










-
Walnut is one of the most important economic oil tree species and plays a vital role in rural revitalization and poverty alleviation in China[1]. Currently, the planting area of walnut trees ranks first among the four dried fruits (walnuts, almonds, cashews, and hazelnuts). However, the various stresses caused by unstable environmental changes can affect the yield and quality of walnuts, especially drought, abnormal temperature changes, and soil salinization are the main abiotic stimuli restricting the development of the walnut industry in the northwest region of China, which is the main production area. During severe 'late spring cold', all newly sprouted tender branches could be frozen; long-term drought and high temperature can cause different types of diseases and burns, and the commodity rate decreases; saline-alkali soil directly affects the growth and quality of walnuts[1,2]. To survive in poor environments, plants have evolved a variety of sophisticated strategies to alleviate damage via various pathways, such as releasing stress signals, regulating related genes' expression, physiological responses[3]. In regulation mechanism, plant transcription factors (TFs), such as MYB (myeloblastosis), ERF (ethylene-responsive element binding factor), WRKY (containing the WRKYQK protein domain), NAC (NAM/ATAF/CUC domain), bHLH (basic helix-loop-helix), perform an important role in transmitting stress-induced signals and coordinating the functional genes' expression in cells[4,5]. In signal transduction networks of developmental processes and stress conditions, plant protein kinases and protein phosphatases play key functions[6]. Reactive oxygen species (ROS) signaling is a prominent pathway for plants to respond to stress[1, 5]. Protein phosphatase 2A (PP2A), a group of serine/threonine (Ser/Thr) phosphatases, was verified to be involved in the ROS pathway to regulate metabolic changes and cell death[7]. Clearly, identification of key factors bound to stress response, such as PP2A, is an important basis for revealing the molecular mechanism of walnut response to osmotic stress.
PP2A exists as a trimer made up of three subunits (A, B and C) in eukaryotic cells. The A, B and C subunits are the structural/scaffold, regulatory, and well-conserved catalytic subunit, accordingly[8]. Subunit A is a complex formed by a series of 15 helical HEAT repeats, each repeat consists of approximately 40 leucine residues, which provide a scaffold for the binding of B and C; Subunit C is strongly conserved among different species[9]. Subunits A and C shape the core enzyme which interacts with B-subunit to produce the holoenzyme; Subunit B is the 'specificity unit' that determines the specificity of PP2A holoenzyme[10]. The B subunit could be further classified into three sub-classes, they are B', B'' and B''' (B55)[11]. Genes encoding PP2A subunits have been identified and characterized in some plant species. In Arabidopsis thaliana, the A, B and C subunit of PP2A contains three, 17 and five members, accordingly[12]. In Hevea brasiliensis, four, 24 and eight members of A, B and C subunit were identified[13]. In other species, the B subunit was also discovered to contain the most members of the three[8−11]. There are differences in the quantity of the PP2A family among different species, but the B subunit group covers most members, which may be related to the functions of various types of subunits.
PP2A proteins are reported to be involved in several notable biological processes, such as stress-related signaling, which includes abundant hormone-related signal transduction pathways and enzymes[7−12]. In Arabidopsis, PP2A dephosphorylation and auxin efflux proteins' correct orientation are necessary for auxin polar transport[14]; the PP2A B subunit B′α and B′β activate brassinosteroid signaling by dephosphorylating the BZR1 TF in the nucleus[15]; the RCN1, a PP2A A subunit gene, was reported to be involved in the regulation of MeJA/ABA/ethylene signaling[16]. In rice, the expression of all catalytic subunit genes (OsPP2A-1-5) was significantly up-regulated by salinity stress[17]. In wheat, over-expression of TaPP2AC-1 in transgenic tobacco enhanced drought tolerance through drought-responding signal transduction pathways[18]. While the expression of TaPP2Ac-4B and TaPP2Ac-4D negatively regulated the defense response to R. cerealis infection may modulate the expression of certain pathogen-response (PR) and ROS-scavenging-related genes[19]. These studies indicated that PP2As have diverse functions in plant stress response, and the walnut PP2A family deserves deep attention for revealing the molecular mechanism of stress adaptation.
In recent years, the diverse roles of PP2A in herbaceous plants, especially its positive roles in response to some abiotic stresses, have attracted widespread attention. However, little is known about the identification and characterization of PP2A genes in woody plants. In this study, we identified the walnut PP2A gene family members and further analyzed their phylogenetics, gene structure, chromosome location, conserved motifs, conserved domains, expression patterns, and interaction proteins. Moreover, transgene lines overexpression of candidate walnut PP2A genes were obtained to confirm the roles in response to osmotic stress. The results of the current study revealed that walnut PP2A genes play positive roles in osmotic stress response by interacting with JrVIP1 protein to control ROS scavenging in a Ca- and ABA-dependent manner.
-
Three-year-old 'Xiangling' walnut (a variety widely planted in China) grafting seedlings were planted in flowerpots and grown in a greenhouse at the College of Forestry, Northwest Agriculture and Forestry University (China) with a 14/10 h photoperiod and relative humidity 70% ± 5% under 22 ± 2 °C and used as the materials for the stress treatments. The soil and growth environment as well as the genetic background are consistent. Considering that osmotic stress always resulted from drought, and saline-alkali conditions and PP2A genes probably relate to ABA signaling, the plants were treated with 20% (w/v) PEG6000, 300 mmol/L NaCl, 20 mmol/L CaCl2 and 30 μmol/L ABA by watering the roots, respectively. Normally watered plants were used as controls. After treatment for 5 d, the leaves were collected and stored at −80 °C for RNA isolation. All treatments were replicated three times and each replicate included five seedlings.
Identification of PP2A members from walnut transcriptomes
-
The walnut leaf transcriptomes under stresses of drought, salt, and ABA (same as above treatments) were sequenced and a preliminary sequence set was obtained. Then firstly 'protein phosphatase' was subjected to search the transcriptomes' database to obtain PP2A family candidate sequences, which were further separately blasted (BLAST: Basic Local Alignment Search Tool (nih.gov)) to judge what homologous proteins they belong to. Next, the open reading frame (ORF) of each potential walnut PP2A (marked as JrPP2A) was confirmed using ORF Finder (
www.ncbi.nlm.nih.gov/orffinder ). The conserved domains of JrPP2A proteins were confirmed based on the online tools including CD-Search (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ), Pfam (http://pfam.janelia.org/ ), and SMART (http://smart.embl-heidelberg.de/ ). The conservative motifs were found using MEME online tools (https://meme-suite.org/meme/ ) and Tbtools[20] with the following parameters: the motif number was 20, any repetition with motif width 17~50. After these analyses, members of the JrPP2A family were confirmed. The amino acid number, molecular weight and theoretical isoelectric point (pI) of JrPP2A proteins were analyzed by ExPASy (https://web.expasy.org/protparam/ ). To clarify the evolutionary relationship of JrPP2A proteins, a neighbor-joining phylogenetic tree with a bootstrap replicate value of 1,000 was constructed in MEGA7 using the JrPP2A proteins, 25 Arabidopsis PP2A proteins downloaded from TAIR and 63 H. brasiliensis PP2A proteins downloaded from NCBI (Hevea brasiliensis (ID 503)-Genome-NCBI (nih.gov)). The phylogenetic tree was modified using Evolview (www.evolgenius.info/evolview ) and the JrPP2A members were grouped referring to the topology of the phylogenetic tree.Chromosomal location, gene structure and promoter analysis of JrPP2As
-
The genomic DNA sequence of JrPP2As were confirmed according to the walnut (Juglans microcarpa × J. regia) genome (
www.ncbi.nlm.nih.gov/genome ) and the chromosomal location information of JrPP2As was determined. The exon-intron structure of each JrPP2A was defined using Gene Structure Display Server 2.0 (GSDS 2.0:http://gsds.gao-lab.org/ ). An up-stream 2,000 bp promoter sequence of each JrPP2A was obtained according to the genome, and the potential cis-acting regulatory elements of the promoters were predicted by PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ).Expression analysis of JrPP2As under osmotic stress
-
The leaves of 3-year-old walnut plants treated separately by PEG6000, NaCl, CaCl2 and ABA for 5 d were sampled for RNA isolation. The total RNA of each sample was isolated using the cetyltrimethylammonium ammonium bromide (CTAB) method[1, 5] and digested with DNase (Takara, Dalian, China) to ensure the RNA quality. The RNA concentration was tested using the Thermo Scientific™ NanoDrop™ One. 0.5 μg RNA of each sample was reversely transcribed into cDNA using the Prime Script™ RT reagent Kit (CWBIO, Beijing, China). The cDNA was diluted 10-fold with ddH2O and used as the template of real-time fluorescent quantitative PCR (qRT-PCR). The 20 μL reaction solution contains 10 μL SYBR Green Real-time PCR Master Mix (CWBIO), 0.5 μM each forward and reverse primer, and 2 μL cDNA template. StepOne™ Real-Time PCR System was adopted to perform the qRT-PCR. The amplification parameters: 94 °C/30 s, followed by 44 cycles of 94 °C/12 s, 60 °C/30 s, 72 °C/40 s, then 81 °C/1 s. The internal reference gene is 18S rRNA (HE574850)[21]. All related primers are listed in Supplemental Table S1. The relative expression levels were calculated using the 2−ΔΔCᴛ method[22].
Yeast two hybrid assay
-
Yeast two hybrid (Y2H) assay was used to clarify whether JrPP2A proteins could interact with basic leucine zipper protein (bZIP) JrVIP1. JrVIP1 was cloned into pGBKT7 vector (marked as BD) to form the bait recombinant (BD-JrVIP1). Each JrPP2A was independently inserted into the pGADT7_Rec vector (marked as AD) to generate the prey recombinants (AD-JrPP2As). Meanwhile, JrVIP1 was inserted into pGADT7_Rec to form AD-JrVIP1, while each JrPP2A gene was independently cloned into pGBKT7 to generate BD-JrPP2As. Then the interaction between BD-JrVIP1 and each AD-JrPP2A as well as AD-JrVIP1 and each BD-JrPP2A were confirmed in yeast Y2H by grown on the SD/-Ade/-His/-Leu/-Trp/X-α-Gal/Aureobasidin A (QDO/X/A) medium plates. The empty AD and BD functioned as interaction control[1,5]. The related primers were listed in Supplemental Table S2.
Qualitative and quantitative analysis of reactive oxygen species
-
To understand the osmotic stress response function, the JrPP2A genes were separately inserted into the pROKII vector to generate recombinant vectors 35S::JrPP2As. Then each 35S::JrPP2A was transformed into Arabidopsis using the Agrobacterium tumefaciens-mediated method[1,5]. The kanamycin-resistant transformed seedlings were further confirmed by PCR and qRT-PCR methods. The most overexpression lines were chosen for osmotic stress response analysis. The seeds of WT and JrPP2A transgenic lines were sown on 1/2MS (Murashige and Skoog) agar medium for 12 d, then the seedlings were transfered into soil to grow to one month old for treatments, including 10% PEG6000, 10% PEG6000 + 10 mol/L CaCl2, 10% PEG6000 + 10 μmol/L ABA, 100 mmol/L NaCl, 100 mmol/L NaCl + 10 mol/L CaCl2, 100 mmol/L NaCl + 10 μmol/L ABA. After 3 d of treatments, the leaves were harvested for reactive oxygen species (ROS) determination. The 3,3'-diaminobenzidine (DAB) and nitroblue tetrazolium (NBT) staining were applied to qualitative confirmation of the ROS generation. The H2O2 content was determined using the Hydrogen Peroxide Assay Kit (colorimetry, A064-1, NJJCBIO, Nanjing, China). The total ROS content was tested using the chemiluminescence method according to the manufacturer's instructions of the Reactive Oxygen Species Assay Kit (E004, NJJCBIO).
Determination of calcium content in transgenic Arabidopsis plants
-
The leaves of WT and JrPP2A transgenic lines were ground and dried into a powder, independently. Then each sample powder was placed into a centrifuge tube, in which 1.9 mL of 1% HCl was added and mixed well with a vortex shaker. The mixture was centrifuged (15,000 r/min) after digested at 37 °C for 48 h. 1.5 mL of the supernatant was taken to measure the concentration of Ca2+ using a TAS-990 atomic absorption spectrometer at 422.7 nm.
Pull-down assay
-
The CDSs of JrPP2As and JrVIP1 were independently cloned into the vectors of pET30a and pGEX4T-1 and then transformed into Rosetta for expression of JrPP2A-His and GST-JrVIP1 proteins by using 0.1 mM IPTG (isopropyl-b-thiogalactopyranoside). Soluble GST or GST-JrVIP1 fusion proteins were extracted and immobilized using a glutathione HiCap matrix (Qiagen). JrPP2A-His was incubated with immobilized GST or GST-JrVIP1, and the interaction was checked by western blotting analysis. The related primers were included in Supplemental Table S2.
Statistical analysis
-
All the data were organized and analyzed using Excel 2023 and SPSS (Chicago, Illinois, USA). The sample variability was described by standard deviation (S.D.) from three repeated assays. The differences between WT and JrPP2A transgenic lines were evaluated using Tukey's multiple comparison test (p < 0.05).
-
A total of 18 putative JrPP2A genes were screened from walnut transcriptome, among these 18, three lacked PP2A catalytic domain. Therefore, 15 genes in J. regia were identified as PP2A family members in the current study. To better describe these 15 JrPP2A genes, their positions on the walnut chromosomes were analyzed, and found that these 15 walnut PP2A genes were unevenly distributed on 10 chromosomes: Four were distributed on the 15th chromosome, which has the maximum number of JrPP2As, followed by the 01st and 16th chromosome, those have two JrPP2As, accordingly; The chromosomes of 02nd, 03rd, 05th, 09th, 10th and 12th, each covers only one JrPP2A gene. In addition, there was no JrPP2A member presented on the chromosomes of the 04th, 06th, 07th, 11th, 13th and 14th. Considering the conciseness of the description, the JrPP2A genes were named JrPP2A01 to JrPP2A15 according to their orders in the chromosomes (Fig. 1, Table 1). The ORFs of the 15 JrPP2As were between 651 bp (JrPP2A12) and 1,764 bp (JrPP2A10), consisting of 216~587 amino acids. The molecular weight of the proteins ranged from 24.15 kDa (JrPP2A12) to 65.61 kDa (JrPP2A10), and the pI ranged from 4.80 (JrPP2A02) to 8.37 (JrPP2A01) (Table 1).
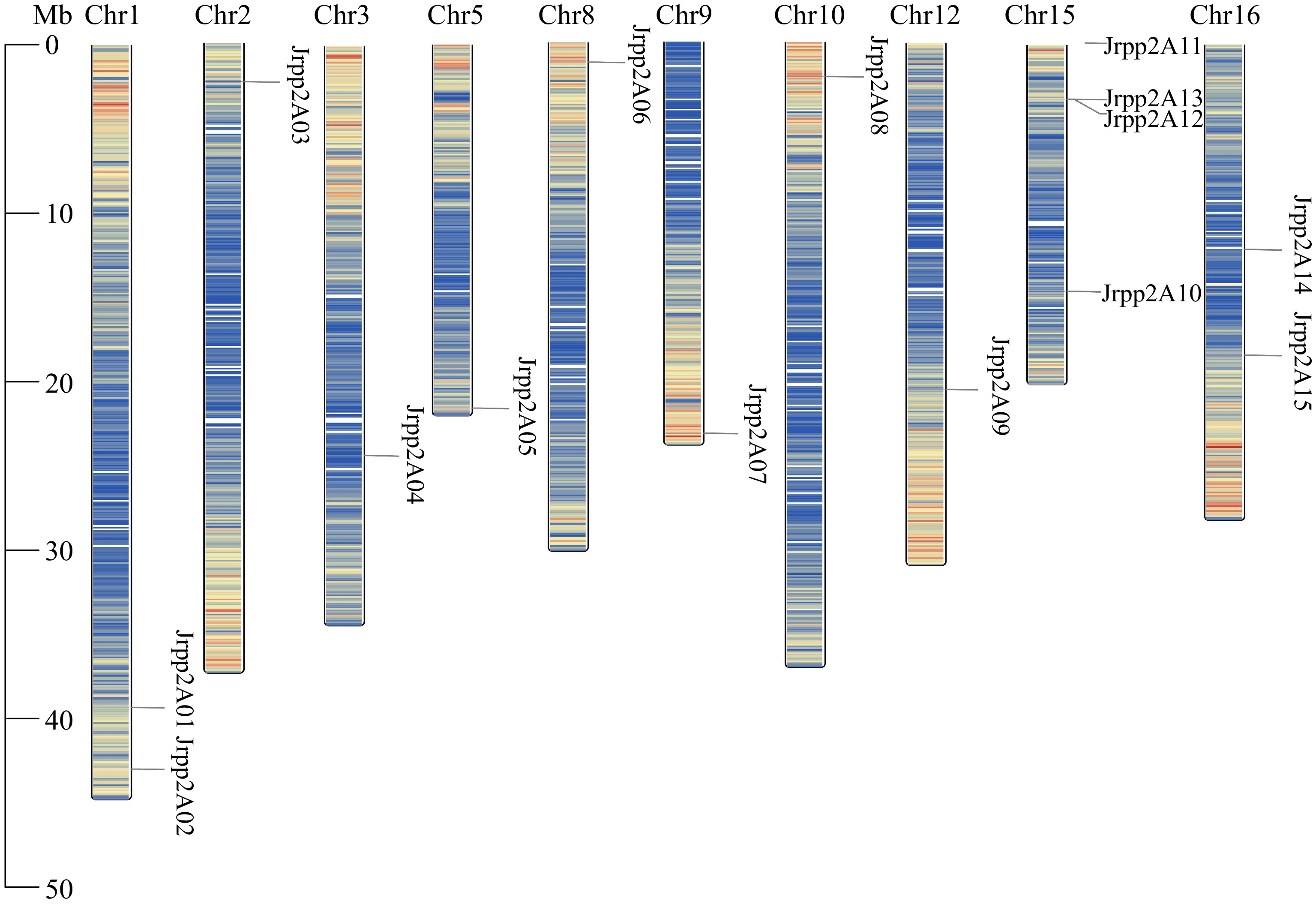
Figure 1.
Distribution of the JrPP2As on chromosomes of the J. regia genome. The chromosome number is shown on the top side of each chromosome.
Table 1. Sequence characteristics of 15 JrPP2As.
Gene names Type Transcriptome No. Gene Bank accession No. Gene ID Chromosome ORF length (bp) Number of amino acids Molecular weight (kDa) pI JrPP2A01 B' comp30409_c0 XM_018967286.2 LOC108992675 ch1 1,452 483 55.13 8.37 JrPP2A02 B'' comp28147_c0 XM_018971752.2 LOC108996025 ch1 1,254 417 49.44 4.8 JrPP2A03 B55 comp26329_c0 XM_018982199.2 LOC109003870 ch2 1,506 501 56.72 6 JrPP2A04 B' comp20037_c0 XM_018993415.2 LOC109011996 ch3 1,503 500 57.52 6.24 JrPP2A05 C comp26715_c0 XM_018952439.2 LOC108981323 ch5 681 226 25.7 4.94 JrPP2A06 B' comp24655_c0 XM_018979195.2 LOC109001785 ch8 1,503 500 56.94 7.61 JrPP2A07 C comp32898_c0 XM_018969187.2 LOC108994090 ch9 918 305 34.78 5.24 JrPP2A08 B' comp25187_c0 XM_018958784.2 LOC108986228 ch10 1,575 524 59.87 7.97 JrPP2A09 B'' comp9850_c0 XM_018985083.2 LOC109005962 ch12 1,431 476 54.74 4.93 JrPP2A10 A comp28348_c0 XM_035685846.1 LOC109013629 ch15 1,764 587 65.61 4.88 JrPP2A11 B55 comp28413_c1 XM_018989098.2 LOC109008856 ch15 918 305 34.06 5.16 JrPP2A12 B55 comp28413_c1 XM_018989097.2 LOC109008856 ch15 651 216 24.15 6.21 JrPP2A13 C comp27670_c1 XM_018980847.2 LOC109002920 ch15 912 303 34.84 4.88 JrPP2A14 C comp23892_c0 XM_018963562.2 LOC108989812 ch16 921 306 35.01 4.83 JrPP2A15 B' comp13497_c0 XM_018951013.2 LOC108980163 ch16 1,530 509 58.52 7.18 Classification of JrPP2A proteins according to phylogenesis and gene structure
-
Classification may have a potential relationship with gene function. Therefore, the 15 JrPP2A proteins were classified according to the widely accepted approaches mainly including evolutionary relationship and gene structure. To investigate the genetic relationship, total 25 Arabidopsis PP2As (three A subunits-AtPP2AA, nine B' subunits-AtPP2AB', six B'' subunits-AtPP2AB'', two B55 subunits-AtPP2AB55, and five C subunits-AtPP2AC), 36 rubber tree PP2As (four A subunits-HbPP2AA, 14 B' subunits-HbPP2AB', six B'' subunits-HbPP2AB", four B55 subunits-HbPP2AB55, and eight C subunits-HbPP2AC) as well as the 15 walnut PP2As were aligned to construct a phylogenetic tree using NJ method. As shown in Fig. 2, these PP2A proteins were divided into five clusters according to the branch of the evolutionary tree and subunits covered in Arabidopsis and rubber PP2As. The B' cluster covered most PP2A proteins, including five JrPP2As (JrPP2A01, JrPP2A04, JrPP2A06, JrPP2A08, JrPP2A15), nine AtPP2As and fourteen HbPP2As. Group A had the least number of PP2As, containing one JrPP2A (JrPP2A10), three AtPP2As and four HbPP2As. Subfamily B'' covered 2 JrPP2A proteins (JrPP202, JrPP209), while subclass B55 included three JrPP2As (JrPP203, JrPP211, JrPP212). The other four JrPP2As (JrPP2A05, JrPP2A07, JrPP2A13, JrPP2A14) were clustered in C subgroup (Fig. 2).
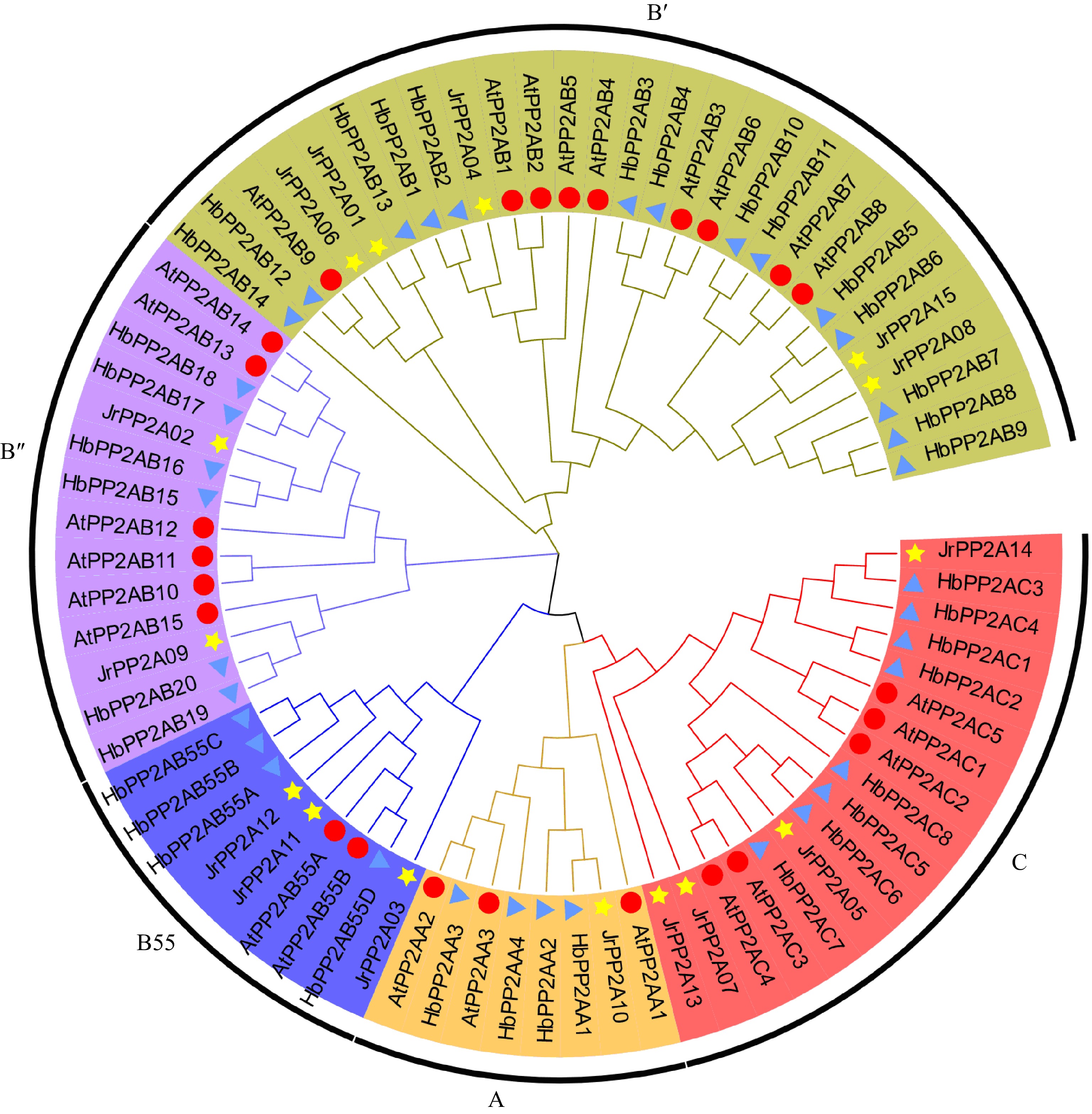
Figure 2.
Phylogenetic relationship of PP2A proteins from J. regia, A. thaliana and H. brasiliensis. A, C, B55, B', B" means five sub-family of PP2As, respectively, which are displayed in different colors. A total of 15 walnut PP2As are represented by yellow five-pointed stars, 25 Arabidopsis PP2As are represented by red circles, 36 H. brasiliensis PP2As are represented by blue triangles.
Considering that the exon-intron structure can provide prominent evidence supporting the phylogenetic relationships of a gene family, the diversity of the intron-exon construction leads to the gene structure being varied. Thus, a structural analysis was performed using these 15 CDSs and genomic sequences of JrPP2A genes in the GSDS online. As shown in Fig. 3a, the intron numbers of the 15 JrPP2A genes varied widely among the five subfamilies. In detail, the A and B" sub-family JrPP2A genes contained 12 and 11 introns, accordingly. The intron numbers of C subclass JrPP2A genes ranged from 6 (JrPP2A14) to 10 (JrPP2A13). The B55-like JrPP2A genes possessed the most introns (14), while the B' sub-group JrPP2A genes had the fewest introns (only 1~2). The performance of the introns validated that the structure of exon–intron was similar in the same subfamily, despite some differences in the length of exons. The showing exon–intron structure was consistent with the evolutionary relationship.
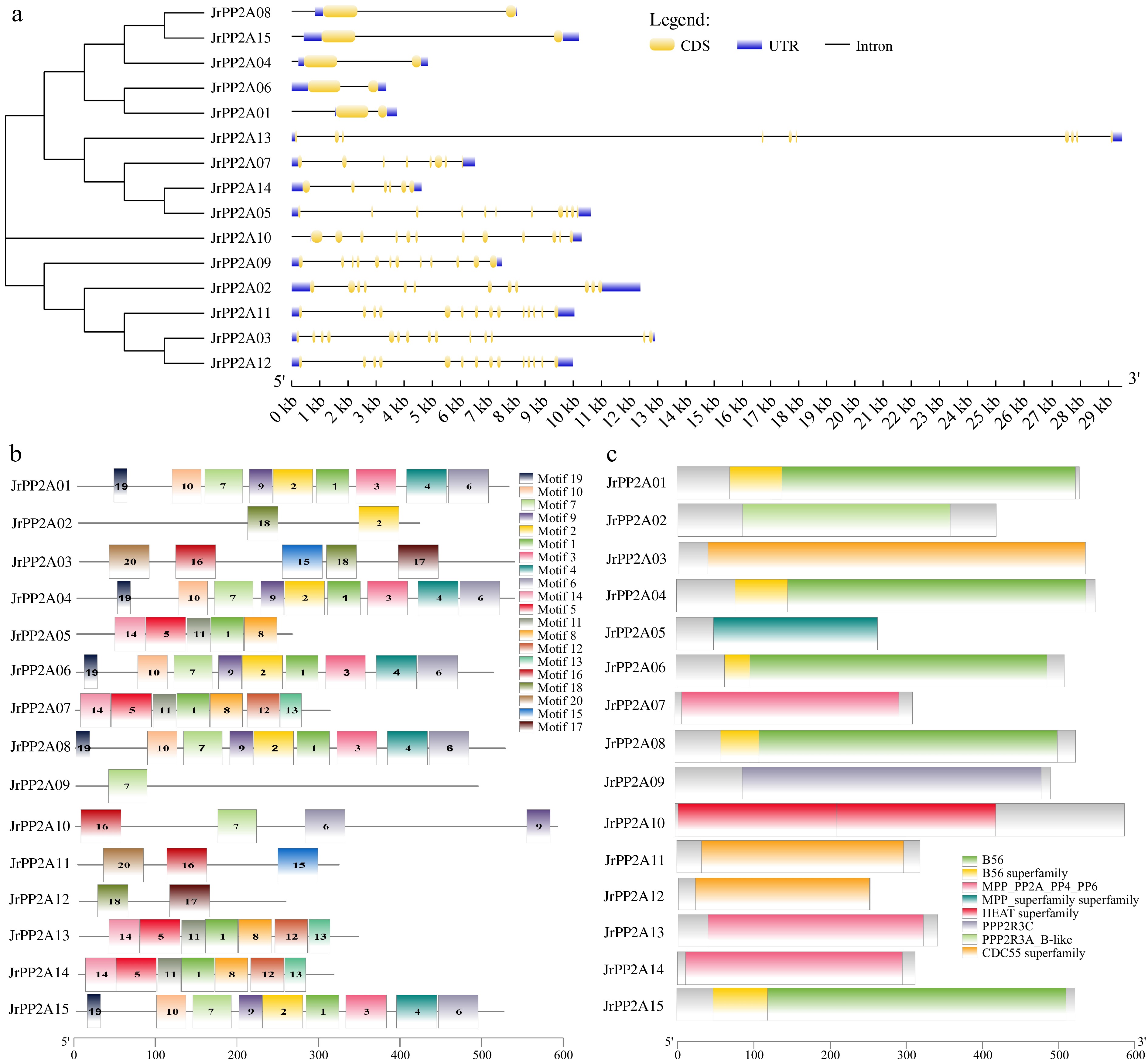
Figure 3.
Gene structure of walnut PP2As. (a) Exon-intron distribution map. The vertical phylogenetic tree and gene structure of JrPP2As was constructed by GSDS online software. Yellow boxes indicate exons; blue boxes indicate upstream or downstream; black lines indicate introns. (b) Conserved motif analysis: 20 separate patterns were identified with the MEME suite and each pattern was depicted with different colors. (c) Distribution of conserved domains.
Conserved motifs and domains of walnut PP2A proteins
-
A total of 20 conserved motifs were identified from the 15 walnut PP2A proteins using an online MEME tool and the basic information (width and best possible match sequence) was downloaded and displayed in Table 2. Each motif included 17–50 amino acids. The proteins categorized within the same group shared a similar motif composition, which further verified the group definitions. As shown in Fig. 3b, some unique motifs could be detected only in specific subgroups. In detail, motif 19, motif 10, motif 7, motif 9, motif 2, motif 1, motif 3, motif 4 and motif 6 was detected in B' subunit JrPP2A proteins; Motif 14, motif 5, motif 11, motif 1 and motif 8 was found in all C subunit JrPP2As. Motif 16, motif 7, motif 6 and motif 9 was contained in A subunit of JrPP2As; Motif 20, motif 6, motif 15, motif 18 and motif 17 was located in subgroup B55 JrPP2As; while subgroup B'' JrPP2A proteins only contained motif18, motif2 and motif7.
Table 2. Motif sequences identified by the MEME tool.
Motif Width Motif consensus 1 41 TIVYGFYDETERHNGIAELLEIFGSIIDGFALPLKEEHKIF 2 50 KVAKRYIDHSFVLRLLDLFDSEDPREREYLKTILHRIYGKFMVHRPFIRK 3 50 HKPKSIGLYHQQLSYCITQFVEKDPKLADTVIRGLLKYWPVTNSQKEVMF 4 50 PAEFQRCMVPLFRQIGCCLNSSHFQVAERALFLWNNDHIVNLIAQNRNVI 5 50 NVQPVKSPVTICGDIHGQFHDLIELFRIGGNCPDTNYLFMGDYVDRGYYS 6 50 PIIFPALEKNARSHWNQAVQNLTLNVRKIFSEMDPELFEECQRQFQEDEA 7 48 DIKRQTLIELVDFVASGSGKFTETAIQEMIKMVSVNLFRVLPPKPREN 8 41 CLHGGLSPSIETLDNIRVIDRIQEVPHEGPMCDLLWSDPDD 9 29 EPSFDPAWPHLQJVYELLLRFVSSSETDA 10 37 VEALPAFKDVPNSEKQNLFISKLNLCCVVFDFSDPTK 11 29 ETFTLLLALKVRYPDRITJLRGNHESRQI 12 41 WGVSPRGAGYLFGGDVVSQFNHTNNLDLICRAHQLVMEGYK 13 27 WFQDKGIVTVWSAPNYCYRCGNVAAIL 14 38 SHADLDRQIEQLKECKPLPEAEVKVLCDKAKEILVEES 15 50 AHAHDFNINSISNNSDGETFISADDLRINLWNLEISNQCFNIIDMKPANM 16 50 YKTEFQSHEPEFDYLKSLEIEEKINKIRWCQTQNGALFLLSSNDKTIKFW 17 50 MDSGPVATFKVHENLRPKLCELYENDSIFDKFECCJSGDGJHFATGSYSN 18 38 EVITSAEFHPIHCNLLAYSSSRGFIRLIDMRCSALCDQ 19 17 TMIKQILSKLPRKPSKS 20 50 PLEWKFSQVFGERPAGEEVQEVDIISAIEFDKSGDHLAVGDRGGRVVJFE The conserved domains of JrPP2A proteins further confirmed the sub-classifications that were presented by the motifs. As shown in Fig. 3c, the different subunits of JrPP2As were obviously varied, the A sub-family members covered the 'HEAT superfamily' domain, the B sub-family members covered the 'B56 superfamily' domain, the C sub-family members covered the 'MPP superfamily' domain, the B55 sub-family members covered 'CDC55 superfamily' domain, the B'' sub-family members covered 'PPP2R superfamily' domain.
The cis-acting regulatory elements in JrPP2As' promoters
-
To analyze the potential transcriptional regulation of JrPP2A genes in abiotic stress response, putative cis-acting elements in JrPP2A promoters were identified. As a result, a total of 55 cis-acting elements in the promoters were identified and the JrPP2A genes in the same subfamily possessed similar cis-elements. These elements belonged to four types: hormone responsiveness, light responsiveness, abiotic stress response, plant growth, and development (Supplemental Table S3). The elements in 'abiotic stress response' and 'light-responsive' were abundant with 15 and 22 hits (Supplemental Tables S4 & S5), accordingly. The elements Box 4 and G box (light responsive related), MYC (drought and ABA responsiveness related), and MYB (drought-inducible) could be found in most JrPP2As' promoters. In addition, 70% of the JrPP2A genes covered ABA-responsive element (ABRE), suggesting that JrPP2As may be associated with the regulation of the ABA pathway. Meanwhile, some elements such as CAT-box and O2 were predicted to be involved in plant growth and development (Supplemental Tables S3−S5). These results implied that the walnut PP2A genes may play abundant functions in plant growth, development and response to abiotic stresses.
Expression patterns of JrPP2A genes under osmotic stresses and ABA
-
To validate the possible functions of JrPP2A genes in osmotic stress response and whether involving in ABA signaling, the transcript abundance of 15 selected JrPP2A genes were analyzed under drought (PEG6000), salt (NaCl), calcium (CaCl2) and ABA treatment.
Under PEG6000 treatment
-
The response of JrPP2A genes to drought stress was different. JrPP2A15 was up-regulated to the highest level while JrPP2A01 was the lowest one. JrPP2A15 was induced to 1.12-~27.86-fold of other genes. JrPP2A14 was also up-regulated to exceed 4.00 and ranked just after JrPP2A15. The expression value of JrPP2A12, JrPP2A04, and JrPP2A03 exceeded 3.00, while the transcription level of JrPP2A02, JrPP2A05, and JrPP2A10 was 1.54~1.68. The other five (JrPP2A06, JrPP2A08, JrPP2A09, JrPP2A11, JrPP2A13) genes differed not obviously with a level between 2.26 and 2.85 (Fig. 4a).
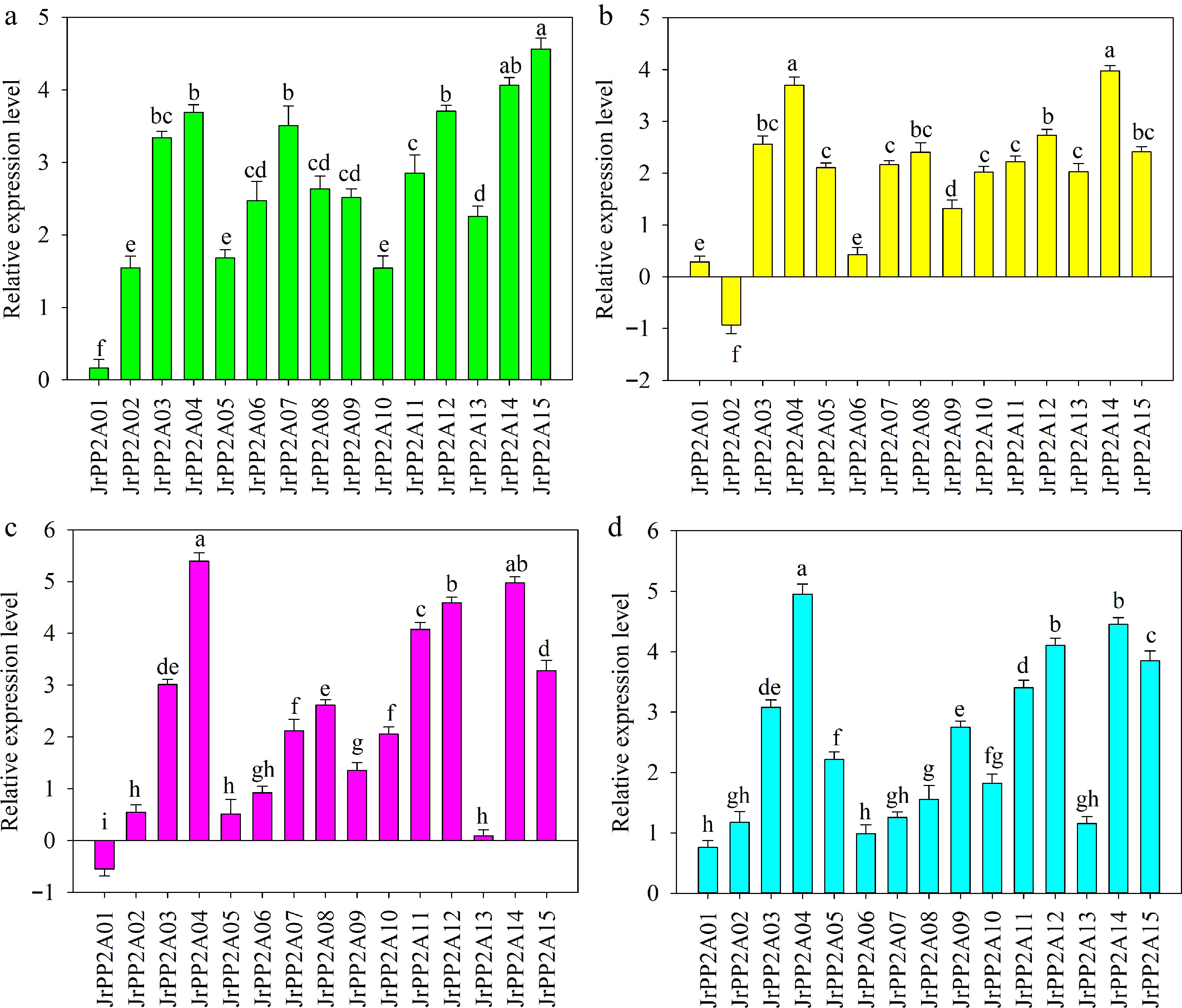
Figure 4.
QRT-PCR analysis of the expression of JrPP2A genes under drought, NaCl, CaCl2, and ABA treatments. The relative expression level is expressed as relative to the internal reference gene and at 0 h. Error bars represent the SD (n = 3). Lowercase letters indicate significant differences among different JrPP2A genes under each treatment according to the Student's t-test (p < 0.05). (a) PEG6000 stress. (b) NaCl stress. (c) CaCl2 treatment. (d) ABA treatment.
Response to NaCl stress
-
Most of the JrPP2A genes except JrPP2A02 were induced by NaCl with varied levels. The expression level of JrPP2A14 was the maximum while not significantly different from the second gene JrPP2A04. The transcription values of JrPP2A12, JrPP2A03, JrPP2A08, and JrPP2A15 were similar and ranged from 2.74 to 2.42. The expression levels of JrPP2A05, JrPP2A07, JrPP2A10, JrPP2A11, and JrPP2A13 were changed not significantly, their values were 2.01~2.22. The expression value of JrPP2A09 (1.32) was significantly lower than the above 10 up-regulated genes while higher than the other three (JrPP2A01, JrPP2A02, and JrPP2A06). Meanwhile, JrPP2A01 and JrPP2A06 displayed similar expression levels (Fig. 4b).
Under CaCl2 treatment
-
Most JrPP2A genes excluding JrPP2A01 were induced by CaCl2 with a large range (0.55~5.39). The expressions of JrPP2A02, JrPP2A05, JrPP2A06, and JrPP2A13 were at a level less than 1.00. JrPP2A09, JrPP2A07, JrPP2A08, and JrPP2A10 were up-regulated to 2.06~2.62, in which JrPP2A08 was significantly higher than JrPP2A07 and JrPP2A10. JrPP2A03 and JrPP2A15 were similar with the expression values 3.01 and 3.28, accordingly. Meanwhile, the difference between JrPP2A03 and JrPP2A08 was not significant. The other four, JrPP2A04, JrPP2A11, JrPP2A12, and JrPP2A14, was showed most obviously transcription and exceed 4.00 (Fig. 4c).
Under ABA treatment
-
All JrPP2A genes were induced by ABA. JrPP2A04 was transcribed highest and significantly differed from all others. The expression levels of JrPP2A14 and JrPP2A12 also exceeded 4.00 and ranked second and third, accordingly, with a non-significant difference. JrPP2A15, JrPP2A11 and JrPP2A03 were up-regulated to a level ranging from 3 to 4. The expression values of JrPP2A09 and JrPP2A05 were 2.22~2.75 with significant difference. JrPP2A01 and JrPP2A06 were transcribed with a level lower than 1.00. The other five genes (JrPP2A02, JrPP2A07, JrPP2A08, JrPP2A10 and JrPP2A13) were expressed in the level between 1.16~1.82 (Fig. 4d).
Overexpression of JrPP2As promote osmotic stress tolerance involving in Ca and ABA signal
-
To confirm the roles of JrPP2As in osmotic stress response, three members, JrPP2A07, JrPP2A9, and JrPP2A14, were chosen according to classification and expression under PEG6000, NaCl, CaCl2, and ABA treatments. JrPP2A07, JrPP2A9, and JrPP2A14 were separately overexpressed in A. thaliana. The transgenic lines with the highest expression value (160.89-, 112.20-, 101.83-fold of WT, accordingly) were selected for analysis (Supplemental Fig. S1a). The results showed that the 42-d old of WT and transgenic seedlings were grown normally without ROS generation under control condition, however, when exposed to PEG6000 and NaCl stress, NBT straining (represent O2−) of WT was deeper than that of transgenic lines (Fig. 5a). Quantitative determination of H2O2 also revealed a similar pattern as NBT staining (Supplemental Fig. S2a). The total ROS content of WT was 1.49-~1.68-fold and 1.37-~1.48-fold of JrPP2A07, JrPP2A9, and JrPP2A14 transgenic lines under PEG6000 and NaCl stress, accordingly (Fig. 5b). Moreover, the cell damage reflected by electrolyte leakage (EL) rate was similar to ROS accumulation. The EL rate of WT was 1.53-~1.91-fold and 1.57-~1.73-fold of JrPP2A07, JrPP2A9, and JrPP2A14 transgenic lines under PEG6000 and NaCl stress, accordingly (Supplemental Fig. S1b). However, the changes of catalase (CAT) activity were opposite to that of ROS content and EL rate (Supplemental Fig. S1c). These results suggested that JrPP2A07, JrPP2A9, JrPP2A14 could positively improve plant drought and salt inducing osmotic stress tolerance.
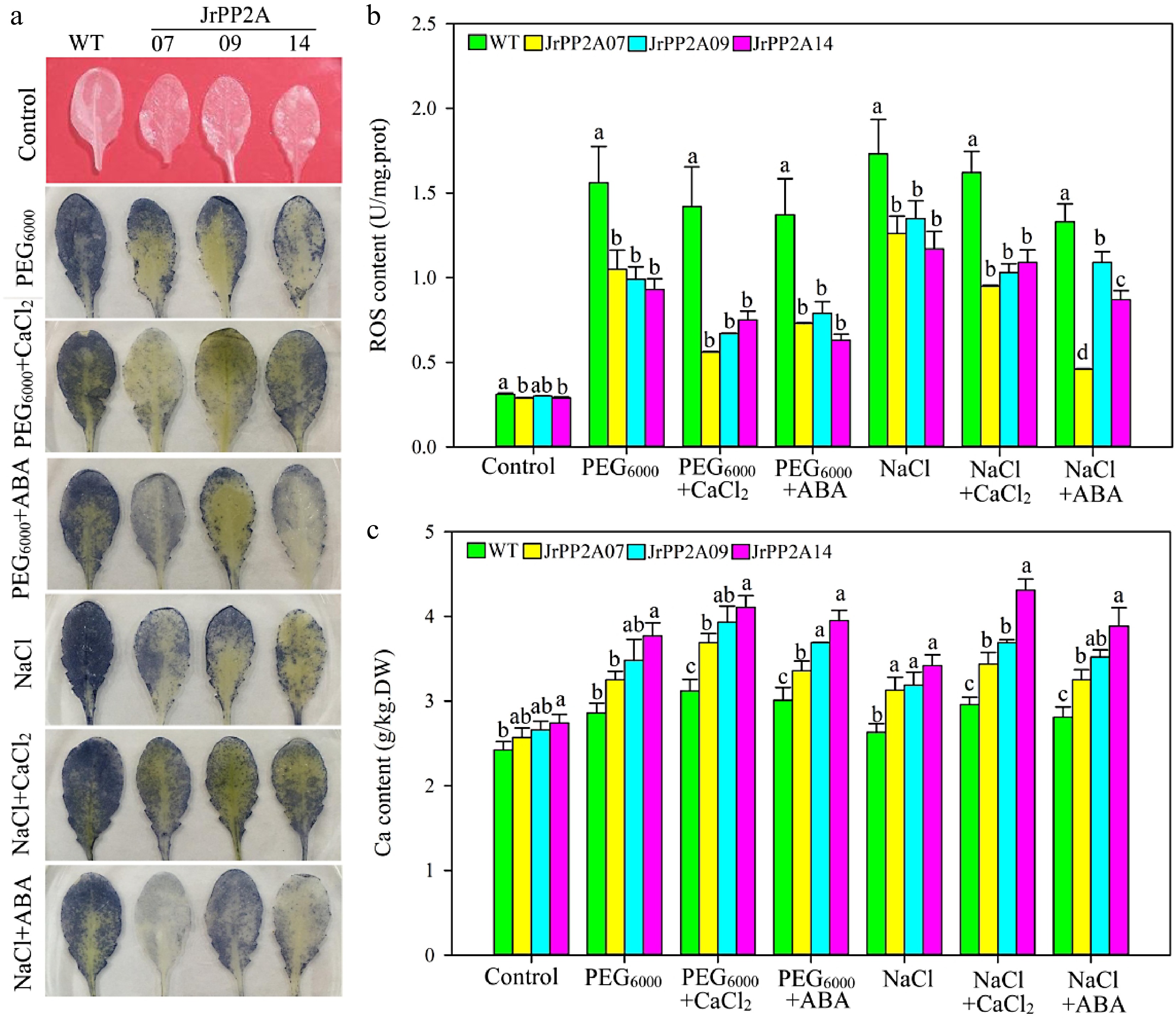
Figure 5.
Osmotic stress response function of JrPP2As. WT, wild type. JrPP2A07, 09, 14, the transgenic lines overexpression of JrPP2A07, JrPP2A09, JrPP2A14. Fourty two-day old seedlings were treated with PEG6000, PEG6000 + CaCl2, PEG6000 + ABA, NaCl, NaCl + CaCl2, NaCl + ABA for 5 d. Control was normally watered. The significant differences among WT, JrPP2A07, JrPP2A09, and JrPP2A14 were marked with lowercase (p < 0.05). (a) NBT staining. (b) Total ROS content. (c) Total Ca content.
Considering the up-regulating expression by CaCl2 and ABA, we want to know whether the JrPP2A genes are associated with Ca and ABA signaling when responding to osmotic stress. Therefore, the 42-day old seedlings were also treated simultaneously with PEG6000 and CaCl2, PEG6000 and ABA, NaCl and CaCl2, NaCl and ABA. Interestingly, we observed that compared to pure PEG6000 and NaCl treatments, the addition of CaCl2 and ABA resulted in changes in resistance physiological indicators of JrPP2As' transgenic plants, suggesting further improvement in osmotic stress resistance (Fig. 5a & b, Supplemental Fig. S1b & S1c). The NBT staining, H2O2 content and total ROS accumulation under PEG6000 + CaCl2 and PEG6000 + ABA was less than under PEG6000, under NaCl + CaCl2 and NaCl + ABA was less than under NaCl, accordingly (Fig. 5a & b, Supplemental Fig. S2a). The cell damage was alleviated after adding CdCl2 and ABA. The EL rate of transgenic lines under NaCl + ABA was 52.56%~84.00% of that under NaCl (Supplemental Fig. S1b), while the CAT activity was increased by adding CdCl2 and ABA (Supplemental Fig. S1c). Moreover, the Ca uptake in JrPP2A transgenic plant cells was improved compared to WT. Under control conditions, the Ca content in transgenic plants' cells was 1.06-~1.13-fold of WT. When exposed to PEG6000 and NaCl, the degree of Ca increases in transgenic plants was also higher than that of WT. The Ca content of WT and transgenic lines under PEG6000 stress were 1.18-~1.38-fold of themselves under control. Adding CaCl2 and ABA significantly improved the Ca accumulation compared to a single treatment of PEG6000 and NaCl (Fig.5c), further confirmation of the relationship of Ca with osmotic stress in JrPP2A genes' response pathway.
Interactions between JrVIP1 protein and JrPP2As
-
JrVIP1 is a basic leucine zipper protein (bZIP) and homologous to the VirE2-INTERACTING PROTEIN1 (VIP1) from A. thaliana (Supplemental Fig. S3), whose dephosphorylation can be mediated by PP2A in osmotic and other stresses[11, 23]. In our previous study, to analyze the relationship between walnut response to drought and demethylation, we constructed BD-JrVIP1, which was used to screen the possible interacting proteins from the walnut yeast two hybrid library, and found that there were many JrPP2A members. To very whether the biological function of the JrPP2A family members was associated with JrVIP1, in this study, AD-JrPP2As were constructed and submitted to the Y2H system to confirm the interaction with BD-JrVIP1. The results showed that JrPP2A02, JrPP2A05, JrPP2A07, JrPP2A09, and JrPP2A14 could interact with JrVIP1, respectively (Fig. 6a). The interactions were also tested by AD-JrVIP1 and BD-JrPP2As (Fig. 6b). Moreover, JrPP2A07, JrPP2A09, and JrPP2A14 were selected to further verify their interactions with JrVIP1 by an in vitro pull-down assay (Fig. 6c). These data confirmed that JrVIP1 could interact with JrPP2A07, JrPP2A09, and JrPP2A14. Since VIP1 homologous were believed to be a vital protein for phosphorylation or dephosphorylation in stress response[11, 23], we are more convinced that the JrPP2A genes are related to stress response involving in phosphorylation.
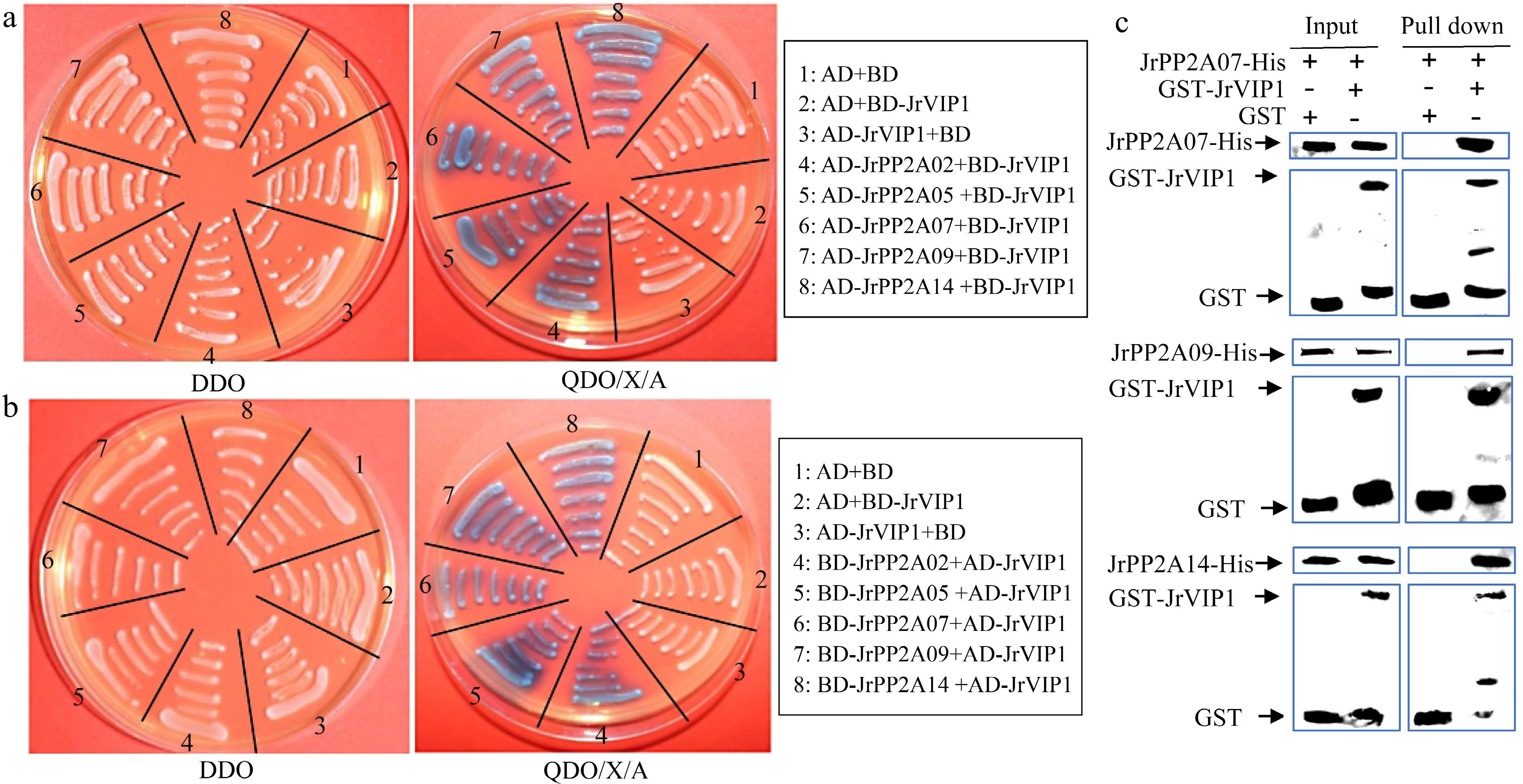
Figure 6.
Interaction analysis of JrVIP1 and JrPP2A proteins using yeast two-hybrid (Y2H) and in vitro pull-down assay. AD+BD, AD+BD-JrVIP1, AD-JrVIP1+BD, negative control. The DDO plate was used as positive control for growth. (a) JrVIP1 was used as the bait. (b) JrVIP1 was used as the pray. (c) An in vitro pull-down assay demonstrates the interaction between JrVIP1 with JrPP2A07, JrPP2A09, JrPP2A14. JrPP2A-His protein was incubated with immobilized GST or GST-JrVIP1 protein, and immunoprecipitated fractions were detected by anti-His antibody. The assay was performed three times with the same result.
-
The plant PP2A gene has been regarded as important members in response to various external stimulus[24, 25]. To provide scientific guidance in the cultivation and management of walnut trees and then to guarantee the yield and quality of walnuts under adverse conditions, the molecular mechanism of adaptation to adversity is urgently to be revealed. Many environmental factors may lead to osmotic stress, so the regulation of osmotic stress response is particularly noteworthy. Therefore, the walnut transcriptomes under different abiotic stresses were sequenced to establish a basic database for identifying key genes that adapt or respond to environmental stresses. In the current study, 15 JrPP2A genes with prominent expression levels that can respond to various stressors were identified from the transcriptome data mentioned above. This quantity, 15 JrPP2A genes, is less than the number of members of the PP2A family in other plants such as Arabidopsis and rubber trees. The possible reason is that the JrPP2A family genes selected in this study are shared genes screened from transcriptomes under various stresses, and mainly transcriptionally expressed genes that respond to osmotic stress. It cannot be guaranteed that all members can be induced to express under stress, nor can these genes be screened in the transcriptome database. Walnuts are likely to have other PP2As that may not be clear about their response to osmotic stress. In terms of evolutionary relationship, the 15 JrPP2A proteins shared a high similarity with the PP2As from other species, such as A. thaliana[11, 23], O. sativ[26], and H. brasiliensis[13] and grouped into five subfamilies (Fig. 1, Table 1). The ORF length, pI, amino acid number, and molecular weight of JrPP2As were similar to those from Arabidopsis and H. brasiliensis[11,13,27], confirming that these 15 JrPP2As belong to PP2A protein family.
To understand the biological functions of PP2A family members, originally, both intron/exon organizations and protein motif patterns were analyzed, and found that JrPP2As displayed diverse gene and protein structures. The intron/exon structure is an important pathway for gene functional evolution and the main reason for functional differences with its homologous proteins[28]. Despite the conserved distribution pattern of exons in subgroups B and C, many other subgroups displayed diversity in exon structure and number, which was consistent with the protein pattern (Fig. 3). Most of B' subunit type JrPP2As contained motifs of 19, 10, 7, 9, 2, 1, 3, 4, and 6, which were related to B56 domain[7, 13, 24]; while JrPP2ACs were related to MPP domain[7, 24] (Fig. 3c). Because different motifs and the number of motifs were related to functions[25−27], our results suggested that JrPP2As have potential abundant roles in walnut. Next, given that plant promoter cis-acting elements perform essential roles in regulating gene expression and may imply metabolic pathways and stress response[29,30], the cis-acting elements in JrPP2As promoters were detected. We screened abundant cis-elements related to drought and salt stress response (Supplemental Tables S3−S5), such as MYC elements, W-box, MBS elements, MYB recognition site, TCA-element. The results preliminarily suggested that these JrPP2As may be involved in stress responses such as drought and salt regulated by the upstream promoter elements.
Consequently, the response patterns of 15 JrPP2A genes exposed to PEG6000 and NaCl were determined and found that most of the JrPP2As were significantly induced by PEG6000 and NaCl. The transcription activity of genes in response to different stresses may effectively predict their potential functions. Arabidopsis PP2AC5 was reported to be induced by drought and salt stresses, when it was overexpressed, the transgenic Arabidopsis was verified to improve the drought and salt stress tolerance[31]. Compared with the loss-of-function mutant pp2a-c5-1, PP2AC5 overexpression lines were conferred with better root and shoot growth under salt treatments[32]. The drought and salt inducible GmPP2A-B"71 could also enhance plant tolerance to drought and salt stresses via overexpression in soybean[33]. A novel Ca2+-binding protein, named AtCP1 (AtPP2A-B"43), can be up-regulated by NaCl treatment, was also believed to positive on slat stress response[34]. The mRNA levels of Solanum tuberosum PP2Ac1, PP2Ac2a, PP2Ac2b and PP2Ac3 in leaves were up-regulated by salt stress, suggesting that the subunits might have vital roles in response to drought and salt stress[35]. These reports and the inducible expression of JrPP2As let us believe that JrPP2A genes are likely to play roles in drought and salt stress response. From the expression level, JrPP2A04, JrPP2A14 and JrPP2A15 were the top three while JrPP2A01 and JrPP2A02 were the bottom two those may deserve further attention (Fig. 4a−c).
To confirm the functions of JrPP2A genes in drought and salt-inducing osmotic stress response, JrPP2A07, JrPP2A09, and JrPP2A14 were independently overexpressed in A. thaliana and found that the transgenic plants showed less ROS accumulation, and lighter cell damage while higher antioxidant enzyme activity than those of WT under PEG6000 and NaCl stress (Supplemental Figs S1 & S2a, Fig. 5), determining that JrPP2A07, JrPP2A09, and JrPP2A14 are positive genes in walnut osmotic stress tolerance. Additionally, the process that plant PP2A responding to osmotic stress is implicated in ABA signaling[36]. In this study, the transcription of JrPP2As under ABA treatment was tested and showed that B subunit JrPP2A genes were induced obviously by ABA, especially JrPP2A04. In other species, such as wheat, TaPP2AB-α, a novel B subunit of PP2A, was induced by the response to NaCl, PEG6000, cold, and ABA at the transcriptional level[37]. Transgenic Arabidopsis overexpression of TaPP2AB-α displayed more lateral roots under mannitol or NaCl treatment[37]. These results indicated that TaPP2AB-α could promote plant lateral root growth under osmotic conditions[37]. TIP41, an interactor of PP2A present in Arabidopsis, was induced by long-term NaCl, polyethylene glycol and ABA treatments, proving that TIP41 mediates the participation of PP2A in ABA-mediated mechanisms[38]. The mRNA levels of A. thaliana PP2AA3 in roots and shoots were up and down-regulated by drought and ABA treatments, suggesting that the subunits might have vital roles in response to drought and ABA treatments[39]. Therefore, we believed that JrPP2As in response to osmotic stress involving in ABA signaling.
Under external stimulation, Arabidopsis PP2A protein stimulates the Ca accumulation or transport within cells and in turn activates B'' and C class PP2A proteins[11, 23]. To investigate whether the JrPP2A family genes are involved in calcium signaling in osmotic stress response, we also invested the transcription activity of the JrPP2A genes under the condition of adding exogenous CaCl2. We were surprised to find that members of JrPP2A genes were significantly induced by CdCl2 (Fig. 4d). Overexpression of JrPP2A07, JrPP2A09, and JrPP2A14 in Arabidopsis promoted the Ca accumulation within plant cells under PEG6000 and NaCl stress. Moreover, under CaCl2 mediation conditions, the ROS accumulation and cell damage were reduced, while antioxidant protection was enhanced, indicating the effective improvement of the plant osmotic stress tolerance with increasing Ca uptake (Supplemental Figs S1 & S2a, Fig. 5). These performances confirmed the positive mediation of Ca in JrPP2As' abiotic stress response. The activation of B'' and C subunits of PP2A proteins mediates the dephosphorylation of VIP1[11, 23]. Hypo-osmotic stress regulates the dephosphorylation and nuclear-localization of VIP1. When cells confronted with mechanical stress, VIP1 was transiently accumulated in the nucleus, where it controls the expression of the target genes and inhibits mechanical stress-induced root waving[11]. In this process, Ca signal was accompanied to play necessary roles[23]. These reports told that the interaction between PP2A and VIP is an important pathway for osmotic stress response. In walnut tree, we also discovered that JrVIP1, a homology of Arabidopsis VIP1, could interact with B'' subunit type JrPP2A02 and JrPP2A09 as well as C subunit type JrPP2A05, JrPP2A07 and JrPP2A14 (Fig. 6). Moreover, JrVIP1 was also up-regulated by osmotic stress (Supplemental Fig. S2b & S2c). Therefore, we believe that the JrPP2A genes also mediate VIP1 dephosphorylation in response to osmotic stress and are involved in Ca signaling. Meanwhile, an osmotic stress responsive pathway mediated by JrVIP1 and JrPP2As was summarized as Fig. 7.
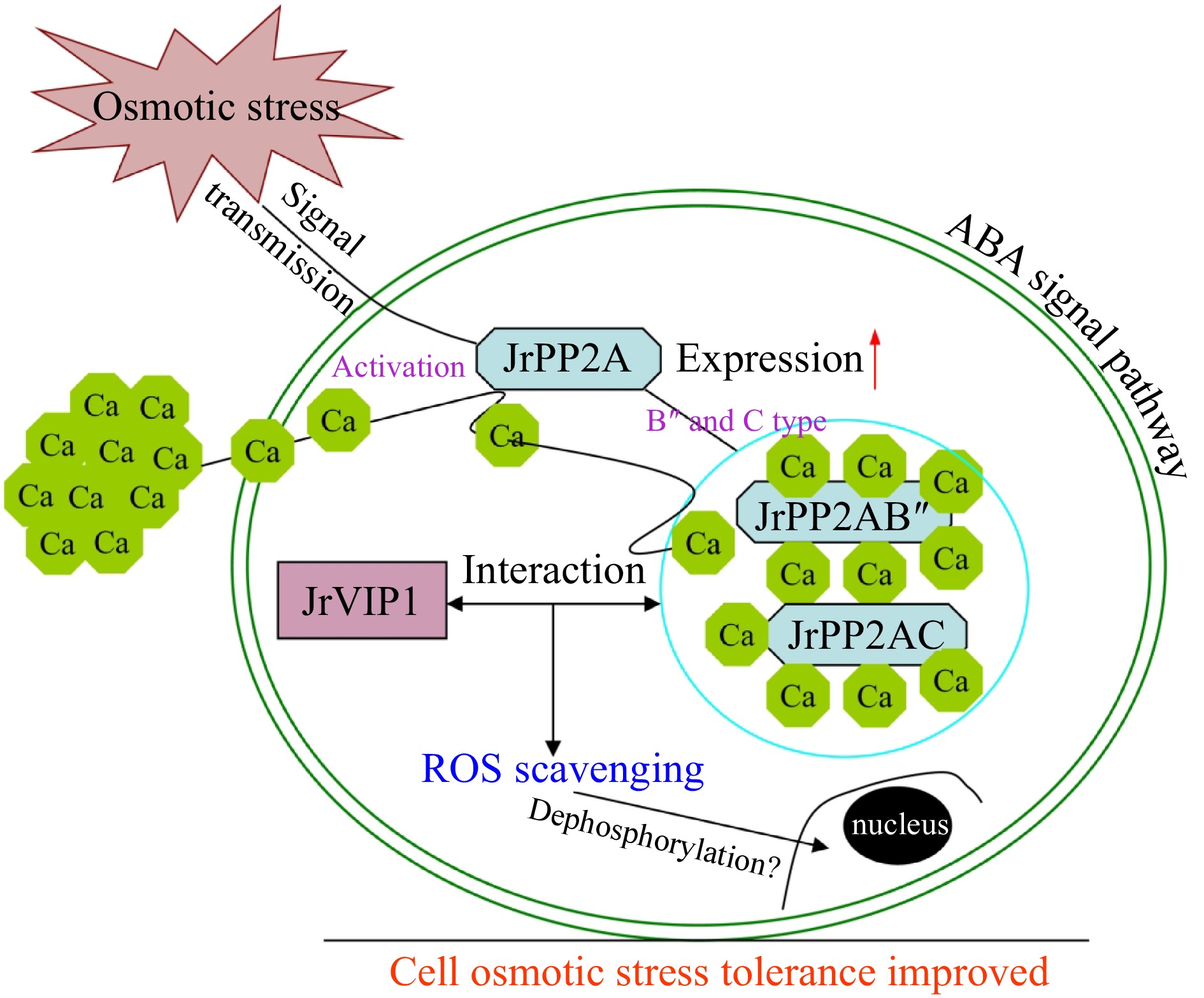
Figure 7.
The JrVIP1 and JrPP2As mediated osmotic stress responsive pathway in walnut trees.
-
In this study, we identified the PP2A family genes from the walnut transcriptome and a total of 15 JrPP2As were screened to be unevenly distributed on 10 of the walnut chromosomes. The JrPP2As genes were grouped into five subfamilies and members of the same subfamily shared similar gene structures and conserved protein motifs. Promoter element compositions imply that JrPP2As may be involved in hormone, light, growth, development, and abiotic stress responses. Most of the JrPP2A genes exhibited various expression levels to drought and salt-inducing osmotic stress, among which JrPP2A07, JrPP2A09, and JrPP2A14 were improved to play positive roles in osmotic stress response involving Ca and ABA signaling. Moreover, B'' and C types of JrPP2A proteins mediate the dephosphorylation of JrVIP1 in osmotic stress response. Our findings generate novel insights into PP2A family genes in walnut and lay a foundation for further understanding their biological functions.
-
The authors confirm contribution to the paper as follows: study conception and design: He Y, Yang G; performing experiments: He Y, Chen S, Li C, Yang S; data analysis: Li C, Yang S, Sun Z, Hou S; draft manuscript preparation: He Y, Yang G; data analysis checking and manuscript revision: Yang G, Xu Z. All authors reviewed the results and approved the final version of the manuscript.
-
All the data were presented in the main manuscript and additional supporting files. The Arabidopsis and H. brasiliensis related datasets generated and/or analyzed during the current study are available in the TAIR database (
www.arabidopsis.org ) and NCBI (Hevea brasiliensis (ID 503)-Genome-NCBI (nih.gov)). This work was supported by Chinese Universities Scientific Fund (2452022117); National Natural Science Foundation of China (32171804, 31800510); Experimental Demonstration Station (Base) Science and Technology Innovation and Achievement Transformation Project from Northwest Agriculture and Forestry University (TGZX2021-41). The funding agency was not involved in the design of the study, collection, analysis, interpretation of data or writing the manuscript.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yi He, Shuwen Chen
- Supplemental Table S1 The primers used for qRT-PCR analysis.
- Supplemental Table S2 The primers used for construction of overexpression vectors and interaction related recombinants.
- Supplemental Table S3 The cis-acting elements of 15 JrPP2A promoters.
- Supplemental Table S4 Distribution of the number of cis-acting elements of 15 JrPP2As.
- Supplemental Table S5 The number of cis-elements in the JrPP2A promoters.
- Supplemental Fig. S1 The expression, EL rate and CAT activity of JrPP2A07, JrPP2A09, JrPP2A14 transgenic lines. A, The expression level of JrPP2A07, JrPP2A09, JrPP2A14 in transgenic lines. B-C, the EL rate and CAT activity under the treatments of PEG6000, NaCl, PEG6000+CaCl2, PEG6000+ABA, NaCl, NaCl+ CaCl2, NaCl+ ABA for 3 days, which was consistent with figure 5. The significant differences among WT, JrPP2A07, JrPP2A09 and JrPP2A14 were marked with lowercase (p < 0.05).
- Supplemental Fig. S2 The generated H2O2 contents of JrPP2A transgenic plants under osmotic stress same as Fig. 5 (A) and the expression patterns of JrVIP1 under PEG6000 (B) as well as NaCl (C) stress. The significant differences among WT, JrPP2A07, JrPP2A09 and JrPP2A14 were marked with lowercase (p < 0.05) in A. * in B and C means the differences between the treatment time points and 0 h (control) was significant (p < 0.05). For the stresses, 20% (w/v) PEG6000 and 0.3 mol/L NaCl was independently watered to the 3-year-old 'Xiangling' walnut for 3 days, and the leaves were sampled at 0 (Control), 3, 12, 24, 48, 72 h.
- Supplemental Fig. S3 Similarity and conservative domain analysis of JrVIP1. A, Alignment of JrVIP1 and AtVIP1. B, Similarity analysis using online blast in NCBI. C, The conservative domains existed in JrVIP1 and AtVIP1.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
He Y, Chen S, Li C, Yang S, Sun Z, et al. 2024. Walnut phosphatase 2A proteins interact with basic leucine zipper protein JrVIP1 to regulate osmotic stress response via calcium signaling. Forestry Research 4: e016 doi: 10.48130/forres-0024-0012 

Walnut phosphatase 2A proteins interact with basic leucine zipper protein JrVIP1 to regulate osmotic stress response via calcium signaling
- Received: 28 November 2023
- Revised: 21 March 2024
- Accepted: 07 April 2024
- Published online: 06 May 2024
Abstract: Walnut is an important economic tree species that is susceptible to osmotic stress. Scientific cultivation management is an important way to improve the yield and quality of walnuts, which requires understanding the regulatory mechanisms in response to osmotic stress. Therefore, in this study, 15 protein phosphatase 2A (PP2A) genes were identified from the walnut transcriptome (named JrPP2A01~15) and their potential function responses to osmotic stress were elucidated. The open reading frame (ORF) of JrPP2A01~15 ranges from 651 to 1,764 bp in length, the molecular weight of the encoded proteins are 24.15−65.61 kDa, and the theoretical isoelectric points are 4.80−8.37. These JrPP2As were unevenly distributed on 10 chromosomes and divided into five groups based on the composition of conserved domains, motifs, and exon/intron organizations. The five groups are JrPP2AAs, JrPP2AB's, JrPP2AB''s, JrPP2AB55s, and JrPP2ACs, including 1, 5, 2, 3, and 4 members, accordingly. The cis-elements in JrPP2As' promoters were involved in responses to hormone and abiotic stress. Most JrPP2A genes, excluding JrPP2A01, JrPP2A02, JrPP2A05, JrPP2A06, and JrPP2A13, could be induced significantly by PEG6000, NaCl, CaCl2 and ABA. JrPP2A02, JrPP2A05, JrPP2A07, JrPP2A09, and JrPP2A14, could independently interact with a bZIP transcription factor JrVIP1. Moreover, overexpression of JrPP2A07, JrPP2A09, and JrPP2A14 could significantly decrease ROS accumulation while increasing calcium (Ca) uptake exposed to PEG6000 and NaCl stresses, which was mediated by exogenous CaCl2 and ABA. These results suggested that JrPP2A genes play potential key roles in walnut response to drought and salt-inducing osmotic stress involving Ca- and ABA-dependent signaling pathways.
-
Key words:
- Juglans regia /
- Abiotic stress /
- Expression analysis /
- Protein interaction /
- ROS signal