










-
An increasing number of reports have highlighted the essential roles played by UDP-UGT (Uridine diphosphate-Glycosyltransferases) in plant growth, development and the response to stress[1]. The UGT family catalyze the glycosylation of a wide range of secondary metabolites with differing levels of specificity[2−4]. UGT are also involved in the detoxification of both endogenous metabolites such as pest deterrent volatiles[5] and exogenous metabolites, such as pesticides[6]. The role of UGT in pesticide detoxification is mentioned above. However, the connection between the role for UGT in the prevention of plant infection with viruses and pesticide detoxification is unclear.
Assessing the overall influence of UGT in abiotic stress responses proves challenging due to the multitude of available reports with varying complexities. Meta-analysis involves the systematic aggregation, evaluation and quantitative analysis of separate studies or multiple datasets using statistical methods[7−9]. Applied to specific questions, meta-analyses frequently unveil statistical significances between experimental variables and physiological/biochemical indicators that are not evident from individual studies alone[10].
A comprehensive and unbiased comparative meta-analysis of available data could yield further insights into the roles of UGT in abiotic stress responses. In this study, the data from the UGT studies was divided (parameterized) based on the experimental conditions employed (eight), encompassing the UGT class and secondary metabolites studied, and the type and duration of the stress applied. Distinct physiological and biochemical indices were employed to assess the impact of UGT overexpression on the abiotic stress response.
This study aims to address the following questions: (1) What is the general effect of UGT overexpression on the abiotic stress response? (2) Were there any differences related to UGT overexpression between stressed and unstressed plants? (3) Is there any relationship between the experimental parameters utilized and the physiological indices of abiotic stress? We intend that this study provide insights into the roles of UGT in abiotic stress responses and their potential utility in bioengineering approaches for improved crop tolerances. By influencing the expression of UGT, we can have a positive impact on specific physiological indicators of plants, so as to help plants resist external stress.
-
Research articles related with UGT were identified through a systematic search of 12 electronic literature databases using Endnote and ISI Web of Science (
http://apps.webofknowledge.com ) between January 1st, 2010 and March 31st, 2022, with the search terms ('Glycosyltransferase gene + plant' / 'UGT gene + plant') and ('transgene' / 'transgenic' / 'overexpression'/ 'over-expressed' / 'over-expressed' / 'over-expressed' / 'over-express'; Supplemental File S1). The initial list contained 222 papers, out of which 97 were removed as they were determined to contain duplicate data. Of the remaining 125 articles, 112 articles were subsequently excluded for the following reasons: data unrelated to plants (23 articles); non-original research articles (4 articles); no UGT were overexpressed (58 articles); data unrelated to abiotic stress (27 articles). Finally, 13 articles were retained for meta-analysis (Supplemental File S1). Where raw data was unavailable, GetData Graph Digitizer (http://getdata-graph-digitizer.com ) was used to extract data from digitized graphs. Excel (2010) was used to organize the data and calculate the response ratio. Each study was considered as an independent unit which contains the average value and sample size of the experimental group and the control group. A total of 617 independent studies were extracted from the 13 articles, including five kinds of abiotic stress (salt, osmotic, drought, temperature and oxidation stress), with a total of 38 plant characteristics. Fifteen genes from eight groups were included in the analysis, five of which were from the G group. Eight genes were overexpressed in native plants, four genes were overexpressed in heterologous plants, and three genes were overexpressed in native and heterologous plants. (Supplemental File S1)Effect size and moderators
-
For each study, the mean of the treatment group relative to that of the control group was chosen as the response ratio for inclusion in the analysis, and its natural logarithm was used for meta-analysis to compare the effect sizes of the treatment. And the natural logarithm of the response ratio (ln R) calculated[10]:
$\rm ln\;R=ln(Y_{TC}/Y_{NC}) $ Where YTC and YNC represent the average values in transgenic plants (TC) and non-transgenic plants (NC) in abiotic stress and control, respectively.
In order to determine if the effects of UGT overexpression on the ln R values differed between the experimental conditions utilized, the variables was categorized based on various studies, which can be divided into two types: the first considered the experimental conditions, including the experimental medium type (solid, soil or liquid), stress degree and stress duration. The second category considered the experimental materials, including the plant taxonomic unit of the UGT donor and recipient (monocotyledon or dicotyledon), whether the donor and recipient are from the same species and the type of gene donor and recipient (monocot or dicot). Each variable contains at least two categorical levels of information, and each level contains at least three studies from more than two articles.
Meta-analysis
-
Comprehensive Meta-Analysis (CMA) (v. 2.2.023 Biostat, Englewood, NJ, USA; 2018) and GraphPad Prismsoftware (v.7.00) were used to construct forest plots, and random effects models were used for all analyses. The non-parametric variance method was used to weight individual studies:
$\rm V\; ln\; R = (n_{NT} + n_{NC})/(n_{NT} × n_{NC}) $ Where V ln R is the variance of the natural logarithm of the response ratio, nTC is the sample size from transgenic plants and nNC is the sample size used for the non-transgenic, control plants. The summary effect size was considered significant at p < 0.05, Q statistics were used to assess whether the effect size was showed significant heterogeneity between different data sources, and p values < 0.1 were taken as significant. I2 values were calculated to quantify the estimated influence of (real) heterogeneity in the effect size[7].
Publication bias can be problematic in meta-analyses and arises mainly due to the difficulty in publishing negative results, sampling errors in small sample studies and missing data. Therefore, funnel plots were used to allow an initial visual assessment of plot asymmetry to be made. Subsequently, three statistical methods were used to test the influence of publication bias in the data. Begg and Mazumbar rank (Kendall) correlation and Egger's regression test were used to assess the deviation revealed in the funnel plots. The Trim and Fill methods of Duval and Tweedie were used to assess the potential impact of missing studies and bias [9]. When all three tests indicated the existence of publication bias, the index was considered to have significant publication bias.
-
A total of 15 variables were screened to be included in the analysis of publication bias (Table 1). Evidence for the existence of publication bias in the meta-analysis was assessed. Five funnel plots in our displayed asymmetry, indicating that publication bias should be considered. In Kendall's test 12 of 15 summary effects displayed p values > 0.05, indicating little bias influence (no tendency for effect sizes to increase as study size decreased). The remaining three summary effects showed p ≤ 0.05, indicating publication bias. The e standard of Egger's two–tailed significance test (Egger's p value) showed that eight summary effects may be biased. The Trim and Fill method (shear and supplement method) show that there are seven summary effects with deviation from the variance of the natural logarithm ratio of response rate. The adjusted value of six of these effects was farther from zero than the original value (Ln R of summary effect), and that of the remaining four summary effects was closer to zero which jointly indicates that the summary effects have bias influence. As shown in Table 1, none of the summary effect sizes of the indicators tested were suggested to contain bias by all three statistical methods. Therefore, all indicators were included in the meta-analysis.
Table 1. Measures used in characterizing publication bias for each effect size.
Indicator effect size Summary Eeffecta Funnelb Kendallc Egger'sd Duval & Tweediee N LnR p plot tau p β p Adjusted #trim Relative anthocyanin contents 33 0.487 0.001 YES 0.39 0.00 −2.59 0.00 0.487 5 Root length 80 0.269 0 NO 0.02 0.77 0.64 0.00 −0.072 31 Germination rate 181 0.158 0 NO 0.12 0.03 −0.03 0.96 0.184 0 Survival rate 151 0.535 0 NO 0.04 0.49 0.46 0.00 0.255 37 Flavonols content 11 0.353 0.151 YES −0.15 0.59 −299.13 0.43 0.336 1 Water content 4 0.408 0.791 NO −0.60 0.50 −320.03 0.22 0.108 0 Water loss 48 0.014 0.9 NO −0.38 0.00 −6.71 0.00 0.014 0 Stomatal aperture 11 0.435 0 YES −0.36 0.14 −0.62 0.01 0.436 0 completely closed Stomatal 4 0.280 0.492 YES −0.50 0.31 −1277.39 0.05 0.280 0 Partially closed Stomatal 4 0.229 0.575 YES −0.50 0.50 −1851.04 0.04 0.229 0 completely open Stomatal 4 0.408 0.467 NO −0.17 1.00 −513.78 0.60 −0.392 1 Electrolyte leakage 78 −0.31 0.001 NO −0.01 0.87 −274.24 0.55 −0.413 18 chlorophyll content 12 0.164 0.488 NO −0.14 0.58 −382.07 0.14 0.164 0 Proline content 14 0.25 0.253 NO −0.01 1.00 −151.85 0.79 0.250 0 Soluble sugar contents 12 0.097 0.682 NO −0.17 0.49 −707.59 0.03 0.097 0 a Summary effect: N = number of studies, ln R = natural log of the overall summary effect, p = probability that the summary effect ≠ 0; b Funnel plot appears asymmetrical; c Begg and Mazumdar Kendall rank correlation: tau = rank correlation coefficient (with continuity correction), two-tailed p = probability that the study effect sizes are correlated with their sampling variances; d Egger's linear regression: β = intercept of the regression line, p = probability of significant asymmetry in the study effect size/study size association. The regression runs through zero if the funnel plot is symmetrical. The size of the deviation of the intercept from the origin is a measure of asymmetry, with a two-tailed p < 0.05 indicating significant asymmetry. e Duval and Tweedie trim and fill: adjusted summary effect after imputing missing studies using an iterative trim and fill procedure, #trim = number of studies imputed in the trim and fill exercise. Analysis of heterogeneity analysis
-
In meta-analysis, the observed variations are considered to consist of (real) heterogeneity between the studies and random error equally distributed across studies. Table 2 shows the results of Cochran tests for heterogeneity between the data sources. Several authors have advised caution in the interpretation of the resulting associated p and I2 values. Substantial real dispersion of real effects may also lead to a Phetero > 0.1. Therefore, according to the observed pattern, the random effects model was selected to conduct subgroup analysis of the summary effect values of the different variables.
Table 2. Heterogeneity statistics for the 30 summary effect sizes under overexpressing UGT genes.
Category Qta Pheterob I2c Change (%) Relative anthocyanin contents (S) 2.07 1 0 73 Root length (S) 112.47 0 45.763 44 Germination rate (S) 2303.961 0 93.747 23 Survival rate (S) 190.658 0 38.109 115 Flavonols content (S) 0.471 1 0 44 Water content (S) 0.07 0.995 0 50 Water loss (S) 11.917 1 0 2 Stomatal aperture (width/length) (S) 2.743 0.949 0 56 Completely closed stomatal (%) (S) 0.3 0.584 0 73 Partially closed stomatal (%) (S) 1.389 0.239 27.982 49 Completely open stomatal (%) (S) 0.498 0.48 0 −37 Electrolyte leakage (%) (S) 4.731 1 0 −33 Chlorophyll content (S) 0.291 1 0 21 Proline content (ug·g−1·FW) (S) 0.365 1 0 62 Soluble sugar contents (ug·g−1·FW) (S) 0.126 1 0 30 Relative anthocyanin contents (N) 0.662 0.995 0 28 Root length (N) 0.96 1 0 −1 Germination rate (N) 58.821 0.005 42.197 −4 Survival rate (N) 0 1 0 0 Flavonols content (N) 0.161 0.923 0 37 Water content (N) 0 Water loss (N) 0 1 0 0 Stomatal aperture (width/length) (N) 0 0.994 0 −3 Completely closed stomatal (N) 0.017 0.897 0 1 Partially closed stomatal (N) 0.02 0.887 0 6 Completely open stomatal (N) 0.039 0.844 0 −12 Electrolyte leakage (N) 0.684 1 0 4 Chlorophyll content (N) 0.003 0.957 0 2 Proline content (N) 0.088 1 0 −6 Soluble sugar contents (N) 0.062 1 0 −7 a Qt, total observed variation among studies; b phetro, probability that Qt was due entirely to sampling error and not to real variation among studies; c I2, percentage of heterogeneity due to variation among true effects. Positive values indicating UGT overexpression promotion and negative values indicating UGT overexpression inhibition. (S) represents the heterogeneity of stressed plant; (N) represents the heterogeneity of non-stressed plant. Overall summary effects
-
The summary effect value reflects the degree of influence of UGT overexpression on the effect value of the physiological indicators under stress and without stress, and the p-value reflects its significance. The 15 summary effect values were derived from 566 independent studies involving five different species. Most of the UGT were from monocotyledonous sources (368 studies) The plant species used for UGT overexpression included O. sativa (60 studies) and A. thaliana (506 studies). Figure 1 shows a forest plot summarizing the effects of UGT overexpression on the 15 indicators in plants under abiotic stress (left) and control conditions (right). The summarized effect responses of 6/15 indicators were significantly altered by abiotic stress in UGT overexpressing plants, including anthocyanin content, root length, survival rate, germination rate, stomatal aperture and electrolyte permeability (p ≤ 0.05). There was no significant effect on the comprehensive effect value of six indicators such as water loss. Relative to WT plants, the overexpression of UGT significantly increased the survival rate of plants under abiotic stress by 97%, The overexpression of UGT significantly decreased electrolyte leakage by 33% in plants under abiotic stress conditions, indicating that the overexpression of UGT was beneficial to early growth of plants and prevent damage of plasma membrane. However, under unstressed conditions, none of the indicators studied were significantly affected by overexpression of UGT.
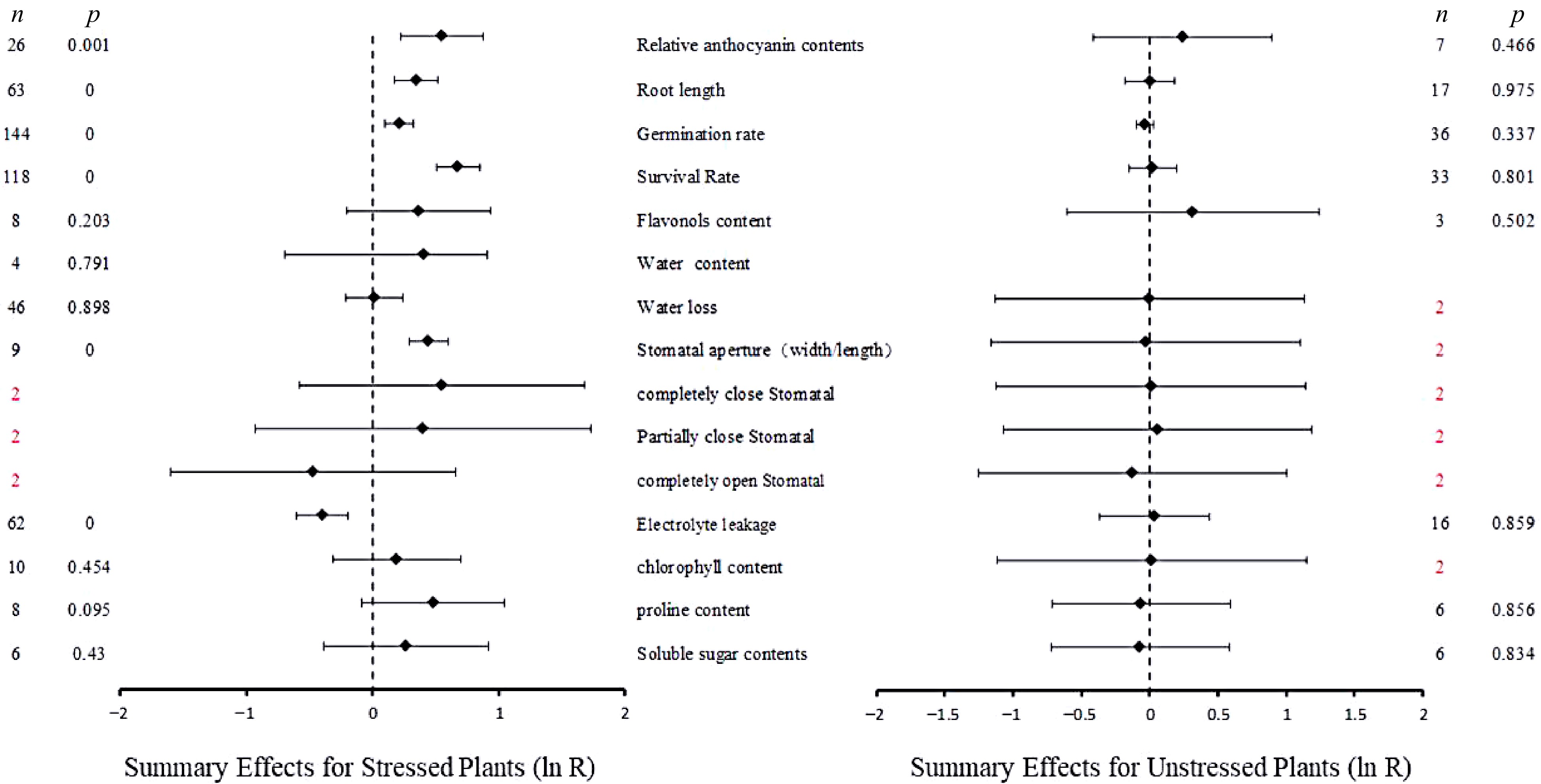
Figure 1.
Summary effect sizes (ln ratio of the response in UGT overexpressing plants/ WT plants; ln R). Horizontal bars associated with summary effects represent the 95% confidence intervals. n is the number of studies contributing to each summary effect. p-values ≤ 0.05 were taken as significant.
Subgroup analysis
-
The six indicators significantly displaying significantly alterations after both abiotic stress and the overexpression of UGT (p ≤ 0.05; Fig. 1) were subjected to subgroup analysis by categorizing the data according to the different experimental materials and conditions utilized in their measurement. Of these, the response effect on stomatal aperture showed no obvious differences between the different data sources, likely due to the low number of replicates available for this indicator. The analyses of the remaining five indicators are presented below.
Root length
-
The influences of 10 experimental conditions on the response effect UGT overexpression are shown in Fig. 2. UGT overexpression had a significant and positive effect on root length (71%) under salt stress, but negative under osmotic stress (−14%). Under abiotic stress, soil as culture medium had significant influence on root length (34%). Considering the donor source of the UGT, the overexpression of those from Sporobolus stapfianus were seen to have the greatest positive effect (95%) on root length under abiotic stress, whereas those from A. thaliana had a small, but negative effect (−15%) and those of O. sativa and Carex rigescens had positive, but insignificant effects. UGT derived from monocots or dicots (donor type) showed positive or negative effects on root length under abiotic stress, respectively. Where the plant system used for expression (recipient) was a monocotyledon, it had a significant effect on the increase of root length . The effects of UGT over expression on root length under abiotic stress conditions was obviously reduced when the donor and recipient were both from either monocots or dicots (−13%) than when they differed (84%).
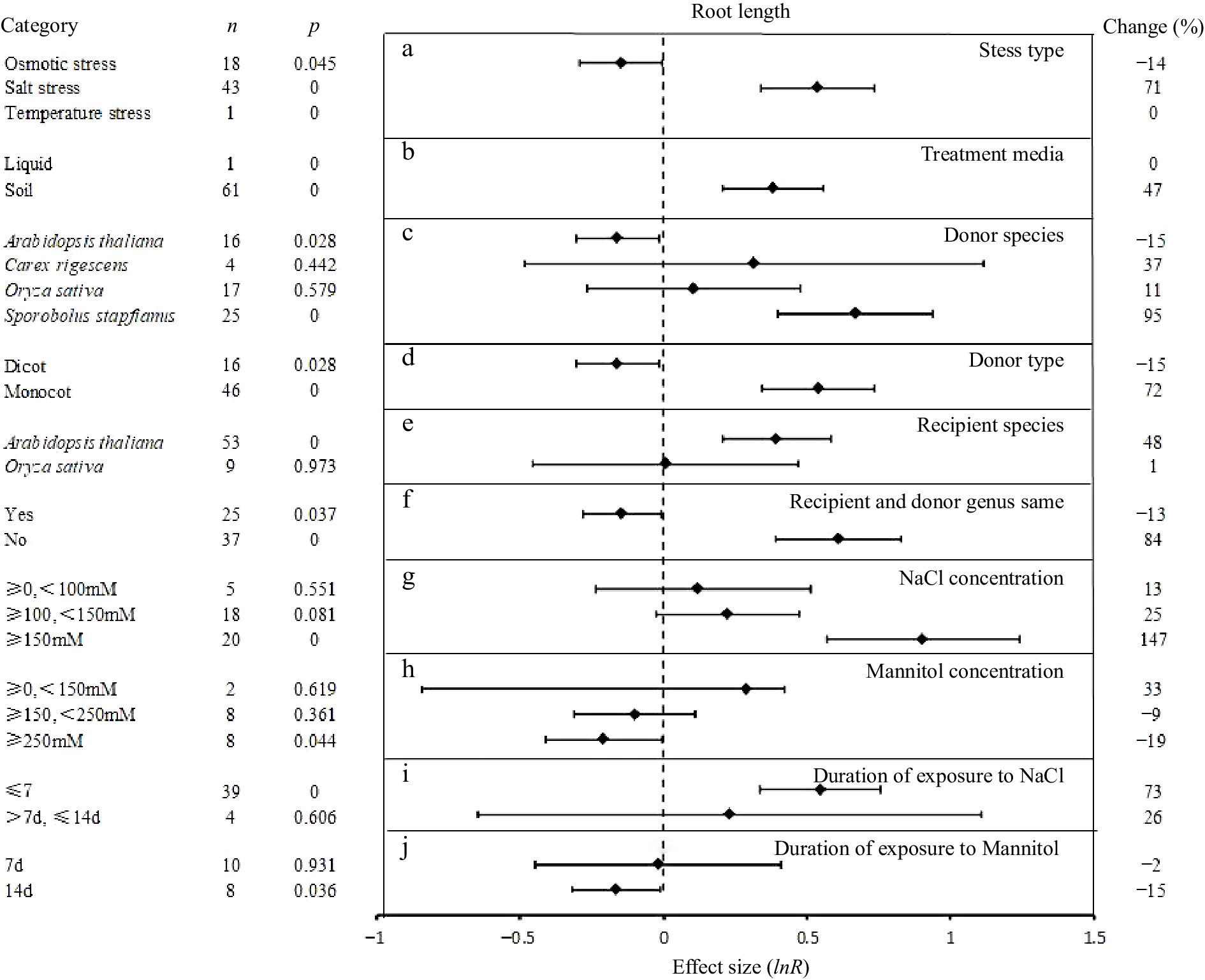
Figure 2.
The effect sizes of UGT overexpression on root length under different experimental conditions of abiotic stress. The horizontal error bars represent the 95% confidence interval. .
Under salt stress, the overexpression of UGT has a positive and significant effect in treatments with ≥ 150 Mm NaCl over 7 d of treatment. In osmotic stress induced by mannitol, UGT overexpression had a significant, negative effects on root length at ≥ 250 mM (−19%), and at longer times of exposure (−15%, 14 d).
Germination rate
-
The overexpression of UGT was seen to have a significant negative effect on germination rate under osmotic stress (−27%), which can be seen in Fig. 3. The inhibitory effects (30%) were seen with mannitol concentrations between 150−250 mM. When mannitol concentration was above 250 mM, the inhibitory effect on germination rate was 24%. Conversely, significant and positive effects on the germination rate was seen under salt stress (41%) and this effect was greater at concentrations above 150 mM (66%) compared to 100−150 mM (22%). In addition, the beneficial effect was greater in salt treatments of longer duration (0−7 d, 37%; 7−14 d, 173%).
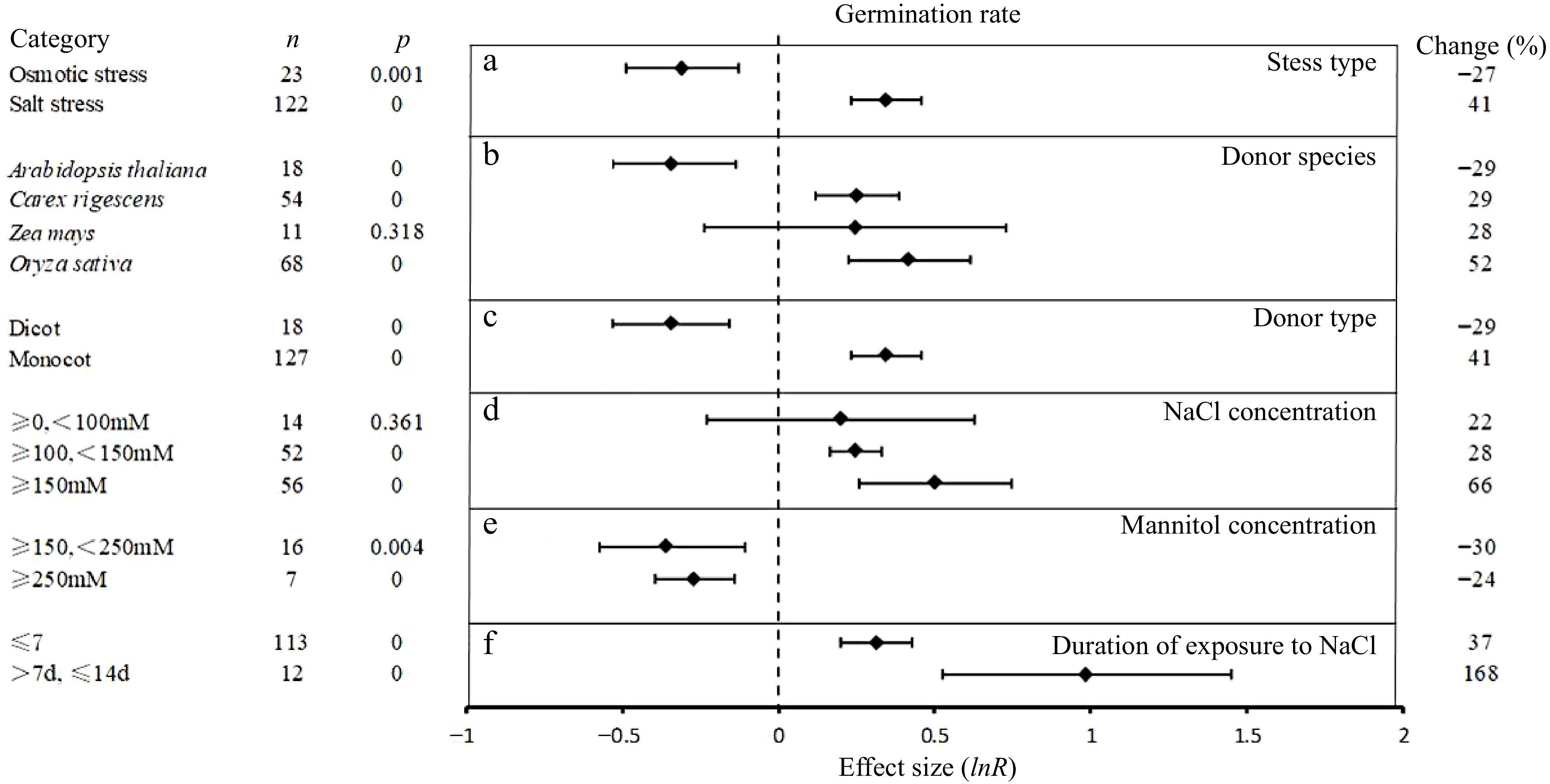
Figure 3.
The effect sizes of UGT overexpression on germination rate under different experimental conditions of abiotic stress. The horizontal error bars represent the 95% confidence interval.
By distinguishing the taxonomic source of the UGT we observe that those from C. rigescens and O. sativa increased the average germination rate of by 29% and 52%, respectively, but only those of O. sativa had significant effects. Conversely, UGT from Arabidopsis had a significant negative effect (−29%).
Survival rate
-
Overexpression of UGT had an overwhelmingly significant and positive effect on the survival rate of plants subjected to abiotic stress (Fig. 4). In 151 independent studies we examined the influences of 12 experimental variables on this survival. Relative to the WT control, the overexpression of UGT had the largest effect on the oxidative stress response (501%), followed by salt (171%), osmotic (137%), drought (76%) and temperature (62%) stresses. The different growth media had similar positive effects on survival rate, but the data from in plants grown in soil or solid media was more significant. UGT from Zea mays promoted the survival rate in transgenic plants by 232%. UGT from monocots and dicots had similar positive effects on the plant survival rate. UGT overexpression in O. sativa had a similar effect on survival rate under abiotic stress (142%) than in A. thaliana (113%). Similar positive effects on plant survival were seen whether the UGT was endogenous or exogenous to the overexpressing species.
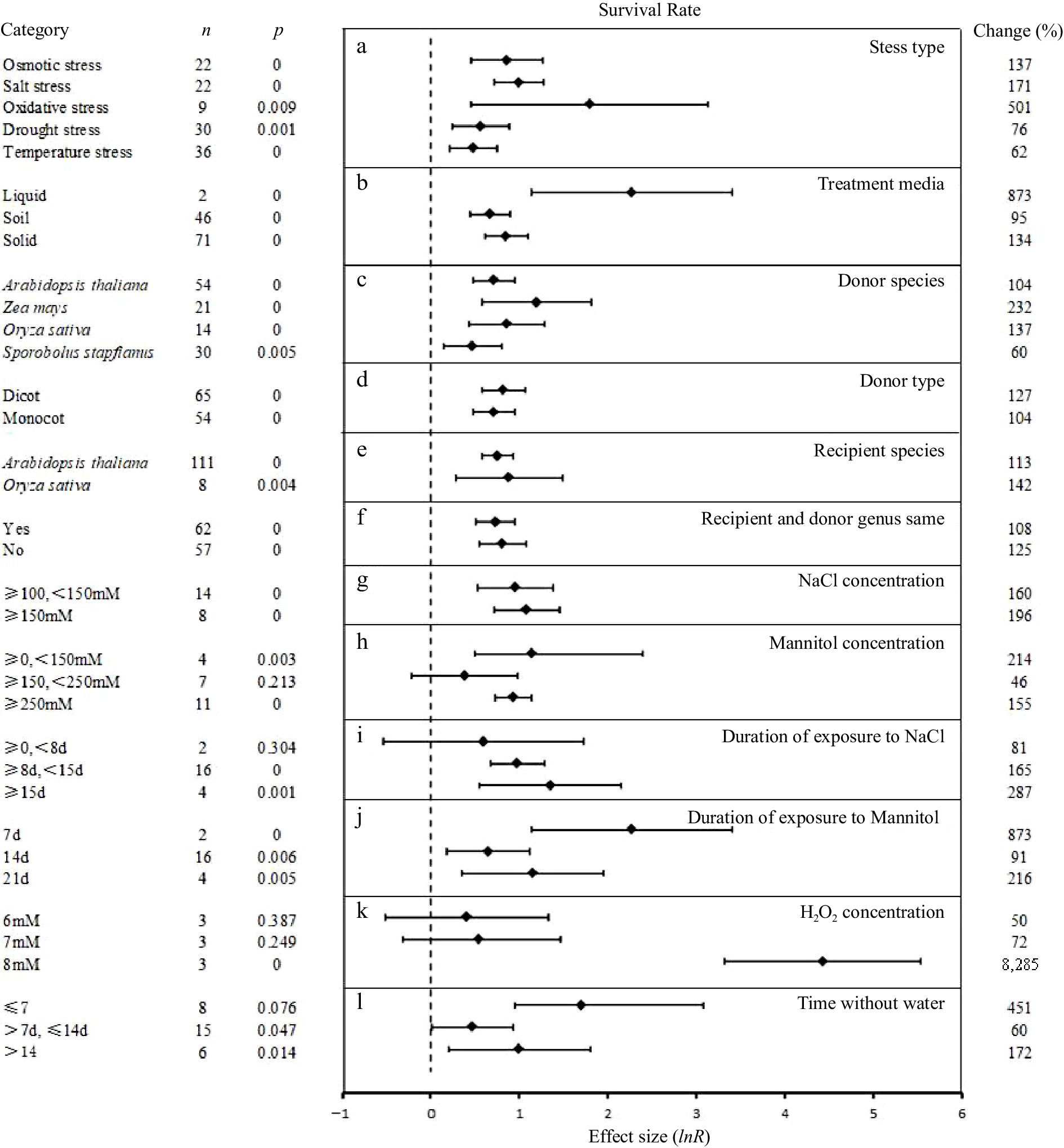
Figure 4.
The effect sizes of UGT overexpression on survival rate under different experimental conditions of abiotic stress. The horizontal error bars represent the 95% confidence interval..
Under salt stress, UGT overexpression had on average, larger impacts in plants subjected to NaCl concentrations above 150 mM and for more than 2 weeks. The overexpression of UGT in plants treated with H2O2 (to simulate oxidation stress) at 6 and 7 mM had relatively small (50%−72%) and less significant positive effects on the plant survival rate. However, larger (8,000%+) and more significant protective effects were observed in plants exposed to 8 mM.
Under osmotic stress, the survival rate of the transgenic plants increased by 214% when the concentration of mannitol was between 0−150 mM, and was higher in plants exposed for a shorter period (7 d; 873%) than over longer periods (14−21 d; 15%−216%) Similarly, UGT overexpression exerted a larger measurable protective effect against short-term drought stress (≤ 7 d; 451%) than over longer periods (> 7 d, ≤14 d; 60%).
Relative anthocyanin contents
-
UGT promoted a greater accumulation of anthocyanins in plants subjected to abiotic stresses (Fig. 1). An analysis of the influence of experimental conditions on this promoting effect revealed that although there was some differences in the average size effects, no significant differences between the treatments were found (Fig. 5). Figure 5 shows the influences of four experimental conditions on the effects of UGT overexpression on anthocyanin contents in plants under abiotic stress (other variables are excluded because they do not satisfy the inclusion of at least two categorical levels). Under salt stress, UGT overexpression increased anthocyanin content by 113%. Plants growing in soil had significant positive effects on anthocyanin content (165%). Larger effects of UGT overexpression on anthocyanin levels were obtained when osmotic stress was simulated with mannitol (106%) than with sucrose (24%). In cold stressed plants, UGT overexpression promoted higher anthocyanin content under chill (113%) than freezing temperatures (34%).
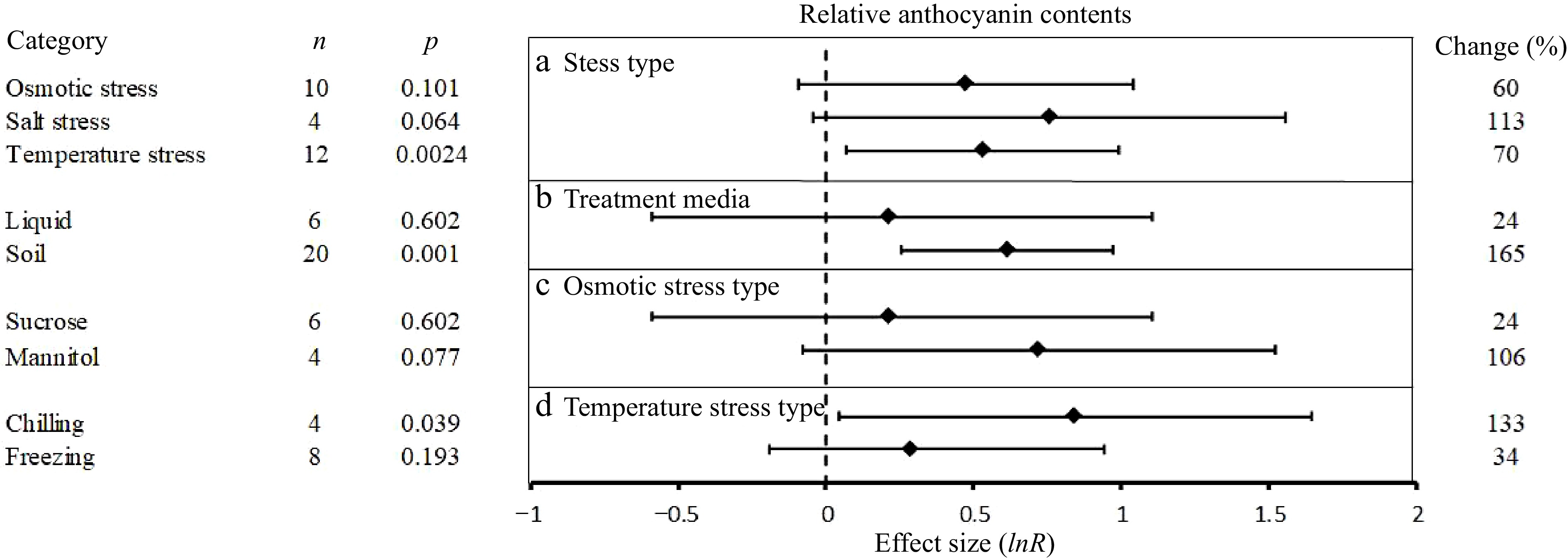
Figure 5.
The effect sizes of UGT overexpression on relative anthocyanin contents under different experimental conditions of abiotic stress. The horizontal error bars represent the 95% confidence interval.
Electrolyte leakage
-
The influence of seven experimental variables on the effect of UGT overexpression on electrolyte leakage is presented in Fig. 6. Under temperature and salt stress, UGT overexpression significantly reduced the electrolyte leakage by 34% and 25%, respectively. UGT from O. sativa effected the largest reduction in electrolyte permeability (−36%), while C. rigescens had no significant effect. The effect was not influenced by the taxonomic source of the UGT. The reduction in electrolyte leakage by UGT overexpression was not significantly affected by the choice of A. thaliana or O. sativa, as the host plant species, this effect was larger in O. sativa (−43%). UGT in endogenous and exogenous overexpression had a similar effect on electrolyte leakage. On average, UGT overexpression promoted a reduction in electrolyte leakage under chilling stress (−43%) and freezing stress (−33%). In salt stress, the effect on the electrolyte leakage of overexpressed plants containing exogenous UGT had no significant between different concentrations.
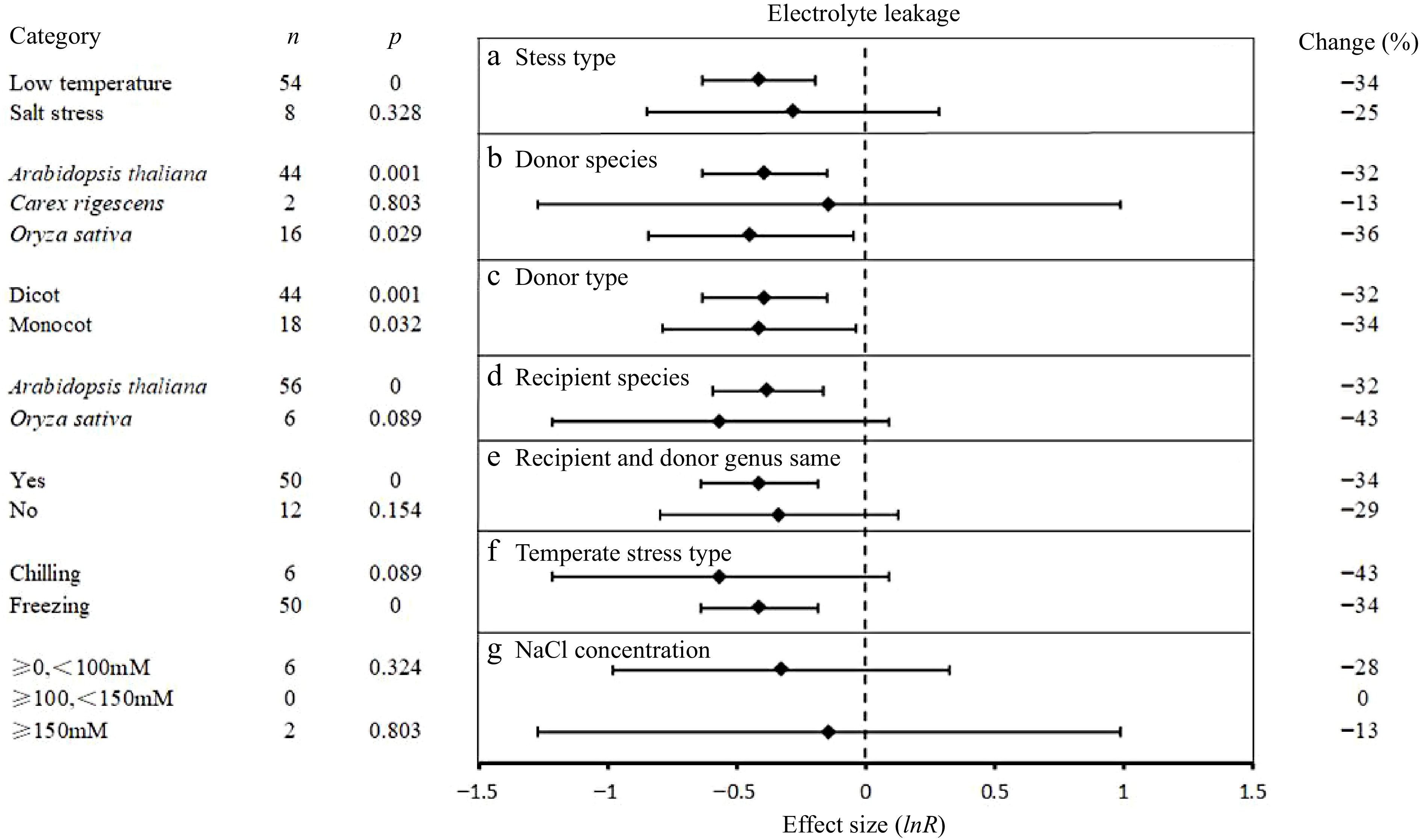
Figure 6.
The effect sizes of UGT overexpression on electrolyte leakage under different experimental conditions of abiotic stress. The horizontal error bars represent the 95% confidence interval.
-
Previously reported studies of UGT have primarily focused on their effects on secondary metabolite levels and their substrate preferences[11−14]. However, there has not been a systematic and statistics-based assessments effect of UGT overexpression on the plant response to abiotic stresses. Therefore, through meta-analysis, we analyzed the effects of UGT overexpression on specific indicators related to plant response to abiotic stress, so as to reveal potential contributions of UGT to abiotic stress tolerance. We also examined the relationship between these indicators and other experimental conditions (such as species, abiotic stress type, experimental medium, experimental concentration, experimental time, etc.) to understand how these variables effect the subgroup indicators and the underlying source of heterogeneity. This approach aims to reduce the ambiguity in current research on the functions of the plant UGT family.
The summarized effects of UGT overexpression on indicators (response effect sizes) were analyzed in plants under stress and non-stress conditions. Due to the low study number of studies (n < 3) available for some indicators, we selected 15 for subsequent analysis. As a modifier enzyme involved in secondary metabolic processes, overexpression of UGT under normal conditions can influence the content of their substrates[15−17]. It can be seen from Fig. 1 that UGT had no significant effect on any indicator under unstressed conditions, but had a positive effect on the content of anthocyanin, which may be related to the synthesis of UGT substrates[18,19]. Abiotic stress wildly recognized in mobilizing phenylpropanoid pathway[20], often resulting in accumulation of anthocyanin precursors. UGT is a pivotal enzyme for the formation of stable anthocyanin[21, 22]. Studies have shown that there is a significant positive correlation between the content of anthocyanin in pericarp and UGT mRNA levels[23]. In comparison to WT plants, UGT overexpression resulted in higher anthocyanin levels in plants subjected to abiotic stress, but not in unstressed plants. Presumably, the lack of anthocyanin precursors is rate-limiting for anthocyanin synthesis, which is indicated under stressed conditions. UGT levels become the rate-limiting step in anthocyanin synthesis in the WT plants, which means that the overexpressed UGT selected for expression in these studies were clearly functional as UDP-glucose flavonoid glucosyltrasferases (UFGTs). The increased anthocyanin content resulted either directly from an increased partition of flavonoid and anthocyanidin precursors into anthocyanin synthesis, or indirectly through positive feed-back effects on precursor biosynthesis.
The overexpression of UGT under abiotic stress did not appear to have a noticeable effects on the rate of water loss, Unexpectedly, we observed that the overexpression of UGT in stressed plants promoted stomatal opening. This observation contradicts numerous reports that indicate restricting stomatal opening is a common response to reduce water loss under various stress conditions, including drought[5, 24] and related stressors like osmotic and saline stresses. Furthermore, the overexpression of UGT was also seen to enhance electrolyte leakage under abiotic stress. Electrolyte leakage is indicative of changes in the membrane permeability and/or perturbations in ion homeostasis, and can be used as an important indicator of membrane damage by stress, such as freezing temperatures, or plasmolysis after drought or osmotic stress. The role of UGT in stomatal movements and ion flux homeostasis under abiotic stress conditions are therefore of potential interest for further study. In the overall summary effect, it can be seen that overexpression of UGT has significant positive effects on root length, germination rate and survival rate, so UGT may play an important role in plant growth and development regulation.
The overexpression of UGT is known to impact on many resistance-related indicators. For example, proline can function as an osmotic regulator under drought conditions and as a free radical scavenger under conditions of oxidative stress[25]. Soluble sugars can maintain the stability of plant proteins and reduce the water potential of cells by vitrifying the fluid around chloroplasts[25]. However, data for these metabolites were not underrepresented in the selected studies so were not included in this meta-analysis. In fact, there is a general lack of statistically relevant data of key physiological indicators of abiotic stress. Numerous studies have demostrated that the expression of related synthetic genes is up-regulated under drought stress[26], salt stress[27] and low temperature stress[28]. This often leads to an increase in anthocyanin content in plants as a mechanism to combat adversity. Anthocyanins are usually included in experiments as resistance indicators. Two of the papers involved in this meta-analysis included anthocyanin content in the determination range, but the UGT gene substrate in these two studies was related to anthocyanin. Therefore, it is worth further consideration whether the overexpression of UGT gene with active substrate non-anthocyanin can enhance the ability of plants to withstand adversity and take the change of anthocyanin content as a reference index.
The overexpression of UGT was seen to moderate the impact of salt stress on most indicators, such as root length, germination rate and survival rate. The ability of plants to maintain the intracellular K+/Na+ ratio is crucial for plant tolerance to salt stress[29]. In apple, the overexpression of a UGT was shown to reduce salt-stress damage to the antioxidant system and enhance the efflux of sodium ions and the influx of potassium ions, thus helping maintain intracellular ion homeostasis[30]. However, the studies on transgenic plants under stress conditions involved in this analysis did not involve relevant ion property experiments. With the increase of salt concentration and treatment time, the response to UGT gene overexpression was also enhanced in germination rate, suggesting that UGT gene was closely related to salt stress in the early stages of plants.
Interestingly, the effects of UGT overexpression on the germination rate and root length were opposite in plants under saline and osmotic stress. This suggests that UGT may be differentially employed in the response to these two stresses and requires further study.
UGT possesses a highly conserved PSPG domain in the C-terminal, responsible for selectively binding of nucleoside sugar donors[31, 32]. The binding of the aglycone acceptor is chiefly determined by multiple positions, mostly in the N-terminal region. UGT genes with high homology genes may have similar functions in response to abiotic stress. The overexpression of UGT from monocotyledons showed a very obvious positive effect on germination rate and root length, survival rate and electrolyte permeability in plants under abiotic stress, while those from Arabidopsis had a negative effect on germination rate and root length. It important to note in this study, data from dicotyledonous plants were mainly from A. thaliana, while data from monocotyledonous plants came from a wider range of species. In most of the studies involved, the UGT were from monocotyledons (368 of 566 studies) and overexpressed in dicotyledonous (505 of 566 studies). The effects of UGT overexpression on electrolyte leakage, root length and survival rate were found to be more obvious when the taxonomic origin of the UGT (monocot or dicot) was different from the plant system used for expression. The reason behind this observation is not clear and further experimental exploration is needed.
A large number of investigations have provided evidence for important roles for UGT in secondary metabolism, and the analysis provided insights into focused aspects of this important field of UGT in the response and tolerance to abiotic stress. But the research on the relationship between UGT and abiotic stress is still lacking. In this study, an in-depth analysis of how various variables affect the index after overexpression of UGT under abiotic stress is conducted. The meta-analysis provides certain basis for how to adjust experimental variables to maximize the function of UGT, and can help us further our understanding of UGT function in abiotic stress, and suggest directions for the functional exploration of UGT in future plant breeding efforts for the improvement of abiotic stress tolerance.
-
The authors confirm contribution to the paper as follows: study conception and design: Zhu X, Fang W; data collection: Chen Y, Duan Y, Shen Q; analysis and interpretation of results: Chen Y, Cao Y, Deng D, Gao Q; draft manuscript preparation: Chen Y, Zhu X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are not publicly available due to management requests, but are available from the corresponding author on reasonable request.
This research was supported by the National Natural Science Foundation of China (31972460, 32160729), the China Agriculture Research System of MOF and MARA (CARS-19), Science and Technology Support Project of Guizhou Province (ZK[2021]154, [2022]07), and the Priority Academic Program Development of Jiangsu Higher Education Institutions.
-
The authors declare that they have no conflict of interest.
- Supplemental File. S1 Studies.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen Y, Cao Y, Duan Y, Deng D, Gao Q, et al. 2023. The effects of overexpressing UDP-Glycosyltransferases genes on the plant response to abiotic stress: a meta-analysis. Beverage Plant Research 3:28 doi: 10.48130/BPR-2023-0028 

The effects of overexpressing UDP-Glycosyltransferases genes on the plant response to abiotic stress: a meta-analysis
- Received: 11 August 2023
- Revised: 17 October 2023
- Accepted: 19 October 2023
- Published online: 02 November 2023
Abstract: Uridine diphosphate (UDP)-Glycosyltransferases (UGT) play essential roles in modifying secondary metabolites during the plant life cycle and are also involved in the response to abiotic stresses. However, the plant UGT family is vast and the available data describing their role in abiotic stress responses is varied and intricate, so that their potential roles are often obscured. To address this, a meta-analysis was conducted to assess the effects of overexpression of UGT on various plant physiological indicators under abiotic stress. Out of the 15 plant characteristics examined in UGT overexpressing plants, 10 showed an increase of over 30% , while two plant characteristics decreased by more than 30%, while only three indices were significantly affected under non-stressed conditions. Notably, UGT had a significant and positive effect in salt-stressed plants. This study sheds light on the complex role of UGT in abiotic stress and can provide valuable guidance for future research on UGT functions and their genetic manipulation in crop breeding programs for improved abiotic stress tolerances.
-
Key words:
- Meta-analysis /
- UGT gene /
- Overexpression /
- Abiotic stress