









-
Drought, as an abiotic stress, is a primary environmental factor that restricts plant growth and development. Under drought stress conditions, plants usually show a wilting phenotype and typically experience reduced growth rates, leaf area, and number, along with decreased photosynthetic rates. Concurrently, there is an accumulation of reactive oxygen species (ROS) in plants which worsens lipid peroxidation of cell membranes and leads to cellular damage. To adjust to the drought environments, plants will physiologically increase the activity of antioxidant enzymes and the abundance of osmotic regulation substance[1]. At the molecular level, plants response to drought stress is often related to regulating the transcriptional expression of stress defense genes[2].
Gene expression is usually regulated by the interaction between cis-acting elements and transcription factors, and the latter usually requires some cofactors to jointly regulate gene expression. Valine-glutamine (VQ) protein is a special transcription cofactor that is widely found in plants and contains the highly conserved VQ motif FxxhVQxhTG[3]. In 2002, Morikawa et al. discovered VQ protein in plants for the first time and named it SIB1 (SIGMA FACTOR-BINDING PROTEIN1)[4]. Subsequently, VQ genes have been identified in various plant species, such as rice, maize, grape, and so on[5−7]. The expression of many VQ genes changed significantly when exposed to environmental stress, pathogen invasion, and plant hormone treatment[5,6,8]. These indicated they were involved in responses to different biological or abiotic stresses. Extensive evidence suggests that VQ proteins participate in these processes through interactions with proteins such as WRKY transcription factor (WRKY) and mitogen-activated protein kinase (MAPK). In this way, they regulated plant growth and development in response to environmental stresses[9−12].
Some progress has been made in how VQ proteins function in the response to drought stress. In Ipomoea batatas, the IbWRKY2 gene positively regulated drought tolerance. IbVQ4 could interact with IbWRKY2, and PEG treatment remarkably induced IbVQ4 expression. This suggested that IbVQ4 might helpfully assist transcription factors to respond to drought[13]. A recent study showed that the overexpression of TaVQ4-D significantly improved the tolerance of Triticum aestivum plants to drought stress. Compared with wild type, the levels of superoxide dismutase and proline in transgenic plants increased, the levels of malondialdehyde decreased, and the expression of drought-responsive genes was up-regulated. In addition, TaVQ4-D interacted with two mitogen-activated protein kinases (MPK3 & MPK6), and as their phosphorylated substrates to response stress[14].
The VQ motif, the most prominent feature of the VQ family proteins, plays an important role in the performance of these functions. Firstly, the VQ motif affected the function of VQ proteins in regulating growth and development. Mutating IVQQ to EDLE in the VQ motif of AtIKU1 resulted in smaller seeds of Arabidopsis thaliana, while mutating other locations did not show phenotypic changes[15]. Secondly, VQ motifs could affect the subcellular localization of VQ proteins. The fusion protein ATVQ9-GFP was primarily localized in the nucleus. However, when VVQK within the VQ domain was mutated to EDLE, its subcellular localization altered and it became distributed in both the nucleus and cytoplasm[16]. In addition, the VQ motif played a crucial role in influencing interactions between VQ proteins and other proteins. In the absence of the VQ motif, the interaction between AtVQ20 and AtWRKY34/AtWRKY2 was completely cut off in A. thaliana. Similarly, the substitution of EDLE for LVQK in the VQ motif also prevented any interaction, suggesting that the VQ motif's integrity was crucial for its interaction with WRKY[17]. It was interesting that the VQ motif of AtMKS1 was not only involved in nucleus localization but also influenced its interaction with AtWRKY33 and AtWRKY25 in A. thaliana[18].
In conclusion, VQ proteins are a class of proteins with conserved VQ motif structure. They are widely involved in plant growth and development as well as in the response to various stresses. The functions of VQ genes have been identified and studied in many plants, but the role of stress resistance in the woody plant P. ostii 'Fengdan' is unknown. To clarify the function of VQ, the PoVQ31 gene was cloned, the protein features were analyzed, and the PoVQ31-silenced plants were produced to explore the drought resistance mechanism. This study deepened the understanding of the function of VQ proteins, opened up new horizons in the study of stress tolerance in the woody plant P. ostii 'Fengdan', and provided valuable theoretical support and practical guidance for the improvement of plant stress tolerance.
-
The one-year-old P. ostii 'Fengdan' plants were selected as the experimental material and transplanted in pots with dimensions of 15 cm × 14 cm × 10 cm. The cultivation substrate was a 1:1:1 mixture of garden soil, peat and perlite. The materials were grown under long sunlight conditions (16 h light/8 h dark) with temperature maintained at 25 °C. Collecting leaf tissues for gene cloning and expression pattern analysis after exposure to natural drought conditions for 0, 4, 8, and 12 d. After the VIGS experiment, plants were cultured in the above-growth environment for 30 d, and then drought-treated for 15 d, and the tissue leaves were collected respectively before and after treatment. The 4-week-old Nicotiana benthamiana was employed in subcellular localization observation experiments.
Gene cloning and sequence analysis
-
Total RNA was extracted from P. ostii 'Fengdan' leaves using the FastPure Universal Plant Total RNA Isolation Kit (Vazyme, China) and using the HiScript II Reverse Transcriptase (Vazyme, China) to reverse transcribe RNA into cDNA according to the manufacturer's protocol. The CDS sequence of PoVQ31 (Unigene0032765) was downloaded from the existing P. ostii 'Fengdan' transcriptome database (SRA: SRP161474) to design primers (forward primer: 5'-TGCGAACCTTTTCTCCTG-3'; reverse primer: 5'-CTTCAATTTCAACAGGTGCA-3'). After amplifying the PoVQ31 CDS sequence, recovering and inserting the PCR product into the T-plasmid vector, then transferring the recombinant plasmid into Escherichia coli for sequencing. Though MEGA 11.0 software to construct the phylogenetic tree, which using the neighborhood connection method and 1,000 guided repeats. The conserved motifs of PoVQ31 and its homologous sequences were predicted on the MEME online program (
https://meme-suite.org/meme/ ). Applying ESPript 3.0 website (https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi ) to present multiple sequence comparisons. Conserved motifs logo was drawn using WebLogo 3 website (https://weblogo.threeplusone.com/create.cgi ).Bioinformatics analysis
-
Using the ProtParam and the ProtScale analysis tool on the Expasy website (
www.expasy.org ) to predict protein hydrophilicity/hydrophobicity and the protein physicochemical characteristics of PoVQ31. The DeepTMHMM website (https://dtu.biolib.com/DeepTMHMM ) was used to predict transmembrane structural domains. The SignalP-5.0 website (https://services.healthtech.dtu.dk/services/SignalP-5.0/ ) was used for identifying signal peptides. The NetPhos-3.1 webpage (https://services.healthtech.dtu.dk/services/NetPhos-3.1/ ) was used to forecast phosphorylation sites. Protein secondary and tertiary structures of PoVQ31 were predicted using SOPMA (https://npsa-prabi.ibcp.fr/NPSA/npsa_sopma.html ) and AlphaFold (https://alphafold.ebi.ac.uk/ ).Quantitative real-time PCR (qRT-PCR) analysis
-
RNA was extracted from the P. ostii 'Fengdan' leaf tissues using FastPure Universal Plant Total RNA Isolation Kit (Vazyme, China), and RNA was reversed to cDNA HiScript II Reverse Transcriptase (Vazyme, China). qRT-PCR was carried out on the Bio-Rad CFX96 Real-Time PCR Detection System (Bio-Rad, USA) using the 2×PerfectStart® Fast Green qPCR SuperMix (TransGen, China) according to the manufacturer's instructions. The expression level was calculated base on the 2−ΔΔCᴛ method[19]. The P. ostii Ubiquitin (JN699053) was used as an internal reference gene. Special primer pairs designed for the qRT-PCR are listed in Supplemental Table S1.
Subcellular localization
-
Using primers (forward primer: 5'-CGGGGATCCTCTAGAGTCGACATGGAAAAACAGTACTACAACACCGT-3'; reverse primer: 5'-CACCATGGTACTAGTGTCGACACGAGTTTTCTGACTCGGAGAACT-3') to amplify the CDS sequence of PoVQ31, and the purified amplification product was inserted into the p35S:eGFP vector. The empty vector p35S:eGFP and p35S:PoVQ31-eGFP plasmids were transformed into Agrobacterium strains GV3101 via the freeze-thaw method. The transformed strains were mixed with the nucleus labeled strains containing mCherry protein in a 1:1 ratio. The mixture was injected into the N. benthamiana and incubated in the dark for two days before being observed under laser confocal microscopy (Leica TCS-SP8, Wetzlar, Germany). The excitation light wavelengths of 488 and 561 nm were used to detect the signals of green fluorescent protein (GFP) and red fluorescent protein (RFP), respectively.
Virus-induced gene silencing (VIGS)
-
The tobacco rattle virus (TRV)-based virus-induced gene silencing was utilized to silence PoVQ31 with two vectors, TRV1 and TRV2. After using the following primers (forward primer: 5'-AAGGTTACCGAATTCTCTAGACTTGGAGTTTGAACGGTGACCG-3'; reverse primer: 5'-CGTGAGCTCGGTACCGGATCCATGGAAAAACAGTACTACAACACCGT-3') to amplify a 241 bp PoVQ31 gene-specific fragment and connect it to the TRV2 vector, the vectors of TRV1, TRV2, and TRV2-PoVQ31 were transformed into the Agrobacterium strain GV3101. Suspension of TRV2 and TRV2-PoVQ31 were each mixed in equal proportions with a suspension of TRV1 to prepare the infiltration solution. The P. ostii 'Fengdan' plants with 1~2 buds were washed with sterile water and then infected for 30 min based on vacuum suction filtration method. These plants were transplanted into pots and grown in the greenhouse. Plants were routinely cultured for 30 d before the 15-d drought treatment, and then phenotypic observations and physiological data were tested on the gene-silenced plants. DNA was extracted from drought-treated leaves to identify transgenic plants, and RNA was isolated for qRT-PCR to calculate gene silencing efficiency. All primer sequences are presented in Supplemental Table S1.
Determination of physiological indexes
-
Plant leaves were collected for measuring physiological indices before and after drought. For relative electrical conductivity (REC) measurements, 0.1 g fresh leaves were cut into 10 mL deionized water and left it overnight at room temperature to measure the initial conductivity C1. The sample was then heated in a boiling water bath for 30 min, cooled sufficiently, and C2 measured. REC was calculated as C1/C2 × 100%. For water content, the fresh weight of the leaves (FW) were weighed, then the leaves were placed in the oven at 105 °C, dried out for 15 min, then baked at 80 °C until constant weight, then the dry weight (DW) was weighed. The formula for calculating the water content of leaves is: (FW−DW)/FW × 100%. Diaminobenzidine (DAB) and nitroblue tetrahydroxide (NBT) staining kits were purchased from the Maclin Biochemical Technology (Shanghai, China) and leaf staining was performed according to the instructions. Malondialdehyde (MDA) content and four protective enzyme activities, including peroxidase (POD), superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX), were all measured according to the instructions for the reagent kit.
Statistical analysis
-
All measurements were conducted in triplicate, and the error bars represent the standard deviation. Significant differences were determined by Student's t-test (* p < 0.05, ** p < 0.01, *** p < 0.001).
-
Previous studies have shown that VQ family proteins, as transcriptional regulatory cofactors, can participate in plant response to abiotic stress. Here a VQ protein was identified from the transcriptome, PoVQ31, whose expression was significantly upregulated under drought conditions. First, the full-length of the PoVQ31 gene was cloned. It had an open reading frame (ORF) of 540 bp. The gene encoded a polypeptide containing 179 amino acids with a molecular weight of 19.65 kDa. The phylogenetic tree was constructed using MEGA 11.0 to further explore the evolutionary relationship between PoVQ31 and other VQ proteins. The findings indicated that the VQ31-like protein of Cornus florida, which belonged to the Cornaceae family was the closest relative to PoVQ31 (Fig. 1a). According to multiple sequence comparisons, these VQ proteins were highly conserved and all of them contained a typical VQ structural domain (FxxhVQxhTG). This domain might be essential for the functional access of VQ proteins in these species (Fig. 1b & c).
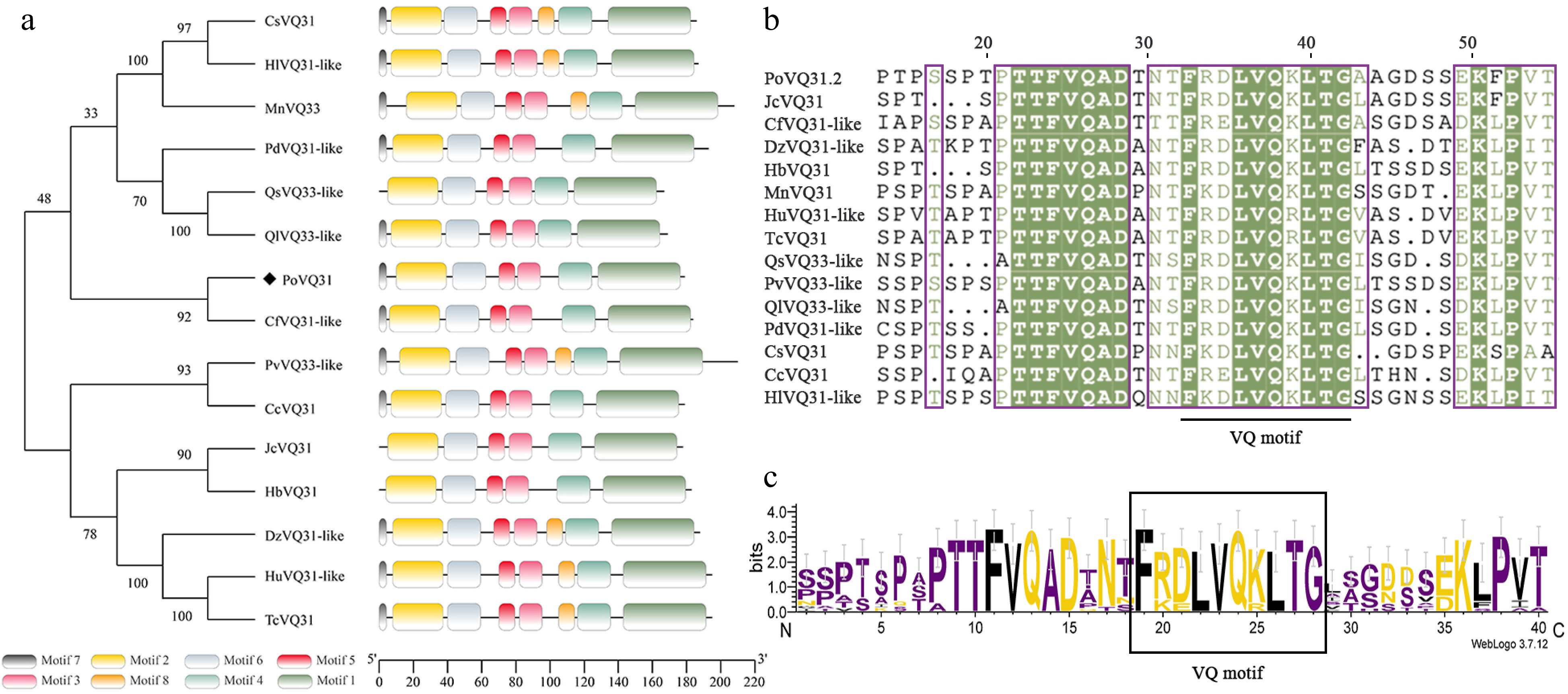
Figure 1.
Phylogenetic analysis and structure characterization of PoVQ31. (a) Phylogenetic tree constructed by Paeonia ostii 'Fengdan' PoVQ31 and other species's VQ proteins. Conserved motifs of VQ proteins, each denoted by a distinct color. (b) Multiple sequence alignment of the PoVQ31. (c) The specific highly conserved motif of VQ protein.
Then the physicochemical properties and protein structure of PoVQ31 were predicted. It was predicted by DeepTMHMM that PoVQ31 was a non-secretory protein as it lacked the signal peptide and the transmembrane structure (Fig. 2a & b). ProtScale prediction showed that the content of the hydrophilic region was higher than that of the hydrophobic region, indicating that PoVQ31 was a hydrophilic protein (Fig. 2c). Based on NetPhos-3.1 prediction, the PoVQ31 protein included 39 phosphorylation sites, of which 26 were for serine, 10 were for threonine, and three were for tyrosine (Fig. 2d). SPOMA predicted that the proportions of the alpha helix, beta sheet, beta turn, and random coil in the secondary structure were 13.97%, 12.29%, 2.23%, and 71.51% (Fig. 2e). SWISS-MODEL was used to predict the protein's tertiary structure. The results showed that the protein had a lot of α-helical and irregularly coiled structures (Fig. 2f). This matched the secondary structure prediction results. In short, PoVQ31 was a hydrophilic non-secretory protein of the VQ gene superfamily.
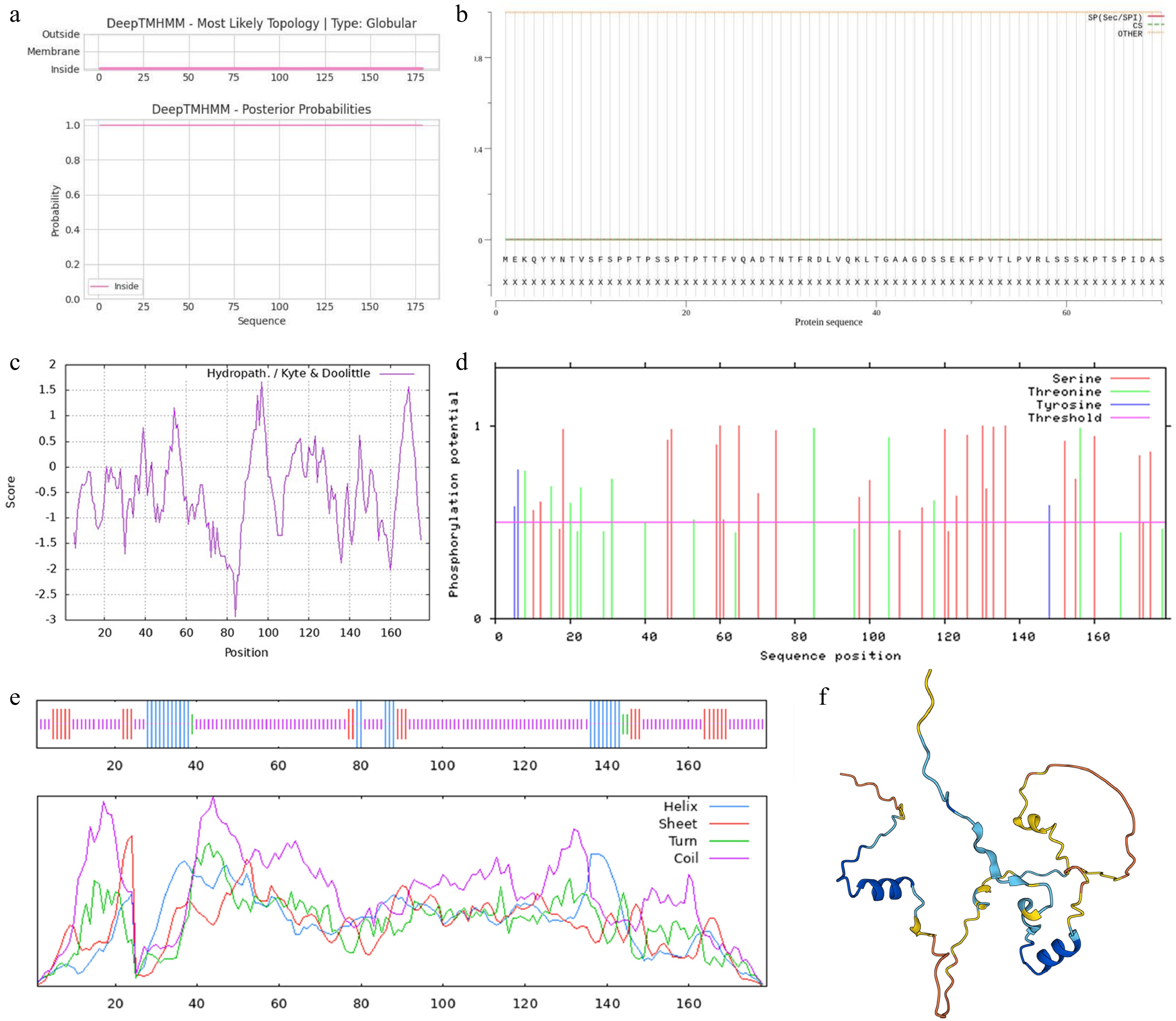
Figure 2.
Bioinformatics analysis of PoVQ31. Prediction of (a) transmembrane structure, (b) signal peptide, (c) hydrophilicity/hydrophobicity, (d) phosphorylation sites, (e) secondary structure, and (f) tertiary structure of PoVQ31 protein.
PoVQ31 is a nucleus-localized protein and positively responds to drought stress
-
To identify the subcellular localization of PoVQ31, the 35S: PoVQ31-eGFP fusion vector and empty eGFP vector were expressed in N. benthamiana. The observation results showed that the green fluorescence signal of the 35S: PoVQ31-eGFP fusion vector merged with the red fluorescence signal of the nucleus localization vector, confirming that PoVQ31 was located in the nucleus (Fig. 3). To study the function of PoVQ31 in response to drought stress, the expression pattern of PoVQ31 at different time points (0, 4, 8, and 12 d) under drought conditions were analyzed. Study findings revealed a significant upregulation of PoVQ31 expression when exposed to drought stress and the expression level constantly increased over time (Fig. 4). Briefly, these results suggested that PoVQ31 was localized in the nucleus and might play a crucial role in regulating drought tolerance in P. ostii 'Fengdan'.
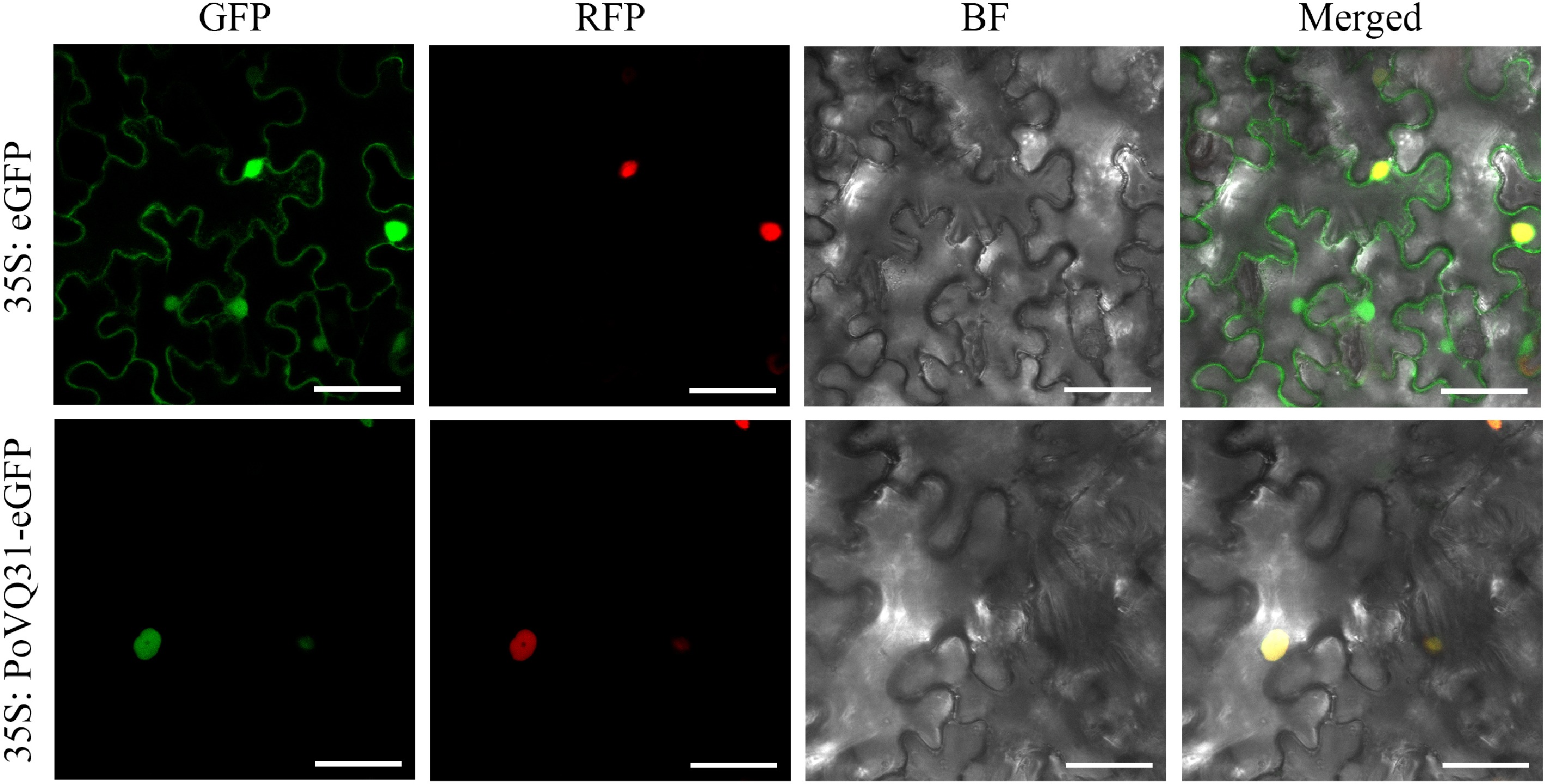
Figure 3.
Subcellular localization of PoVQ31. GFP, green fluorescent protein; RFP, nuclear localization signal exhibited by the red fluorescent protein; BF, bright-field image; merged, overlay images. Scale bars = 50 μm.
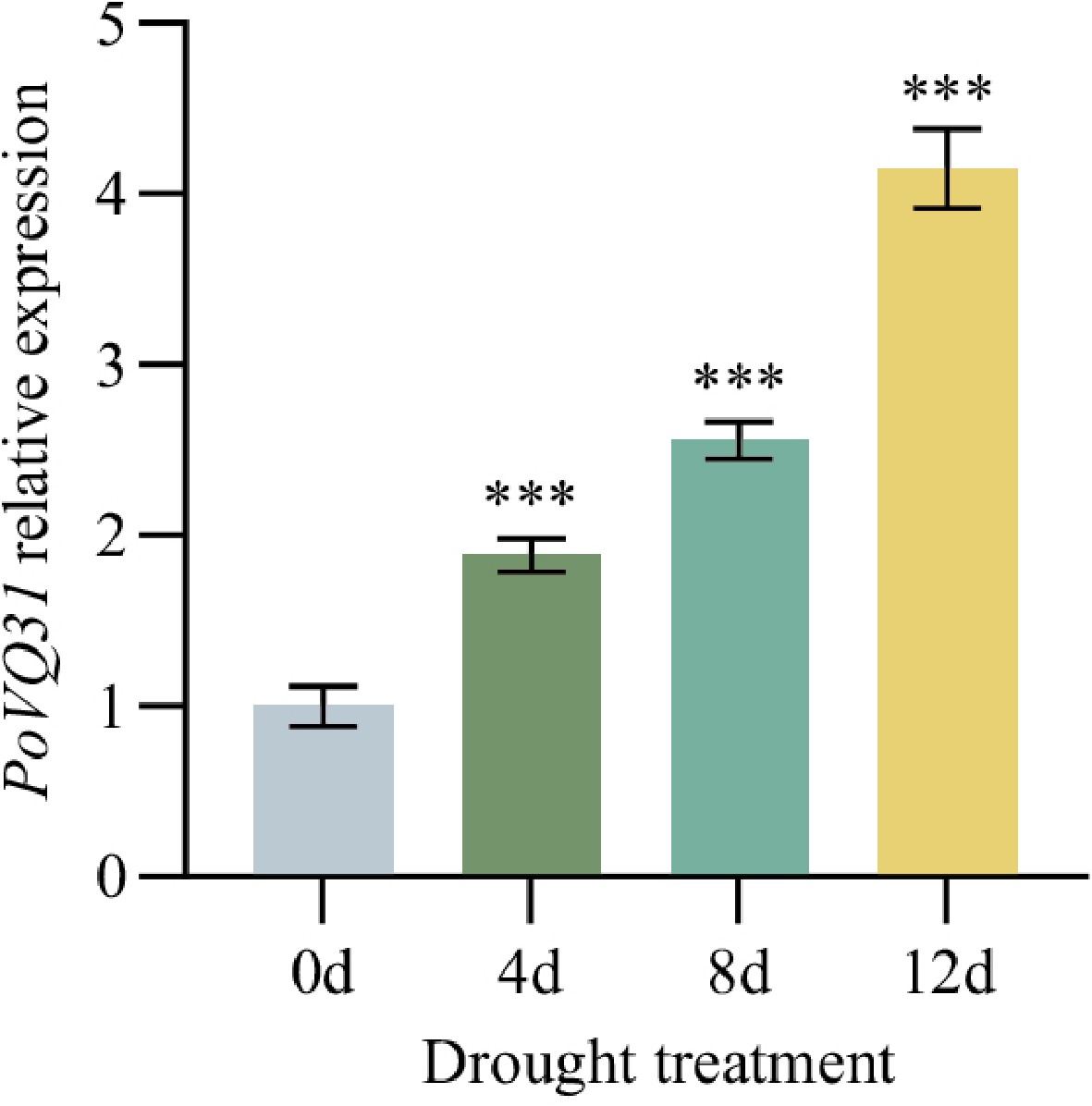
Figure 4.
Relative expression level of PoVQ31 under drought stress at different times. Data are presented as the mean ± standard error of the mean. Significant differences from the 0d were determined by Student's t-test (*** p < 0.001).
Silencing of PoVQ31 decreases drought tolerance in P. ostii 'Fengdan'
-
To investigate the biological function of PoVQ31 in drought stress, virus-induced gene silencing technology (VIGS) was used to generate two PoVQ31-silenced P. ostii 'Fengdan' lines, designated as Line 1 and Line 2, and empty TRV2 was defined as the control (Fig. 5a). Then it was verified that whether these vectors were transferred into the plant. When PCR amplification of plant DNA was performed using primers of PoUbiquitin and TRV1, a single bright band was observed in both TRV2 and TRV2-PoVQ31 plants. However, when TRV2 primers were used, there was a significant length difference between TRV2 and TRV2-POVQ31 plants. These results indicated that TRV1 and TRV2 vectors were successfully inserted into P. ostii 'Fengdan' (Fig. 5b). The qRT-PCR experiments suggested that the expression level of TRV2-POVQ31 plants was only half of that in the control, indicating the PoVQ31 gene was successfully silenced in P. ostii 'Fengdan' (Fig. 5c). Finally, we subjected the plants to a short-term drought treatment for 15 d and evaluated their drought-resistant phenotypes. As shown in Fig 5a, compared with TRV2, TRV2-POVQ31 plants had impaired drought tolerance and a higher degree of leaf and stalk wilt. These results suggested that silencing PoVQ31 in P. ostii 'Fengdan' weakened the drought tolerance of the plants.
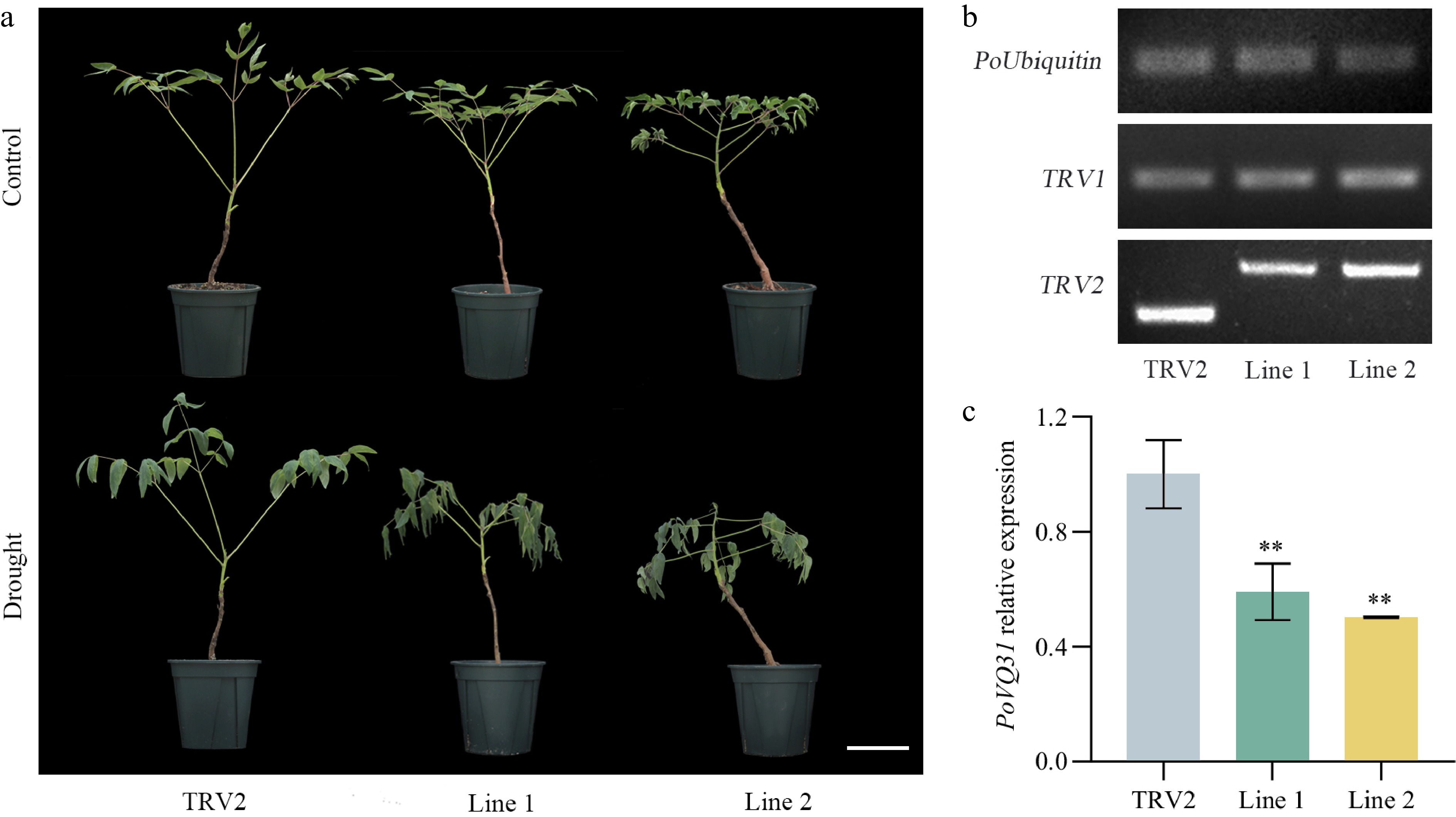
Figure 5.
Virus induced gene silencing of PoVQ31 in Paeonia ostii 'Fengdan'. (a) Phenotypic comparison of TRV2 (empty vector-transformed) and silenced plants after 15 d of drought treatment. Scale bars = 10 cm. (b) Identification of positive plants by PCR technology. (c) Silencing efficiency detection of positive lines by qRT-PCR technology. Data are presented as the mean ± standard error of the mean. Significant differences from the TRV2 were determined by Student's t-test (** p < 0.01).
PoVQ31 enhances P. ostii 'Fengdan' drought resistance by ROS
-
After observing the phenotypic differences, physiological indices of the transgenic plants were measured to further illustrate the function of PoVQ31 in plant drought tolerance. The leaf water content of TRV2-PoVQ31 plants was significantly lower than that of TRV2 plants under drought treatment, while there was no difference under normal conditions (Fig. 6c). On the contrary, under drought stress, the REC of TRV2-POVQ31 plants was significantly greater, indicating that the cell membrane of TRV2-POVQ31 plants was more damaged under drought stress (Fig. 6d). In addition, TRV2-PoVQ31 plants accumulated higher levels of MDA than TRV2 plants under drought conditions (Fig. 6e). Correspondingly, DAB staining and NBT staining showed larger and deeper stains in the leaves of TRV2-PoVQ31 plants. It meant that drought treatment induced a greater accumulation of hydrogen peroxide and O2·− in TRV2-PoVQ31 plants than in TRV2 (Fig. 6a & b). POD, SOD, CAT, and APX are the main enzymes in plants' defense system against stress. They cooperate to eliminate excess reactive oxygen species from the plant body and increase the plant's resistance to adversity[20]. The figure showed no significant difference in four antioxidant enzymes activity between TRV2 and TRV2-PoVQ31 plants before treatment. All four enzyme activities increased under drought conditions, but enzyme activity was significantly lower in TRV2-PoVQ31 plants than in TRV2 plants. This indicated that the antioxidant enzyme system of PoVQ31-silenced plants were severely damaged, making them inefficiently scavenge the excess accumulation of ROS under drought stress (Fig. 7). Various physiological indexes proved that PoVQ31 gene-silenced plants were more sensitive to drought stress than the blank control. PoVQ31 could effectively activate the antioxidant enzyme activity system to scavenge ROS stress, which positively regulated the drought response mechanism of P. ostii 'Fengdan' and enhanced its drought tolerance.
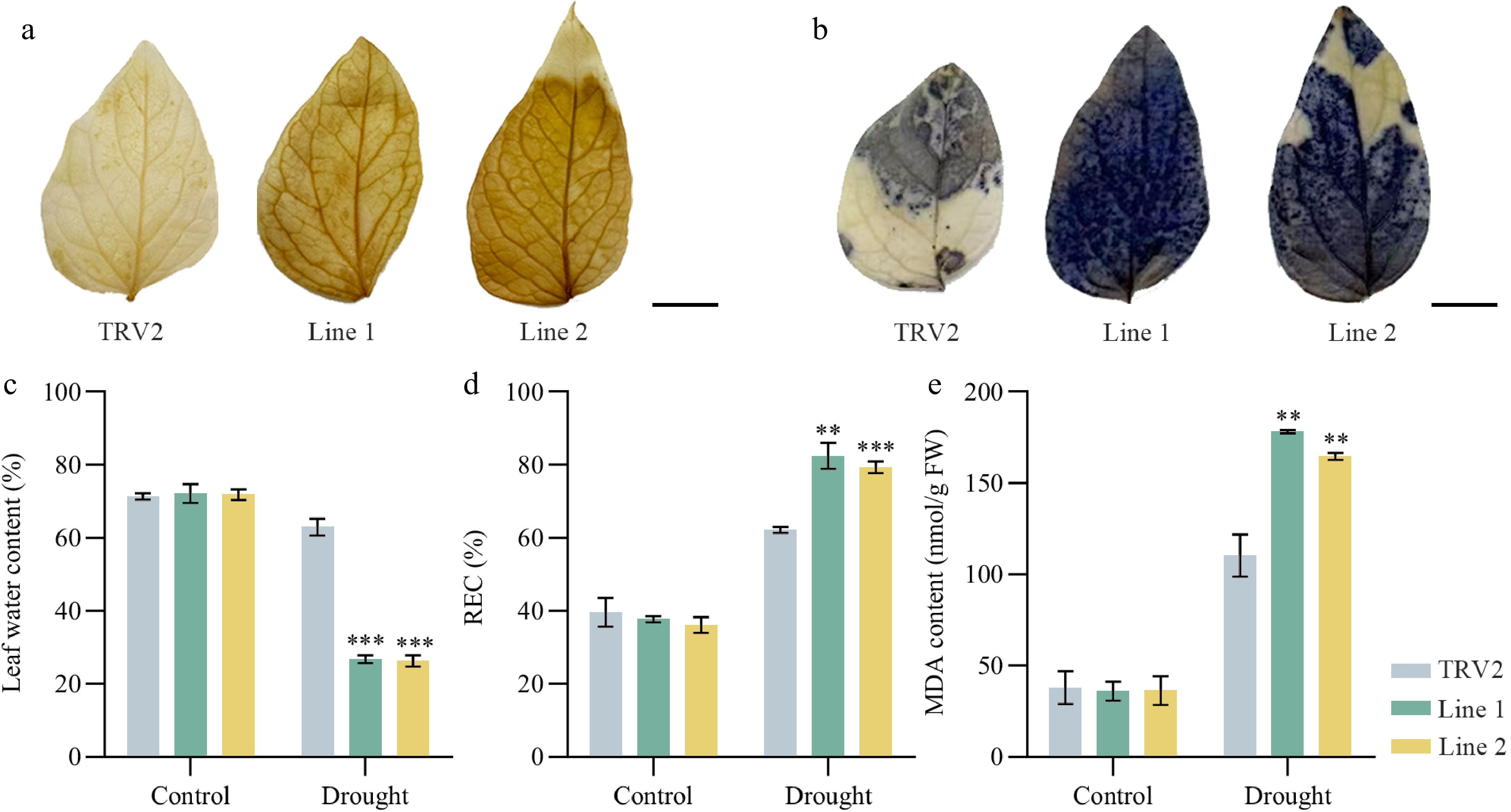
Figure 6.
Physiological indices of TRV2 and PoVQ31-silenced plants under drought treatment. (a) DAB and (b) NBT staining of Paeonia ostii 'Fengdan' leaves. Greater superoxide accumulation is indicated by darker staining. Scale bars = 1 cm. (c) Leaf water content. (d) Leaf REC. REC, relative electric conductivity. (e) Leaf MDA content. MDA, malondialdehyde. Data are presented as the mean ± standard error of the mean. Significant differences from the TRV2 were determined by Student's t-test (** p < 0.01, *** p < 0.001).
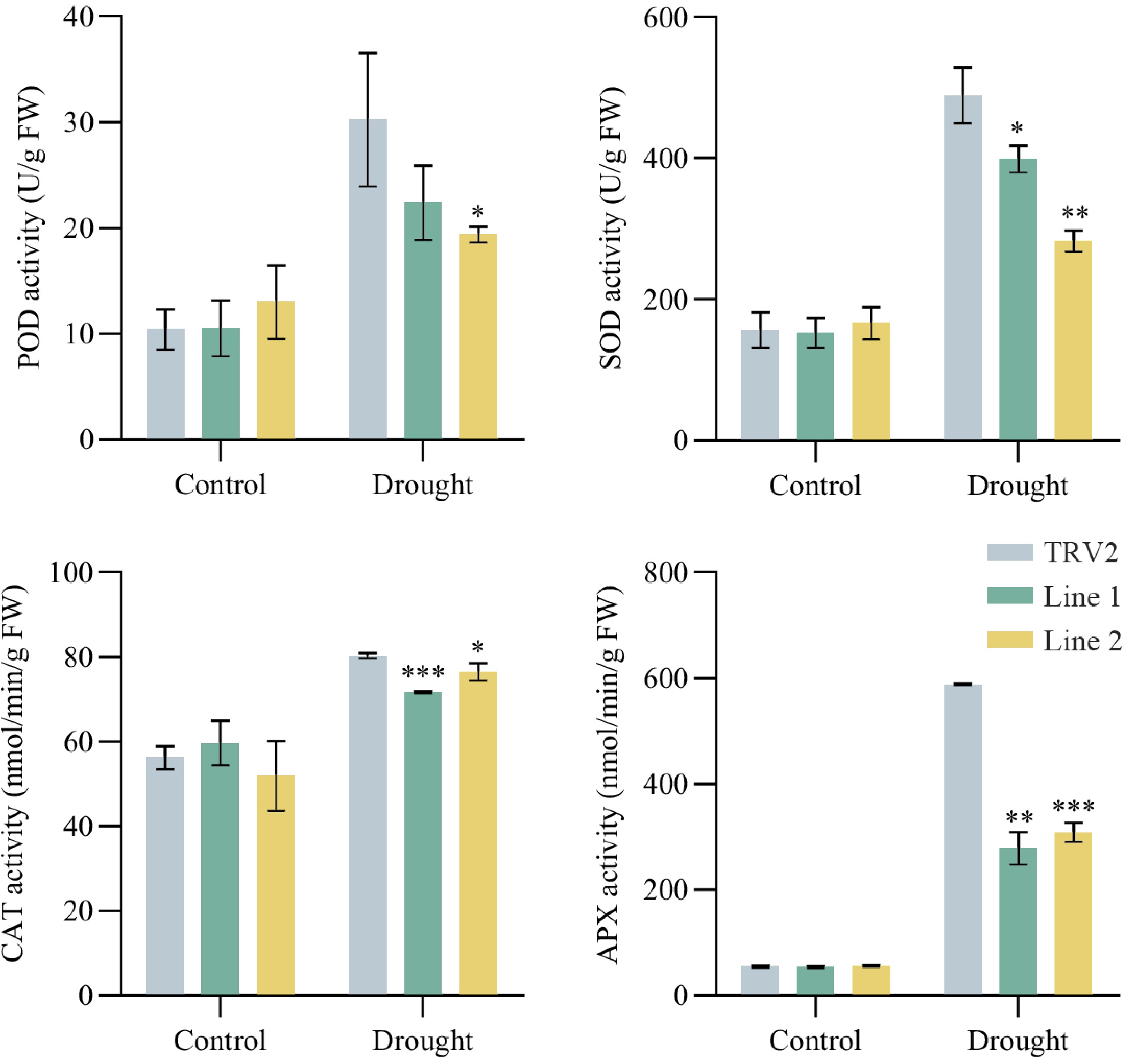
Figure 7.
Enzyme activities of TRV2 and PoVQ31-silenced plants under drought treatment. POD, peroxidase; SOD, superoxide dismutase; CAT, catalase; APX, ascorbate peroxidase. Data are presented as the mean ± standard error of the mean. Significant differences from the TRV2 were determined by Student's t-test (* p < 0.05, ** p < 0.01, *** p < 0.001).
-
Drought is a destructive abiotic stress factor that exerts a profound impact on the growth and development of plants[21]. Transcription factors can regulate the expression of stress-related genes by interacting with cis-acting elements or other proteins. In this way, they engage in plants physiological reactions to drought stress[22]. At present, several different families of transcription factors have been identified in higher plants, including myeloblastosis (MYB), basic leucine zipper (bZIP), and WRKY[23]. These transcription factors play a key role in plants' fight against drought environment. For example, when SpWRKY1 was overexpressed in Nicotiana tabacum, transgenic plants showed lower ROS levels and higher photosynthetic rates. The expression of downstream stress-related genes was also enhanced, which endowed transgenic plants with stronger drought resistance[24]. However, not all WRKY transcription factors have positive regulatory effects. For example, overexpression of GhWRKY68 in cotton resulted in reduced plant tolerance to drought stress[25]. To enhance plants through genetic engineering, a comprehensive grasp of transcription factor roles and regulatory mechanisms are necessary.
As a transcriptional coregulator, VQ proteins can interact with transcription factors and participate in a variety of biological processes, including responses to biotic and abiotic stresses, seed development and light morphogenesis[26]. For example, in A. thaliana, deletion mutants of AtVQ8 presented growth retardation and yellow-green leaf color. While overexpression of AtVQ17, AtVQ18 or AtVQ22 resulted in severe inhibition of plant growth[9]. AtVQ10 was involved in the JA-mediated signaling pathway through its interaction with AtWRKY8, which positively regulated resistance in Botrytis cinerea[27]. It has been demonstrated that in Raphanus sativus, the interaction between RsVQ4 and RsWRKY26 enhanced the DNA binding capacity of RsWRKY26, consequently stimulating the expression of the downstream RsHSP70-20 gene. Moreover, RsWRKY26 could bind to the RsVQ4 promoter and activate its transcription, forming a positive feedback loop that jointly regulated heat stress response[28]. At present, most of the research on VQ protein focuses on cloning and identification of VQ whole family genes. However, little is known about how VQ proteins respond to abiotic stress, particularly drought stress. Although there are studies on the expression of the VQ gene under drought stress, there is a lack of functional verification. Therefore, it is important to explore the response mechanism of VQ protein under drought stress.
VQ proteins, which are prominently characterized by their VQ motifs, display unique biological functions. In this study, the 32 to 41 amino acids of PoVQ31 protein have the typical conserved motif (FxxhVQxhTG) of the family. Mutations in the VQ motif may affect its molecular function, such as a single mutation in AtVQ29 (V70A) or AtVQ29 (V70D) that relieved AtVQ29 of its transcriptional repressive activity[10]. VQ proteins also rely on the VQ motif to interact with WRKY transcription factors and MAPKs. Under normal conditions, AtVQ4/MVQ1 interacted with AtWRKY33 and repressed AtWRKY33 transcriptional activity. However, when the pathogen invaded, AtMPK3/6 mediated phosphorylation led to instability and degradation of AtVQ4/MVQ1, diminishing or eliminating repression of AtWRKY33 activity. With increasing levels of phosphorylation, the expression of disease resistance genes downstream of AtWRKY33 rose, thereby regulating the immune response in A. thaliana[29]. Bioinformatics predictions in this study showed that the PoVQ31 protein interacted with several WRKY family proteins and had multiple phosphorylation sites (Supplemental Fig. S1; Fig. 2d). Whether there is a MAPK-VQ-WRKY-like ternary regulatory mechanism involved in drought stress response in P. ostii 'Fengdan' remains to be further verified. VQ motifs have been hypothesized to be associated with the nucleus location of proteins[30]. Consistent with this, PoVQ31 was also shown localized to the nucleus in this study.
Several studies have shown that VQ genes can be significantly induced in expression by abiotic stresses. Transcriptome data of A. thaliana showed that 56% (19/34) of VQ gene expression was up-regulated after ozone treatment[31]. AtVQ9 was strongly induced by NaCl treatment, and its mutants showed higher seed germination and better seedling growth under NaCl treatment. Whereas overexpression lines showed the opposite effect, suggesting that AtVQ9 negatively regulated A. thaliana resist to NaCl stress[16]. Similarly, TaVQ14 expression was up-regulated in the seeds of Triticum aestivum under drought and salt stress, and heterologous overexpression of TaVQ14 increased A. thaliana seed resistance to salt and drought stress[32]. This study found that the expression of PoVQ31 was significantly raised with drought time migration, suggesting that PoVQ31 might be involved in the regulation of response to drought stress.
The drought resistance function of VQ protein has not been systematically studied in the woody plant P. ostii 'Fengdan'. The purpose of this study was to reveal the specific role of PoVQ31 in drought stress response by producing POVQ31-silenced transgenic plants of P. ostii 'Fengdan'. The experimental data showed that drought could significantly induce the expression of PoVQ31. Compared with the control group, transgenic plants were more sensitive to drought. Under drought stress, they showed lower water content, protective enzyme activity, and higher REC and MDA accumulation. These results indicated that PoVQ31 played a positive regulatory role in the drought resistance mechanism of P. ostii 'Fengdan'.
Drought stress can intensify the production and accumulation of ROS in plants. Excess ROS can cause oxidative stress, resulting in damage to cell components such as lipids, nucleic acids, metabolites, and proteins, and ultimately plant death[20]. Several studies have shown that the ROS regulation system is crucial for P. ostii 'Fengdan' to cope with drought stress[33,34]. In this study, to verify the biological function of PoVQ31, we produced PoVQ31 gene silencing lines using VIGS technology. The related ROS-scavenging enzyme activities of the control group and the silent strain were detected. It was found that the POD, SOD, CAT, and APX activities of the transgenic strain were significantly lower under drought stress. In contrast, DAB and NBT staining showed that transgenic plants were subjected to more severe oxidative stress. These results demonstrated that PoVQ31 was indispensable for activating the antioxidant enzyme system to scavenge ROS damage to alleviate drought stress.
In summary, PoVQ31 was cloned and identified, a member of the P. ostii 'Fengdan' VQ family, and its function was verified. It was found that PoVQ31 can alleviate ROS accumulation in response to drought stress, but the specific molecular regulatory mechanism needs to be further investigated. In previous studies, VQ protein can interact with WRKYs and MAPKs to regulate downstream defense gene expression. In the future, yeast dual-hybrid technology can be used to find transcription factors interacting with POVQ31 and further clarify its regulatory network.
-
In this study, PoVQ31 was successfully cloned and identified, a member of the VQ family in P. ostii 'Fengdan'. Its protein spatial structure and physicochemical properties were predicted. The PoVQ31 protein was specifically localized in the nucleus and its expression was significantly induced under drought stress. Silencing of the PoVQ31 gene by VIGS technology revealed enhanced drought sensitivity. Reduced leaf water content, and increased REC and MDA accumulation were presented in the gene silenced plants. We speculated that drought stress injury in P. ostii 'Fengdan' was associated with ROS accumulation, and the results of MDA and NBT staining supported this conclusion. In addition, the reduced activities of protective enzymes in transgenic plants indicated that PoVQ31 responded to drought stress by alleviating ROS accumulation. In conclusion, PoVQ31 was a positive regulator of P. ostii 'Fengdan' against drought stress.
-
The authors confirm contribution to the paper as follows: conceptualization, supervision, funding acquisition: Tao J, Zhao D; software, investigation, data curation, writing-original draft, visualization: An H; formal analysis: An H, Luan Y; writing-review & editing: An H, Luan Y, Zhao D; methodology: Zhao D. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This work was supported by Modern Agriculture (Flower) Industrial Technology System of Jiangsu Province (JATS[2023]446).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Supplementary data of gene-specific primers used in this study.
- Supplemental Fig. S1 Interacting protein prediction for PoVQ31.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
An H, Luan Y, Zhao D, Tao J. 2024. The VQ motif-containing PoVQ31 protein positively modulates drought stress tolerance in Paeonia ostii 'FengDan'. Ornamental Plant Research 4: e017 doi: 10.48130/opr-0024-0015 

The VQ motif-containing PoVQ31 protein positively modulates drought stress tolerance in Paeonia ostii 'FengDan'
- Received: 22 March 2024
- Revised: 04 May 2024
- Accepted: 13 May 2024
- Published online: 18 June 2024
Abstract: VQ proteins are transcription co-regulators that play important roles in plant growth, development, and response to biotic and abiotic stresses. However, the specific relationship between VQ proteins and drought stress has not been thoroughly studied in Paeonia ostii 'Fengdan'. In this study, PoVQ31 was successfully cloned, a gene with a total length of 540 bp and encoding 179 amino acids. Through bioinformatics analysis, the sequence characteristics, physical and chemical properties, and spatial structure were analyzed. It was observed that PoVQ31 had a typical VQ motif and was localized in the nucleus. Furthermore, its expression could be significantly upregulated in response to drought stress. PoVQ31-silenced lines showed more drought sensitivity compared to controls. Experiments further showed that transgenic lines had decreased leaf water content, but relative electrical conductivity (REC) and malondialdehyde (MDA) content increased compared with controls under drought stress conditions. The nitroblue tetrazolium (NBT) and diaminobenzidine (DAB) staining results showed that the transgenic plants accumulated excess reactive oxygen species (ROS). Meanwhile, the reduced antioxidant enzyme activity in PoVQ31-silenced plants prevented them from scavenging excessively accumulated ROS. All these results suggested that PoVQ31 might be involved in the activation of the ROS scavenging system, which in turn positively regulated the tolerance of Paeonia ostii 'Fengdan' to drought stress. This study provided novel insights into the function of VQ proteins and further enriched the understanding of this important protein in Paeonia ostii 'Fengdan'.
-
Key words:
- Paeonia ostii 'FengDan' /
- Drought stress /
- VQ31 /
- Reactive oxygen species