










-
Pollen dormancy is of great significance to plant breeding which utilizes pollens as germplasm resources. To achieve double fertilization in plant, it is very crucial for pollen to maintain a dormancy state during spreading and preservation, and to only initiate pollen germination and tube growth after contacting the female stigma, then immotile sperm cells rely on pollen germination and tube tip growth to penetrate long distance female tissue and to reach the embryo sac.
However, pollen dormancy is often disrupted by various factors including temperature, humidity, nutrients (minerals and carbohydrates), etc.[1−3]. Similar to seeds, desiccation is also an important prerequisite for maintaining pollen dormancy before hydration. Humidity is a major climatic factor because dry mature pollen can remain dormant for extended periods in dry-air environments until it acquires water from stigma[4,5]. So far, little is known about how dormant mature pollen senses humidity to initiate germination in plant. The mutant of raring-to-go (rtg) was firstly identified as defective in pollen dormancy in Arabidopsis, with premature germination of pollens within the anther under high humidity conditions[4]. To date, the underlying mechanism remains unknown as the mutated gene for rtg has yet to been identified. The metabolic state of pollen is crucial for maintaining dormancy. Hashida et al. discovered that the nmnat mutant, lacking the nicotinate/nicotinamide mononucleotide adenyltransferase, exhibits precocious pollen germination within the anther under high humidity due to a deficiency in NAD+ (nicotinamide mononucleotide)[5]. Another mutant with a mutation in the inositol polyphosphate 5-phosphatase gene displays abnormal accumulation of Ins (1,4,5) P3 and Ca2+, resulting in the breaking of pollen dormancy and premature pollen germination within the Arabidopsis anther[6]. Additionally, FLA14 encoding a fasciclin-like arabinogalactan protein has been implicated in maintaining pollen dormancy under high humidity conditions in Arabidopsis, although the exact mechanism remains unknown[7]. In 2016, Ju et al. discovered that JGB as a negative regulator was involved in JA biosynthesis-mediated pollen dormancy in moist environments[8].
Termination of dormancy in pollen means the beginning of germination, characterized by pollen tubes with polarized tip growth. This process is intricately regulated and involves various cellular events, such as cytoskeleton organization, cell wall modification, and ion transport[9]. ROP1, a small GTPase, acts as a central molecular switch in determining pollen tube tip growth by orchestrating downstream events such as actin filament (F-actin) dynamics, Ca2+ oscillation, and vesicle exocytosis and endocytosis[10]. The regulatory mechanism of RHO small GTPase, directly determined by RhoGEF (guanine nucleotide exchange factors), RhoGDI (guanine dissociation inhibitors), and RhoGAP (GTPase activating proteins), is highly conserved not only in tip-growing pollen tubes but also in other polarized cells of plants and animals[11,12]. Furthermore, ROP family members in plants have been reported as rheostats in response to various abiotic and biotic stresses, such as hypoxia, salt, and pathogen[13−16]. For pollen tube tip growth, vesicular distrubited REN1(ROP1 enhancer 1) as a RhoGAP protein, which relies on the actin mediated exocytosis, plays a critical role in globally inhibiting ROP1 activity on apical PM of pollen tube by stimulating GTP hydrolysis[17]. RhoGDI2a, on the other hand, acts as the major RhoGDI responsible for maintaining the polarized distribution of active ROP1 in apical plasma membrane (PM) by removing active ROP1 from PM. In Arabidopsis, the pollen tube of ren1 and gdi2a exhibited depolarized growth due to the lateral propagation of active ROP1. In 2018, JGB was identified as REN4 (ROP1 enhancer 4), dynamically localized to the apical and subapical PM of pollen tubes, to maintain the polarized distribution of active ROP1 in the apex PM of pollen tube through endocytosis[18]. Hence, the above studies raises the question of whether ROP1 is involved in pollen dormancy and germination in response to humidity.
Here, we investigated the role of ROP1 in pollen dormancy and germination response to humidity. In ROP1-OE, ren1-3, and gdi2-ami, we found that pollen germination was promoted in vitro or inside anther under high humidity. Conversely, the inhibition of ROP1 activity by overexpression of RhoGAP REN1 and RhoGDI2 prolonged pollen dormancy. Further observation revealed that a close association between the polarization distribution and accumulation of active ROP1 and the germination site of the pollen tube. In conclusion, our work provided genetic evidence that ROP1 pathway is involved in pollen dormancy, and manipulation of ROP1 activity holds potential for preserving pollen as genetic germplasm resource.
-
Arabidopsis thaliana ecotype Columbia (Col-0) was used as the wild type plant material. The seeds were surface-sterilized with 75% ethanol for 15 min, rinsed four-five times with sterile water, and germinated for 5 d on half-strength Murashige and Skoog medium plates with 0.1% MES, 1.2% (for vertical growth or 0.8% (for horizontal growth) agar (pH 5.8). The seeds were vernalized in darkness at 4 °C for 2 d before being transferred to a greenhouse kept at 22 °C with 65% humidity and a 16-h-light/8-h-dark photoperiod.
Plasmid construction
-
The genomic DNA of GDI2 containing 1315 bp promoter and 1410 bp genic DNA was amplified using with Phanta®Max Super-Fidelity DNA Polymerase (Vazyme) and specific primers GDI2-207F/GDI2-207R. The 2.4 kb promoter and full length CDS of REN1 were amplified using REN1-pro-F/REN1-pro-R and REN1-CDS-F/ REN1-CDS-R respectively, then ligated with GFP for pREN1-GFP-REN1. GDI2 amiRNAs were designed according to the WMD (
http://wmd3.weigelworld.org/cgi-bin/webapp.cgi ) procedure[19]. Six primers, pRS300-A, pRS300-B, GDI2 I miR-s, GDI2 II miR-a, GDI2III miR*s, and GDI2 IV miR*a were used to amplify specific fragments from the template of pRS300 vector (gift from the Detlef Weigel group). These amplified fragments were respectively cloned into the vector of pDONR™207 (Invitrogen) using BP Clonase™II Enzyme Mix (Invitrogen, catalog number: 11789020). Then, GDI2 gDNA was shuttled into the gateway destination vectors PGWB653 and PGWB4 respectively, pREN1-GFP-REN1 fragment was shuttled into the modified gateway destination vectors pCAMBIA3300:GW, gdi2-Ami fragment was shuttled into the modified gateway destination vector pCAMBIA3300:Lat52 pro:GFPNOS:Lat52:GW using LR Clonase II enzyme mix (Invitrogen, catalog number: 11791019)[18]. The primers used here are listed in Supplemental Table S1.Pollen tube germination assay on medium
-
In vitro pollen tube germination was performed according to previously published methods, with slight modification[17]. Briefly, mature Arabidopsis pollen grains were isolated from newly opened flowers and placed onto pollen germination medium (18% sucrose, 0.01% boric acid, 1 mM CaCl2, 1 mM Ca (NO3)2, 1 mM MgSO4, pH 6.4 and 0.5% agar). The plates were incubated at 28 °C for 1−2 h. To determine the pollen germination percentage, pollen grains and pollen tubes were observed under an inverted microscope, and digital images were taken by the use of a camera (Nikon). To calculate germination percentage, a minimum of 500 pollen grains were counted in each experiment. At least three experiments were conducted. The pollen germination percentage of each sample was measured using Image J software.
RNA extraction and qRT-PCR
-
Total RNA was extracted from mature pollens of Arabidopsis opening flowers using TRIzol reagent (Invitrogen). Reverse transcription and cDNA synthesis were performed using oligo primers and PrimeScript RT Enzyme Mix I (TaKaRa) from 1 µg of total RNA. Quantitative PCR was performed using an SYBR Premix Ex-Taq (TaKaRa). The PCR program was as follows: 95 °C for 15 s, followed by 45 cycles of 95 °C for 5 s, 55 °C for 20 s, and 72 °C for 30 s. The expression data were normalized using Actin2. The PCR primers used for real-time PCR are shown in Supplemental Table S1.
Pollen tube germination assay in response to humidity
-
Four-week old Arabidopsis with opening flowers were used for humidity treatment and phenotypic analyses as described by Ju et al.[8] and Miao et al.[7]. A plant growth box with a cover was used to generate a humid environment for Arabidopsis flowers. A subset of plants was grown under controlled conditions at 65% relative humidity, while another subset of plants was transferred to a growth box with 85% relative humidity via periodically spraying water[7]. After treatment of 3−4 d, the phenotypic was analyzed using microscopy (Nikon-SMA800).
Scanning electron microscopy observation
-
Scanning electron microscopy observation was performed as described by Li et al.[20]. Dehiscent anthers of plants under normal and high humidity conditions were harvested and fixed by FAA, followed by dehydration with a graded ethanol series. Ethanol gradients were set to 35%, 50%, 75%, 85%, 95%, and 100% and treated for 10 min each. Specimens were dried at crucial points and coated with gold before examination with a Scanning Electron Microscope. The germination of pollen in the anther and stigma was observed under scanning electron microscope (Zeiss Merlin Compact).
Pollen tube staining with Aniline Blue
-
The dehiscing anther of the opening flower was stained in the 108 mM K3PO4 solution with 0.1% decolorized Aniline Blue (DAB), pH = 11.0, for 2 h, then observed using a microscope with ultraviolet light (Nikon-SMA800).
Confocal microscopy observation
-
To visualize the distribution of active ROP1, mature pollen grains of the CRIB4-GFP marker line isolated from dehiscing anther or germinated pollens of CRIB4-GFP after 1 hour cultivation on pollen germination medium were observed using confocal microscope LEICA TCS SR5 (Zeiss), GFP signals were analyzed with the excitation and emission wavelengths set to 488 nm/505 to 550 nm, respectively.
-
Given REN4/JGB participates in pollen dormancy under high humidity conditions in Arabidopsis[8,18], we proposed that REN4-mediated ROP1 was involved in pollen dormancy and germination. To test our hypothesis, we first observed the in vitro pollen germination of ROP1 overexpression line (Lat52p:GFP-ROP1, hereafter ROP1-OE)[21], and found the pollen germination ratio of ROP1-OE showed 55.06% ± 5.20% compared to the 20.75% ± 2.21% germination ratio of wild type after 1 h cultivation (Fig. 1a & b). To further investigate whether ROP1-OE mediated pollen germination is a response to humidity, pollen dormancy state was compared between wild type and ROP1-OE under standard (65%) and high humidity (85%) conditions. Under 65% relative humidity, most pollens inside the dehiscing anther remained dormant and ungerminated in both the wild type and ROP1-OE (Fig. 1c & Supplemental Fig. S1). However, with the humidity increasing via periodically spraying water, ROP1-OE showed precious pollen germination within the anther, as revealed by aniline blue staining (Fig.1c). The situation of pollen germination within opening flower was further observed via high-resolution SEM (Scanning electron microscopy) analysis. Under high humidity conditions, many germinated pollen tubes were found both within the anther and on the pistil surface of ROP1-OE, while only very few pollen tubes germinated in the wild type anthers (Fig. 1d). The above results suggested that ROP1 overexpression accelerated the release of pollen dormancy under moist conditions in Arabidopsis.
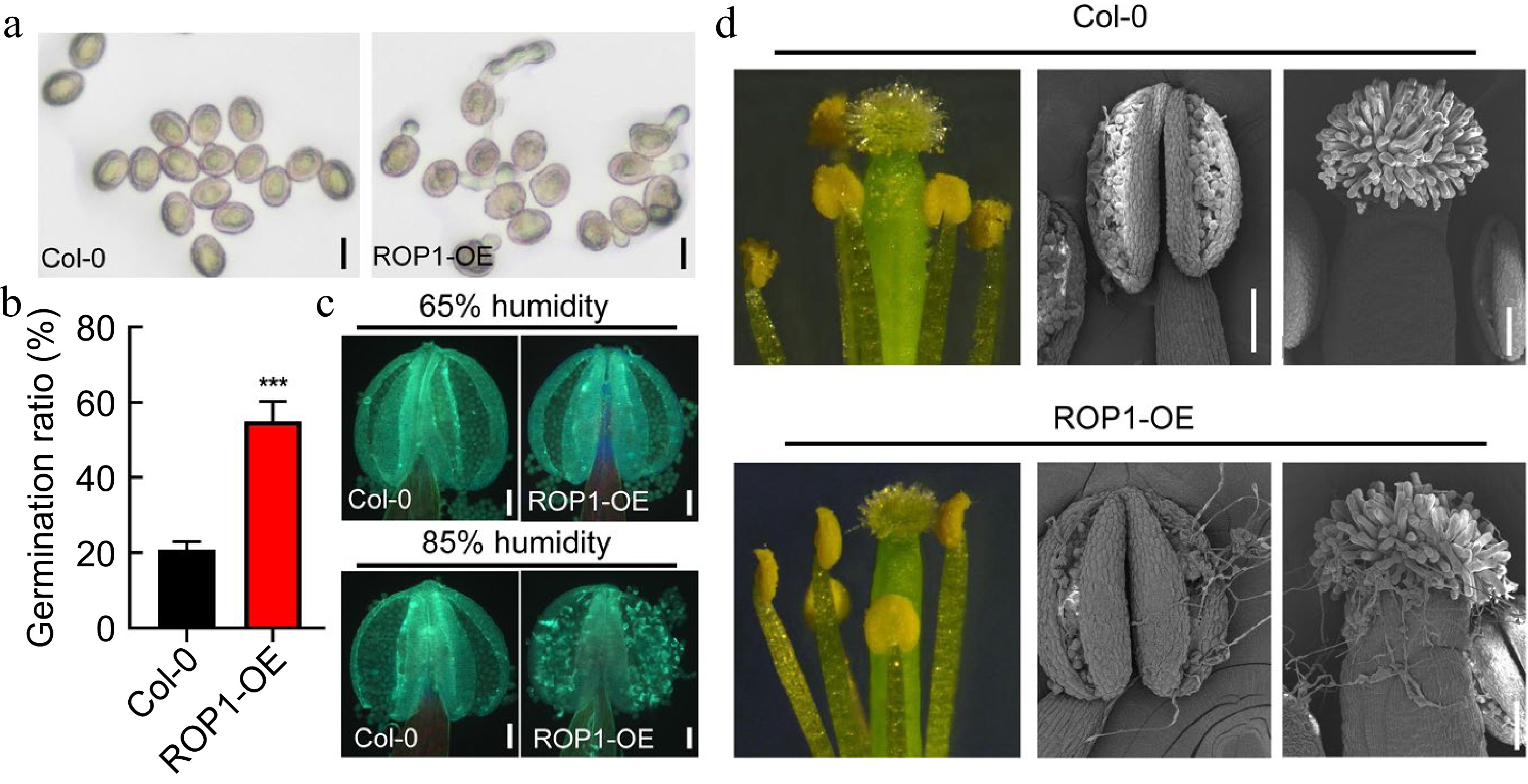
Figure 1.
ROP1 overexpression disrupts dormancy of pollen inside anther under moist environments. (a), (b) Images and germination ratio of Col-0 and ROP1-OE pollen 1 after imbibition. Scale bars = 20 μm. Error bars represent SD. Statistical significance was determined by Student's t-test (*** p < 0.001, n = 3). (c) Fluorescence images of Col-0 and ROP1-OE anther with or without pollen tubes stained with aniline blue. Scale bars = 100 μm. (d) Image of Col-0 and ROP1-OE anther and stigma under high humidity conditions. The red arrow points to the pollen tube. Scale bars = 500 μm in the left images. Scale bars = 100 μm in the images of the middle and right.
Active ROP1 marks the germination site of the pollen tube
-
Based on the presence of two ROP1 forms: inactive GDP-bound and active GTP-bound forms, we next sought to understand whether active ROP1 plays a critical role in pollen dormancy and germination. ROP1 effector CRIB4 was reported as a visualized marker of active ROP1 in pollen tube[22], and the subcellular distribution of active ROP1 was investigated in mature dormant pollen and germinating pollen using CRIB4-GFP marker line. In mature dormant pollen within the dehiscing anther, CRIB4-GFP mainly distributed in the pollen cytoplasm (Fig. 2a). After incubation on the germination medium for 1 h, CRIB4-GFP signaling was further observed in pollens with different germinated and elongated states. In the pollen without visible signs of a germination site, strong GFP signals shifted to a restricted PM region and appeared with asymmetrically polarized distribution (Fig. 2b). Then, the polarized distribution of CRIB4-GFP at the PM became more pronounced at the germination site and was maintained at the apical PM of emerging pollen tubes (Fig. 2c−f). The observations indicated that ROP1 activation and polarized recruitment in PM is closely associated with the initiation of pollen germination.
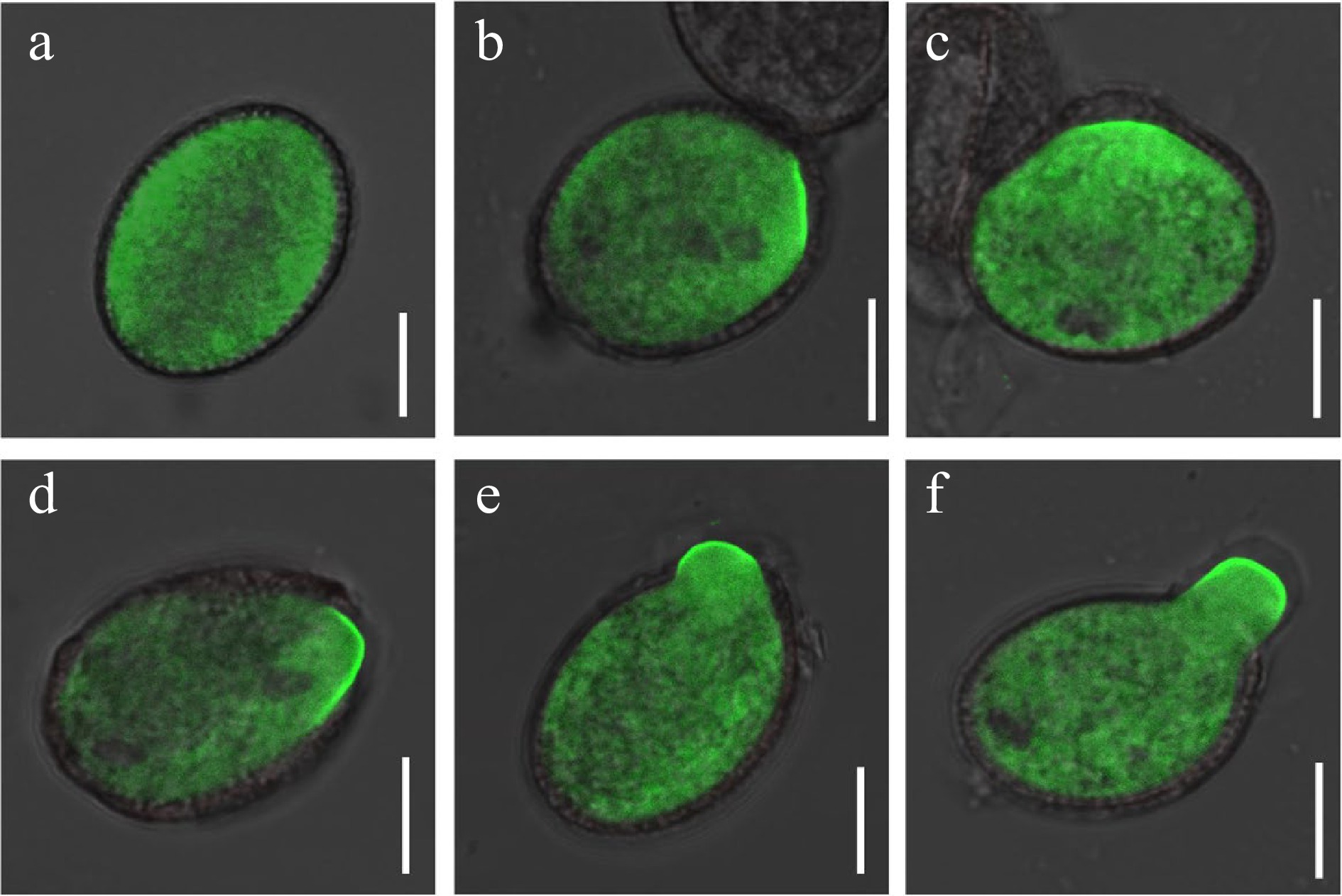
Figure 2.
Active ROP1 localization in the dormant pollen and germinated pollen of Arabidopsis. (a) CRIB4- GFP distribution in the dormancy mature pollen grain. (b)−(d) CRIB4-GFP polar distribution in the germinated site of pollen during germination. (e), (f) CRIB4-GFP polar distribution in the germinated and elongated pollen tube. Scale bars = 10 μm.
ROP1 activation releases pollen dormancy
-
The role of active ROP1 in pollen germination was further analyzed using the ren1-3 mutant (knockdown of RhoGAP EN1)[17]. In vitro pollen germination assay showed that the ren1-3 pollen exhibited a significantly higher germination ratio (55.17% ± 6.78%) compared to the wild type (8.86% ± 4.68%) after 1 h incubation on medium (Fig. 3a, b). Additionally, under 65% humidity conditions, only a few elongated pollen tubes were observed within the ren1-3 anther, while pollen tube germination from the dehiscing ren1-3 anther was dramatically accelerated, and pollen tubes attached to the stigma under high humidity conditions (Fig. 3c, d). This suggested the suppression of ROP1 activity by REN1 is required for maintaining pollen dormancy in Arabidopsis.
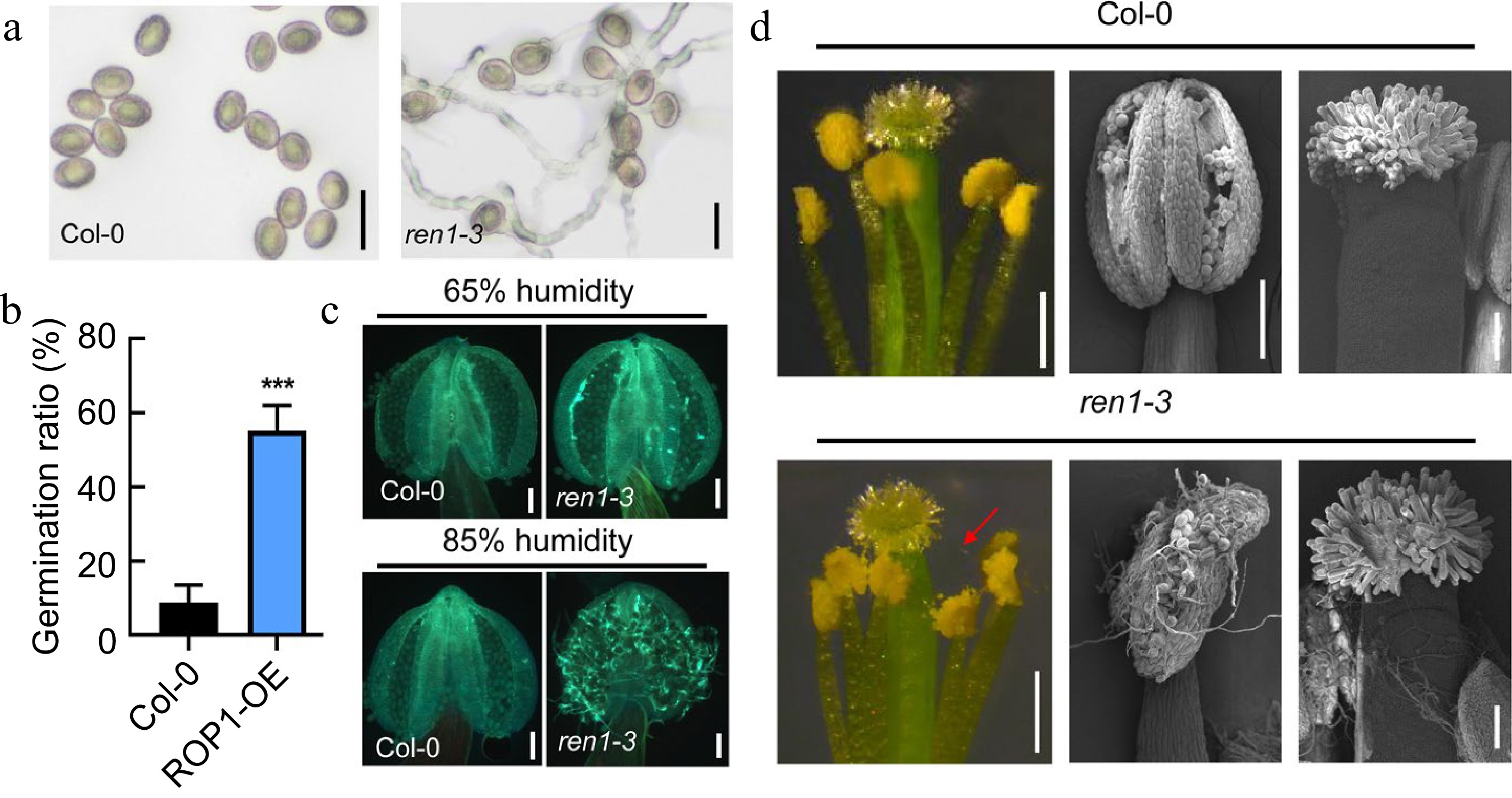
Figure 3.
Pollen dormancy was interrupted in the ren1 mutant under moist environments. (a), (b) Images and germination ratio of Col-0 and ren1-3 pollen 1 h after imbibition. Scale bars = 20 μm. Error bars represent SD. Statistical significance was determined by Student's t-test (*** p < 0.001, n = 3). (c) Fluorescence images of Col-0 and ren1-3 anther with or without pollen tubes stained with aniline blue. Scale bars = 100 μm. (d) Image of Col-0 and ren1-3 anther and stigma under high humidity conditions. The red arrow points to the pollen tube. Scale bars = 500 μm in the left images. Scale bars = 100 μm in the images of the middle and right.
PM localized-active ROP1 favors humidity-induced pollen germination
-
Active ROP1 is distributed on the apex PM of pollen tube, and loss function of RhoGDI caused accumulation of active ROP1 in PM distribution[35]. We hypothesized that the distribution of active ROP1 determined its role in pollen germination. To test our hypothesis, two knockdown lines (gdi2 ami) of GDI2 were investigated (Supplemental Fig. S2). After 1 h of in vitro cultivation, compared to 20.75% ± 2.2% pollen germination of wild type, the pollen germination ratio of gdi2-ami2 and gdi2-ami8 showed 56.45% ± 2.18% and 50.98% ± 6.34% respectively (Fig. 4a). The pollen germination inside anther of wild type and gdi2 ami were further compared. Although there is no difference in pollen dormancy between wild type and gdi2 ami at standard humidity (Supplemental Fig. S1), with the increase of humidity, pollen tubes of gdi2 ami germinated prematurely within the anther and were randomly distributed on the pistil surface, while almost no pollen germination was observed within the wild type anther (Fig. 4c & d). These suggested that subcellular distribution of active ROP1 is critical for pollen dormancy and surviving inside the anther under unfavorable humidity conditions and ensures pollen germination and tube growth within the pistil after contact with the stigma.
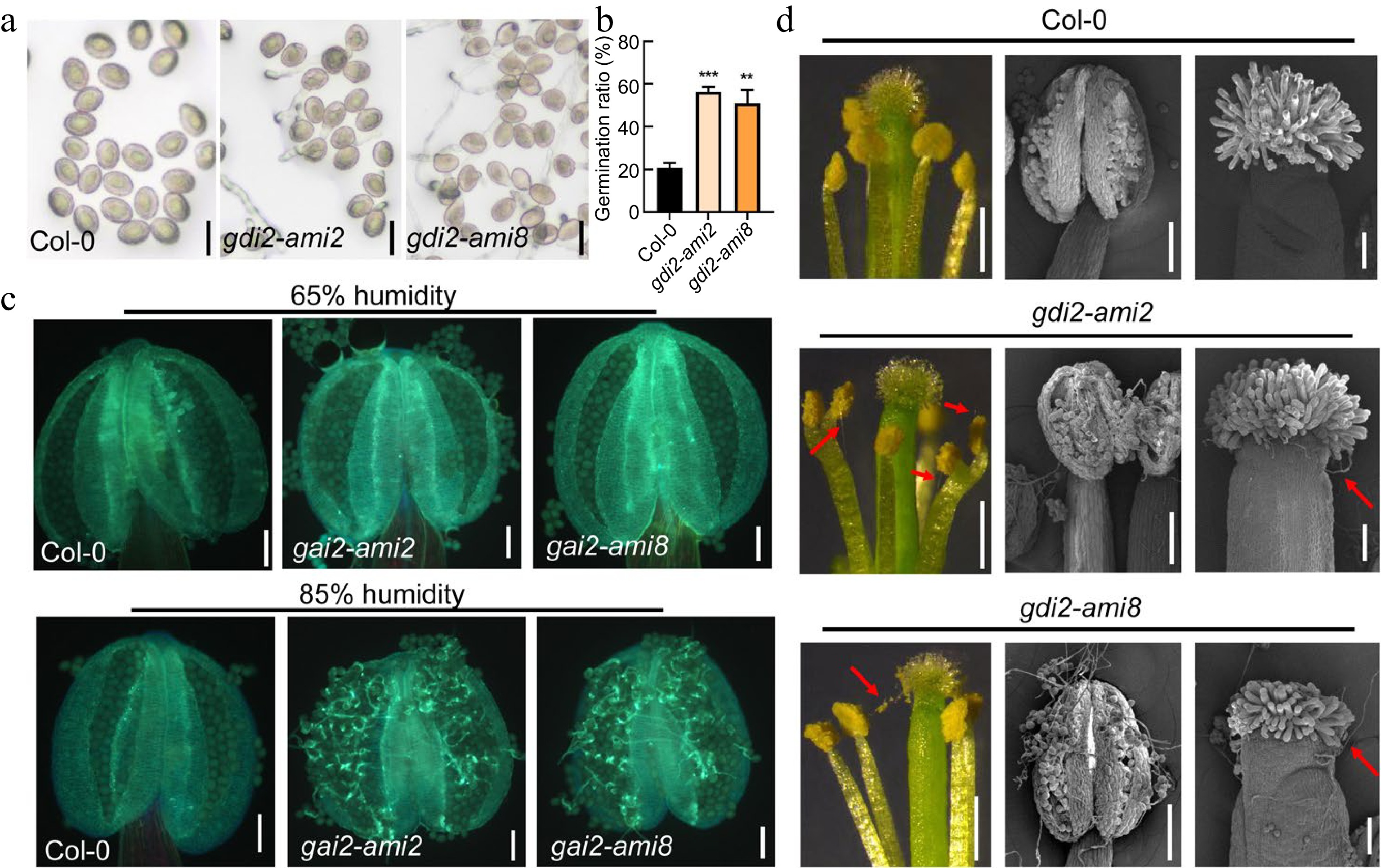
Figure 4.
Pollen dormancy was interrupted by the knockdown of GDI2 under moist environments. (a), (b) Images and germination ratio of Col-0, gdi2-ami2, and gdi2-ami8 pollen 1 hour after imbibition. Scale bars = 20 μm. Error bars represent SD. Statistical significance was determined by Student's t-test (*** p < 0.001, n = 3). (c) Fluorescence images of Col-0 and gdi2-ami anther with or without pollen tubes stained with aniline blue. Scale bars = 100 μm. (d) Image of Col-0, gdi2-ami2 and gdi2-ami8 anther and stigma under high humidity conditions. The red arrow points to the pollen tube. Scale bars = 500 μm in the left images. Scale bars = 100 μm in the images of middle and right.
Suppression of ROP1 activity reinforces pollen dormancy
-
Three pollen-specific expressed ROPs (ROP1, ROP3, and ROP5) in Arabidopsis were previously suspected to be functionally redundant in pollen tube growth[23], and Xiang et al. reported that the mutation combination of ROP1, ROP3, ROP5, and ROP9 in Arabidopsis caused inhibition of pollen tube germination in vitro to some extent[24]. To further confirm the role of active ROP1 in pollen dormancy and germination, the transgenic lines overexpressing GFP-REN1 or GDI2-RFP fused protein driven by their native promoter were generated. We first examined pollen viability before pollen germination assay, and found that pollen viability of REN1 and GDI2 overexpressing lines is similar to that of wild type (Supplemental Fig. S3). Three lines with different REN1 expression levels in wild type background were selected for pollen tube germination assay (Fig. 5). After 3 h cultivation, we observed that GFP-REN1 overexpression significantly inhibited pollen germination, as evidenced by the high ungerminated ratio (98.99% ± 0.32%, 98.9% ± 0.27%, and 97.39% ± 0.61%) compared to the wild type (54.48% ± 5.21%, 53.81% ± 6.09%, and 62.31% ± 6.11%) (Fig. 5a−d).
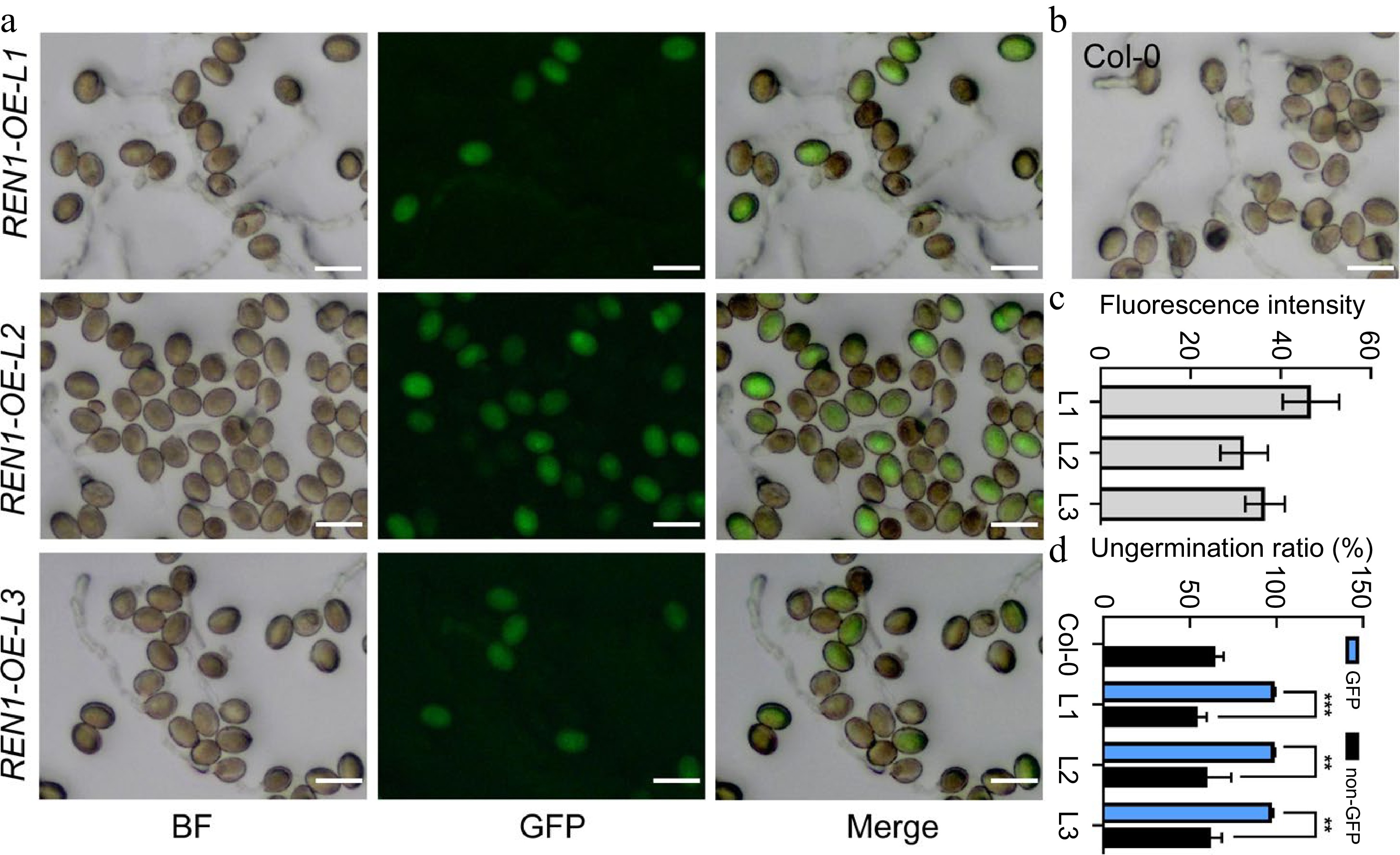
Figure 5.
Inhibition of pollen germination by REN1 overexpression. (a), (b) In vitro pollen germination of Col-0 and three REN1-OE transgenic lines (pREN1: GFP-REN1 CDS) after 3 h cultivation on PGM. Scale bars = 50 μm. (c) Fluorescence intensity assay of GFP-REN1 expressed pollens from three REN1-OE. (d) Germination ratio of Col-0 and REN1-OE pollens after 3 h cultivation on PGM. Statistical significance was determined by Student's t-test (** p < 0.01; *** p < 0.001), error bars represent SD, n = 3.
Meanwhile, the pollen tube germinated ratio of three GDI2 overexpression lines with different fluorescence intensities of 132.44 ± 8.20, and 112.93 ± 4.95, 48.74 ± 8.28 were further analyzed (Fig. 6). Similarly, after 3 h cultivation, we observed that pollen germination was also obviously inhibited by GDI2 expression (Fig. 6a−d), and the ungerminated ratio of pollens in three GDI2 overexpression lines were 99.42% ± 0.41%, 99.36% ± 0.47%, and 98.73% ± 0.34% respectively. Comparatively, the ungerminated ratio of pollens without RFP fluorescence was 49.27% ± 4.0%, 71.91% ± 5.90%, and 55.62% ± 8.81% respectively (Fig. 6d). Although GDI2-OE3 showed much less expression than the other two lines, they showed similar inhibition on pollen germination, this indicated that GDI2 is highly functional for pollen dormancy in Arabidopsis. Pollen germination inhibition by REN1&GDI2 overexpression is consistent with REN4/JGB overexpression[8,18]. These results confirm that ROP1 activity modulation is critical for the maintenance of pollen dormancy before attaching to the stigma in Arabidopsis.
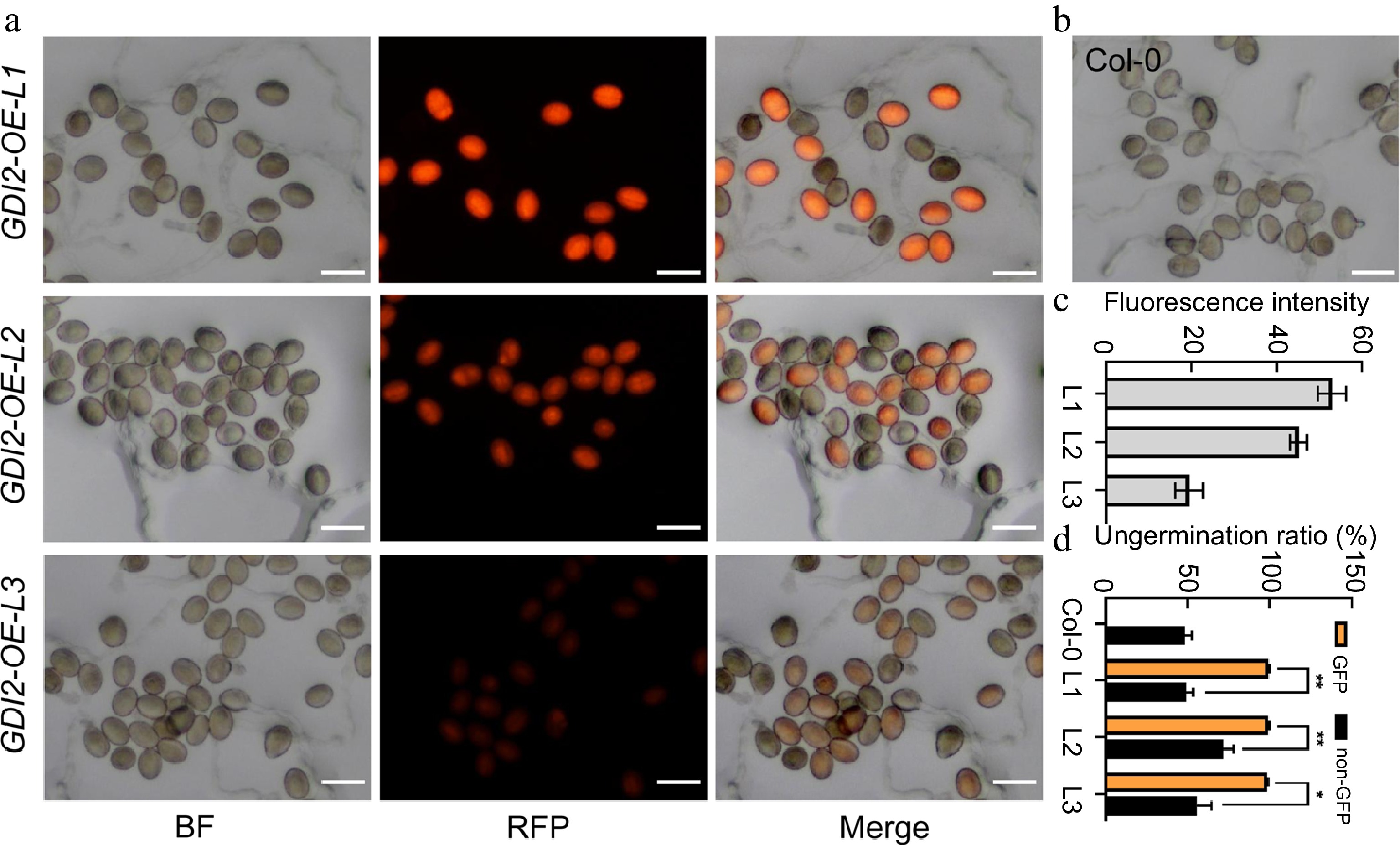
Figure 6.
Inhibition of pollen germination by GDI2 overexpression. (a), (b) In vitro pollen germination of Col-0 and three GDI2 OE transgenic lines (pGDI2:GDI2-RFP) after 3 h cultivation on PGM. Scale bars = 50 μm. (c) Fluorescence intensity assay of GDI2-RFP expressed pollens from three GDI2 OE transgenic lines. (d) Germination ratio of Col-0 and GDI2-OE pollens after 3 h cultivation on PGM. Statistical significance was determined by Student's t-test (* p < 0.05; ** p < 0.01; *** p < 0.001), error bars represent SD, n = 3.
Inhibition of ROP1 activity doesn't affect pollen germination on the stigma
-
To illustrate the specific role of ROP1 activity in detached pollen dormancy, in vivo pollen germination on stigma and seed development were examined in REN1-GFP and GDI2-GFP overexpressed lines. The mature pollens of heterozygote REN1- and GDI2-GFP overexpressed line were pollinated into stigmas of emasculated wild type flowers, as shown in Fig. 7a, b, the GFP-labeled pollen tube overexpressing REN1 and GDI2 were observed on the stigma at 1 h after pollination. Furthermore, the subsequent fertilization and seed development in their homozygous transformants proceeded normally (Fig. 7c, d & Supplemental Fig. S4). This indicates that pollen dormancy state by inhibiting ROP1 activity can be successfully overcame upon receiving signal from female stigma. Hence, this trait offers potential for pollen germplasm preservation and facilitating plant breeding.
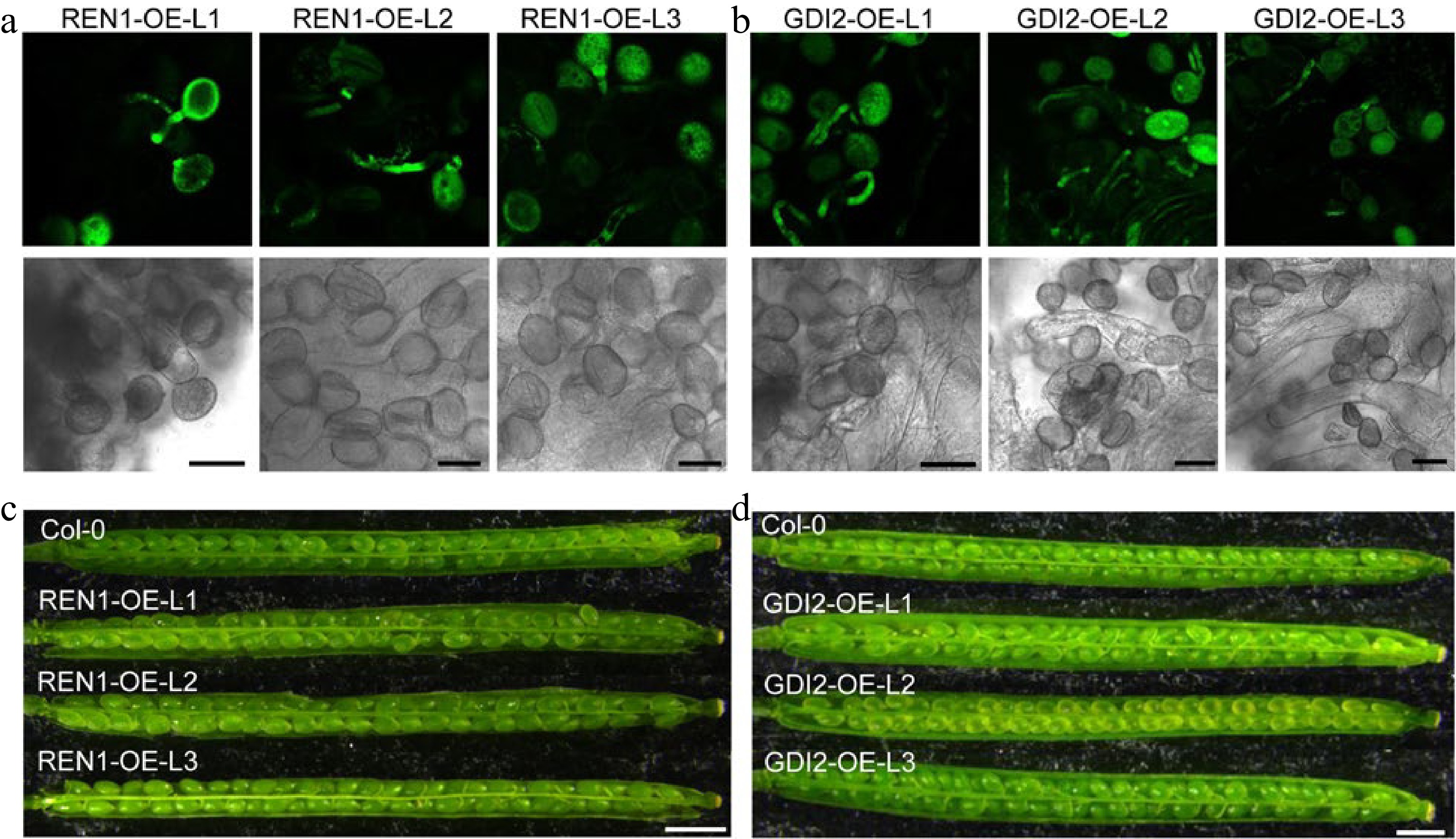
Figure 7.
Pollen germination on stigma and silique of REN1 or GDI2 overexpression lines. (a) Germination of GFP labeled REN1-overexpressed pollens on Arabidopsis stigma. Scale bars = 25 μm. (b) Germination of GFP labeled GDI2-overexpressed pollens on Arabidopsis stigma. Scale bars = 25 μm. (c) The developed silique of self-fertilized wild type and REN1-overexpressed lines. Scale bars = 2 mm. (d) The developed silique of self fertilized wild type and GDI2-overexpressed lines. Scale bars = 2 mm.
-
In this study, we investigated the role of active ROP1 in pollen dormancy and germination response to environmental factors. Our results provide evidence that active ROP1 is involved in disrupting pollen dormancy and promoting germination under high humidity conditions (Figs 1, 3, 4 & Supplemental Fig. S1). ROP1 overexpression accelerated pollen germination (Fig. 1), while inhibition of ROP1 activity through overexpression of REN1 or GDI2 prolonged pollen dormancy (Figs 5 & 6). Furthermore, the distribution of active ROP1 was closely associated with the germination site of pollen tubes (Fig. 2), indicating its importance in the initiation of pollen germination.
The findings of this study contribute to our understanding of the regulatory mechanisms underlying pollen dormancy and germination. Previous studies have focused on the role of ROP1 in pollen tube growth and polarized tip growth[17,18,21,25−35], but its involvement in pollen dormancy has been less explored. Our results highlight the dual role of ROP1, where its activation promotes pollen germination, while its inhibition maintains dormancy. The activity balance of ROP1 is crucial for the pollen survival and successful reproduction under varying environmental conditions.
It is worth noting that although the in vitro pollen germination was inhibited by overexpression of RhoGAP REN1, GDI2, and JGB/REN4 (Figs 5 & 6), the germination on the stigma and subsequent fertilization were not affected, as evidenced by the normal seed development (Fig. 7). This suggests that the inhibition of ROP1 activity specifically regulates pollen dormancy during the dispersal phase, ensuring pollen survival in moist conditions. The redundancy of multiple pollen-expressed ROPs may provide a protective mechanism to ensure pollen germination on the stigma and guarantee successful sexual reproduction[24].
The role of humidity in modulating ROP1 activity and pollen germination is still not fully understood, as our analysis did not reveal significant changes in the expression levels of ROP1 and its upstream regulators in response to high humidity (Supplemental Fig. S5). This suggests that humidity may exert its effect on ROP1 activity through post-transcriptional or post-translational mechanisms. Notably, previous research has shown that exogenous application of MeJA or Ca2+ effectively disrupted pollen dormancy and stimulated pollen germination in Arabidopsis[6,8]. Additionally, the depolarized pollen tube resulting from the overexpression of calcium depended proteins kinase CPK32 mimicked the pollen tube defective induced by expressing constitutively activated ROP1[36]. Furthermore, JGB/REN4 has been found to inhibit JA biosynthesis and concurrently curtail ROP1 activity in pollen. Does high humidity activate ROP1 through MeJA or Ca2+ to disrupt pollen dormancy? Further investigation is needed to elucidate the precise molecular mechanisms by which humidity regulates ROP1 activity in pollen germination.
-
In conclusion, our study sheds light on the role of ROP1 in pollen dormancy and germination, emphasizing the importance of fine-tuning ROP1 activity to balance dormancy and germination in response to environmental cues. Manipulation of ROP1 activity has the potential to enhance pollen germplasm preservation and facilitate the breeding of new crop and fruit varieties. Furthermore, our findings may provide insights into preventing pre-harvest sprouting or vivipary in crop seeds.
-
The authors confirm contribution to the paper as follows: research design: Li H; research assistant: Zhou X, Liu Z; Experiment performance: Yan J; data analysis: Li H, Hu J; manuscript preparation: Li H, Gong J; manuscript review: all authors. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
We thank ZP Zhang and WJ Cai of Core facility for technical support with microscopy work in the Center for Excellence in Molecular Plant Sciences, Chinese Academy of Sciences. This work was supported by grants from the National Natural Science Foundation of China (31970744) and the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB27020101).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primer Sequences Used in the Gene Cloning.
- Supplemental Fig. S1 Image of Col-0, ROP1-OE, ren1-3, gdi2-ami2 and gdi2-ami8 flower (a), anther (b) and stigma (c) under 65% relative humidity conditions. Scale bars = 500 μm in the images of the first row, and = 100 μm in the SEM images of the 2nd and 3rd rows.
- Supplemental Fig. S2 The relative expression level of GDI2 in the opening flower of wild type and two gdi2-ami homozygous lines. The opening flowers were collected for qRT-PCR analysis of gene expression. Error bars represent SD from 4 biological replicates. Statistical significance was determined by Student’s t-test (*p < 0.05), n = 4.
- Supplemental Fig. S3 Pollen viability assay of REN1 overexpressed lines (a) and GDI2 overexpressed lines (b). Anthers were dissected from buds just before opening, stained with Alexander’s staining, mounted on glass slides and observed under a light microscope. Viable pollen is colored in red, scale bar = 100 μm.
- Supplemental Fig. S4 Resistance assay of transgenic Arabidopsis lines overexpressing REN1 (a) and GDI2 (b). Seedling resistance of Col-0 and REN1-OE homozygotes on selection 1/2 MS plate supplemented with 10 mg/L phosphinothricin. Seedling resistance of Col-0 and GDI2-OE homozygotes on selection 1/2 MS plate supplemented with 50 mg/L Kanamycin.
- Supplemental Fig. S5 The expression level of ROP1 and its regulators under different humidity conditions. The effect of high humidity treatment 1 day (a) and 4 days (b) on the expression of ROP1, REN1, GDI2, REN4, and GEF12, respectively. Error bars represent SD. Statistical significance was determined by Student’s t-test (*p < 0.05), n = 3.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yan J, Zhou X, Liu Z, Hu J, Li H, et al. 2023. Inhibition of ROP1 activity is essential for pollen dormancy under moist conditions in Arabidopsis. Seed Biology 2:18 doi: 10.48130/SeedBio-2023-0018 

Inhibition of ROP1 activity is essential for pollen dormancy under moist conditions in Arabidopsis
- Received: 24 July 2023
- Accepted: 20 October 2023
- Published online: 07 November 2023
Abstract: Pollen dormancy is a vital trait for plant breeding, enabling the use of pollen as a valuable germplasm resource. However, pollen dormancy can be easily disrupted by humidity and the underlying regulatory mechanisms remain poorly understood. Here we report a novel finding that highlights the involvement of ROP1 in the termination of pollen dormancy triggered by humidity. ROP1 overexpression significantly promoted in vitro pollen germination, and shortened pollen dormancy period under high humidity conditions, leading to premature pollen germination within the anthers. Consistently, the knockdown of Rho GAP-REN1 and Rho GDI2 caused premature pollen germination on medium, and the additive effect on pollen germination was further observed when subjected to increased humidity in the ren1 and gdi2 mutants. Conversely, inhibition of ROP1 activity through the expression of Rho GAP-REN1 and Rho GDI2 suppressed in vitro pollen germination. In summary, our work demonstrates that the suppression of ROP1 activity is crucial for maintaining pollen dormancy, will contribute to facilitating the preservation of pollen as a valuable genetic germplasm resource and advancing development of plant breeding.
-
Key words:
- Pollen dormancy /
- Humidity /
- ROP1 GTPase /
- Germplasm resource /
- Pollen preservation