeast transformed with CsTCP5 or CsTCP18 grew poorly on SD/-His/-Ade medium with X-α-gal, indicating that neither CsTCP5 nor CsTCP18 was transcriptionally active. Thus, CsTCP5 and CsTCP18 are more likely to exert their regulatory functions via protein interactions or transcriptional repression.
Protein–protein interaction assays
-
Because CsTCP5 and CsTCP18 showed no direct transcriptional activation activity, we next performed Y2H and BiFC assays to determine whether they might exert their functions through interaction with CsMYB46/CsMYB105 and/or CsJAZ3/CsJAZ10/CsJAZ11. The Y2H results confirmed that CsMYB46 and CsMYB105 interacted with CsTCP18, CsJAZ3 interacted with CsTCP5, and CsJAZ11 interacted with both TCP transcription factors (Fig. 2f).
We further examined the interactions among the TCP, MYB, and JAZ proteins using BiFC assays. The control combinations of CsTCP5N or CsTCP18N with unloaded YFPC and CsMYB46C/CsMYB105C or CsJAZ3C/CsJAZ10C/CsJAZ11C with unloaded YFPN generated no fluorescence signals when transiently transformed into tobacco cells (Fig. 3). However, the combinations CsTCP5N with CsJAZ3C and CsTCP18N with CsMYB46C or CsMYB105C produced fluorescent signals that overlapped with those of the nuclear marker P19 (tagged with mCherry). Thus, both Y2H and BiFC assays indicated that CsTCP5 interacted with CsJAZ3 and that CsTCP18 interacted with CsMYB46/CsMYB105.
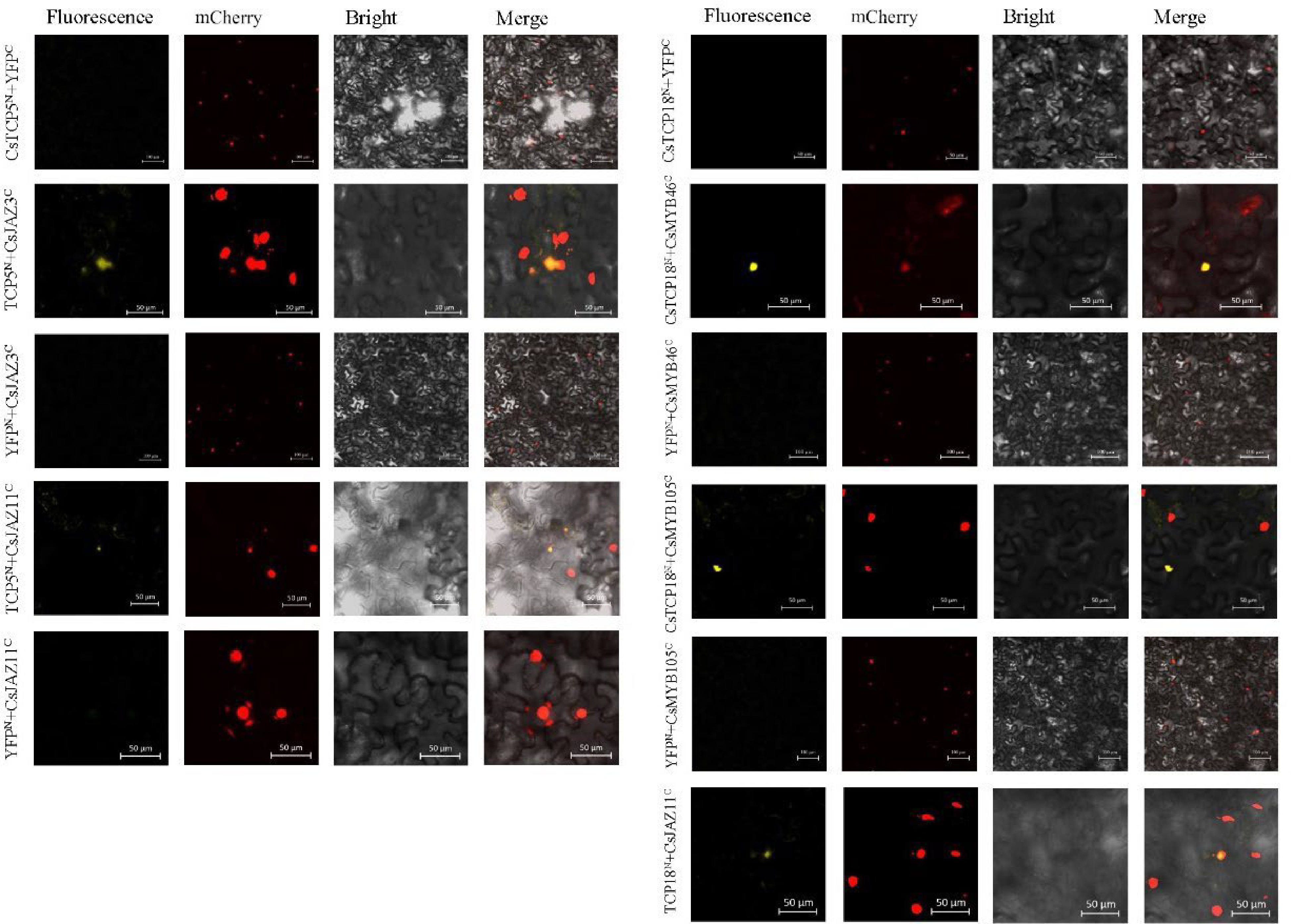
Figure 3.
BiFC analysis of protein–protein interactions of CsTCP5 and CsTCP18 with CsMYBs and CsJAZs in N. benthamiana cells. Fluorescence: yellow fluorescence channel. mCherry: red fluorescence channel. Bright: bright field channel. Merge: superimposed channel. Bar = 100 µm.
Functional analysis of transgenic Arabidopsis overexpressing CsTCP5/CsTCP18
Phenotypic analysis of transgenic Arabidopsis
-
Transgenic Arabidopsis overexpressing CsTCP5 or CsTCP18 were grown together with wild-type (WT) seedlings and seedlings transformed with the empty vector (EV) on plates with or without MeJA and ABA, and changes in cotyledons (Supplemental Figs S3 & S4), roots (Fig. 4), and germination rate (Fig. 5) were observed. Under normal growth conditions, the CsTCP5-overexpressing (OE) lines OE5.2 and OE5.4 had smaller cotyledons and fewer lateral roots than WT and EV plants, whereas the OE18.5 and OE18.7 lines were virtually indistinguishable from control seedlings (Supplemental Fig. S4a–b, Fig. 4a, c).
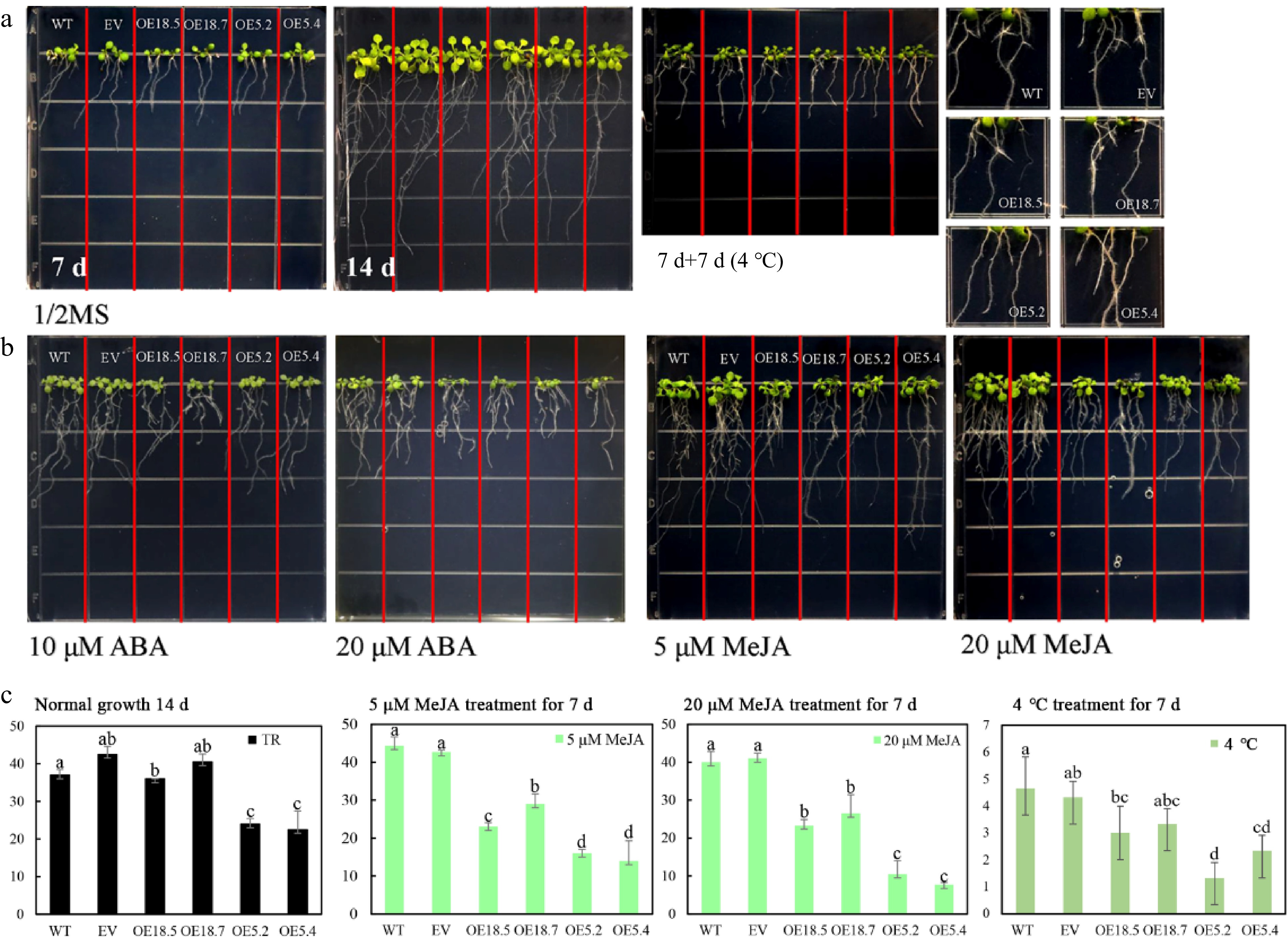
Figure 4.
Root growth of WT, EV, CsTCP18-overexpressing (OE18.7 and OE18.5), and CsTCP5-overexpressing (OE5.2 and OE5.4) Arabidopsis under different treatments. (a) Root growth after 7 d at normal temperature, 14 d at normal temperature, and 7 d at normal temperature followed by 7 d at low temperature (4 °C). (b) Root growth after 7 d with 10 μM ABA, 20 μM ABA, 5 μM MeJA, or 20 μM MeJA. Each square in the grid measures 1.5 cm × 1.5 cm. (c) Lateral root numbers of control and transgenic Arabidopsis lines under different treatment conditions. Error bars represent the SE of three replicates. Different letters indicate significant differences between measurements (p < 0.05). TR: total lateral roots.
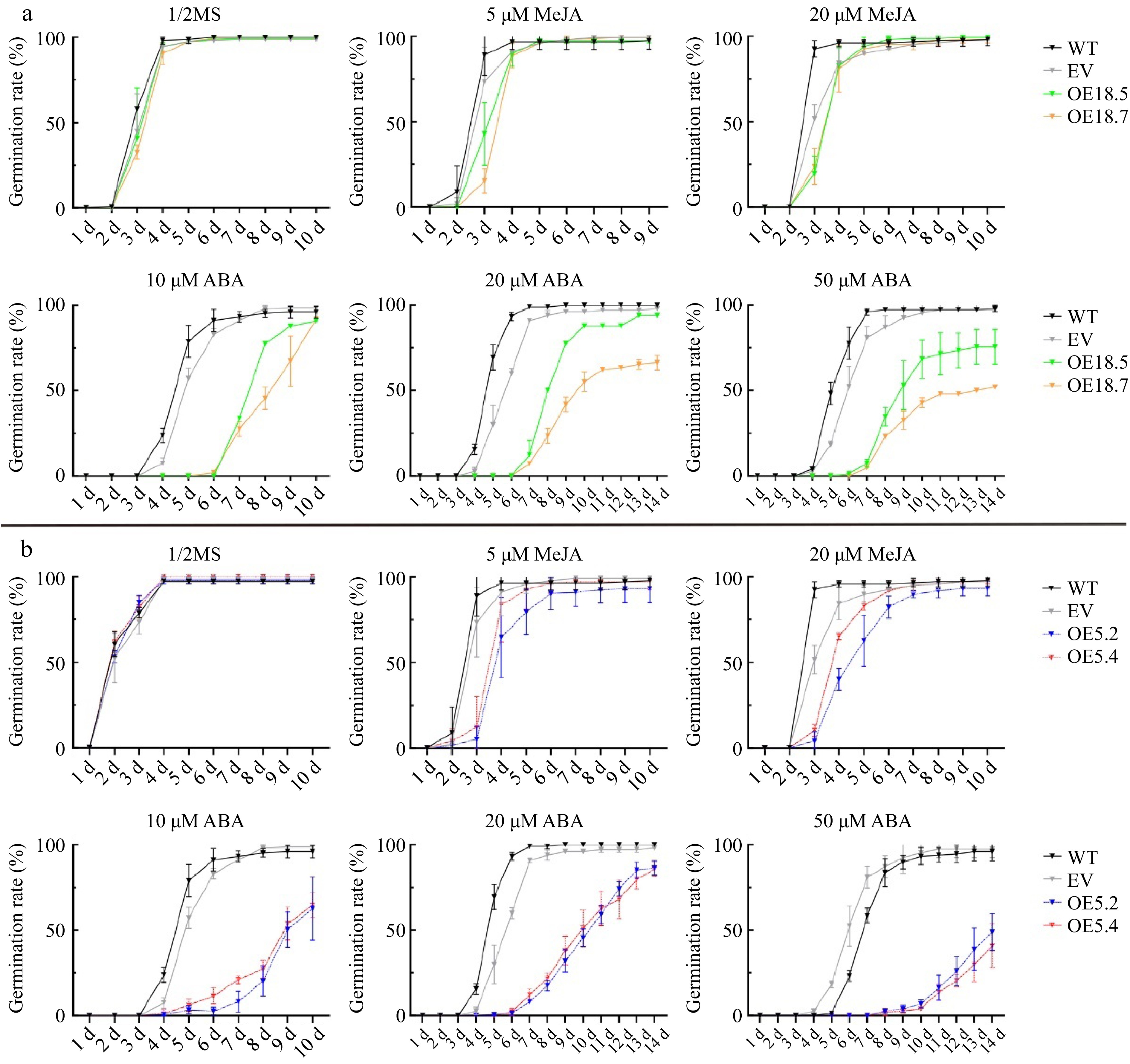
Figure 5.
Germination rates of WT, EV, CsTCP18-overexpressing (OE18.5 and OE18.7), and CsTCP5-overexpressing (OE5.2 and OE5.4) Arabidopsis exposed to different concentrations of MeJA and ABA. (a) Germination rates of CsTCP18-overexpressing Arabidopsis. (b) Germination rates of CsTCP5-overexpressing Arabidopsis. Error bars represent the SE of three replicates.
Compared with WT and EV seedlings, the CsTCP5-OE and CsTCP18-OE lines showed a greater decrease in cotyledon size under MeJA treatment, and the degree of reduction increased with increasing MeJA concentration (Supplemental Fig. S4c). MeJA treatment also inhibited lateral root production in the OE lines, particularly OE5.2 and OE5.4, and this effect was greater with 20 μM MeJA than with 5 μM MeJA (Fig. 4b–c). Compared with WT and EV Arabidopsis, the OE5.2 and OE5.4 lines showed lower germination rates under MeJA treatment, and this effect was more pronounced with 20 μM MeJA (Fig. 5b). ABA treatment also reduced the germination rates of CsTCP5 and CsTCP18 OE lines to a greater extent than WT and EV lines, and this effect increased with increasing ABA concentration (Fig. 5b). Under low-temperature treatment (4 °C for 7 d), the production of lateral roots slowed and root elongation stopped (Fig. 4c). The root hair density of OE5.2 and OE5.4 lines was clearly lower than that of WT and EV Arabidopsis under these conditions (Fig. 4a).
Analysis of drought resistance in CsTCP5/CsTCP18-OE Arabidopsis
-
To investigate the roles of CsTCP5 and CsTCP18 in plant drought resistance and their possible association with ABA signaling, transgenic Arabidopsis lines were exposed to drought and drought with exogenous ABA treatment (AD). Drought treatment reduced plant height and rosette leaf size and delayed flowering; ABA spraying did not rescue these effects (Fig. 6a; Table 1). Their higher survival rates under drought stress suggested that CsTCP5-OE and CsTCP18-OE Arabidopsis had enhanced drought resistance (Table 1).
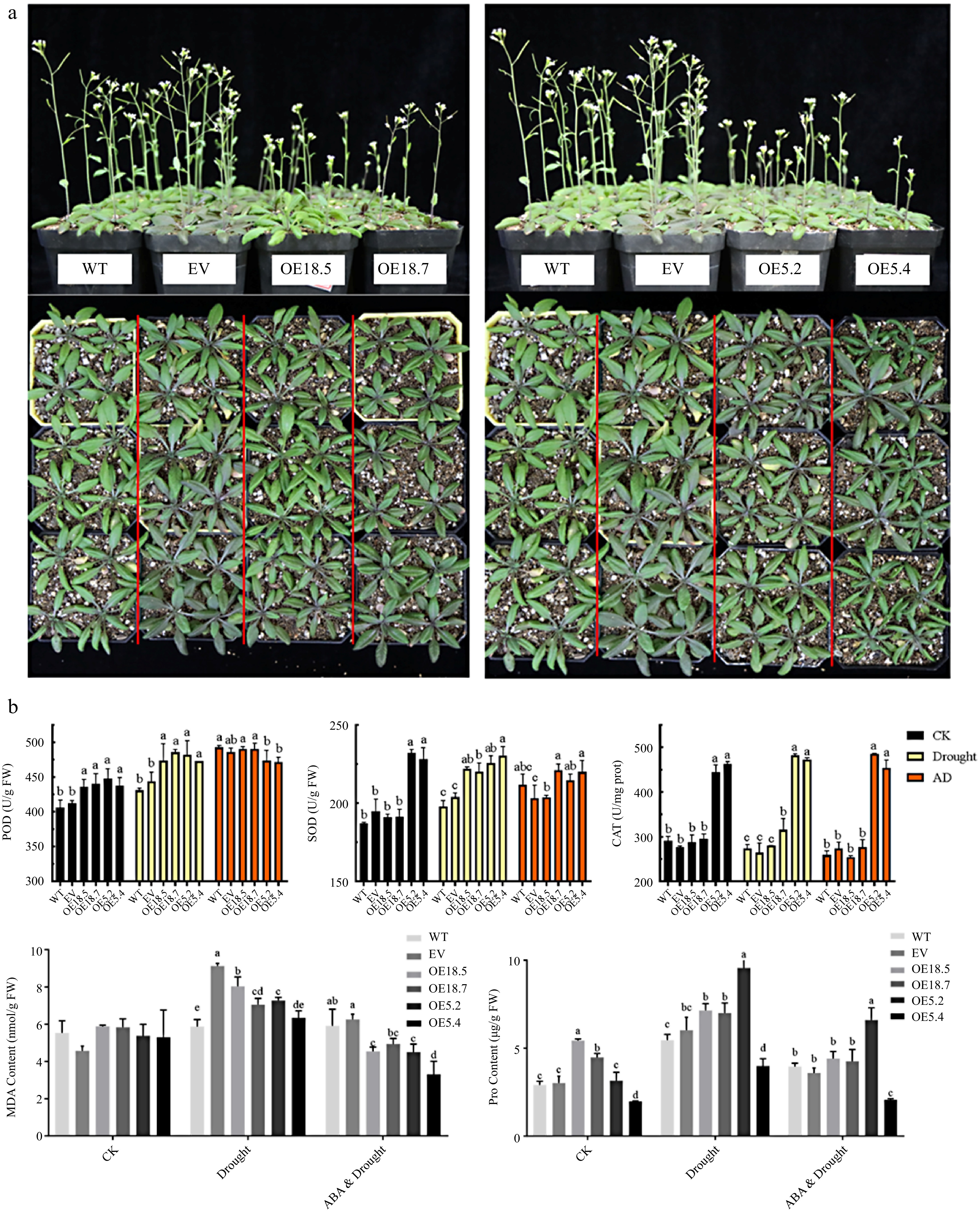
Figure 6.
Phenotypes and physiological verification of WT, EV, CsTCP18-overexpressing (OE18.5/OE18.7), and CsTCP5-overexpressing (OE5.2/OE5.4) Arabidopsis exposed to different conditions. (a) Phenotypes observations of all lines exposed to drought conditions. Bar = 7 cm. (b) Antioxidant enzyme activities and contents of malondialdehyde (MDA) and proline in Arabidopsis under control conditions, drought, or drought plus ABA application. CK: control group. AD: ABA and Drought. Different letters indicate significant differences between groups (p < 0.05).
Table 1. Numbers of plants that survived and flowered in WT, EV, CsTCP5-overexpressing, and CsTCP18-overexpressing Arabidopsis lines under drought conditions.
WT EV OE18.5 OE18.7 OE5.2 OE5.4 Survived 3/24 (12.5%) 5/24 (20.8%) 7/24 (29.1%) 9/24 (37.5%) 13/24 (54.1%) 16/24 (66.7%) Flowered 13/24 (54.1%) 7/24 (29.1%) 4/24 (16.7%) 7/24 (29.1%) 0/24 (0%) 4/24 (16.7%) The drought resistance of the WT, EV, CsTCP5-OE, and CsTCP18-OE lines was also assessed by measuring the activities of several antioxidant enzymes: peroxidase (POD), superoxide dismutase (SOD) and catalase (CAT) (Fig. 6b). Under normal growth conditions, activities of these antioxidant enzymes were significantly higher in CsTCP5-OE lines than in control plants. Under drought treatment, POD, SOD, and CAT activities of CsTCP5-OE lines remained significantly higher than those of WT and EV Arabidopsis. Under AD treatment, POD activity of the CsTCP5-OE lines was slightly lower than that observed under drought treatment and was also slightly lower than that of WT and EV Arabidopsis. SOD activity did not differ among WT and CsTCP5-OE lines under AD treatment, but CAT was significantly higher in CsTCP5-OE lines than in WT and EV lines under these conditions.
POD activity was significantly higher in CsTCP18-OE lines than in control lines under normal growth conditions, but SOD and CAT activities did not differ among these lines (Fig. 6b). Under drought, POD and SOD activities were significantly higher in CsTCP18-OE lines than in WT and EV Arabidopsis, but CAT activity was higher only in OE18.7. Under AD treatment, there were no significant differences in POD, SOD, or CAT activity between WT and CsTCP18-OE lines.
We next measured MDA and proline levels in CsTCP5/18-OE plants to assess membrane lipid peroxidation damage and the intensity of osmoregulatory metabolism (Fig. 6b). Under the drought and ABA plus drought treatments, MDA content (which is indicative of membrane damage) was significantly lower in CsTCP5/18-OE lines than in WT and EV plants, further supporting the increased drought resistance of these overexpression lines. Compared with drought alone, drought plus exogenous ABA application significantly reduced the MDA content of the overexpression lines. Apart from OE5.4, the overexpression lines had significantly higher proline contents than the WT and EV plants, and the increase in proline content was significantly greater in OE5.2 than in other OE lines. Proline content was also significantly lower in the ABA plus drought treatment compared with the drought treatment alone.
-
The tea plant is an economically valuable leaf crop, and the development and growth of its leaves strongly influence the yield and economic value of a tea garden. Natural stresses such as low temperatures and cold waves before tea harvest in the spring and high temperatures and drought in the summer and autumn have a significant effect on tea plant growth. Given the threat of global climate change, it will be essential to develop ways of mitigating abiotic stress during tea production. Numerous studies have shown that TCP transcription factors have important roles in stress response and hormone signal transduction[2−5,7], and their manipulation through breeding or genetic engineering represents a potential avenue for increasing tea stress tolerance. Nonetheless, there have been few studies on TCP transcription factors in tea. Previous research on TCP gene expression in various tissues of the tea plant revealed that CsTCP5 and CsTCP18 were highly expressed in stems and leaves, and their promoter regions contained numerous hormone-response and growth-response elements[38]. Because gene expression patterns tend to be closely related to gene function, CsTCP5 and CsTCP18 may play roles in tea leaf development, stem elongation, and stress response. In the present study, we overexpressed CsTCP5 and CsTCP18 in Arabidopsis in order to characterize their functions.
When CsTCP5 and CsTCP18 were cloned (Supplemental Fig. S1), we discovered that the CsTCP5 coding sequence contained a mutation that caused an H/Y substitution at amino acid 63 of the encoded protein and that two amino acid residues (GG) were inserted between residues 302 and 303 (Supplemental Fig. S2). These mutations were not in a conserved domain, and we considered it unlikely that they would have an effect on protein function, although this possibility cannot be excluded. The CsTCP18 coding sequence also contained a mutation (T/C) at the 293rd base, but because of codon degeneracy, the translated protein sequence was unchanged. Subcellular localization results revealed that CsTCP5 and CsTCP18 were present in the nucleus, and similar results have been reported in Pak-choi and buckwheat[25,47]. However, transcriptional activation assays in yeast cells indicated that CsTCP5 and CsTCP18 do not exhibit direct transcriptional activation activity. Overexpression of ApTCP2 from Acer palmatum and CnTCP13 from Chrysanthemum nankingense in Arabidopsis produced similar results[48,49]. We concluded that CsTCP5 and CsTCP18 were more likely to regulate plant growth and development through protein interactions.
A number of studies have examined the role of TCP transcription factors in plant morphogenesis, particularly in model plants[49−52]. Here, overexpression of CsTCP5 from the tea plant in Arabidopsis resulted in smaller cotyledons and fewer lateral roots, and these phenotypic changes were not rescued by exogenous ABA and MeJA sprays. These results demonstrate that the phenotype produced by CsTCP5 overexpression in Arabidopsis involves a regulatory pathway distinct from hormone signal transduction or depends on events downstream of initial hormone perception. Like that of CsTCP5, overexpression of OsTCP19 in Arabidopsis also led to abnormal lateral root development and reduced numbers of lateral roots[17]. PavTCP17 overexpression-Arabidopsis lines in sweet cherry had considerably fewer main branches and rosette leaves, and lateral bud development was greatly suppressed[53]. AtTCP4 has also been shown to regulate cell number and area through effects on cell proliferation, thereby influencing leaf size[54]. At the same time, the study showed that AtTCP4 overexpression in Arabidopsis increased ABA sensitivity and inhibited the promotion of leaf growth by endogenous ABA[29]. CsTCP5 may regulate the leaf development and root growth of Arabidopsis in a similar way. In this study, root growth and germination rate of CsTCP5-OE Arabidopsis were inhibited in a dose-dependent manner by ABA and MeJA treatment. Likewise, AtTCP14 is a positive regulator of embryonic growth potential during germination, but its expression is inhibited under ABA treatment; attcp14 seeds showed delayed germination compared with those of the wild type and were hypersensitive to exogenous ABA[28]. ABA can inhibit seed germination and post-germination growth of Arabidopsis, and JA can enhance the function of ABA[55]. In contrast to CsTCP5, CsTCP18 produced no discernable phenotypic differences when it was overexpressed under normal growth conditions. However, ABA and MeJA treatments caused development of some phenotypes that were not observed in the controls. Cotyledon size, germination rate, and root growth were inhibited by ABA treatment in CsTCP18-OE lines in a dose-dependent manner. MeJA treatment had little effect on cotyledon size and germination rate of CsTCP18-OE lines, but it did produce a dose-dependent inhibition of lateral root number. CsTCP5 and CsTCP18 thus showed distinct phenotypic responses to hormone application.
Tea plants thrive in high temperatures and humidity, and weather conditions in the early spring have a strong influence on tea yield and quality. MeJA plays an important role in tea plant resistance to low-temperature stress, and the MYB transcription factors CsMYB46 and CsMYB105 have been shown to participate in the low-temperature stress response[44]. We therefore investigated the expression patterns of CsTCP5 and CsTCP18 in response to MeJA treatment and low temperature stress. Low temperature significantly reduced expression of both CsTCP5 and CsTCP18, but application of MeJA after cold stress affected the two genes differently. CsTCP5 expression was significantly reduced by MeJA and low temperature treatments, particularly the combined MeJA-low temperature treatment. On the other hand, CsTCP18 expression was not altered by MeJA alone, was decreased by low temperature, and did not decrease further when MeJA and low temperature were combined. These results indicate that CsTCP5 and CsTCP18 display similar trends in response to low temperature stress in tea plants but have different response patterns to MeJA. These results are consistent with previous studies showing that some TCP family members play roles in the MeJA response[56−58], although others do not[59]. In the present study, CsMYB46 and CsMYB105 were also influenced by low temperature and MeJA, although their responses differed, suggesting that they may respond to these stimuli through different mechanisms. Low temperature stress can rapidly alter the expression of genes involved in JA biosynthesis, including a gene encoding the JAZ protein, a key inhibitor of the JA signaling pathway[60]. JAZ mediates a number of JA-regulated secondary metabolic processes, including flavonoid biosynthesis, by interacting with various transcription factors (MYB/MYC/WRKY/bHLH/COI)[32]. Here, both CsJAZ3 and CsJAZ11 were significantly downregulated by MeJA but significantly upregulated by MeJA and low temperature, consistent with previous research in which CsJAZ3, CsJAZ10, and CsJAZ11 were differentially expressed in response to various stresses and hormones[61]. Correlation analysis provided evidence for interactions of CsTCP5 and CsTC18 with CsMYB46/CsMYB105 and CsJAZ3/CsJAZ10/CsJAZ11 proteins, and a number of these interactions were confirmed by Y2H and BiFC assays: CsTCP5 interacted with CsJAZ3 and CsJAZ11, and CsTCP18 interacted with CsMYB46, CsMYB105, and CsJAZ11.
Previous research has shown that the TCP transcription factor family is important in drought stress response[62]. Yeast one hybrid and EMSA assays demonstrated that the stress/ABA-responsive gene BT2 was the direct target gene of PeTCP10[63]. Here, CsTCP5 and CsTCP18 had distinct expression patterns in response to ABA and drought. CsTCP5 expression did not change significantly under drought, increased significantly in response to ABA, and decreased significantly in the ABA and drought combination. CsTCP18 expression was significantly downregulated by individual ABA and drought treatments but increased to more than twice the control level under the combination treatment. CsTCP5 and CsTCP18 may therefore participate in ABA-dependent drought stress responses in different ways. CsTCP5/CsTCP18-OE Arabidopsis showed reduced plant height, smaller rosette leaves, and delayed flowering under drought stress compared with WT and EV plants. Additional TCP transcription factors also cause smaller leaves, shorter plants, and fewer branches when overexpressed in Arabidopsis, including GhTCP62[52], PavTCP17[53], BrrTCP2, and ZjTCP16[51,64]. We hypothesize that drought treatment may increase such phenotypic changes based on the dissimilar phenotypes of previously CsTCP5/CsTCP18-OE Arabidopsis in normal growth condition. Nonetheless, CsTCP5/CsTCP18-OE Arabidopsis showed greater survival and flowering than WT and EV plants under drought stress. OE lines generally showed higher activities of the antioxidant enzymes POD, SOD and CAT, as well as lower MDA content, which was indicative of reduced membrane lipid peroxidation and plant damage. OE lines also showed higher proline contents suggestive of enhanced osmoregulation. Thus, overexpression of CsTCP5 and CsTCP18 increased the resistance of Arabidopsis to drought stress, consistent with previous TCP overexpression studies that have supported a role for TCP proteins in plant drought resistance[19,26,63,65]. In the combined drought and ABA treatment, MDA content of CsTCP5-OE and CsTCP18-OE lines was significantly lower than that of WT and EV Arabidopsis and lower than under drought treatment alone. Except for OE5.4, the OE lines had higher proline contents than WT and EV Arabidopsis under three treatments; however the proline content of all lines was reduced compared with that measured under drought treatment alone. These results are consistent with the role of ABA in regulating plant drought resistance and thus preventing membrane lipid peroxidation. At the same time, osmotic adjustment via proline synthesis may lag behind other drought responses, and ABA application could improve plant drought resistance, thus explaining why proline content was low. For example, treatment with both ABA and abscinazole (an inhibitor of ABA 8'-hydroxylase) increased endogenous ABA concentrations, maintained leaf water potential, and reduced proline concentrations[66]. We speculate that proline content was lower because ABA treatment promoted stomatal closure, helping plants to conserve water and obviating the need for osmotic adjustment through proline accumulation. These results provide further evidence that CsTCP5 and CsTCP18 play roles in drought stress regulation mediated by ABA.
-
In conclusion, we cloned CsTCP5 and CsTCP18 from the tea cultivar 'Longjing 43' and characterized their nuclear localization, transcriptional activation activity, and interactions with other transcription factors. Seed germination, root growth, and responses to abiotic stress and hormones were significantly altered in transgenic Arabidopsis overexpressing CsTCP5 and CsTCP18, although there were differences between CsTCP5 and CsTCP18 lines. Drought caused reduced plant height, smaller leaves, and later flowering in CsTCP5/CsTCP18-OE Arabidopsis, but the OE lines showed enhanced survival and flowering compared with WT plants. OE lines also showed increased antioxidant enzyme activity, reduced membrane lipid peroxidation, and greater accumulation of the osmolyte proline, all strong evidence that these plants possessed improved drought tolerance. Overall, this study characterized the tea plant TCP transcription factors CsTCP5 and CsTCP18 and demonstrated their important roles in plant growth, development, hormone response, and stress response, providing a basis for future research into modifying the susceptibility of tea plants to environmental stimuli.
-
The authors confirm contribution to the paper as follows: project design and management: Shang X, Zhang H, Ma Y, Fang W; data analysis: Shang X, Zhang H, Zhou N, Han Z, Zhang C; manuscript preparation and revision: Zhang H, Ma Y; partial funds and consultant: Ma Y, Fang W. All authors read and approved the final manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This research was supported by the National Key R & D Program of China (2020YFE0202900), the National Natural Science Foundation of China (31972460, 32160729), the China Agriculture Research System of MOF and MARA (CARS-19), Science and Technology Support Project of Guizhou Province (ZK[2021]154), and the Priority Academic Program Development of Jiangsu Higher Education Institutions.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Huan Zhang, Xiaowen Shang
- Supplemental Table S1 Full-length primers.
- Supplemental Table S2 Treatments applied to tea plants.
- Supplemental Table S3 Treatments applied to transgenic Arabidopsis.
- Supplemental Table S4 Other primers used in this study.
- Supplemental Table S5 Quantitative primers.
- Supplemental Fig. S1 Amplification products of CsTCP5 and CsTCP18.
- Supplemental Fig. S2 Sequence alignment of CsTCP5 and CsTCP18. The top line is the cloned sequence, and the second line is the sequence from TPIA.
- Supplemental Fig. S3 Identification of T3 generation lines by genomic PCR.
- Supplemental Fig. S4 Cotyledons of WT, EV, CsTCP18-overexpressing (OE18.7 and OE18.5), and CsTCP5-overexpressing (OE5.2 and OE5.4) Arabidopsis grown with and without MeJA. a Seedlings of EV, WT, OE18.5, and OE18.7 lines under normal growth conditions. b Seedlings of EV, WT, OE5.2, and OE5.4 lines under normal growth conditions. c Seedlings of EV, WT, OE18.5, OE18.7, OE5.2, and OE5.4 lines treated with 5 μM or 20 μM MeJA.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang H, Shang X, Zhou N, Han Z, Zhang C, et al. 2023. The effective role of CsTCP5 and CsTCP18 transcription factors from Camellia sinensis (L.) O. Kuntze under drought and low-temperature. Beverage Plant Research 3:29 doi: 10.48130/BPR-2023-0029 

The effective role of CsTCP5 and CsTCP18 transcription factors from Camellia sinensis (L.) O. Kuntze under drought and low-temperature
- Received: 05 September 2023
- Revised: 24 September 2023
- Accepted: 07 October 2023
- Published online: 10 November 2023
Abstract: TCP transcription factors are essential for plant growth and environmental response, but little is known about their specific roles in growth, hormone response, and stress response in tea plants. Here, CsTCP5 and CsTCP18 were cloned from the tea variety 'Longjing 43', and subcellular localization revealed that both CsTCP5 and CsTCP18 proteins were present in the nucleus. Exogenous abscisic acid (ABA) and methyl jasmonate (MeJA) had different effects on CsTCP5 and CsTCP18 expression, but both genes were repressed by drought and low-temperature treatment. Yeast-two hybrid (Y2H) and bimolecular fluorescence complementation (BiFC) assays demonstrated that neither transcription factor was transcriptionally active but that each interacted with members of the CsMYB and CsJAZ families. Transgenic Arabidopsis plants overexpressing CsTCP5 had smaller cotyledons and fewer lateral roots compared with wild-type (WT) and empty vector (EV) Arabidopsis, and their root growth and germination rate were inhibited by ABA and MeJA treatment. Lateral root numbers also decreased significantly in CsTCP18-OE Arabidopsis after MeJA treatment. These results demonstrate that CsTCP5 and CsTCP18 have regulatory effects on cotyledon development, lateral root growth, and germination rate, and these effects can be modulated by ABA and MeJA. Under drought stress, the CsTCP5-OE and CsTCP18-OE lines exhibited greater survival, lower plant height, smaller rosette leaves, delayed flowering, increased activities of antioxidant enzymes, decreased MDA content, and increased proline content compared with WT and EV Arabidopsis. These findings suggest that CsTCP5 and CsTCP18 are important for tea plant growth, interact with ABA and MeJA pathways, and play roles in stress response.
-
Key words:
- Gene functions /
- Characterizations /
- CsTCP5 /
- CsTCP18 /
- Transcription factors