









-
Leucine-Rich Repeats Extensins (LRX) are a class of cell wall-anchored proteins, with an N-terminal leucine-rich repeat (LRR) domain and a C-terminal extensin domain. In plants, LRR domains are typically present in receptor proteins that can recognize and bind various external signaling molecules, such as hormones and pathogenic proteins. This binding triggers downstream signaling pathways, thereby regulating plant growth, development, and responses to the environment[1−3]. Extensins are highly glycosylated cell wall proteins, with multiple repetitive sequences and hydroxyproline-rich domains[4−6]. These structural features enable extensins to interact with other components within the cell wall, forming a cross-linking network, thereby increasing the strength and stability of the cell wall and playing an important role in cell wall formation and mechanical support for cells[7,8]. There are 11 LRX genes identified in Arabidopsis, and based on their tissue-specific expression patterns, they can be roughly divided into two subfamilies. The first subfamily includes LRX1 to LRX7, which are highly expressed in rapidly growing tissues such as root tips, shoot tips, and young leaves[9,10]. LRX genes play an essential role in plant cell wall formation and cell elongation processes, suggesting their involvement in the regulation of plant development. Specifically, LRX1 is crucial for root hair cell wall formation[11]. Additionally, LRX3/4/5 are crucial for salt tolerance in Arabidopsis. LRX interacts with Rapid Alkalinization Factors (RALF) peptides RALF22/23, which in turn interact with the plasma membrane-localized receptor-like protein kinase FERONIA (FER), forming the LRX3/4/5-RALF22/23-FER module. This module controls plant growth and salt stress responses by regulating hormone homeostasis and reactive oxygen species (ROS) accumulation[12−14]. The second subfamily includes LRX8 to LRX10 (also known as AtPEX1-4, Pollen extension-like), which are expressed in specific reproductive tissues such as flowers and pollen[15−17]. Plant reproduction relies on tightly regulated pollen tube growth to deliver sperm. This process is controlled by secreted RALF peptides, which have been shown to be perceived by Catharanthus roseus RLK1-like (CrRLK1Ls) receptor-like kinases/LORELEI-like GLYCOLPHOSPHATIDYLINOSITOL (GPI)-anchored protein (LLG) complexes or leucine-rich repeat extensins. LRX8 to LRX11 can interact with RALF4/19 on the pollen tube cell wall and enter the signaling pathway mediated by ANNEXIN1/2 (ANX1/2), thus regulating pollen germination and pollen tube growth[18−20]. Disruption of LRX protein function leads to unstructured pectin deposition and changes in wall mechanical properties, resulting in pollen tube defects[18].
In addition, in contrast to the tissue specificity of the Arabidopsis PEX genes in reproductive organs, rice OsPEX1, besides being expressed outside of reproductive tissues, also exerts negative regulation on root growth in gibberellin-mediated pathways and regulates root growth by influencing cell wall plasticity[21]. The LRR domain of maize and tomato PEX proteins is highly conserved, and the tomato PEX gene exhibits pollen specificity[1]. Comparison of PEX genes in maize, tomato, tobacco, or Arabidopsis revealed that PEX genes not only play a role as structural proteins in polarized pollen tube growth, but may also act as gender recognition factors in the process of interacting with the pistil[22].
The Cucurbitaceae plants are a pivotal group of economic crops, including many species of agricultural and horticultural significance such as cucumber, pumpkin, and watermelon. These crops are widely distributed across the world, particularly in tropical and subtropical regions of Asia, Africa, and the Americas[23,24]. Cucurbitaceae plants are rice in nutritional and medicinal value[25], containing substances such as cellulose, vitamins, minerals, and others[26−28], and they are used in traditional medicine to treat various diseases, including cancer[29], inflammation, diabetes, and detoxification[30]. In recent years, there has been an increasing amount of research on demonstrating that gene families play an essential role in the adaptive changes and physiological function of plants. Although the LRX gene families has been discovered in some plants, research on the LRX gene families in Cucurbitaceae plants remains relatively limited.
Here, the LRX family members were first identified from seven cucurbit species in Cucurbitaceae at the genome-wide level and obtained a total of 40 LRX genes. The gene structure, conserved motifs, conserved domains, chromosome localization, cis-acting elements, and phylogeny of these 40 LRX genes were then systematically analyzed. The synteny relationship of LRX genes between cucumber and six other cucurbit species was also discussed. Finally, the expression patterns of CsLRX genes in different tissues and under drought stress and salt stress in cucumber were analyzed by qRT-PCR. The results of this study will provide a theoretical basis for the functional identification of LRX genes in Cucurbitaceae crops.
-
Download the LRX protein sequences of Arabidopsis and rice from the Ensemble Plants database (
https://plants.ensembl.org/index.html ). The AtLRX1 (At1G12040) gene in Arabidopsis was used as a template to search for homologous genes of seven cucurbit species in the Cucurbitaceae database (http://cucurbitgenomics.org/search/genome/2 ), and further identified by BLAST and HMM Search in TBtools[31]. Additionally, a BLAST search of these LRX genes in the TAIR database were performed (www.Arabidopsis.org ) to obtain their homologous genes. The molecular weight (MW) and Isoelectric point (PI) of the identified proteins were studied by ExPASy online software (https://web.expasy.org/compute_pi/ ). The chromosome distribution map of LRX genes was drawn by TBtools[32].Phylogenetic analysis
-
There are 11 LRX genes known in Arabidopsis[33] and eight in rice[34]. To understand the evolutionary relationship of LRX genes in Cucurbitaceae, multiple sequence alignment was performed using the ClustalW tool for LRX proteins in Cucurbitaceae, as well as LRX proteins in Arabidopsis and rice. A phylogenetic tree was generated by the neighbor-joining (NJ) method with default parameter settings: bootstrap method setting to 1,000, Poisson model, and Pairwise deletion in MEGA 11[35]. Further visualization and refinement of the tree were performed via Interactive Tree Of Life (iTOL) (
https://itol.embl.de/ )[36].Gene structure analysis, identification of conserved domains and conserved motifs
-
Gene structure analysis was performed by Tbtools[32],extract the protein sequences of 40 LRX genes from seven species in Cucurbitaceae, and submit them to the NCBI-CDD (
www.ncbi.nlm.nih.gov/cdd ) and MEME (https://meme-suite.org/meme/tools/meme ) websites for predicting conserved domains and motifs[37,38]. Merge and visualize the results by TBtools[32].Synteny analysis
-
The genome and annotation files of Arabidopsis, rice, and seven cucurbit species were downloaded. The intraspecific collinearity analysis of the cucumber CsLRX gene family and the synteny analysis between the cucumber CsLRX genes and the LRX genes of other six cucurbit species were plotted using TBtools. In addition, TBtools was used to draw the synteny analysis between cucumber CsLRX genes and rice and Arabidopsis[32].
Analysis of cis-acting elements
-
The genomic DNA sequences of 2,000 base pairs (bp) upstream of the transcription start site of 40 LRX genes were extracted from the reference genomes of seven cucurbit species using TBtools, and then submitted to the PlantCare database (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) to identify possible cis-regulatory elements[39]. Visualize, classify, and analyze the prediction results. The heat map of cis-acting element analysis is generated by TBtools[32].Plant materials
-
The North China type (Chinese Long) cucumber inbred line 3667 was grown in a greenhouse of the China Agricultural University in Beijing (Beijing, China). Samples from different tissues such as roots, stems, young leaves, tendrils, female flowers, and male flowers were collected. The samples were rapidly frozen in liquid nitrogen and stored at −80 °C.
RNA extraction and qRT-PCR analysis
-
For analysis of CsLRX gene expression patterns, including salt stress and drought stress. The sample RNA was extracted using the Quick RNA Isolation Kit (Huayueyang, Beijing, China). The FastKing gDNA Dispelling RT SuperMix (TianGen Biotech, Beijing, China) was applied to synthesize the first-strand cDNA with the extractive RNA template.qRT-PCR was performed using the UltraSYBR Mixture (Low ROX) (Cwbio, Beijing, China) on an Applied Biosystems 7500 real-time PCR system (Applied Biosystems, Foster City, CA, USA). The UBIQUITIN EXTENSION PROTEIN (UBI-EP) gene was used as a reference gene. Three biological and three technical replicates were carried out for expression dynamics analysis. The significant differences were analyzed by Student's t-tests (p < 0.05). The primers are listed in Supplemental Table S1.
Subcellular localization
-
Cloning of CsLRX1 and CsLRX3 coding sequences without stop codons were constructed into the pSuper-1300 vector containing the green fluorescent protein (GFP) to create a fusion protein. After selecting the correct vector through sequencing, extract the plasmid and transform it into the GV3101 Agrobacterium strain. The Agrobacterium liquid was then resuspended, which contains the target vector, a nuclear marker vector, and P19, in infection solution to an OD600 of approximately 1.0, then mixed and left to stand for 3 h. Four-week-old N. benthamiana leaves were selected for infiltration, and after a dark treatment of 12−24 h, followed by a light treatment of 48 h, the GFP fluorescence was observed using a confocal microscope. The subcellular localization primers are listed in Supplemental Table S1.
-
The known LRX genes (AtLRX1, At1G12040) protein sequences of Arabidopsis were used as templates to search their homologous genes in the genome database (
http://cucurbitgenomics.org/search/genome/2 ) of seven cucurbit species, respectively. BLAST and HMM Search in Tbtools were used to further identify and finally, 40 LRX genes were obtained as shown in Table 1, including six from cucumber (C. sativus), five from wax gourd (B. hispida), four from watermelon (C. lanatus), eight from pumpkin (C. maxima), eight from moschata pumpkin (C. moschata), five from bottle gourd (L. siceraria), and four from melon (C. melon) (Table 1). The coding sequence lengths ranged from 936 to 4,956 bp. The physical characteristics of these LRX proteins were predicted, revealing significant differences among the 40 LRX family proteins (Table 1). The average amino acid length was 646.4 aa, with predicted protein molecular weights ranging from 42,123.14 to 166,407.81 Da, and theoretical Isoelectric points (PI) ranging from 3.7 to 10.08 (Table 1). The identified genes are named based on their chromosomal locations rather than following uniform naming conventions.Table 1. Genome-wide identification of LRX gene family members in seven cucurbit species.
Gene name Gene ID Gene position CDS (bp) AA MW (Da) PI Arabidopsis homology Start End (+/−) CsLRX1 Csa1G383520.1 14493464 14496369 (+) 2,328 776 42,123.14 4.84 AT4G13340 CsLRX2 Csa2G004760.1 814446 817377 (+) 2,298 766 82,435.4 6.5 AT4G13340 CsLRX3 Csa3G146350.1 9732797 9733957 (+) 1,161 387 43,772.94 5.71 AT3G22800 CsPEX1 Csa6G005160.1 473143 474726 (−) 1,584 528 42,813.05 4.68 AT2G15880 CsPEX2 Csa6G006180.1 487825 489724 (−) 1,854 618 76,300.69 6.17 AT2G15880 CsPEX3 Csa6G006190.1 491897 494318 (+) 2,286 762 41,120.04 5.01 AT2G15880 BhiLRX1 Bhi01M000558 14563184 14564624 (+) 1,158 386 89,384.76 6.46 AT3G22800 BhiLRX2 Bhi02M000924 25237963 25240855 (−) 2,304 768 82,573.58 5.98 AT3G24480 BhiLRX3 Bhi06M001733 55954895 55964421 (+) 1,170 390 76,954.31 5.4 AT4G18670 BhiPEX1 Bhi09M001722 56817604 56820319 (−) 1,176 392 57,276.43 5.83 AT3G19020 BhiPEX2 Bhi12M001681 60171798 60173997 (+) 2,100 700 120,050.2 7.96 AT3G19020 ClaLRX1 Cla97C05G086020.1 4563471 4564598 (+) 1,128 376 41,548.56 4.58 AT3G22800 ClaLRX2 Cla97C06G119550.1 20517876 20520416 (+) 2,541 847 148,150.13 5.41 AT3G24480 ClaLRX3 Cla97C11G208980.1 2574856 2577150 (+) 2,295 765 166,407.81 3.7 AT4G13340 ClaPEX1 Cla97C06G112900.1 3979734 3981893 (−) 2,160 720 81,584.74 6.46 AT3G19020 CmaPEX1 CmaCh01G018740.1 12291107 12293909 (−) 1,590 530 54,054.38 5.13 AT4G33970 CmaLRX1 CmaCh05G013340.1 10085979 10089368 (−) 3,390 1,130 74,989.78 4.43 AT3G24480 CmaLRX2 CmaCh06G013780.1 8987856 8988998 (+) 1,143 381 42,903.13 5.18 AT3G22800 CmaPEX2 CmaCh08G008980.1 5563777 5568808 (−) 4,275 1,425 40,235.09 4.75 AT4G33970 CmaPEX3 CmaCh09G001990.1 826607 831562 (+) 4,956 1,652 35,186.52 4.96 AT3G19020 CmaLRX3 CmaCh10G006020.1 2780772 2788301 (−) 2,328 776 52,221.21 5.8 AT4G13340 CmaLRX4 CmaCh14G017400.1 12740774 12742300 (−) 1,527 509 88,647.32 6.16 AT3G22800 CmaPEX4 CmaCh17G004460.1 2660615 2662696 (+) 2,082 694 83,972.54 5.88 AT2G15880 CmoPEX1 CmoCh01G019320.1 13801421 13802672 (−) 1,182 394 40,916.77 4.78 AT3G19020 CmoLRX1 CmoCh06G013900.1 10200801 10201904 (+) 1,104 368 85,186.25 4.81 AT3G22800 CmoPEX2 CmoCh08G008690.1 5609995 5611160 (−) 936 312 82,799.35 6.04 AT3G19020 CmoPEX3 CmoCh08G008700.1 5621196 5622985 (−) 1,413 471 82,892.28 6.06 AT3G19020 CmoLRX2 CmoCh10G006420.1 2945225 2947750 (−) 2,526 842 42,186.25 4.73 AT4G18670 CmoLRX3 CmoCh12G012710.1 11268328 11273318 (−) 2,328 776 55,986.22 4.62 AT4G13340 CmoLRX4 CmoCh14G017780.1 13715371 13717483 (−) 1,128 376 66,641.59 5.19 AT3G22800 CmoPEX4 CmoCh17G004250.1 2835184 2837965 (+) 2,355 785 79,908.67 4.65 AT2G15880 LsiPEX1 Lsi04G016020.1 23544642 23546786 (−) 1,944 648 67,872.59 5.61 AT2G15880 LsiLRX1 Lsi05G016310.1 23953552 23954682 (−) 1,131 377 41,040.03 4.92 AT3G22800 LsiLRX2 Lsi06G008900.1 18515443 18517854 (+) 2,412 804 85,115.97 6.11 AT4G13340 LsiPEX2 Lsi09G015830.1 23966231 23967994 (−) 1,764 588 62,720.9 4.87 AT2G15880 LsiLRX3 Lsi11G002700.1 2740543 2743776 (+) 2,226 742 79,910.5 5.85 AT4G13340 MELOLRX1 MELO3C004550.2.1 28466831 28470698 (−) 1,770 590 null null AT3G24480 MELOLRX2 MELO3C006506.2.1 3799835 3800974 (+) 1,122 374 40,873.81 4.84 AT3G22800 MELOPEX1 MELO3C021195.2.1 31428500 31430511 (−) 1,947 649 70,632.34 6.37 AT3G19020 MELOPEX2 MELO3C034935.2.1 31442588 31444077 (+) 1,446 482 52,356.41 5.13 AT3G19020 CDS: coding sequence, AA: the number of amino acids, MW: Molecular weight, PI: Theoretical isoelectric point. To clarify the physical locations of LRX genes in the genomes of these cucurbit species, chromosome distribution maps of these genes were generated (Supplemental Fig. S1). Specifically, in cucumber, six LRX genes were distributed on chromosomes 1, 2, 3, and 6; in wax gourd, five were found on chromosomes 1, 2, 6, 9, and 12; in watermelon, four were located on chromosomes 5, 6, and 11; in pumpkin, eight were distributed on chromosomes 1, 5, 6, 8, 9, 10, 14, and 17; in moschata pumpkin, eight were present on chromosomes 1, 6, 8, 10, 12, 14, and 17; in bottle gourd, five were found on chromosomes 4, 5, 6, 9, and 11; and in melon, four were located on chromosomes 5, 6, and 11 (Supplemental Fig. S1).
Phylogenetic analysis of the LRX genes in Cucurbitaceae
-
In Arabidopsis, there are known to be 11 LRX genes[33], while rice contains eight LRX genes[34]. To further explore the phylogenetic relationships and evolutionary patterns of LRX genes in the Cucurbitaceae family, this study utilized the 11 LRX genes in Arabidopsis, the eight LRX genes in rice, and the 40 LRX gene members from seven cucurbit species to construct a phylogenetic tree using the neighbor-joining method (Fig. 1). In Arabidopsis, LRX genes are typically divided into two subfamilies, with genes highly expressed in vegetative tissues represented by LRX, and genes highly expressed in reproductive organs represented by PEX[34]. Consistent with previous studies, all LRX genes were divided into two clades, with the first clade containing 34 LRX homologues (including seven AtLRX, five OsLRX, three CsLRX, three BhiLRX, three ClaLRX, four CmaLRX, four CmoLRX, three LsiLRX, two MELOLRX), while the remaining 25 LRX homologs belonged to the second clade (including four AtPEX, three OsPEX, three CsPEX, two BhiPEX, one ClaPEX, four CmaPEX, four CmoPEX, two LsiPEX, two MELOLRX) (Fig. 1). The evidence shows that these LRX genes are closely related in their evolutionary relationships, with no significant genetic differences.
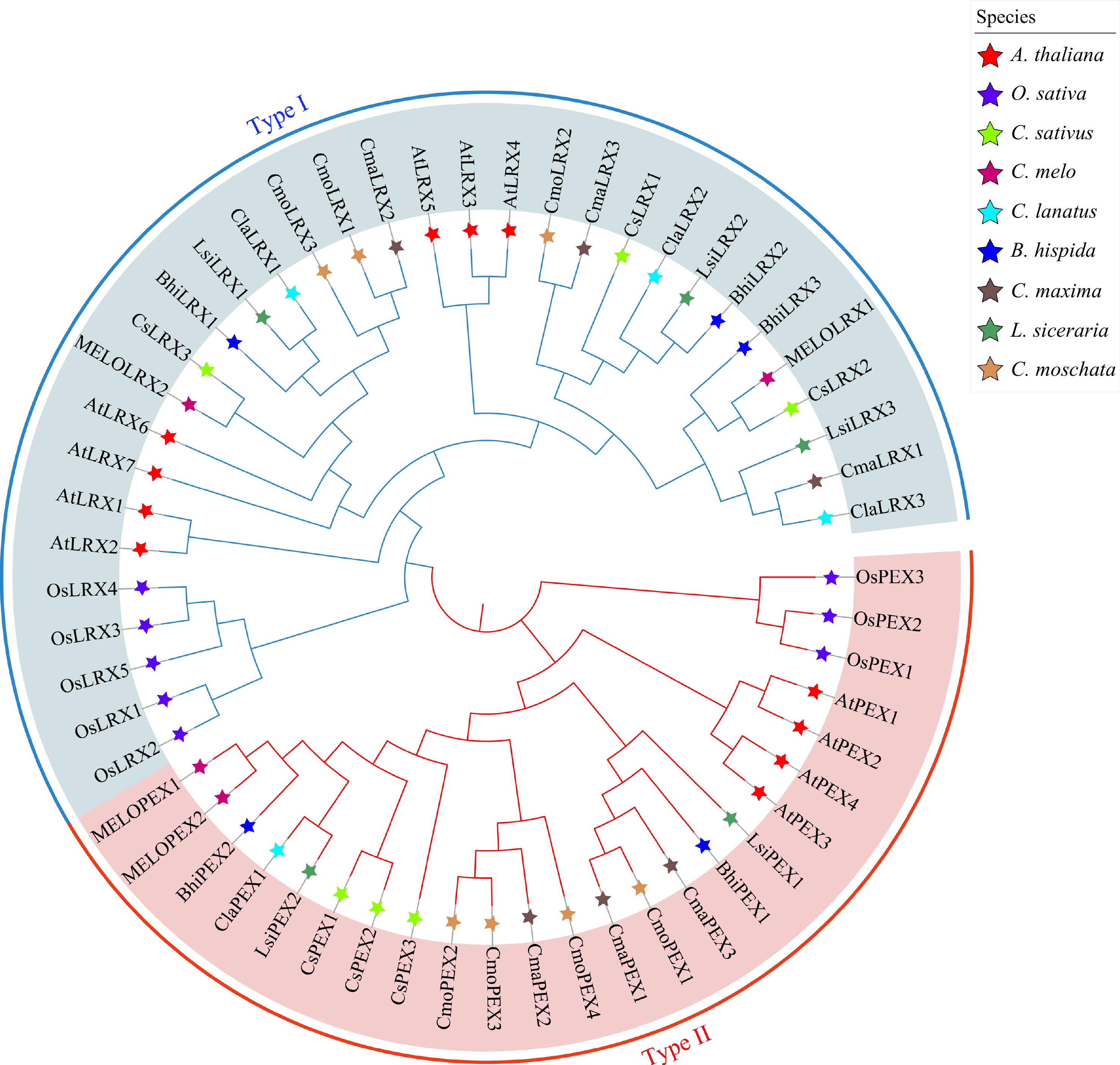
Figure 1.
Phylogenetic tree of the LRX proteins from Arabidopsis, rice, and seven cucurbit species. Red star, purple star, light green star, deep red star, sky blue star, deep blue star, deep brown star, deep green star and light brown star represent Arabidopsis thaliana (A. thaliana), rice (O. sativa), cucumber (C. sativus ), melon (C. melon), watermelon (C. lanatus), wax gourd (B. hispida), pumpkin (C. maxima), bottle gourd (L. siceraria) and moschata pumpkin (C. moschata), respectively.
Analysis of gene structure, conserved motifs, and conserved domains of LRX genes in Cucurbitaceae
-
The gene structure provides important clues for its functional diversification and can also reflect the evolutionary history of the gene family. Therefore, the exon-intron structure of these LRX genes was further analyzed (Fig. 2). Gene structure analysis showed that most genes lack a 5' untranslated region (UTR) or a 3' UTR region, containing 1 to 2 exons, with most genes lacking introns. For example, all four LRX genes in watermelon contain only one exon structure, without introns and UTR regions. Among the six LRX genes in cucumber, only two genes each contain one intron, while the remaining four have no introns (Fig. 2). This is consistent with the prediction of their small number of transcripts. Although the UTR regions of LRX genes within the same subfamily exhibit differences in size and structure, suggesting their involvement in different regulatory processes and potentially functioning in different biological functions. Overall, genes within the same subfamily show a similar gene structure, indicating a higher degree of conservation during the evolutionary process. This conservation may reflect the similar biological functions performed by these genes across different species or the presence of similar evolutionary pressures.

Figure 2.
Phylogenetic clustering, conserved motifs, conserved domains and gene structures of seven cucurbits LRX genes. Left one: The phylogenetic tree of LRX genes. Light green and brownish red represent LRX and PEX subfamilies, respectively. Left two: the conserved motif of LRX genes, different colors represent different motifs; right one: LRX gene structure analysis, green and blue represent the UTR region and the CDS region, respectively; right two: the conserved domain of LRX genes, different colors represent different domains.
Subsequently, the MEME software was used to predict the conserved motifs in these LRX genes (Fig. 2, Supplemental Fig. S2). Among them, Motif1 to Motif7 are conserved motifs, shared by the vast majority of LRX genes and mainly located at the N-terminus. Motif10 appears most frequently and is located at the C-terminal, with 22 tandem repeats in CmaLRX1, suggesting that this repetition indicates the importance of this motif for gene function. Motif9 was not found in the LRX genes of melon, but is present in the other six species of cucurbits, suggesting that the LRX genes in melon have undergone specific changes in function or regulation, which may also reflect the diversity of this motif among different species or individuals (Fig. 2). Considering that LRX is a class of cell wall proteins containing a conserved N-terminal leucine-rich repeat domain[16] and a typical C-terminal extensin protein domain[4], we visualized the conserved domains of 40 LRX proteins and found that all LRX genes contain these two conserved sequences (Fig. 2, Supplemental Fig. S3), indicating the high functional and structural conservation of these genes.
Synteny analysis of LRX genes
-
The existence of collinearity can provide important clues for gene evolution and evolutionary relationships. By comparing genes with collinearity, it is possible to reveal their common ancestors and evolutionary history, and to infer the origin and expansion process of gene families[40]. To better understand the amplification patterns of LRX genes during the evolutionary process, we used TBtools software to conduct collinearity analysis of LRX genes in cucumber and six other cucurbit species (Fig. 3a). The results indicated that cucumber shares collinear genes with 4, 6, 8, 7, 6, and 6 genes in wax gourd, watermelon, pumpkin, moschata pumpkin, bottle gourd, and melon, respectively, suggesting that cucumber, wax gourd, watermelon, pumpkin, moschata pumpkin, bottle gourd, and melon may have common ancestral genes and have retained similar gene structures and functions during the evolutionary process (Supplemental Table S2). Eight three percent of cucumber CsLRX genes showed a synteny relationship with moschata pumpkin, while 75% of cucumber CsLRX genes showed synteny relationship with watermelon, bottle gourd, and melon, with the least relationship found between wax gourd and pumpkin, accounting for 50% (Fig. 3a). Interestingly, CsLRX1 and CsLRX2 in cucumber both show collinearity with LRX genes in the other six cucurbit species, and each has two pairs of collinear gene pairs with melon, watermelon, bottle gourd, and pumpkin. In addition, CsLRX1 and CsLRX2 replicate each other (Fig. 3c), promoting the expansion of the cucumber LRX gene family. The different numbers of collinearity relationships between cucumber LRX genes and LRX genes in other cucurbit species may indicate gene rearrangements, insertions, or deletions during the evolution of these crops, leading to differences in the number of collinear gene pairs in their genomes.
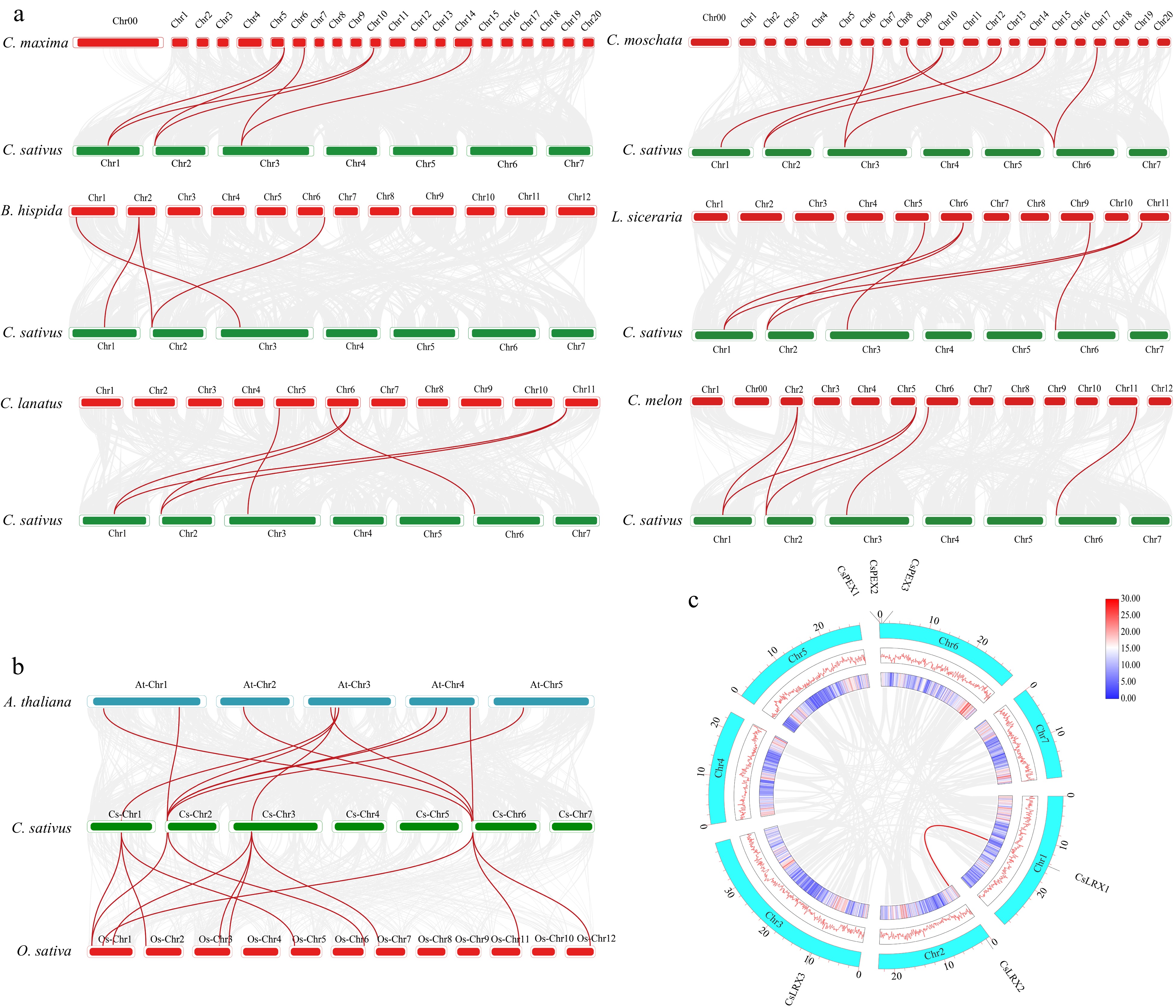
Figure 3.
Synteny analysis of LRX genes. (a) Synteny analysis of LRX genes between cucumber and six other cucurbits. (b) Synteny analysis of LRX genes in cucumber, Arabidopsis thaliana and rice (O. sativa). (c) Colinearity analysis of CsLRX genes in cucumber species. The red and blue lines represent gene pairs with collinearity.
Rice and Arabidopsis are representative model plants in the plant kingdom, and we conducted synteny analysis among cucumber, Arabidopsis, and rice to reveal their evolutionary relationships and genetic changes (Fig. 3b). We identified 11 gene pairs between cucumber and Arabidopsis, and 12 gene pairs between cucumber and rice, a similarity in number that may imply a certain degree of conservation and similarity between cucumber and Arabidopsis, as well as between cucumber and rice. Gene duplication is usually caused by repeat events in the genome, such as whole-genome duplication, segmental duplication, etc., and it can help us understand the diversity and conservation of gene functions[40,41]. Through intraspecific collinearity analysis, we further explored segmental duplication events of CsLRX genes in cucumber. We identified one pair of duplicated genes in the cucumber genome: CsLRX1 and CsLRX2, which provides the potential for the diversity and emergence of new gene function through duplication (Fig. 3c).
Cis-acting elements analysis of Cucurbitaceae LRX gene promoter regions
-
To better understand the transcriptional regulation and potential functions of LRX cucumber (C. sativus), wax gourd (B. hispida), watermelon (C. lanatus), pumpkin (C. maxima), moschata pumpkin (C. moschata), bottle gourd (L. siceraria), and melon (C. melon) genes, we predicted cis-regulatory elements in their promoters. Core elements of promoters such as TATA-box and CAAT-box were found in all LRX genes. The functional cis-regulatory elements in the promoters can be mainly divided into three categories, including biotic and abiotic stress response, plant hormone response, and plant growth and development (Fig. 4). Multiple stress-responsive element (SRE) binding promoter elements ARE were found in the promoters of 38 LRX genes, consistent with the function of LRX as a cell wall protein involved in maintaining cell wall integrity to reduce cell damage caused by external stress. Multiple TAACCA (CCAAT-box) elements, which is bound by transcription factors and regulate gene transcription activity and stress response processes were also discovered[42]. Additionally, low-temperature-responsive (LTR) elements and stress responsiveness (TC-rich repeats) elements were found in individual LRX genes. Interestingly, there are MYB elements in the LRX genes of seven cucurbits, and some MYB transcription factors are involved in the response of plants to abiotic stresses such as drought, high salt, and low temperature. They enhance plant tolerance by regulating gene expression related to stress response[31]. Numerous hormone-related elements were also discovered in the promoter regions of these LRX genes, including multiple ethylene response elements (ERE) in 34 LRX genes and other hormone response elements such as methyl jasmonate response motifs, including CGTCA and TGACG; gibberellin response elements, such as GARE and P-box. LRX genes also contains plant development-related elements, such as G-box. There are also some widely functional cis-regulatory elements such as the binding site W-box for the WRKY family of transcription factors, which are involved in regulating various biological processes including growth and development, hormone signal transduction, and stress response[43,44]. All of the above indicate that LRX plays a crucial regulatory role in physiological processes in plant growth and development, abiotic stress, and hormone signaling.
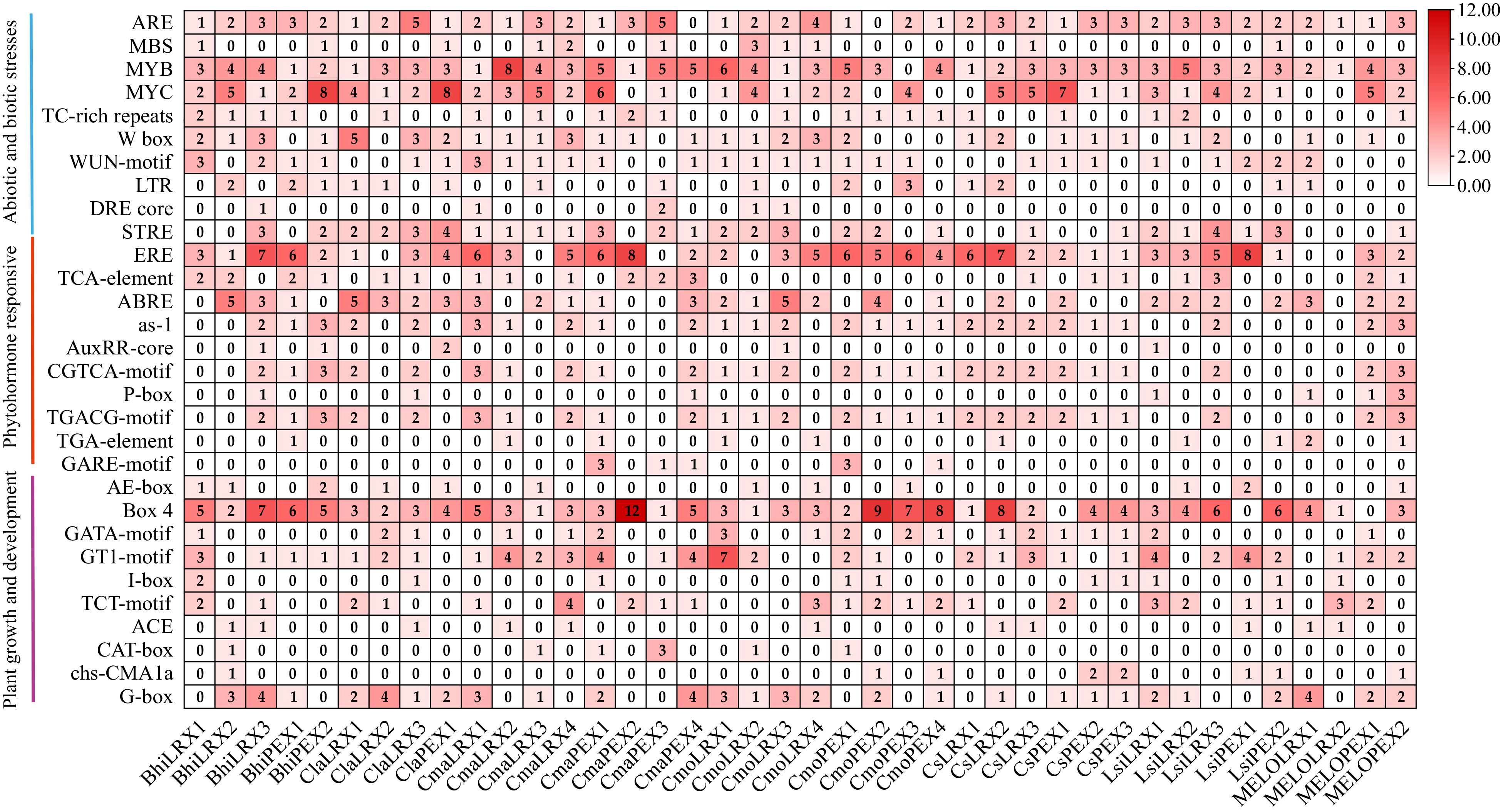
Figure 4.
Cis-acting elements in the promotors of LRX genes in seven cucurbit species. The colors indicate the different cis-elements numbers. Values indicate the statistical number of cis elements.
Expression analysis of LRX genes in Cucurbitaceae different tissues and qRT-PCR analysis in cucumber
-
We plotted the heat map based on the published RNA-seq data (Fig. 5a). CsLRX1 in cucumber was specifically expressed in stems, CsLRX2 was highly expressed in stems, and was also highly expressed in male flowers, roots and ovaries. CsLRX3 was specifically expressed in roots, while CsPEX1, CsPEX2 and CsPEX3 were specifically expressed in male flowers. These findings are consistent with previous reports that LRX is highly expressed in vegetative organs and PEX is highly expressed in reproductive organs[15,17]. The CmoLRXs in moschata pumpkin were specifically expressed in different tissues. The expression of CmaLRXs in pumpkin was low in fruit and specifically expressed in leaves, stems, and roots. In bottle gourd, LsiLRXs had low expression in leaves and specific expression in other tissues. MELOLRX2, MELOPEX1 and MELOPEX2 are strongly expressed in male flowers of melon, indicating that they play an important role in male flower development.
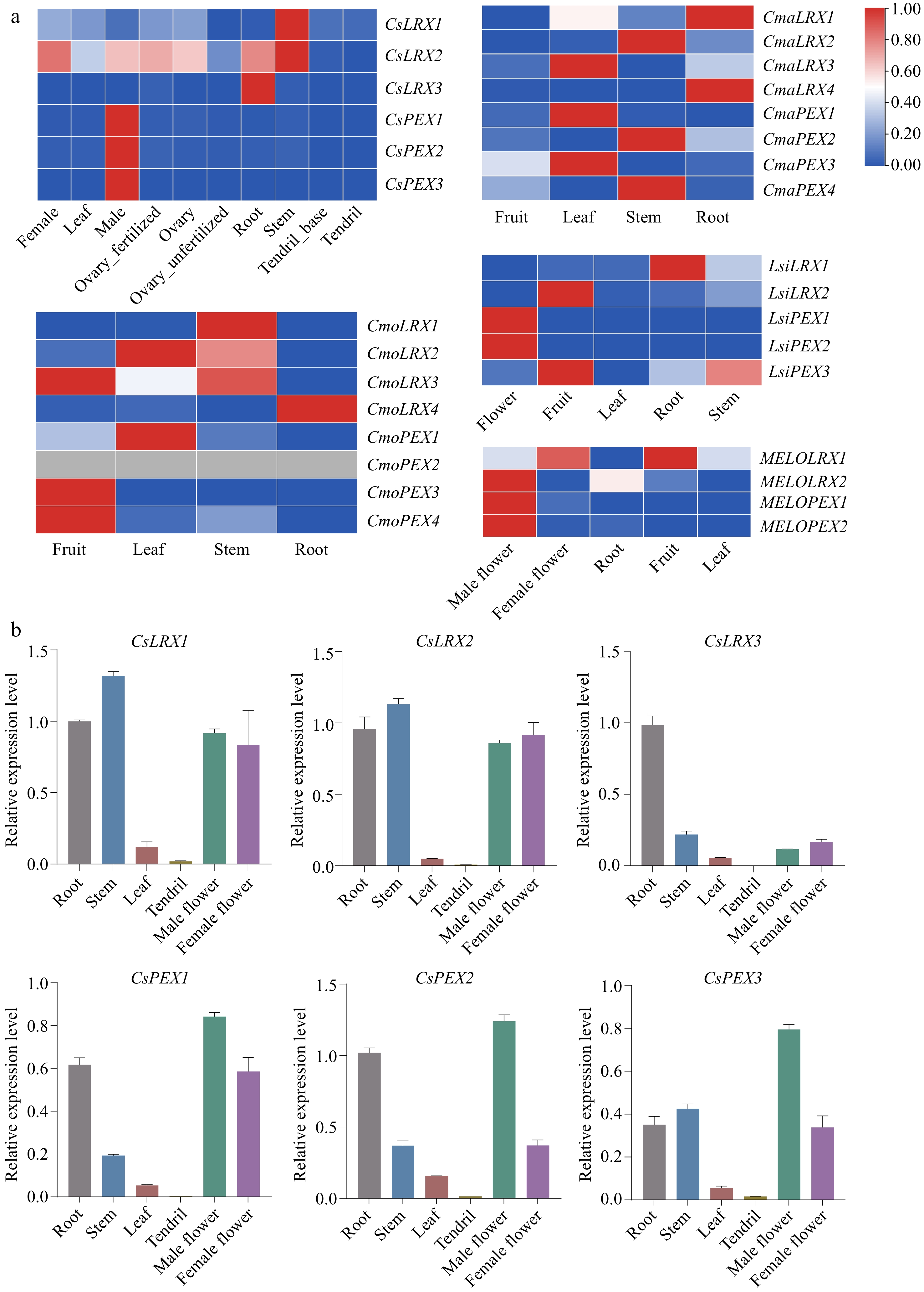
Figure 5.
The expression pattern of LRX genes in different Cucurbitaceae plant tissues. (a) The expression of public RNA-seq data of LRX genes in different tissues of Cucurbitaceae. (b) qRT-PCR analysis of CsLRX genes in different tissues of cucumber. Values are means ± SD of three biological replicates.
Furthermore, we further validated the expression patterns of 6 CsLRX genes in different cucumber tissues (root, stem, leaf, tendril, male flower, female flower) using qRT-PCR (Fig. 5b). The data results showed minor differences compared to the public RNA-seq data, which could be attributed to variations in plant growth environment, sampling methods, and sampling time. However, the overall expression patterns were similar. For example, CsLRX1 and CsLRX2 genes showed relatively high expression in the stem as well as in the root, male flower, and female flower, while CsLRX3 exhibited specific expression in the roots. CsPEX1 to CsPEX3 genes were highly expressed in male flowers, suggesting functional redundancy in regulating male flower development. Additionally, the expression levels of CsLRX genes in cucumber leaves and tendrils were relatively low, especially with almost undetectable levels in tendrils. In conclusion, our results suggest that these genes play a prominent role in the growth of plant vegetative organs and in regulating male flower development.
CsLRX genes respond to salt stress and drought stress in cucumber
-
Plants need to regulate growth and respond to stress by sensing and transmitting cell wall signals. In Arabidopsis, LRX3/4/5 are involved in plant responses to salt stress[12]. To explore whether the CsLRX genes in cucumber are induced by salt stress and drought stress, the roots of cucumber were treated with 100 mM NaCl and 10% PEG6000, and sampled at 0, 3, 6, 9, 12, and 24 h for qRT-PCR to detect its expression (Fig. 6a). The results showed that CsLRX1, CsLRX2 and CsLRX3 involved in vegetative growth responded to salt stress and drought stress. Under salt stress, CsLRX1, CsLRX2 and CsLRX3 all showed a downward trend at 3 h, and then continued to rise to the highest value and then showed a downward trend. The difference is that CsLRX1 and CsLRX2 have the highest expression at 12 h, while CsLRX3 has a peak at 9 h (Fig. 6b). They may help plants maintain the integrity and stability of cell walls in high-salt environments by regulating the morphological and mechanical properties of cell walls. Under drought stress, they all showed an upward trend (Fig. 6c). In fact, cucumbers had wilted after 9 h of treatment, indicating that these genes played an important regulatory role in cucumber response to drought stress. The different expression patterns of LRX genes in cucumber under salt stress and drought stress reflect the different physiological response strategies of plants to these two different types of abiotic stresses. The expression changes of these genes reveal that plants optimize their survival strategies by regulating the expression of cell wall proteins in the face of environmental stress.
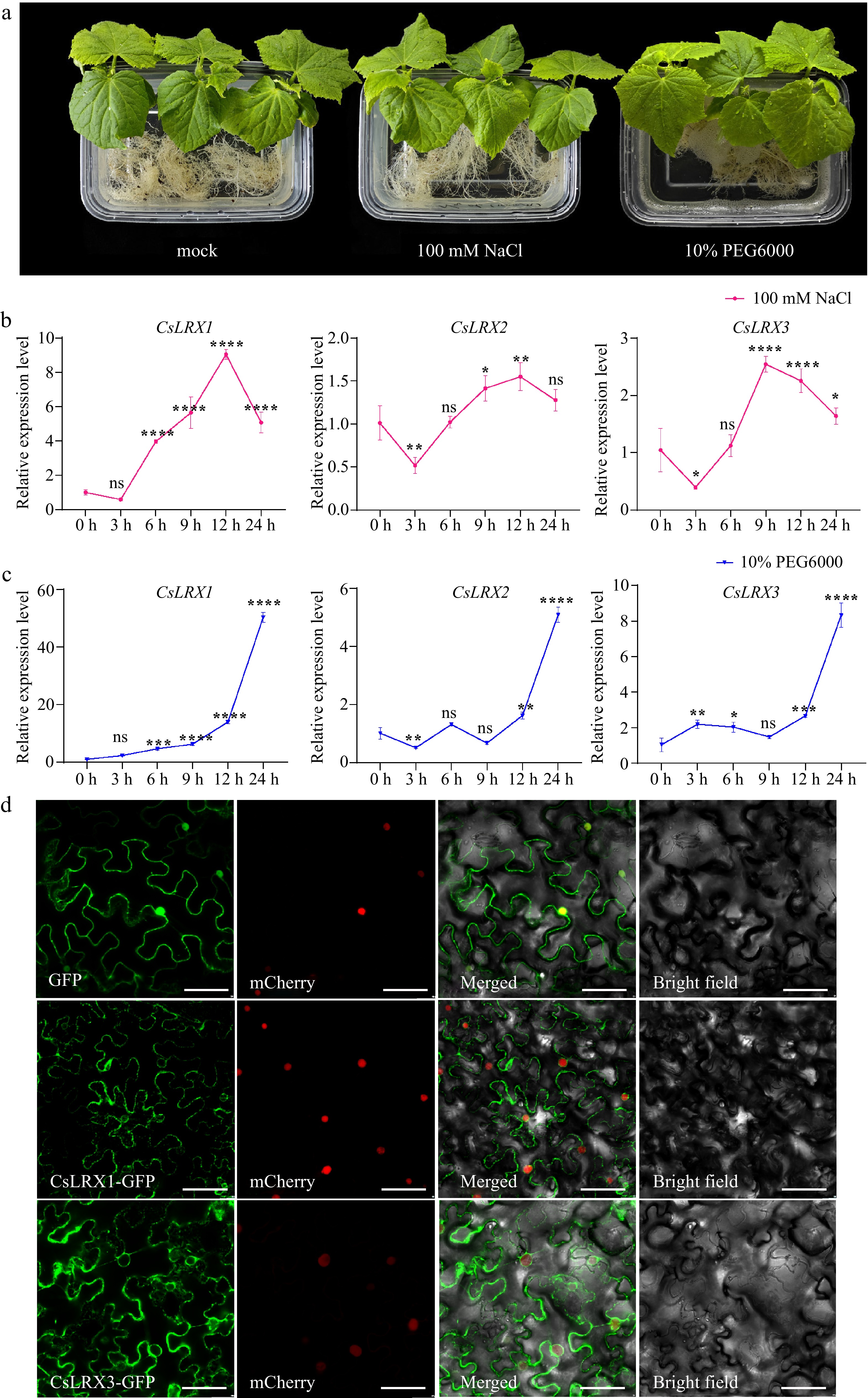
Figure 6.
Expression analysis of CsLRX genes in cucumber. (a) Cucumber roots were treated with 100 mM NaCl and 10% PEG6000, and water was used as a control (mock). (b)−(c) The CsLRX genes in cucumber roots treated with 100 mM NaCl and 10% PEG6000 for different time was analyzed by qRT-PCR. Values are means ± SD of three biological replicates. Significant differences between 0 h and other time points are indicated by asterisks (ns: no significance, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, Student's t test). (d) Subcellular localization of CsLRX1 and CsLRX3. The empty vector was used as a control. These indicated structures were transiently expressed in N. benthamiana leaves. Bar, 50 μm.
To detect the subcellular localization of these genes, the stop codon of these three genes were removed in cucumber and constructed into p1300 vector containing the GFP tag, and transiently expressed in N. benthamiana leaves. Because the CDS sequence of CsLRX2 contains many tandem repeat structures, it is impossible to clone the complete CDS region. Therefore, only CsLRX1 and CsLRX3 proteins were explored. The results showed that GFP fluorescence was observed in the cell wall, cell membrane, and cytoplasm (Fig. 6d). This is consistent with the fact that LRX protein is a kind of plant cell wall protein, which participates in the growth and development of plants by regulating the morphology and properties of the cell wall[10].
Interaction network prediction of CsLRX protein in cucumber
-
To further explore the biological function of CsLRX protein in cucumber, this study used the STRING database to search and analyze its potential interacting proteins. It has been found that many proteins containing the LRR domain may interact with CsLRX protein. The LRR domain has attracted much attention due to its key role in the interaction between various proteins, suggesting that CsLRX protein may play a role in many aspects of plant growth. In addition, according to the expression pattern of the LRX gene, it can be divided into LRX proteins expressed in the vegetative growth stage and PEX proteins expressed in the reproductive growth stage[34]. In particular, most of the proteins interacting with LRX proteins are closely related to the maintenance of cell wall integrity. For example, RLP12 (Csa7G032260), a member of the receptor-like protein family is crucial for the jasmonic acid (JA) signaling pathway induced by coronatine and is involved in the maintenance of cell wall integrity[45]. On the other hand, proteins interacting with PEX proteins are mainly involved in anther development. BAME1 (Csa6G425100) kinase, for example, affects cell division and differentiation by regulating intercellular communication during early anther development, thereby forming cell layers[46]. In summary, CsLRX protein may play a key role in regulating plant vegetative growth and reproductive growth by interacting with proteins containing the LRR domain.
-
The LRX genes play a significant role in plant cell wall formation and plant growth and development[33]. The proteins encoded by LRX genes contain abundant leucine-rich repeat sequences, which form a helical structure and participate in cell-cell interactions and signal transduction[10], regulating the synthesis and remodeling of plant cell walls[5]. LRX genes also possess a unique Extensin domain, rich in amino acids such as leucine, glutamic acid, and cysteine forming a cross-linked polymer that contributes to the formation and stability of plant cell walls[5]. They are chimeric proteins that are insoluble in the cell wall and form a protein-protein interaction platform[6]. LRX protein regulates cell wall expansion by binding to RALF peptide hormones and directly interacts with the transmembrane receptor FERONIA, thereby participating in cell growth regulation[6] In addition, LRX protein also plays an important role in pollen tube growth and pollen germination. Especially in Arabidopsis, LRX protein interacts with RALF4/19 polypeptide to control the integrity and growth of pollen tubes[18,33]. Studies have shown that mutations in the LRX protein can lead to defects in cell wall structures such as root hairs, further confirming its importance in cell wall development[18]. Additionally, LRX genes are involved in responses to stress conditions such as salt stress. In Arabidopsis, LRX has been reported to be involved in regulating various biological processes, including root hair development and resistance to salt stress[12]. Therefore, LRX protein plays an indispensable role in proper plant development and growth regulation.
Cucurbitaceae crops include many important fruit and vegetable that are widely distributed in tropical and subtropical regions and have high economic value[23]. Cucurbit species have various forms and uses, such as food, medicinal, and ornamental purposes, and are extensively cultivated and utilized[25]. In recent years, with the development of high-throughput sequencing technologies, genomic research on cucurbits has been conducted, and the genome sequences of several cucurbit crops have been published[47−52]. Furthermore, ongoing genomic studies are being conducted on other cucurbit crops such as bitter gourd and Buddha's hand fruit[53]. The release of these genome sequences will facilitate a deeper understanding of the genetics, biology and agricultural applications of cucurbit plants. Research on Cucurbitaceae crops has received significant attention, involving various aspects such as growth and development, stress resistance, gene regulation, and more. However, there is currently no reported research on LRX genes in the Cucurbitaceae family. In this study, we identified and characterized the LRX genes in cucurbit plants based on these published whole-genome sequences. We identified 6, 5, 4, 8, 8, 5, and 4 LRX genes in cucumber, wax gourd, watermelon, pumpkin, moschata pumpkin, bottle gourd, and melon, respectively (Table 1). The distribution of these genes on chromosomes is random (Supplemental Fig. S1). Similar to the 11 LRX genes in Arabidopsis, we found that the LRX genes in these seven cucurbit species are also divided into two subfamilies, LRX and PEX (Fig. 1). It is worth noting that the number of LRX genes in these seven cucurbit species vary, but they are less than LRX genes in Arabidopsis. It is speculated that the differences in the number of LRX genes between different cucurbit species may be related to gene duplication or loss during the evolutionary process, as segmental and tandem duplications promote the expansion of gene families[40]. These LRX genes have highly similar structures, with 1 to 2 exons, and the majority do not have introns (Fig. 2). Most LRX genes share conserved motifs Motif1 to Motif7, mainly located at the N-terminus. Motif10 appears most frequently. In general, the repeated occurrence of conserved motifs may indicate their crucial role in regulating gene expression or protein structure, suggesting their importance and diversity in gene function. Conserved domain analysis revealed an LRR domain at the N-terminus and an extensin domain at the C-terminus (Fig. 2), which is consistent with previous reports[33].
To better understand the amplification patterns of genes during the evolutionary process, synteny analysis of LRX genes were conducted in cucumber and six cucurbit species were 4, 6, 8, 7, 6, and 6, respectively (Fig. 3a; Supplemental Table S2). Interestingly, CsLRX1 and CsLRX2 in cucumber have collinearity with LRX genes of six other cucurbits, and there are two pairs of collinearity gene pairs with melon, watermelon, bottle gourd, and pumpkin respectively. Furthermore, CsLRX1 and CsLRX2 replicated each other in cucumber (Fig. 3c), promoting the expansion of the cucumber LRX gene family[40]. These differences in quantity may reflect various genetic changes, including gene rearrangements, insertions, or deletions, that these cucurbit species experienced during the evolutionary process[54]. These differences may be associated with genetic diversity in growth, development, stress resistance, and quality traits. The cis-acting elements on the promoter can reveal the mechanism of gene expression regulation, cell signal transduction network, and the influence of gene-environment interaction[55]. Generally, transcription factors bind to specific cis-acting elements to regulate the expression of target genes. The analysis of cis-acting elements in the promoters of 40 LRX genes showed that these genes may be related to the growth and development of Cucurbitaceae plants, phytohormone response, and response to biotic and abiotic stresses. A large number of stress response elements and hormone response elements were identified, such as ARE, ERE, GARE, P-box and so on. These elements have been reported to be directly involved in plant growth and development and stress (Fig. 4).
LRX gene plays an important role in plant response to drought stress and salt stress. The three genes LRX3, LRX4 and LRX5 in the LRX protein family are essential for plant salt tolerance. When these three genes were mutated at the same time, the plants showed a phenotype of short growth and were very sensitive to salt stress[12]. This indicates that the LRX gene plays a role by participating in the regulation of plant growth and salt stress response, and works together with RALF22/23 and FER[14]. The present results also indicate that CsLRX in cucumber responds to salt stress (Fig. 6b). In addition, the LRXs-RALFs-FER module controls plant growth and salt stress response by regulating a variety of phytohormone. Cell wall integrity is a key factor affecting cell wall integrity and determines the expression pattern of stress response genes. Although there is little evidence directly mentioning the response of LRX genes to drought stress, there is evidence that achieving the best balance between stress response and plant growth is essential for survival in the field environment[56]. Considering the role of LRX genes in regulating plant growth and responding to salt stress, it can be speculated that these genes may also play a role in plant response to drought stress, our results have confirmed that the CsLRX gene in cucumber is induced by drought stress (Fig. 6c). Interestingly, the expression trend of LRX genes was not consistent under salt stress and drought stress (Fig. 6b, c). Under salt stress conditions, plants first enhance the integrity of the cell wall by increasing the expression of cell wall proteins to resist salt-induced cell damage. This may be the reason why the expression of LRX genes increased first. Over time, if plants fail to effectively eliminate accumulated salt or other harmful substances, their physiological state may deteriorate, resulting in a decrease in the expression of cell wall proteins, thereby showing a downward trend in LRX genes expression[14]. In contrast, under drought stress, plants need to quickly adjust their physiological state to adapt to the water shortage environment. In this case, plants may continue to increase the expression of cell wall proteins to maintain the integrity of the cell wall and maintain water. Since drought stress is usually more severe, plants may not be able to return to normal physiological state in time, so the expression of LRX genes continues to rise throughout the stress process[12,14]. In summary, plants adapt to these challenges by regulating the expression of cell wall proteins in the LRX gene family when facing different types of environmental stresses.
Finally, the STRING database was used to search and analyze the potential interacting proteins of CsLRX in cucumber (Fig. 7). Many proteins containing LRR domain had the possibility of interacting with CsLRX protein. The LRR domain has attracted much attention due to its key role in a variety of protein-protein interactions, which may involve a variety of biological functions, including but not limited to signal transduction, cell death, and response to environmental stress. These functions may make CsLRX protein play an important role in cucumber growth and development and response to environmental stress. Through these studies, we not only improved the understanding of the genome structure of Cucurbitaceae, but also provided an important molecular basis for the resistance improvement of cucumber.
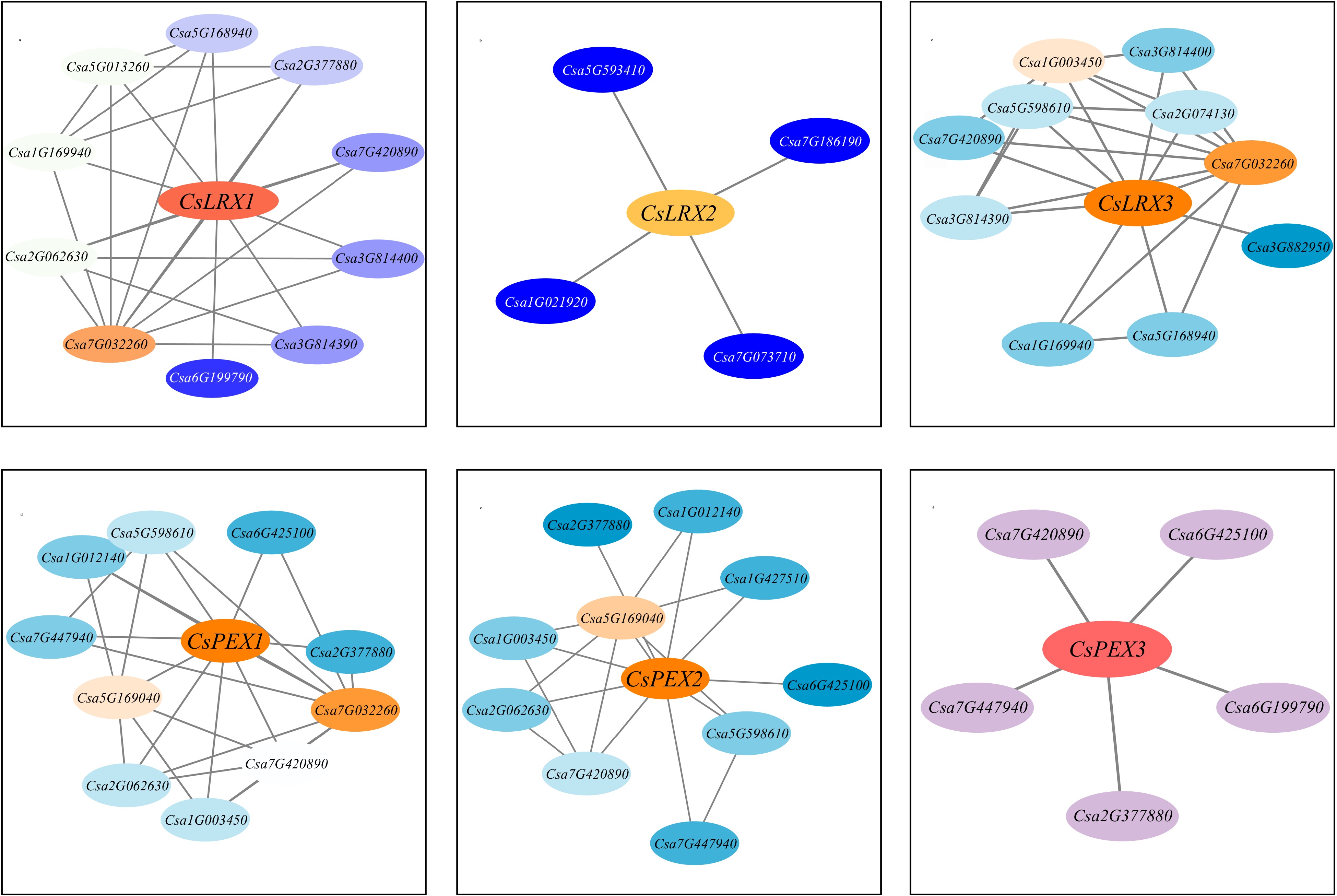
Figure 7.
Interaction network prediction of CsLRX protein in cucumber.
-
In this study, 40 LRX genes were first identified from seven cucurbit species. A comprehensive analysis of these genes were conducted, including chromosome localization, gene structure, conserved motifs, conserved domains, cis-regulatory elements, evolutionary relationships, and gene duplications. The results revealed that LRX genes in the Cucurbitaceae family can be classified into two subfamilies. Gene duplications were observed in CsLRX1 and CsLRX2 genes in cucumber as well as LRX genes in the other six cucurbit species, leading to an expansion of the LRX gene family in these plants. Additionally, CsLRX1 and CsLRX2 genes underwent reciprocal duplication within the cucumber species. Promoter cis-regulatory element analysis suggested that these LRX genes may participate in plant growth and development, phytohormone responses, as well as responses to biotic and abiotic stresses. In addition, the CsLRX genes in cucumber are involved in the response to salt stress and drought stress. The present study provides crucial insights and references for further understanding the functional roles of LRX genes in cell wall formation and growth and development processes in Cucurbitaceae crops.
-
The authors confirm contribution to the paper as follows: study conception and design: Fan S; writing the first version of the manuscript: Fan S; manuscript revision: Yang S, Shi K, Yang L, An M, Wang F, Qi Y, Feng M, Wang M, Gen P, Liu X, Ren H. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this article and its supplementary information files.
This study was supported by the National Key Research and Development Program 'Strategic Science and Technology Innovation Cooperation' Key Special Project (2023YFE0206900).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primers of cucumber CsLRX genes used in qRT-PCR.
- Supplemental Table S2 The orthologous relationship of LRX genes in cucumber and other six cucurbit species.
- Supplemental Fig. S1 Location distribution of LRX genes on chromosomes of seven cucurbit species. The leftmost is the chromosome length scale. The chromosome is represented by a bar in the figure, the length represents the size of the chromosome, and the color depth represents the gene density. The corresponding position of LRX genes on chromosome was marked with red.
- Supplemental Fig. S2 10 Conserved Motifs of LRX Proteins from seven cucurbit species. The height of the letter indicates the frequency of the base in each Site Count sequence.
- Supplemental Fig. S3 The conserved domains of LRX proteins in seven cucurbit species.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fan S, Yang S, Shi K, Yang L, An M, et al. 2024. Genome-wide identification of the LRX gene family in Cucurbitaceae and expression analysis under salt and drought stress in cucumber. Vegetable Research 4: e026 doi: 10.48130/vegres-0024-0025 

Genome-wide identification of the LRX gene family in Cucurbitaceae and expression analysis under salt and drought stress in cucumber
- Received: 07 May 2024
- Revised: 06 June 2024
- Accepted: 12 June 2024
- Published online: 07 August 2024
Abstract: Leucine-Rich Repeats Extensins (LRX) are a type of cell wall protein that participate in the formation of the plant cell wall and play a crucial role in plant growth and development. However, the study of LRX genes in Cucurbitaceae has not been reported. Here, 40 LRX genes were identified from seven cucurbit species in the Cucurbitaceae database, including cucumber (Cucumis sativus), wax gourd (Benincasa hispida), watermelon (Citrullus lanatus), pumpkin (Cucurbita maxima), moschata pumpkin (Cucurbita moschata), bottle gourd (Lagenaria siceraria), and melon (Cucumis melon). These LRX genes were divided into two subfamilies: LRX, controlling vegetative growth, and PEX, controlling reproductive growth. The gene structure, domain, and motif were relatively conserved, indicating that genes within each subfamily have similar functions. The differences in the number of LRX genes among the seven cucurbit species indicate the presence of gene loss or duplication events during evolution. Analysis of cis-regulatory elements showed that these LRX genes may be involved in plant growth and development, phytohormone response, and biotic and abiotic stress responses. In addition, the expression pattern of CsLRX genes in different tissues of cucumber and its expression analysis under salt stress and drought stress were detected by real-time quantitative PCR (qRT-PCR). The results showed that CsLRX genes showed organ-specific expression pattern in cucumber and responded to adversity stress. In summary, the results of the study provide a reference for further understanding the role of these genes in cell wall formation and the growth and development of Cucurbitaceae crops.
-
Key words:
- Cucurbitaceae /
- LRX genes /
- Cell wall protein /
- Expression patterns /
- Cucumber