










-
Tea plant (Camellia sinensis (L) O. Kuntze) is an important leaf economic crop widely cultivated worldwide[1]. Chlorophyll content as a useful agronomic trait, plays a crucial role in tea plant cultivation and breeding[2,3]. Different varieties of tea plants have leaf colors which are often perceived as more visually appealing, and have a higher price due to their delicate appearance, high content of theanine, as well as umami flavor[4]. In recent years, more and more new tea varieties with albino or etiolation phenotypes have been cultivated, including 'Baiye 1 (Anji Baicha)', 'Baicha 1', 'Huangjinya', and 'Zhonghuang 1', etc[5,6]. In general, albino or etiolation buds are known to possess high levels of amino acids, which are essential in enhancing the natural flavor and improving the quality of tea[7,8]. At present, cultivated albino or etiolation tea plants can be categorized into temperature-sensitive, light-sensitive and environmental-insensitive groups according to their response to the environment[9,10]. 'Baiye 1' is a cultivar of temperature-sensitive albino tea, which displays yellow shoots at low temperatures and transitions to green shoots upon exposure to warm temperatures[11]. Previous studies demonstrated that chlorophyll deficiency in albino or etiolation tea plants may be closely related to differential gene expression in the pathways of both chlorophyll biosynthesis and degradation[12].
Chlorophyll is a crucial pigment that plays the central role in light-harvesting during photosynthesis, and facilitates the transfer of electrons to the photoreaction center[13,14]. Typically, chlorophyll biosynthesis involves three primary stages: (i) the production of 5-aminolevulinic acid (ALA), (ii) the formation of protoporphyrin IX from eight molecules of ALA, and (iii) the synthesis of chlorophyll through the Mg-protoporphyrin pathway[15,16]. The first step in chlorophyll biosynthesis is directed by the Mg-branch of the tetrapyrrole pathway, which involves the ATP-dependent insertion of Mg2+ into protoporphyrin IX, performed by the magnesium chelatase enzyme complex. The magnesium chelatase enzyme complex is composed of three subunits: CHLI, CHLD, and CHLH[17,18]. The CHLI subunit is primarily responsible for the hydrolysis of ATP during enzymatic reactions within the chlorophyll biosynthetic pathway[19]. It is equipped with specific structural motifs required for ATP binding, such as Walker A and B (W-A and W-B) motifs, sensors 1 and 2 (S-1 and S-2) motifs, presensor I and II (PS-I and II) inserts, as well as an arginine finger (ARG-finger) motif. PsCHLI1 has been identified by Wu et al.[20] as the main CHLI subunit, shown to have a crucial role in sustaining Mg2+-chelatase activity as well as enhancing the pea's photosynthetic efficiency. Interestingly, the CHLD subunit exhibits an AAA+ module at its N-terminus, which shares significant structural similarity to CHLI. Research has revealed that comparison of Rhodobacter capsulatus subunit CHLI and the AAA+-like module of subunit CHLD (residues 1−238 aa) shows 25% identity for the N-terminal AAA+ core[21]. The CHLD subunit possesses an elongated linker region abundant in acidic residues, as well as a C-terminal integrin I domain (also referred to as von Willebrand factor A or VWFA). In addition, CHLH is a multifunctional protein localized in the plastid that acts as a crucial gene related to leaf pigmentation by regulating chlorophyll biosynthesis and chloroplast ultrastructure. Regarding Mg2+-chelatase, the ATPase function is carried out by the AAA-type CHLI subunit, while the binding of protoporphyrin IX (Proto IX) and metal insertion take place on CHLH[22].
A number of mutations of the Mg2+-chelatase subunit have been reported in chlorophyll-deficient mutants in Arabidopsis, rice and maize[23−25]. However, the interaction mechanism between subunits of Mg2+-chelatase and the function of Mg2+-chelatase in chlorophyll biosynthesis and metabolism in albino tea plants has not yet been investigated. In this study, the relationship between gene expression of CsCHLI, CsCHLD and CsCHLH of Mg2+ chelating enzyme and the chlorophyll content in tea leaves were characterized. Then, the CsCHLI gene was cloned, the subcellular localization and the mechanism in chlorophyll synthesis were analyzed by yeast two-hybrid (Y2H) and bimolecular fluorescence complementation (BiFC). Moreover, transgenic Arabidopsis was used to validate the role of CsCHLI in chlorophyll. This study provides in-depth insights into the interaction mechanism of Mg2+ chelating enzyme, as well as the function of CsCHLI in chlorophyll biosynthesis of tea plant. These results can supply important theoretical information for the albino mechanism, meanwhile serve as valuable industrial support for tea breeding.
-
Two-year-old cuttings of different leaf color varieties, such as, albino tea cultivar, 'Baiye 1' (BY1), two normal green cultivars, 'Shaancha 1' (SC1) and 'Longjing 43' (LJ43), were cultivated in the natural photoperiod greenhouse of Northwest A&F University, Shaanxi, China (70% ± 5% relative humidity, temperature 25 ± 3 °C). A. thaliana Ler-0 (Ler wild type) and AtCHLI1 (At4g18480) mutant atchli1 (Sail_203_D11), Ler wild type background, was provided by Xu Min Lab, Northwest University, Xi'an, Shaanxi, China. A. thaliana were grown in an illuminating incubator, with a temperature of 22 ± 2 °C, with 100 μmol·m−2·s−1 light intensity, and a photoperiod of 16 h of light and 8 h of darkness. Wild-type Nicotiana benthamiana plants were cultured in a growth chamber under 25 ± 2 °C, 16 h light/8 h dark, 100 μmol·m−2·s−1 light intensity, respectively.
Chloroplast ultrastructure observation
-
Chloroplast ultrastructure observations were performed as described previously[26]. The freshly harvested second leaf was cut into 1 mm × 1 mm pieces and subsequently immersed in a 2.5% glutaraldehyde solution (v/v), at 4 °C for 12 h to effectively fix the samples. Dehydration and infiltration were conducted using a gradient mixture of acetone and resin, then the ultrathin sections were observed using an HT-7700 transmission electron microscope (TEM; Hitachi, Japan).
Chlorophyll content determination
-
The leaves of tea and Arabidopsis were isolated using 80% (v/v) ice-cold acetone and homogenization with zirconia beads. The extracts were then centrifuged at 10,000× g for 1 min at 4 °C. The absorbance of the extract solutions were measured using UV-1600 spectrophotometer (Shimadzu, Kyoto, Japan) at 645, and 663 nm, respectively. The content of chlorophyll a and b were calculated as described previously[26]. Three experimental replications were conducted.
RNA extraction and RT-qPCR analysis
-
Total RNA was extracted from tea plant leaves using a Tissue RNA Isolation Kit (Beibei Biotechnology Co., Ltd., Zhengzhou, China) following the protocol. RNA quality and integrity were evaluated by electrophoresis on the 1.2% (w/v) agarose gel. Furthermore, cDNA was obtained using a HiScript II 1st Strand cDNA Synthesis Kit (Vazyme Biotechnology Co., Ltd., Nanjing, China). The sequence of CsCHLI (CSS0004540.1) and CsACTIN (CSS0008920.1) were obtained from the Tea Plant Information Archive (TPIA;
http://tpia.teaplant.org ). The specific primers of RT-qPCR were designed with Primer 7.0 software and listed in Supplemental Table S1. RT-qPCR analysis was conducted with the Bio-Rad Real-time thermal cycler CFX96 with SYBR Premix ExTaq™ Kit (Takara Co. Ltd., Japan). Finally, the 2−ΔΔCᴛ method was used to determine the relative expression levels. Three biological replicates per sample were used.Gene structure and motif analysis
-
The gene structure of CsCHLI, including base pair coordinates, was visualized using TBtools software by integrating the findings from the Pfam search and GFF3 gene structure information. The conserved motifs were analyzed utilizing NCBI Conserved Domains (
www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ). The analysis of conserved domains was conducted on the online MEME software (v. 4.9.1) (Multiple Expectation Maximizations for Motif Elicitation;http://meme-suite.org/tools/meme ). The motifs were carried out in Classic Mode on the online MEME software, with the number parameter set to less than 10 and all other options maintained at default settings. Afterward, the MEME outcomes were downloaded as a MAST file and visualized in TBtools, where the motif positions were shown.Phylogenetic analysis
-
To obtain the amino acid sequences of CsCHLI and ten other homologs, BLASTP was performed on the National Center for Biotechnology Information website (NCBI;
https://blast.ncbi.nlm.nih.gov ). Sequence alignment was accomplished with ClustalW, and the phylogenetic analysis was generated using MEGA-X software by the Neighbor-Joining method with 1000 bootstrap replications. The phylogenetic tree was constructed using Geneious 9.0.Vector construction
-
The sequences of genomic DNA or cDNA of CsCHLI were amplifed from young leaves of 'Shaancha 1'. For subcellular location analysis, the open reading frame (ORF) sequence of CsCHLI without the stop codon was amplified use the primers of 3302Y-CHLI-F and 3302Y-CHLI-R. The target PCR fragment was recovered and combined into the 3302Y vector with enhanced yellow fluorescent protein (EYFP) and 35S promoter to construct the recombinant vectors 35S::CsCHLI-EYFP by the ClonExpress II One Step Cloning Kit (Vazyme Biotech, China). For Arabidopsis transformation, full-length CsCHLI was amplified using the primers of 1300-CHLI-F and 1300-CHLI-R, then the digested fragment with BamHI was recovered and inserted into 1300, resulting in the recombinant plasmids 35S::CsCHLI-GFP. For the Y2H assay, CsCHLI (CSS0004540.1), CsCHLD (CSS0026907.1), and CsCHLH (CSS0016317.1) CDS were cloned by corresponding specific primers, the fragments were recovered and inserted into pGBKT7 (BD) and pGADT7 (AD) vectors by the ClonExpress II One Step Cloning Kit to construct the recombinant plasmids BD-CsCHLI, AD-CsCHLI, BD-CsCHLD, AD-CsCHLD and BD-CsCHLH, respectively. For the BiFC assay, CsCHLI, CsCHLD, and CsCHLH were cloned by corresponding specific primers, the target fragments were recovered and inserted into the pNYFP and pCYFP vectors by the ClonExpress II One Step Cloning Kit to construct the 35S::CsCHLI-YFPC, 35S::CsCHLI-YFPN, 35S::CsCHLD-YFPC, and 35S::CsCHLH-YFPN plasmids, respectively.
Subcellular localization
-
The 35S promoter-driven fusions of CsCHLI-EYFP was transformed into Agrobacterium tumefaciens (strain GV3101). The transient transformation method was employed to inject the cells into the leaves of six-week-old tobacco (N. benthamiana) using a needleless syringe. After 48−72 h, the infiltrated leaves were harvested, and EYFP fluorescence was quantified using a laser scanning confocal microscope (Leica, Wetzlar, Germany).
Yeast-two-hybridization assay (Y2H)
-
The recombinant plasmids were transferred into the yeast strain using Y2H (Weidi Biotechnology Co. Ltd. (Shanghai, China)). The transformed cells were cultivated on SD-Leu-Trp (SD-TL) plates and incubated at 30 °C for 2 d. Following growth of the yeast cells, the putative transformants (OD600 = 0.2) were transferred to selective media (SD/-Leu/-Trp/-His/-Ade/X-α-gal/AbA) (SD-TLHA). Observations on bacterial colony growth were conducted after 3 d.
Bimolecular fluorescence complementation assay (BiFC)
-
BiFC assays were conducted following the previously described methods[27]. The CDSs of CsCHLI, CsCHLD, and CsCHLH were inserted into pSPYNE-35S and pSPYCE-35S, which contain the C-terminal and N-terminal regions of yellow fluorescent protein (YFP), respectively. The two plasmids were co-transformed into A. tumefaciens strain GV3101 cells, then the six-week-old tobacco leaves were transiently infiltrated. After infiltration, plants were grown at 22 °C with 16 h photoperiod before detection. After 2 d, the fluorescence signal in the injected area of the leaves were observed using a confocal laser scanning microscope (Leica, Wetzlar, Germany). YFP was visualized by exciting with an argon laser at 514 nm.
Transgenic Arabidopsis generation
-
The recombinant vector was introduced into A. tumefaciens strain GV3101 using the electroporation transformation method. Using the floral dip method as described previously[28], 35S::CsCHLI-GFP was transformed into Ler-0 wild type (Ler-0) and atchli1 mutant in Ler-0 wild type background, respectively. Transgenic Arabidopsis plants were selected in the T3 transgenic plants for phenotypic identification, chlorophyll content measurement, and gene expression analysis[29]. T1 transgenic plants were identified through the detection of GFP fluorescence as well as PCR amplification. The results were analyzed using DSDecode (
http://dsdecode.scgene.com/home/ ) to decode the sequence and detect mutations.Statistical analyses
-
The determination of chlorophyll and molecular analyses were conducted using at least three independent biological replicates. Statistical analyses were performed using Minitab 17.0 statistical software (Minitab Inc., UK). To detect statistically significant differences between mean values, one-way ANOVA was conducted with Fisher's least significant difference (LSD) test at 5%.
-
To investigate the function of Mg2+-chelatase genes in the chlorophyll biosynthesis of tea leaves. The ultrastructure, chlorophyll content, and the expression levels of the Mg2+-chelatase genes in tea leaves were determined. As shown in Fig. 1a, BY1, which was an albino tea plant, exhibited a pale green or blanched color compared to normal green tea. Ultrastructural analysis revealed a significant reduction in the number of chloroplasts in the mesophyll cells, which exhibited defects and lacked stacked thylakoid structures in BY1 leaves (Fig. 1b). In addition, the plastids of albino BY1 mesophyll cells showed a reduced number of osmiophilic granules and were devoid of starch granules. This was further supported by measurements of chlorophyll a and b content in mature leaves, which showed significant differences between albino BY1 and green SC1 or LJ43 tea plants ( Fig. 1c). Furthermore, the expression levels of CsCHLI, CsCHLD, and CsCHLH genes involved in Mg2+-chelatase were analyzed by RT-qPCR (Fig. 1d). The relative expression levels of CsCHLI were found to be significantly lower in the BY1 cultivars compared to CsCHLD and CsCHLH, while the expression levels of CsCHLH displayed no significant difference in difference cultivars. These results implied that CsCHLI could play a crucial role in the biology of chlorophyll in tea plant. Therefore, the molecular function and mechanism of CsCHLI were conducted in the following research.
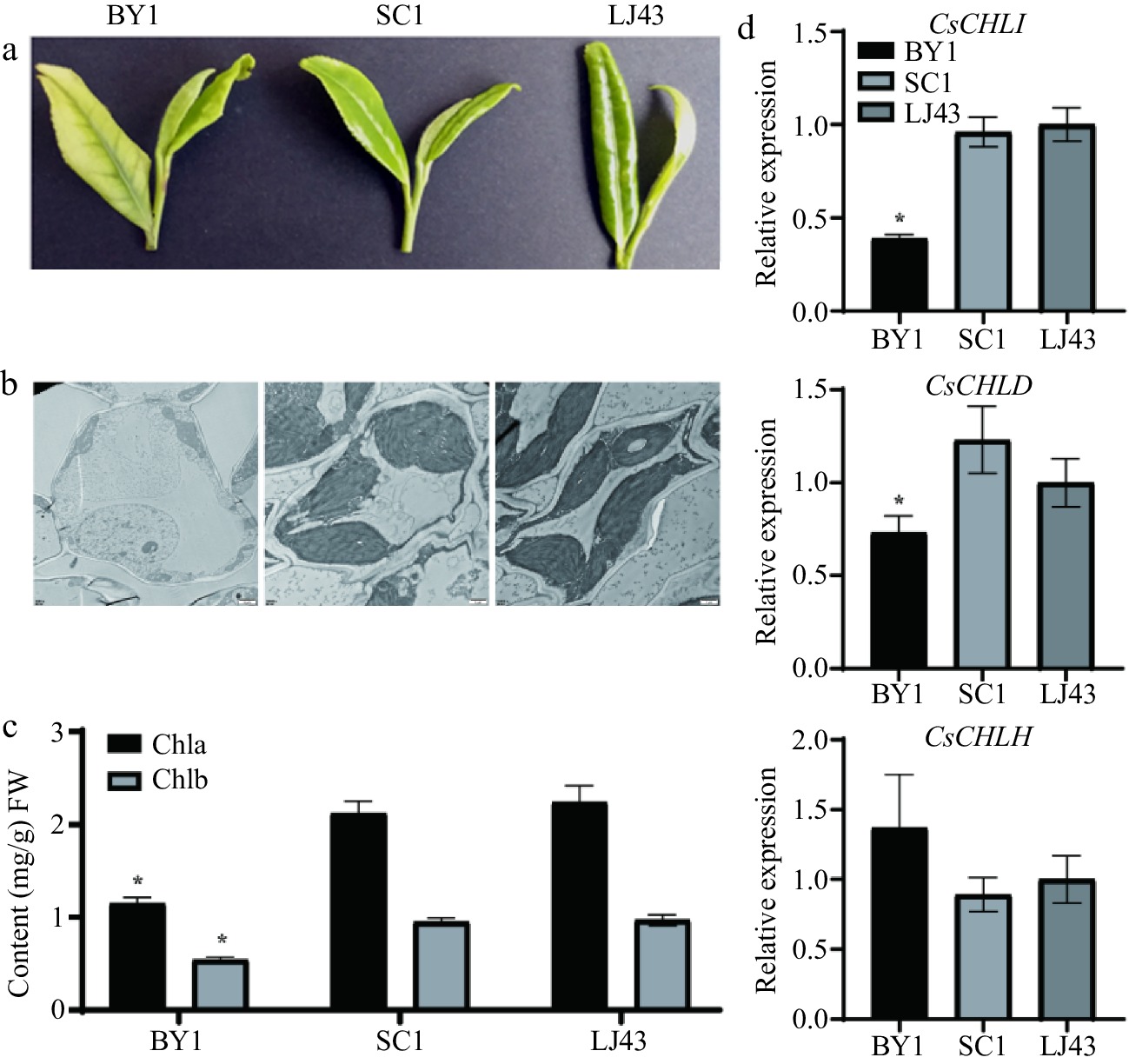
Figure 1.
Leaf phenotypes, leaf ultrastructure, chlorophyll content, and expression levels of CsCHLI, CsCHLD, and CsCHLH genes in albino BY1 and green SC1 and LJ43 tea leaves. (a) Leaf phenotypes of BY1, SC1, and LJ43 plants. (b) Transmission electron microscope images of leaf ultrastructures. (c) Chlorophyll content in the BY1, SC1, and LJ43 leaves. (d) Expression patterns of CsCHLI, CsCHLD, and CsCHLH genes in BY1, SC1, and LJ43 leaves. BY1, Baiye 1; SC1, Shaancha1; LJ43, Longjing43. * means p ≤ 0.05.
Cloning, characterization, and phylogenetic analysis of CsCHLI
-
The full-length of CsCHLI was successfully cloned from the total RNA of 'Shaancha 1'. According to the sequencing results, CsCHLI was 1275 bp, encoded a polypeptide of approximately 46 kDa, and consisted of 424 amino acids (aa), the theoretical pI was 5.42 (Fig. 2a & b; Supplemental Table S2). Based on TargetP analysis, CsCHLI contained a chloroplastic transit peptide (CTP) at the N-terminus.
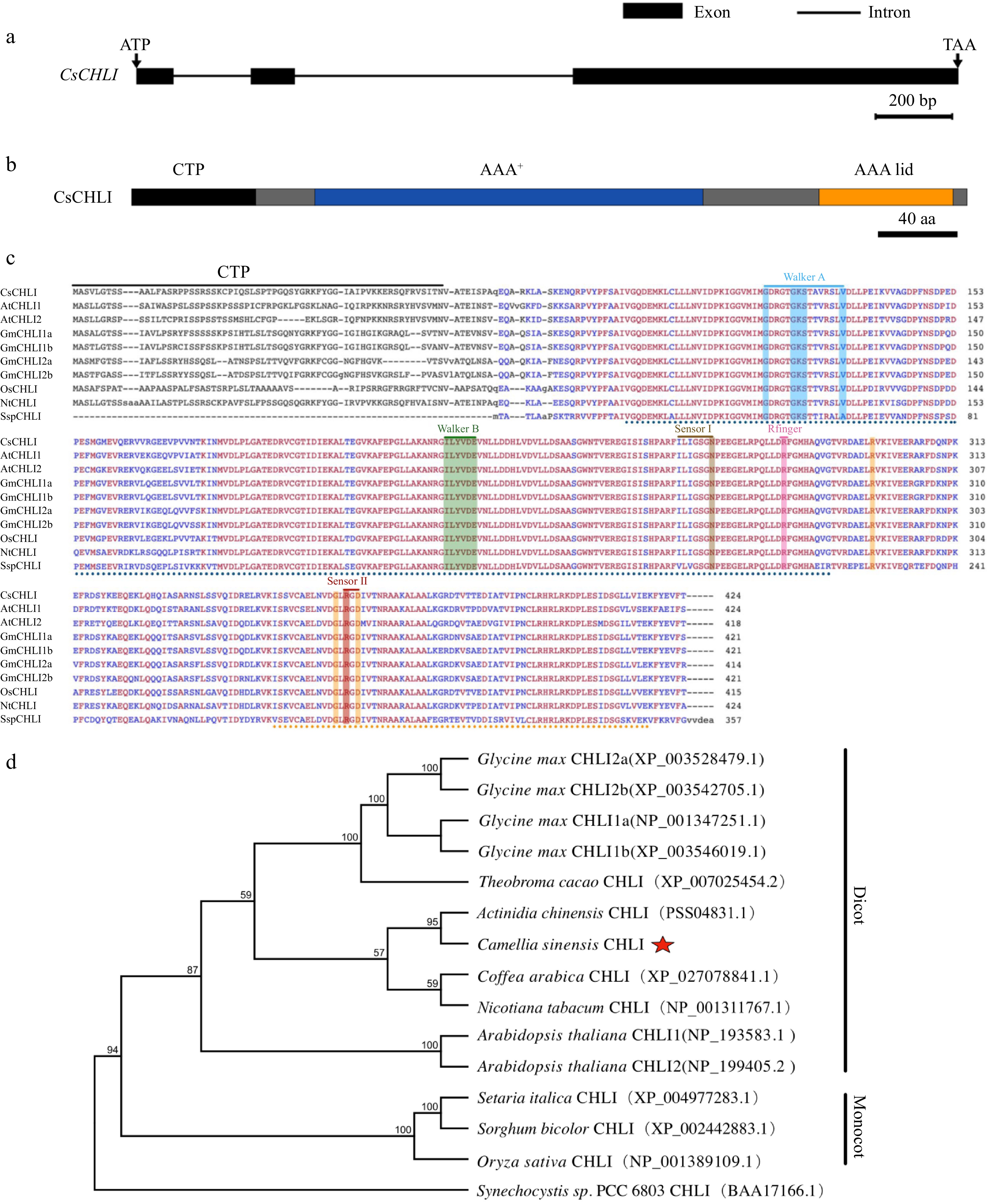
Figure 2.
Schematic structure, multiple sequence alignment and phylogenetic tree of CsCHL. (a) Gene structure of CsCHLI. Dark gray boxes represent coding exons, black lines represent introns, length of nucleotide sequence is represented in the scale. (b) Schematic representation of conserved motifs in CsCHLI. Different coloured rectangles represent different protein domains, length of amino acid sequence is represented in the scale. (c) Multiple sequence alignment of CHLI. The amino acid sequences for analysis include CsCHLI, from Camellia sinensis, GmCHLI1a, GmCHLI1b, GmCHLI2a and GmCHLI2b from Glycine max, OsCHLI from Oryza sativa, NtCHLI from Nicotiana tabacum, AtCHLI1 and AtCHLI2 from Arabidopsis thaliana, and SspCHLI from cyanobacterium Synechocystis sp. PCC 6803. Red and blue highlight the sequences with 100% and 80% identity, respectively. (d) Phylogenetic analysis of CHLI.
To investigate the functional domain and conserved motif of the CsCHLI proteins, multiple sequences were in alignmented. As shown in Fig. 2, CHLI was highly conserved and shared multiple motifs in different species, such as Walker A, Walker B, Sensor I, Arginine fingers (R finger), and Sensor II. Furthermore, the AAA+ (ATPases Associated with Diverse Cellular Activities) domain of CsCHLI was similar to that of AAA lid domain (Fig. 2b & c). Previous studies concluded that the domain of CsCHLI is responsible for binding the ATP α and β phosphates through the Walker-A motif, as well as coordinating the water molecule required for ATP hydrolysis via the Walker-B and sensor I motifs[30]. Therefore, the CsCHLI might have had a specific or exclusive function in ATP hydrolysis activity.
In order to better understand the evolutionary connections between Mg2+-chelatase proteins across various species, a phylogenetic tree was generated using sequence data from cyanobacterium, monocots, and dicots, including one cyanobacteria (Ssp. PCC6803), three monocotyledonous (O. sativa, S. italica, S. bicolor) and six dicotyledonous (A. thaliana, G. max, T. cacao, C. arabica, A. chinensis, N. tabacum) (Fig. 2d). The phylogenetic analysis showed that, CsCHLI was in the branch of dicotyledons, and has a closer genetic relationship with A. chinensis. This result suggested that CHLI may have evolved before the divergence of dicots and monocots, and CsCHLI has more homology to AcCHLI.
Subcellular localization of CsCHLI
-
A gene's function strongly depends on its distribution in cells. For examining the localization of CsCHLI protein, the fused vectors expressing CsCHLI-EYFP under the 35S promoter were constructed. As shown in Fig. 3, free EYFP (35S::EYFP) fluorescence was visualized in both cytosol and nucleus. In contrast, the fluorescent signals of CsCHLI-EYFP was overlapped with those of the auto-fluorescence of chlorophyll in chloroplast. These results indicated that, CsCHLI could be located in the chloroplast.
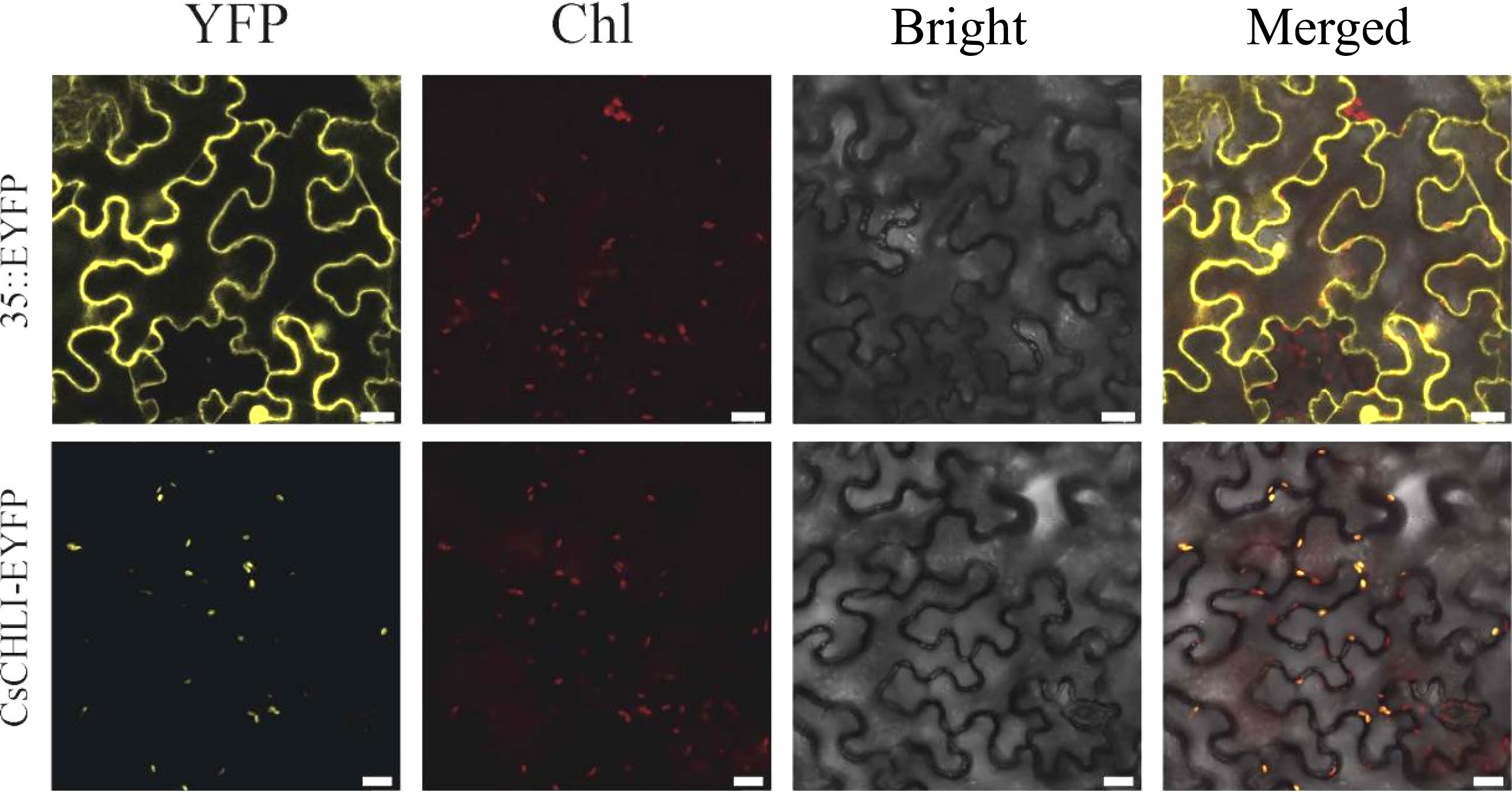
Figure 3.
Subcellular localization of CsCHLI. 35S::EYFP, control vector with 35S promoter; CsCHLI-EYFP, CsCHLI-EYFP fusion protein under 35S promoter; YFP, YFP Fluorescence; Chl, Chlorophyll autofluorescence; Bright, bright field; Merged, Merged images. The scale bar is 20 μm.
Protein-protein interactions of CsCHLI
-
To investigate the interacting partners of CsCHLI in regulatory complexes, the Y2H and BiFC assays were conducted. As shown in Fig. 4, all combination yeast cells had normal growth on the SD-TL medium indicating that the combination vectors were co-transferred into yeast. In the SD-TLHA medium, yeast cells that co-contained a combination of CsCHLI-AD and CsCHLI-BD or CsCHLD-AD and CsCHLD-BD constructs could grow effectively, implying that CsCHLI could interacted with CsCHLI and CsCHLD. However, the yeast carrying CsCHLH-AD or CsCHLH-BD failed to grow on SD-TLHA medium, probably due to it there being more distance between the genetic systems of yeasts and plants.
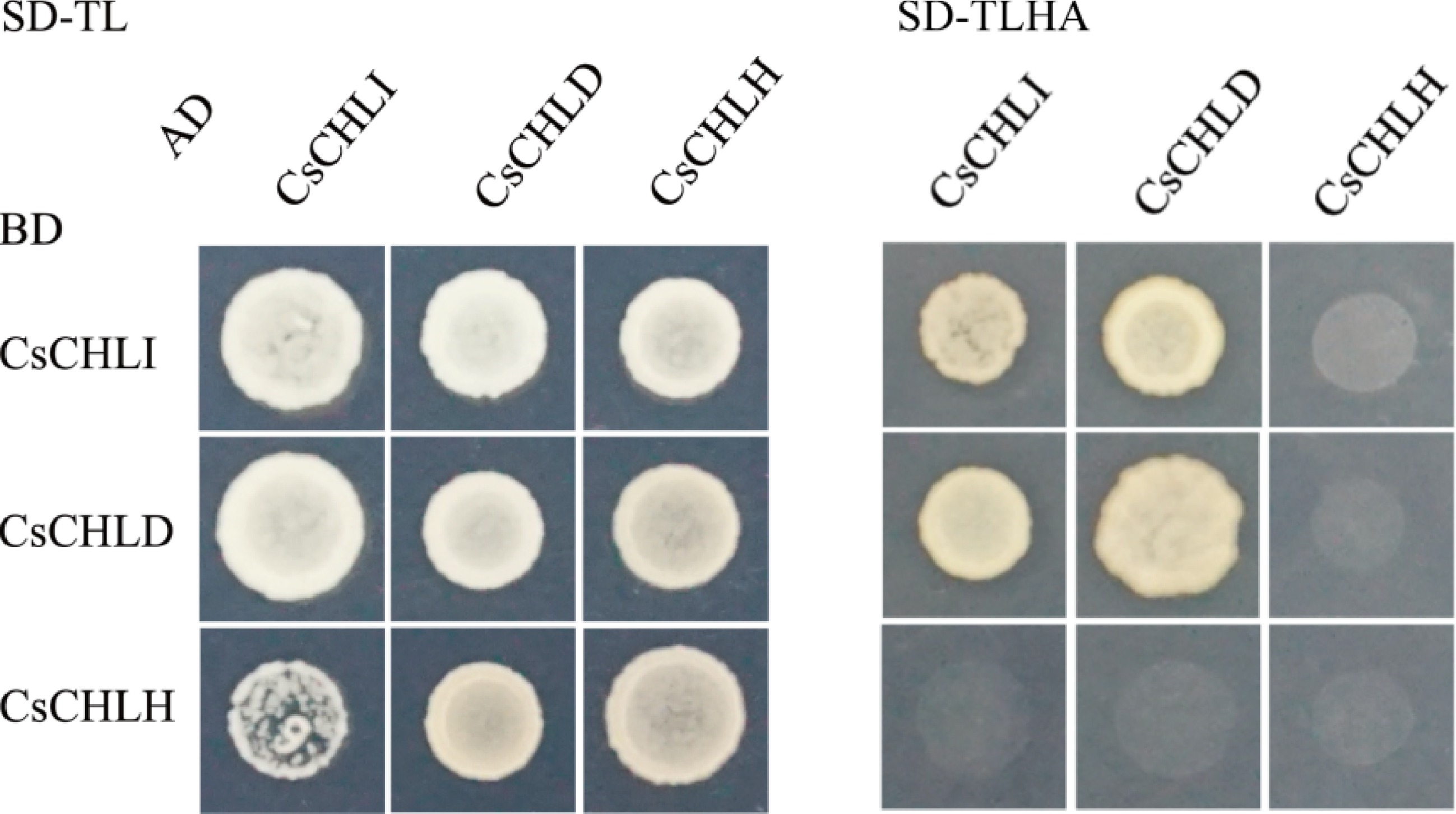
Figure 4.
Yeast two hybrid assay between CsCHLI, CsCHLD, and CsCHLH. pGBKT7 vectors expressing BD-fused CsCHLI, CsCHLD, and CsCHLH were co-transformed with pGADT7 ones expressing AD-fused CsCHLI, CsCHLD, and CsCHLH into the yeast strain Y2H Gold. The transformants were grown on the synthetic dextrose medium (SD) lacking Trp and Leu (SD-TL) or on the SD medium without Trp, Leu, His, and Ade (SD-TLHA).
To further indentify the protein-protein interaction of CsCHLI in plants, the BiFC assays were performed. Following the co-expression of CsCHLI-YFPN and CsCHLI-YFPC, as well as CsCHLH-YFPN and CsCHLD-YFPC, strong YFP fluorescence signals were detected in the leaves of N. benthamiana (Fig. 5). In contrast, the negative control plasmids did not elicit any fluorescent response in the cells (Supplemental Fig. S1). These results demonstrated that the CsCHLI protein could interate with CsCHLD and CsCHLH in plant cells.
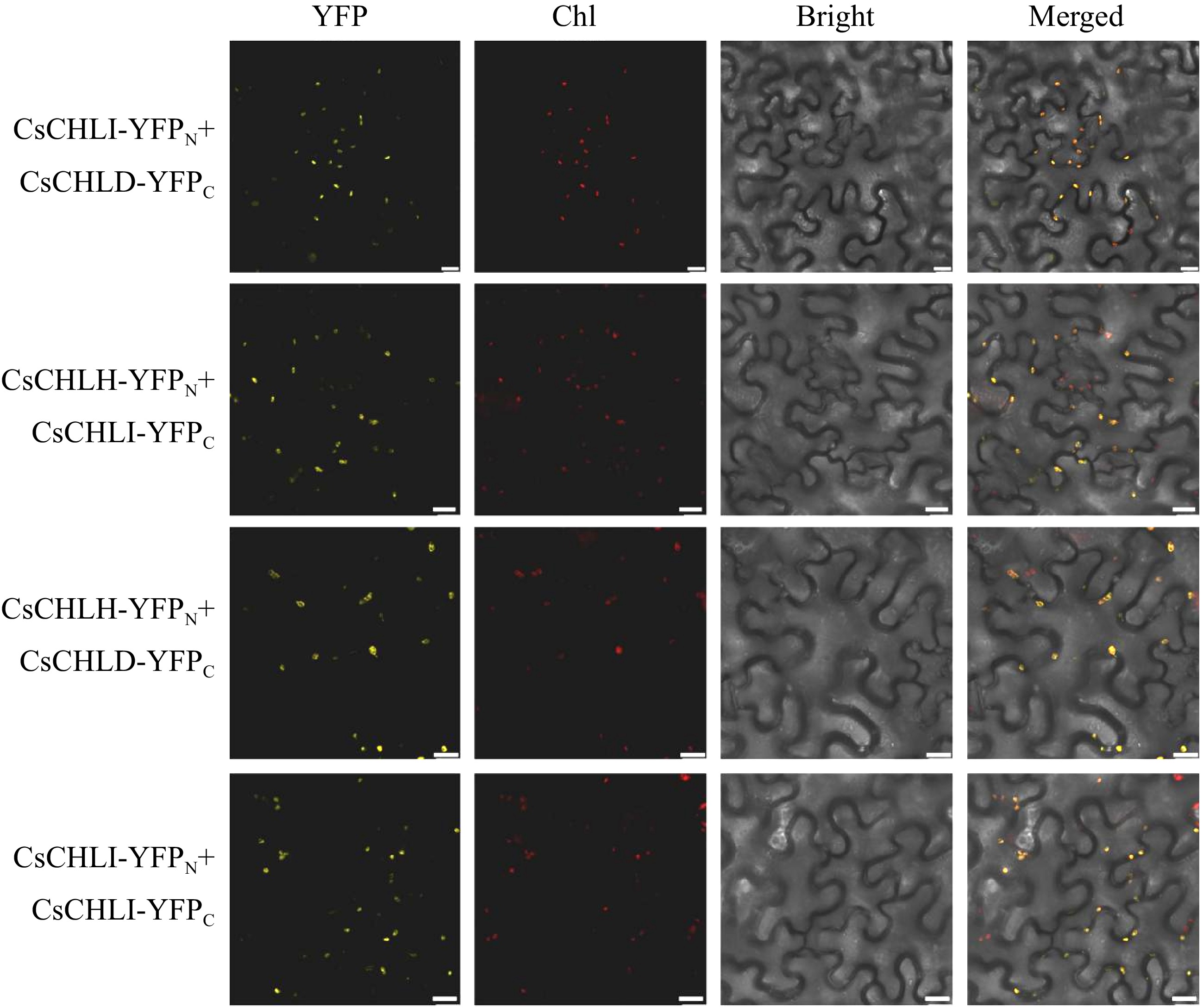
Figure 5.
BiFC assay for examining the interactions of CsCHLI, CsCHLD, and CsCHLH. YFP Fluorescence (YFP), Chlorophyll autofluoresence (Chl), bright-filed (Bright), and Merged images (Merged). The scale bar is 20 μm.
Overexpression of CsCHLI functionally complements atchli1 mutation
-
To further demonstrate the function of CsCHLI in vivo, CsCHLI was overexpressed in Arabidopsis corresponding mutants, atchli1. As shown in Fig. 6, compared to Ler-0, CsCHLI could partially restore the lethal yellowing phenotype of atchli1 in different transgenic lines (Fig. 6a). In addition, 35S::CsCHLI-GFP could be located in chloroplast in Ler-0 and atchli1 (Fig. 6c). Moreover, the contents of chlorophyll-a and chlorophyll-b of transgenic lines atchli1-SI were significantly restored (Fig. 6d). These results indicated that CsCHLI could be located in the chloroplast, and partically restored the reduced chlorophyll content and yellow leaf phenotype.
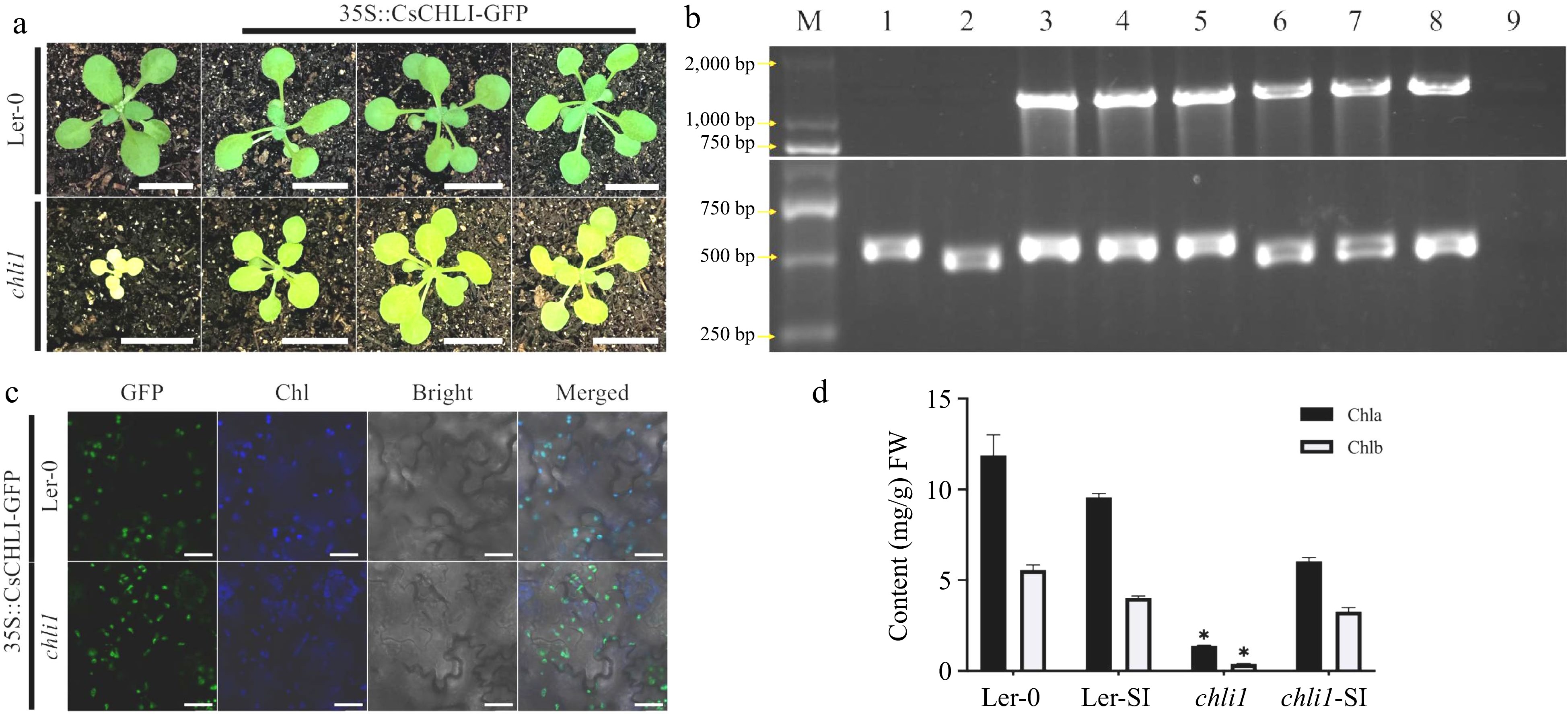
Figure 6.
CsCHLI could restore the yellow leaf phenotype in Arabidopsis. (a) The phenotype of 4-week old Arabidopsis in different lines. Ler-0 wild type background (Ler-0), atchli1 mutant in Ler-0 wild type background (atchli1) and transgenic atchli1 mutants or Ler-0 carrying 35S-driven fused CsCHLI. Scale bar is 1 cm. (b) The identification of different Arabidopsis lines. M: DNA marker, 1: Ler-0, 2: atchli1, 3-5: Ler-0 carrying CsCHLI, 6-8: atchli1 carrying 35::CsCHLI-GFP, 9: water. (c)The fluoresence of different Arabidopsis transgenic lines. YFP Fluorescence (YFP), Chlorophyll autofluoresence (Chl), bright-filled (Bright), and Merged images (Merged). Chlorophyll autofluoresence (Chl) changes to blue. The scale bar is 20 μm. (d) Content of Chlorophyll in different Arabidopsis lines. Ler-SI: Ler-0 carrying 35::CsCHLI-GFP, atchli1-SI: atchli1 carrying 35::CsCHLI-GFP. *: p ≤ 0.05.
-
As is well known, theanine undergoes hydrolysis to form glutamate, which serves as a substrate for chlorophyll synthesis in tea plant[3]. Owing to their special leaf color and high level of theanine, tea cultivars exhibiting albino or etiolation phenotypes have garnered increasing attention among researchers. By the higher theanine content and lower chlorophyll content, 'Baiye 1' has been extensively studied for pigment biosynesis and secondary metabolites. However, there is a limited understanding of the underlying mechanisms of tea leaf color, particularly regarding the accumulation mechanism of chlorophyll. Mg2+-chelatase is a crucial rate-limiting enzyme in the biosynthesis of chlorophyll, and therefore represents a promising target for improving photosynthetic efficiency. Previous studies have demonstrated that knockdown of the gene of CHLI could lead to various physiological and biochemical changes, such as significant reduction in chlorophyll accumulation and yellow-leaf phenotype in Arabidipsis, pea and strawberry[31−33]. In this study, the functional of Mg2+-chelatase I subunits (CsCHLI) in chlorophyll biosynthesis in albino tea plants were investigated. Compared to BY1, SC1 and LJ43, BY1 leaves exhibited the defect of chloroplast structure, including granular stacks and thylakoids, lower chlorophyll content, and significantly decreased expression level of CsCHLI. These results indicate that the gene expression of CsCHLI may be related to the chlorophyll biosynthesis in albino tea cultivars, which could provide an explanation for the differential accumulation of chlorophyll in the tea cultivars.
Analyzing phylogenetic relationships is extremely important for elucidating gene evolution. CHLI was a highly conserved protein, to date, it has been identified in a wide range of plants and bacterial species such as cyanobacteria, Arabidopsis, rice, soybean, tobacco, tomato, etc., indicating that CHLI have important roles in different species[34−36]. As shown in Fig. 2, CsCHLI had closely phylogenetic relationships with dicotyledonous species, suggesting that CsCHLI may potentially serve similar functions in both chlorophyll biosynthesis and metabolism with other dicotyledonous plant. Wei et al.[1] reported that C. sinensis and A. chinensis have a close relationship and diverged from a shared lineage about 80 Mya ago. Meanwhile, a similar result was also found in the multiple sequence alignment and phylogenetic analysis of CHLI, CHLI were highly conserved during the evolution from cyanobacterium to higher plants and the CHLI of C. sinensis and A. chinensis were clustered into one subgroup.
In diverse plant species, the defect of CHLI may cause an etiolation phenotype. Such as, the defect of CHLI Arabidopsis mutant, atchli1, presented yellow leaf and lead to the deficient formation of the photosynthetic apparatus. The silencing of pea CHLI was characterized by underdeveloped thylakoid membranes, altered chloroplast nucleoid structure, and malformed antenna complexes, which resulted in the etiolation phenotype[17]. Many studies have shown that CHLI exhibited the ability to interact with CHLD and CHLH, forming a heterologous complex known as CHLI-D-H. In this study, the analysis of BiFC implied that CsCHLI probably interacted with CsCHLI, CsCHLD, and CsCHLH to form homodimer or heterodimers, however no interaction was detected between CsCHLH and CsCHLI or CsCHLD through Y2H method. Similar results were obtained in the interaction of magnesium chelatase in soybean[37]. This was probably due to the higher molecular weight of CsCHLH (~152 kDa), it maybe unable to fold correctly, thus may not be worked in Y2H. Interestingly, they worked in plants, maybe the interaction between CsCHLH and CsCHLI or CsCHLD is probably required the involvement of other plant proteins. Howerer, it was not presented in yeast strains.
Previous studies have demonstrated that CHLI could restore the etiolation phenotype of corresponding mutants. This was confirmed in wheat mutant chli[38], cucusubmber mutant c528[39], strawberry mutant p240[33]. In order to demonstrate the function of CsCHLI in vivo, overexpression of CsCHLI were transformed into Ler-0 and atchli mutant (Ler-0 wild type background), respectively. As shown in Fig. 6, 35::CsCHLI-GFP is able to locate in the chloroplast, and recover the etiolation phenotype of Arabidopsis mutant atchli1 to a normal green leaf phenotype, meanwhile has no significant affect on the chlorophyll content of Ler-0. Therefore, the increased level of CsCHLI expression observed in the atchli mutant may serve as a compensatory feedback mechanism to counteract the low-chlorophyll stress in tea plant, induced by the decreased activity of Mg-chelatase.
-
Taken together, the function and mechanism of CsCHLI in chlorophyll biosynthesis were identified in tea and Arabidopsis. The analysis between phenotype and gene expression indicated that, the gene expression of CsCHLI has positive correspondence with the chlorophyll content and the chloroplast ultrastructure of albinism tea leaves. Phylogenetic tree analysis confirmed that CHLI were highly conserved from algae to higher plants, and the genetic relationship of CHLI was closer between C. sinensis and A. sinensis. The BiFC analysis showed that CsCHLI has a potential interaction with CsCHLI, CsCHLD and CsCHLH to form protein complexes. Furthermore, the overexpression of CsCHLI could restore the chlorophyll-deficiency phenotype of atchli1 Arabidipsis mutant. This study indicated that CsCHLI located in chloroplast, has a potential protein-protein interaction with CsCHLD and CsCHLH to form a heterologous complex, and performed an important function in the chlorophyll biosynthesis in plants. Our results provide a theoretical basis for the mechanism of albinism tea leaves, and will be beneficial for the albino or etiolation breeding of tea plants.
-
The authors confirm contribution to the paper as follows: Zhao Y and Wang W contributed equally to this work. Study conception and design: Gao Y, Xiao B; experiments operation: Zhao Y, Wang W, Zhan X, Zhang M, Hou X; draft manuscript preparation: Zhao Y, Wang W; manuscript revision: Gao Y, Xiao Y, Gao M. All authors reviewed and approved the final manuscript.
-
The datasets used in this study are available from the corresponding author upon reasonable request (Yuefang Gao, yuefanggao@nwafu.edu.cn).
This work was financially supported by the National Natural Science Foundation of China (31700612), and the Key Research and Development Program of Shaanxi Province (2021ZDLNY04-03, 2023-YBNY-145). We acknowledge Prof. Min Xu (Northwest University, Xi'an, Shaanxi, China) for providing the seeds of atchli1 mutants.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Yiqing Zhao, Wenjing Wang
- Supplemental Table S1 The primers used in this research.
- Supplemental Table S2 Physiological and biochemical properties of CsCHLI.
- Supplemental Fig. S1 BiFC of free YFPC or YFPN coexpressed with CsCHLI, CsCHLD, and CsCHLH. YFP Fluorescence (YFP), chlorophyll autofluoresence (Chl), bright-filed (Bright), and Merged images (Merged). The scale bar is 20 μm.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhao Y, Wang W, Zhan X, Zhang M, Xiao Y, et al. 2024. CsCHLI plays an important role in chlorophyll biosynthesis of tea plant (Camellia sinensis). Beverage Plant Research 4: e004 doi: 10.48130/bpr-0023-0037 

CsCHLI plays an important role in chlorophyll biosynthesis of tea plant (Camellia sinensis)
- Received: 13 September 2023
- Revised: 26 October 2023
- Accepted: 09 November 2023
- Published online: 01 February 2024
Abstract: Chlorophyll biosynthesis is a crucial biological process in plants, and chlorophyll content is one of the most important traits in the yield and quality of tea. Magnesium chelatase is a conserved enzyme complex responsible for chlorophyll biosynthesis, which is composed of the subunit of CHLI, CHLD and CHLH. In this study, there was a positive correlation between the expression of CsCHLI, chlorophyll content and chloroplast structure. The CsCHLI gene structure and functional domain indicated that its cDNA length was 1,275 bp, encodes 424 amino acids, and consists of cTP, AAA+ and AAA lid domains. Meanwhile, the subcellular localization demonstrated that CsCHLI localized in chloroplasts. In addition, protein-protein interaction analysis by yeast two-hybrid (Y2H) and bimolecular fluorescence complementation (BiFC) assays indicated that CsCHLI could interact with CsCHLI to form homodimer, or perhaps interact with CsCHLD and CsCHLH to form heterodimer. Moreover, Arabidopsis transformation displayed that overexpression of CsCHLI could restore the etiolation phenotype of the atchli1 mutant. These findings provide the mechanistic function of CsCHLI and its implications in chlorophyll biosynthesis in tea plant.
-
Key words:
- CsCHLI /
- Albino /
- Chlorophyll biosynthesis /
- Camellia sinensis