









-
Chloroplasts are vital cellular components for photosynthesis in plants and other eukaryotic photosynthetic organisms[1]. Chloroplast development is a complex process regulated by many genes and that can be divided into three stages: the replication of plastid and plastid genomes; the establishment of genetic and translational systems, during which the chloroplast genome is mainly transcribed by nucleus-encoded RNA polymerases (NEPs); and the establishment of the chloroplast photosystem, which involves the primary transcription of photosynthesis-related genes by plastid-encoded RNA polymerases (PEPs), consisting of core subunits encoded by the rpoA, rpoB, rpoC1, and rpoC2 genes in the plastid, along with other nuclear-encoded components[2,3]. Notably, mutations in the rpoA, rpoB, rpoC1, and rpoC2 genes can impair chloroplast development, leading to a mutant leaf color phenotype.
To date, many genes related to chloroplast development — including AtPDS (Arabidopsis phytoene desaturase), a gene related to photosynthetic pigment synthesis that encodes phytoene desaturase, a key enzyme in the carotenoid synthesis pathway — have been identified[4]. The mTERF3 (mitochondrial transcription termination factor3) protein encoded by the SL1 (seedling lethal1) gene interacts with three subunits of the PEP complex, suggesting that it may be active in chloroplast gene expression, and in Arabidopsis, the sl1 mutant shows severe deficiencies in chloroplast development and photosystem assembly[5]. The OsSLC1 (Oryza sativa seedling-lethal chlorosis1) protein, which belongs to the PPR family, is preferentially expressed in chloroplasts and plays a vital role in intron splicing[6]. In asl4 (albino seedling lethality4) mutants, the transcription levels of genes related to chlorophyll biosynthesis, photosynthesis and chloroplast development are severely inhibited, and loss of ASL4 gene function can lead to chloroplast development defects and seedling death[7].
In Arabidopsis, the chloroplast-defective mutant mrl7/ecb1/svr4/rcb exhibits albino cotyledons, cannot grow true leaves, and dies shortly after germination. The MRL7/ECB1/SVR4/RCB gene has been mapped to At4g28590 through T-DNA insertion identification and gene localization[8−11]. The thioredoxin-like fold protein with disulfide reductase activity encoded by MRL7/ECB1/SVR4/RCB is localized in chloroplasts and the nucleus and is a dual-target nuclear/plastidomal photosensitive pigment signaling component necessary for PEP assembly[12]. MRL7/ECB1/SVR4/RCB has not been identified in the PEP complex, but it can interact with the three PEP complex subunits TRXz, FSD2, and FSD3, which implies that it can regulate the expression of photosynthesis-associated plastid-encoded genes (PhAPGs) through interactions with the PEP complex subunits to participate in chloroplast development[10,13]. Chloroplast-deficient mutants have also been found in Brassica crops. The yellow-green leaf (yvl) mutant was isolated from Brassica napus. The causal gene BnaA03.CHLH was identified as encoding the putative H subunit of Mg-protoporphyrin IX chelatase (CHLH)[14]. Xu et al. reported a lethal mutant, 7-521Y, with cotyledon yellowing and determined that this trait was controlled by two genes. One of the candidate genes was located at the 29 kb region of chromosome C06, and BnaC06. FtsH1, which regulates the PSII repair cycle in B. napus, was confirmed to be a target gene by complementary functional verification[15]. A chlorophyll-reduced mutant (crm1) of rapeseed was generated by EMS mutation. Two target genes with single nucleotide replacements, BnaA01G0094500ZS and BnaC01G0116100ZS, were identified via BSA sequencing in crm1. These two genes encode the CHLI1 protein, which is critical for chlorophyll synthesis[16].
In this study, a natural lethal albino mutant, 6-219W, from curly kale that exhibited yellow cotyledons and albino hypocotyls was identified. Fine mapping, functional validation, and phylogenetic analysis were performed to explore the target gene BoAP1. The BoAP1 locus was ultimately mapped between two InDel markers, PW404 and PW406, on chromosome 8, and BolC08g019310.2J, an ortholog of the AT4G28590 gene, was the candidate gene for BoAP1. The present research provides new insights into the molecular mechanism of the lethal albino phenotype and the development of chloroplasts in cole crops.
-
6-219G is a curly kale line; its hypocotyl and leaves are green. 6-219W is a natural mutant discovered from the self-crossing of 6-219G offspring and is characterized by white hypocotyl and yellow leaves. NB11 is a cabbage inbred line with normal hypocotyls and leaves. The heterozygous genotype 6-219G was crossed with NB11 to generate F1 plants. The F2 population for genetic analysis was obtained via self-pollination of the F1 progeny.
Chlorophyll detection
-
Seven- to ten-day-old 6-219G and 6-219W seedlings were collected to determine the chlorophyll content. After rapid freezing, the whole seedlings (0.1 g) were ground into powder in liquid nitrogen. The powder was immersed in 10 mL of an extraction solution (anhydrous ethanol : acetone = 1:2 (v/v)) for 4 h under light-protected extraction until the tissue residue at the bottom turned white. The abovementioned samples were centrifuged at 12,000 rpm for 15 min. The supernatant was poured into a quartz colorimeter using the extraction solution as the negative control, and the absorbance of chlorophyll at 645 and 663 nm was measured using the EV-2200 spectrophotometer[17]. Three biological replicates were performed, and the data were analyzed using the following formula[18]:
$ \mathrm{Chla} =0.01 \times (21.2\times {\rm{OD}}_{ {663}} -\,{4.48\times {\rm{OD}}}_{ {645}} )\times {\rm F} \div {\rm W} $ $ \rm Chlb=0.01\times (38.2\times OD_{{645}} -\,7.8\times OD_ {663} )\times F \div W $ $ \rm Chl=0.01\times (33.7\times A_{ 645} +13.4\times A_{663} )\times F \div W $ In the above formula, F represents the dilution factor and W represents the sample weight (g).
Transmission Electron Microscope (TEM)
-
The chloroplast ultrastructures of fresh cotyledons from 6-219G and 6-219W were examined via TEM. The samples were fixed in 2.5% glutaraldehyde at 4 °C for 16 h, and then dehydration in the ethanol series and embedding was carried out[19]. The samples were prepared into slides and observed on the H-7500 transmission electron microscope (Hitachi, Tokyo, Japan).
Whole-genome re-sequencing, primer design, and DNA extraction
-
Whole-genome resequencing was performed for 6-219W and NB11 using the Illumina HiSeq 2500 platform (San Diego, CA, USA). After quality control, filtering, and removal of adapter contamination using Fastp (v0.21.0) software, the BWA (v0.7.12) software was used to align the resequencing data to the Brassica oleracea cv. JZS V2.0 reference genome (
http://brassicadb.cn )[20,21]. Subsequently, SNP and InDel variations were investigated by employing the GATK (v4.x) toolkit[22]. Finally, 117 InDel primers were designed on primer3 (v4.1.0) to be uniformly distributed on the nine chromosomes, with each InDel primer being spaced 3–5 Mb apart[23]. The design principles of the primers were as follows: Tm of 52–60 °C, GC content of 40%–50%, and amplification length of 100–250 bp[24]. All primers used in the present research are listed in Supplemental Table S1. Total genomic DNA was extracted from the two parental lines, the F2 population, and the transgenic plants using the improved CTAB method[25,26].Fine mapping of the BoAP1 gene
-
Sixty-six specific polymorphic markers between the parents, 6-219W and NB11 were identified from the initial 117 genome-wide markers. Both parents and 30 F2 individuals with mutant phenotypes were used to preliminarily identify linked markers. The markers at both ends of the linked interval were used to screen 96 recessive single plants to verify whether the markers were linked. Subsequently, more linked markers were developed within the linked interval, which was used for genotyping all F2 individuals.
Phylogenetic analysis of BoAP1
-
A BLASTP search in the Ensembl Plants database (
https://plants.ensembl.org/ ) was performed to download other homologous protein sequences with the BoAP1 sequence as the query. The above proteins were aligned using DNAMAN version 9.0 to display the conserved domains. A neighbor-joining phylogenetic tree (1,000 bootstrap replications) was constructed using MEGA11.0 software, with the protein sequences aligned using ClustalW.CRISPR/Cas9 genome-editing vector construction and cabbage transformation
-
CRISPR/Cas9 target prediction was carried out by employing an online website (
http://crispor.tefor.net/ ) to select two targets located in the first exon of the BoAP1 gene: sgRNA1 (AGAACTCCGATGCTTCACACCGG) and sgRNA2 (GTCCCGCGTAAACCGAAGCGCGG). The purified fragments were cloned and inserted into zmplCas9i (derived from PC1300) using T4 DNA ligase to construct the pCas9-BoAP1 editing vector. After the successful construction of pCas9-BoAP1 was confirmed by sequencing, the vector was transformed into the recipient material NB11 using Agrobacterium-mediated methods[27].Subcellular localization
-
To identify the subcellular localization of the BoAP1 protein, the full-length coding sequence of BoAP1 without a stop codon was ligated into the linear vector pBWA(V)HS-ccdb-GLosgfp after double enzyme digestion. After confirming the successful construction of the vector, the plasmid was introduced into tobacco leaves through Agrobacterium-mediation protocols. Then, the tobacco plants were cultured under weak light for 2 d and observed for GFP fluorescence using a laser confocal scanning microscope (Nikon C1, Japan).
Gene expression analysis
-
Total RNA from 7-day-old 6-219W and 6-219G plants was extracted using the TIANGEN RNAprep Pure Plant Kit (Tiangen, Beijing, China). The qRT-PCR system was prepared with the Taq Pro Universal SYBR qPCR Master Mix Kit (Vazyme, Nanjing, China) and amplified in the Bio-Rad CFX96TM Real-Time System (Bio-Rad, Hercules, CA, USA). The relative expression of BoAP1 in the samples was calculated according to the 2−ΔΔCᴛ method, and all experiments were repeated three times for analysis[28].
-
The spontaneous albino mutant was derived from curly kale. Phenotypic observation showed that the hypocotyl of the 6-219W mutant was white, the cotyledon was yellow, and the life cycle was incomplete, with a growth period of only 7–14 d (Fig. 1a−c). The chloroplasts in the 6-219G contained a complete membrane structure and stacked thylakoids under transmission electron microscopy (TEM) (Fig. 1d, e). However, the number of chloroplasts in the 6-219W mutant was considerably reduced, the plastoglobuli were on the contrary, and the thylakoid membranes could not be observed (Fig. 1f, g). The chlorophyll a and chlorophyll b levels were significantly lower in the 6-219W mutant than in the 6-219G treatment group (Fig. 1h).
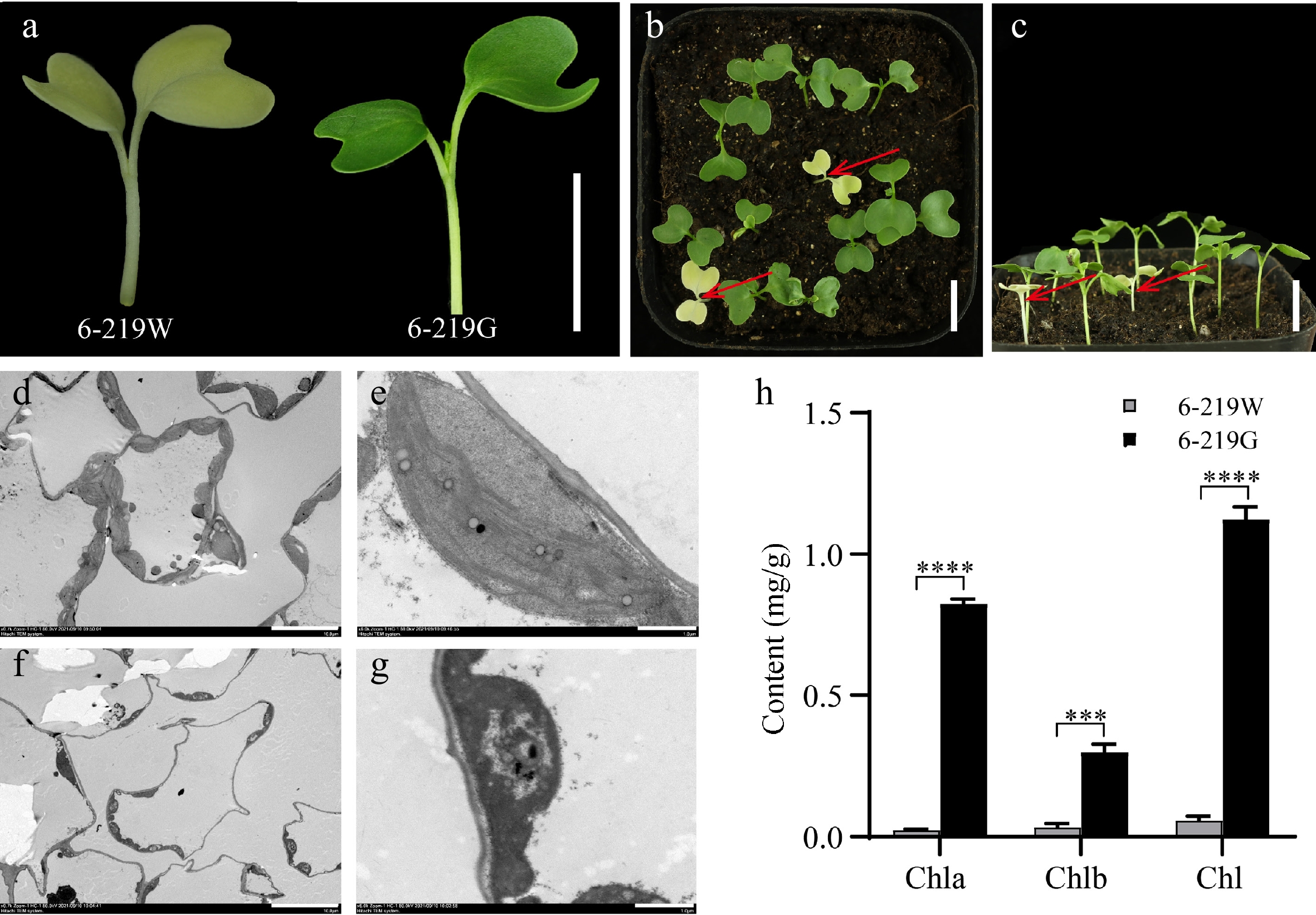
Figure 1.
Phenotypic characterization of wild-type 6-219W and mutant 6-219G. (a) 7-day-old seedlings of 6-219W and 6-219G. (b), (c) 10-day-old seedlings of 6-219W and 6-219G. The red arrows point to 6-219W. (d), (e) Ultrastructure of chloroplasts in 6-219G. (f), (g) Ultrastructure of chloroplasts in the albino 6-219W. Transmission electron microscopy was carried out on 7-day-old leaves of 6-219W and 6-219G. (h) Chlorophyll a (Chla), chlorophyll b (Chlb), and total chlorophyll (Chl) content in the 6-219W and 6-219G groups. The asterisk represents the magnitude of the difference: *** p < 0.001 and **** p < 0.0001. Scale bars: (a)−(c) 1 cm; (e), (g) 10 μm; (f), (h) 1 μm.
Since 6-219W is a lethal albino mutant, heterozygous 6-219G was crossed with NB11, a wild-type cabbage inbred line, to generate the F1 progeny and F2 mapping population. F1 individuals exhibit normal phenotypes. Among the 2,580 plants in the F2 population, there were 652 albino seedlings and 1,928 normal seedlings. The genetic separation ratio between the lethal albino seedlings and the normal individuals was 3:1 (χ2 = 0.101 < χ20.05 = 3.841), demonstrating that a single recessive gene causes this lethal albino trait.
Map-based cloning of the lethal albino gene boap1
-
Based on the genome comparison between 6-219W and NB11, 117 InDel markers evenly distributed across nine chromosomes of cabbage were designed to identify polymorphisms between the parents. A total of 66 markers differed between the two parents. First, 30 albino-lethal individuals were screened with the above 66 differential markers, and we found that the region linked to the albino trait was 12.95 Mb (between InDel markers PW81 and PW304) on chromosome 8 (Fig. 2a). Subsequently, 405 F2 recessive plants with a lethal albino phenotype were mapped based on ten polymorphic InDel markers, indicating a 0.92 Mb region (between the markers PW373 and PW374) on chromosome 8 (Fig. 2a). Finally, eight polymorphic markers were developed 1,642 F2 recessive individuals were selected for fine mapping. The boap1 locus was ultimately identified in an approximately 300 kb region (between the InDel markers PW404 and PW406) on chromosome 8 (Fig. 2a). Three genes related to chloroplast development were determined within the 300 kb candidate region (Supplemental Table S2).
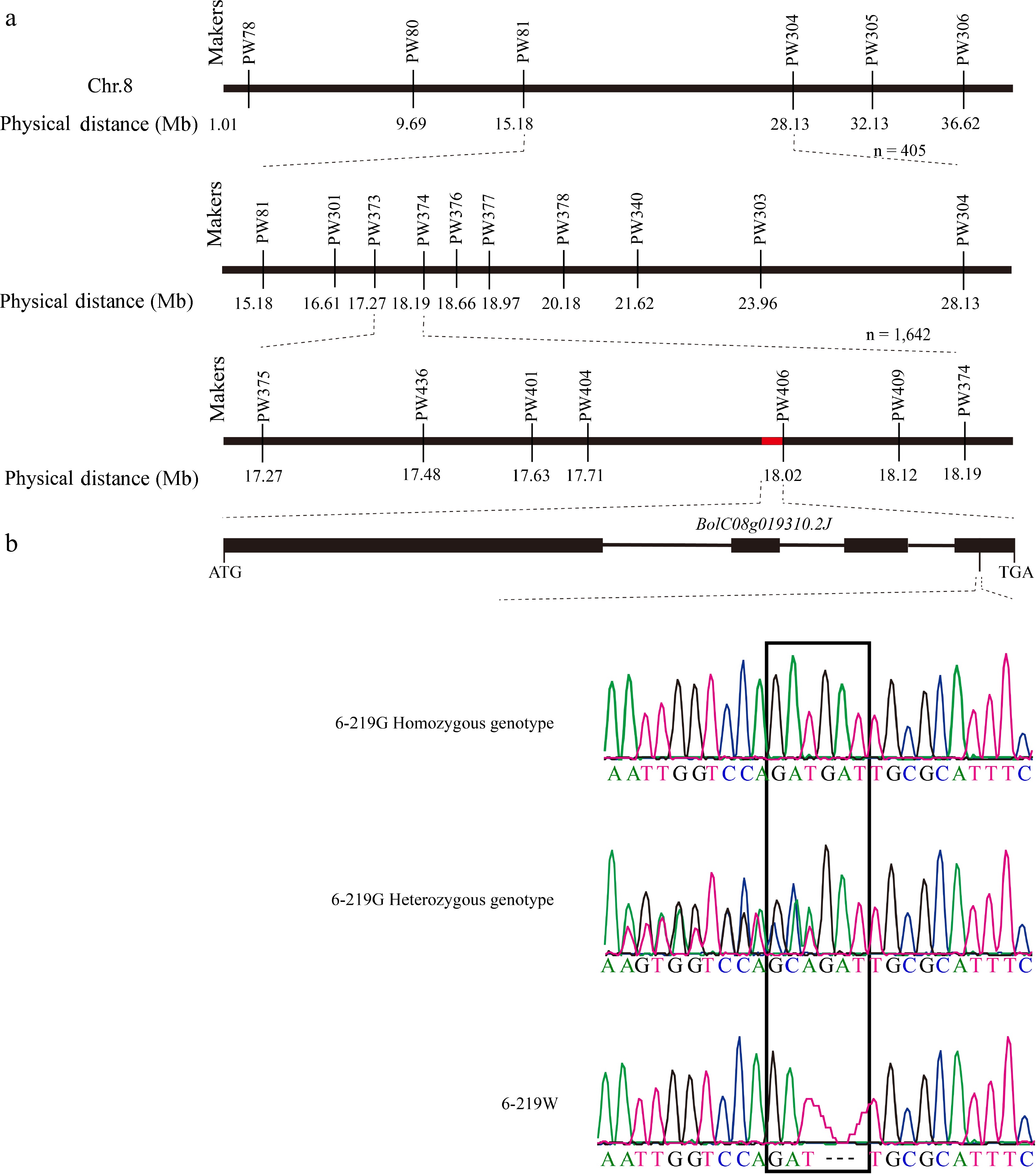
Figure 2.
Map-based cloning of BoAP1. (a) Fine mapping of BoAP1 with InDel markers. The solid red rectangle represents the position of BolC08g019310.2J. (b) The schematic diagram depicts the exons (solid black boxes) and introns (black lines) of BoAP1. The nucleotide sequence and sequence mapping show the point mutation in 6-219W. The black frame represents the location of the boap1 mutation.
The subsequent sequencing analysis of the BolC08g019310.2J gene indicated homozygous deletions of three nucleotides (GAT) in the fourth exon in the 6-219W mutant. In comparison, there were homozygous insertions of three nucleotides (GAT) or heterozygous genotypes in the 6-219G individuals, corresponding to the phenotype (Fig. 2b). The putative protein sequence in the 6-219W mutants had a deletion of a Met at the 298th amino acid position (Supplemental Fig. S1). Protein sequence alignment and conserved motif analysis indicated that the mutation site was located between motif 9 and motif 5 and was conserved in cabbage, Arabidopsis, and Chinese cabbage (Supplemental Fig. S1). Moreover, BolC08g019310.2J shared 77% amino acid sequence identity with AT4G28590, a plastidial phytochrome signaling component in the chloroplast[8]. The above results indicated that deleting the 298th amino acid M might lead to the loss of BolC08g019310.2J function. Therefore, BolC08g019310.2J was designated BoAP1 in this study.
Phylogenetic analysis of BoAP1
-
To elucidate the phylogenetic relationship between BoAP1 and other homologous proteins, the homologous protein sequences of the Brassicaceae, Solanaceae, and Leguminosae families, among others, were downloaded through BLASTP and then a phylogenetic evolutionary tree was reconstructed (Fig. 3b). The results showed that BoAP1 is most closely related to AT4G28590 in Arabidopsis, indicating that BoAP1 plays an essential and conserved role in chloroplast development.
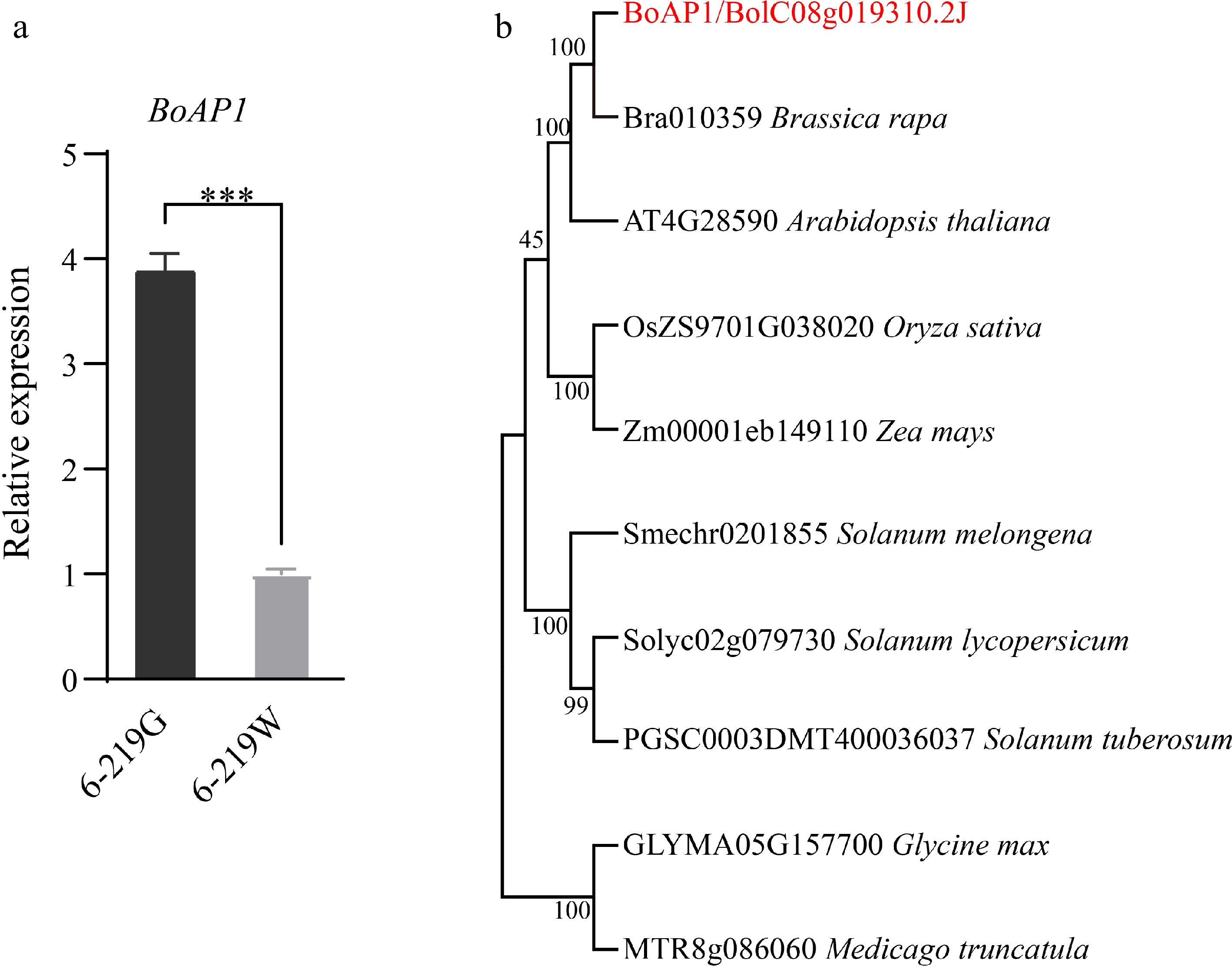
Figure 3.
Gene expression patterns of BoAP1. (a) Expression levels of BoAP1 in the 6-219G and 6-219W. The asterisk represents the magnitude of the difference (***, p < 0.001). (b) Phylogenetic analysis of BoAP1 and its related proteins.
Subcellular localization and expression analysis
-
According to previous reports, AT4G28590 encodes a protein localized in the chloroplast[8], so it was speculated that the protein encoded by BoAP1 is also expressed in the chloroplasts. To verify this prediction, a BoAP1-GFP fusion protein expression vector was constructed and transferred into tobacco leaf cells for transient expression. Laser scanning confocal microscopy showed that the fluorescence signal of the BoAP1-GFP fusion protein overlapped with chloroplast autofluorescence, while the GFP green fluorescence of the empty vector was detected in the nucleus and the cytoplasm (Fig. 4). Taken together, these results indicated that BoAP1 is localized to chloroplasts.
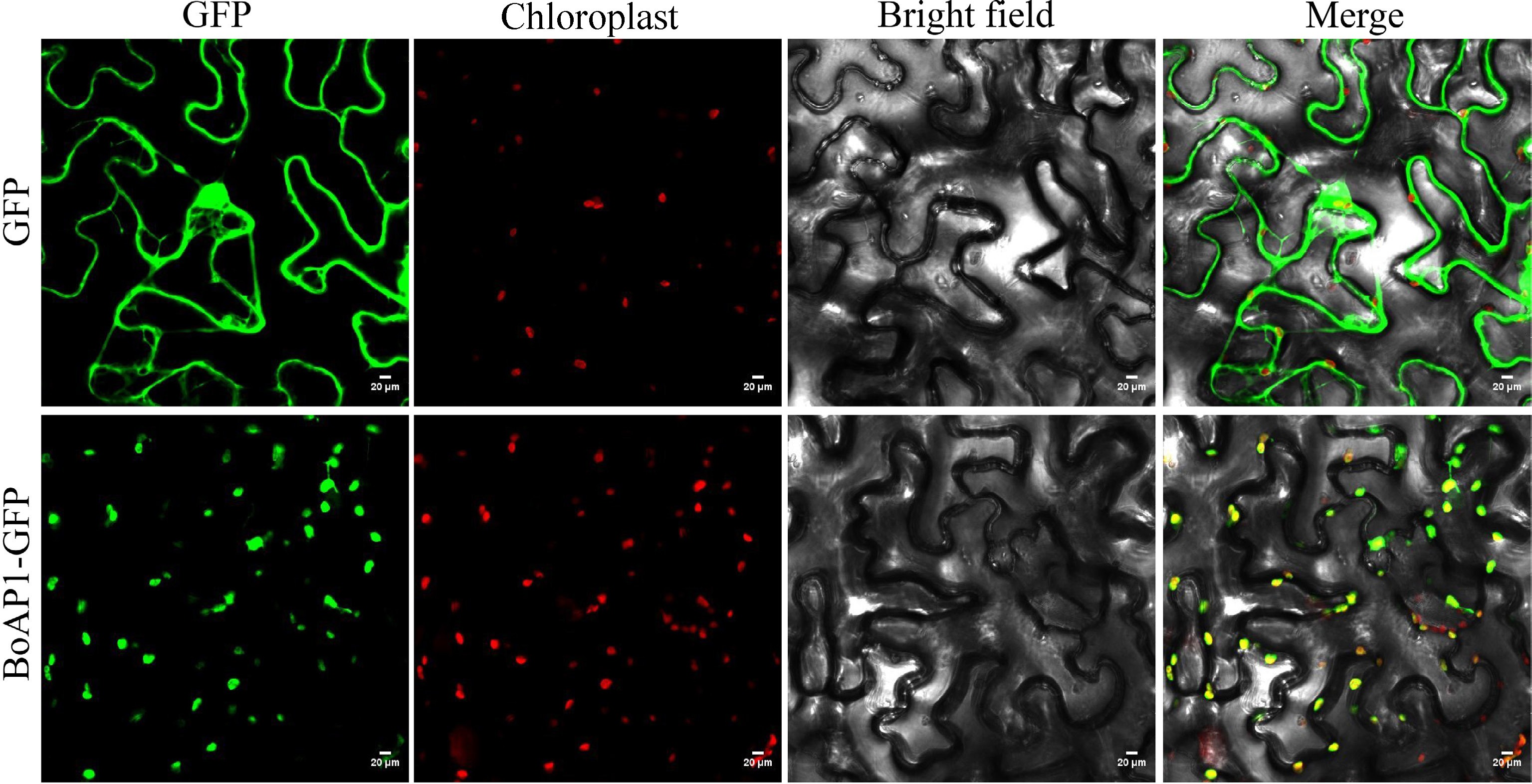
Figure 4.
Subcellular localization analysis of BoAP1. The tobacco leaves were transformed with the empty GFP vector and BoAP1-GFP fusion protein. From left to right: fluorescence of the empty GFP vector and BoAP1; spontaneous fluorescence in chloroplasts; bright-field image; merged image of the first three images. Scale bars = 20 μm.
Quantitative real-time PCR (qRT-PCR) was performed to compare the expression of the BoAP1 transcript in 6-219G and 6-219W. The results indicated that the transcription of BoAP1 was significantly downregulated in the 6-219W mutants, inferring that the expression of BoAP1 is closely related to the albino phenotype (Fig. 3a).
Functional confirmation of BoAP1
-
To confirm whether a functional loss of BoAP1 causes the albinism phenotype, we constructed a pCas9-BoAP1 genome-editing vector with two sgRNAs targeting exon 1 of BoAP1 (Fig. 5a). The pCas9-BoAP1 vector was transferred into wild-type NB11 explants via the Agrobacterium tumefaciens-mediated genetic transformation method. Twenty positive lines were generated by amplifying the Hyg gene. Sanger sequencing analysis of the amplified BoAP1 gene revealed that eight T0-positive transgenic lines were edited and exhibited an albino phenotype (Fig. 5b, c).
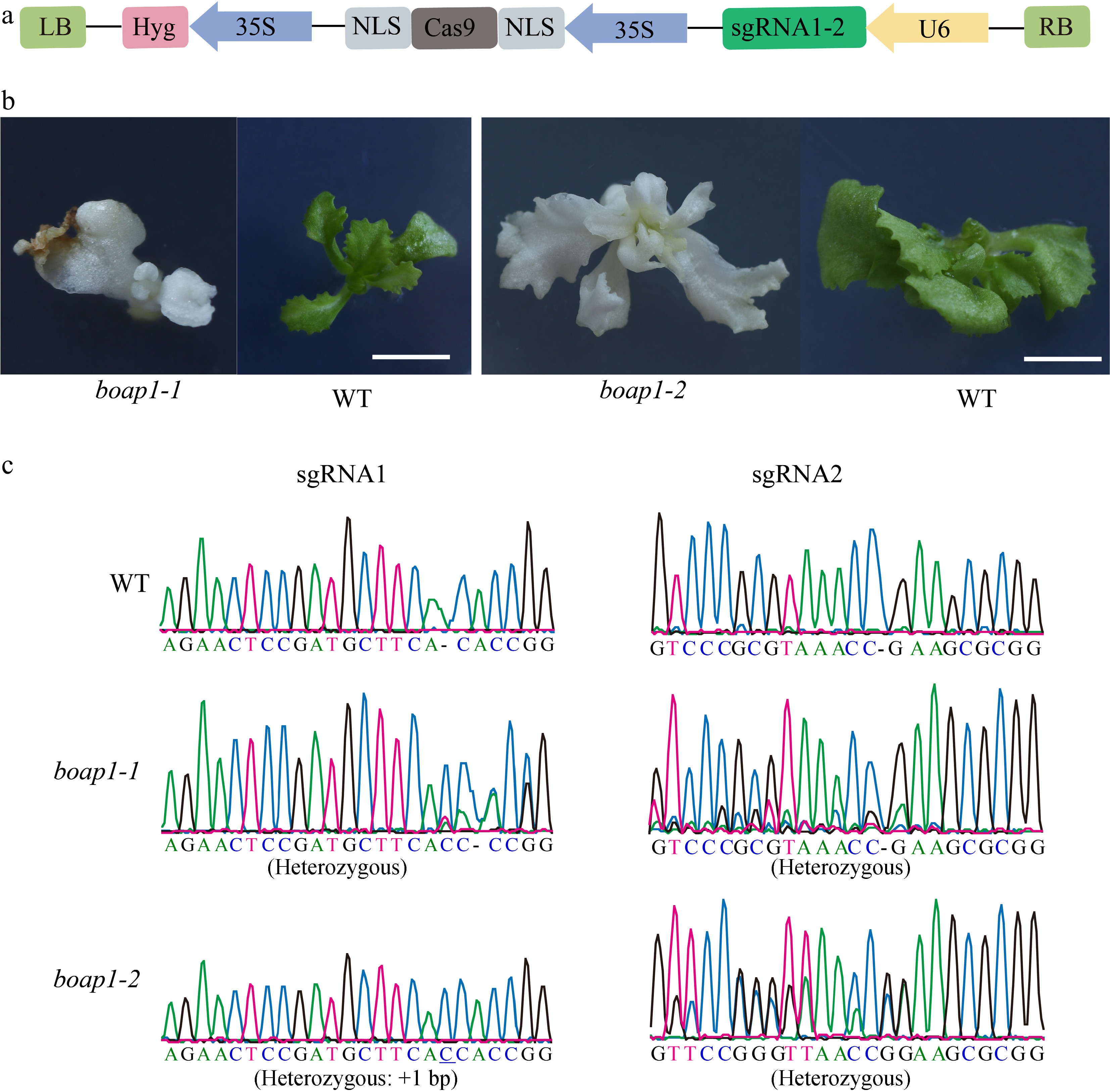
Figure 5.
Functional confirmation of BoAP1. (a) Schematic diagram of the pCas9-BoAP1 genome-editing vector targeting BoAP1. Hyg, hygromycin resistance gene; U6, Arabidopsis U6 promoter. (b) Phenotypes of the WT, boap1-1, and boap1-2. Scale bar: 0.5 cm. (c) Sequence mapping of the WT, boap1-1, and boap1-2 on the target region.
-
Chloroplasts are vital organelles plants need to carry out photosynthesis. They provide organic material and energy for plants and are essential for the growth and development of green plants[29]. Through the in-depth study of chloroplasts, many chloroplast-deficient mutants and corresponding causal genes have been discovered in A. thaliana, Oryza sativa, B. napus, and others, revealing details of the chloroplast growth and development process[5,14,15,30,31]. In this study, a chloroplast-deficient mutant, 6-219W, with yellow cotyledons and white hypocotyls that has an abnormal growth period and survives for only 7–10 d was identified. Its phenotype resembles that of the rapeseed line 7-521Y[15]. An F2 population was used to fine-map the target gene to a 300 kb region on chromosome 8, and through gene sequencing, sequence analysis, and functional verification, it was found that BolC08g019310.2J is a strong candidate for being the gene responsible for the albino trait.
Leaf color is an important target trait in vegetable genetics and breeding, which directly affects its commodity value. Chloroplasts are vital for the shelf life of leafy vegetables. Gene mutations such as BoYgl-2 and BrCAO cause yellow leaf, affecting their yield and commercial value, and shortening their shelf life[32,33]. Mutation in the BrNYM1 gene can maintain leaves greenness of Chinese cabbage during the aging process and extend their shelf life[34]. The full-length introduction of the BoAP1 homologous gene AtMRL7 driven by the 35S promoter into the mrl7 Arabidopsis albino mutant increases the chloroplast number of the mutant and restores the normal phenotype[8]. Therefore, it was speculated that overexpression of the BoAP1 gene might increase the number of chloroplasts and prolong the life and shelf time.
The CRISPR/Cas9 gene editing system is already widely used in plants such as rice, maize, Arabidopsis, tobacco, tomatoes, and cabbage. The lethal albino gene has been successfully applied to test the efficiency of the CRISPR/Cas9 gene editing system because its editing efficiency can be easily observed during the seedling or sprouting stage. CRISPR/Cas9 has been used to knock out the OsPDS gene in rice, and an albino phenotype was observed in the T0 generation[35]. Agrobacterium-mediated CRISPR/Cas9 technology was previously used to knock out the AtPRPL18 and OsPRPL18 genes, and the atprpl18 and osprpl18 mutants exhibited albino seedling phenotypes[31]. Pan et al.[36] and Ma et al.[37] targeted mutations in the PDS homologous genes in tomatoes and cabbage, respectively, and found a high mutation frequency in the T0 transgenic plants. In the present research, a CRISPR/Cas9 editing vector was used to knock out the BoAP1 gene, and the resulting boap1 mutant exhibited a typical lethal albino phenotype. We believe that this gene may be similar to the PDS gene, and the efficiency of various editing systems can be tested by knocking out the homologous genes of BoAP1 in crops.
The PEP complex contains plastid-encoded rpo subunits and other nucleus-encoded proteins, among which FSD2, FSD3, TRXz and FLN1 may play an important role in the redox-mediated regulation of chloroplast development[38−40]. FSD2 and FSD3 are superoxide dismutases[41]. Loss of FSD2 or FSD3 function impairs the growth of Arabidopsis plants and leads to leaf albinism. The fsd2 mutant appears increased superoxide yield, decreased chlorophyll content, and a decreased CO2 assimilation rate, while the fsd3 mutant does not survive beyond the seedling stage[42]. Through CRISPR/Cas9-directed mutagenesis of OsFLN1, a severe lethal albino phenotype was observed in mutants and OsFLN1 was found to regulate the transcription of PEP-related genes and chloroplast growth and development by interacting with OsTRXz[43]. OsTRXz, a chloroplast thioredoxin reductase is indispensable for maintaining the complete structure of the PEP complex and regulating chloroplast biosynthesis. In the present study, the homologous gene BoAP1 of AtMRL7/ECB1/SVR4/RCB was cloned in curly kale. Previous studies have reported that AtMRL7/ECB1/SVR4/RCB can interact with TRXz, FSD2, and FSD3, and that compared to those of untreated RNAi lines, AtMRL7-RNAi lines treated with appropriate concentrations of ROS-related reagents show greater chlorophyll content and a significant recovery of the pale green phenotype, indicating that MRL7/ECB1/SVR4/RCB is involved in the redox-mediated regulation of chloroplast development[10,13]. Therefore, to further explore the regulatory network of the BoAP1 protein, it was hypothesized that BoAP1 interacts with TRXz, FSD2, and FSD3 in Brassica plants and that different concentrations and types of ROS can also restore the green phenotype of 6-219W or boap1 mutants.
-
In conclusion, a spontaneous chloroplast-deficient mutant, 6-219W was identified. A single recessive gene, BoAP1, is responsible for this lethal albino trait. Fine mapping indicated that BolC08g019310.2J was identified as the candidate gene. Through CRISPR/Cas9 technology, it was demonstrated that the BoAP1 mutation disrupted the normal development of chloroplasts in 6-219W, leading to a lethal albino phenotype. The present research provides a basis for the in-depth study of the molecular mechanism regulating chloroplast development.
-
The authors confirm contribution to the paper as follows: study conception and design: Lv H; performing experiments: Zhao X, Yuan K; writing and revising manuscript: Zhao X, Chen L, Fujimoto R; data analyses: Liu Y; assisting experiments: Yang L, Zhuang M, Zhang Y, Wang Y, Ji J. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This work was supported by grants from the National Key R&D Program of China (2023YFD1201501), the Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-IVFCAAS), and China Agriculture Research System of MOF and MARA (CARS-23).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xinyu Zhao, Li Chen, Kaiwen Yuan
- Supplemental Table S1 Primer sequences used in this study.
- Supplemental Table S2 Predicted genes associated with chloroplast development in the candidate region.
- Supplemental Fig. S1 Protein sequence alignment of BoAP1 and its relatives. AT4G8590, MRL7/ECB1/SVR4/AtRCB of Arabidopsis thaliana; Boap1, mutant-type BoAP1 in 6-219W. The 298th amino acid is indicated by the red asterisk.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhao X, Chen L, Yuan K, Liu Y, Yang L, et al. 2024. Genetic mapping and gene editing reveal BoAP1 as a crucial factor regulating chloroplast development in Brassica oleracea. Vegetable Research 4: e031 doi: 10.48130/vegres-0024-0030 

Genetic mapping and gene editing reveal BoAP1 as a crucial factor regulating chloroplast development in Brassica oleracea
- Received: 07 May 2024
- Revised: 16 July 2024
- Accepted: 25 July 2024
- Published online: 08 October 2024
Abstract: Chloroplasts are crucial cellular components that plants need to carry out photosynthesis. Exploring the relevant underlying molecular regulatory mechanism could help elucidate this complex process. In this study, a spontaneous chloroplast-deficient mutant, 6-219W, exhibiting a lethal albino phenotype in curly kale (Brassica oleracea var. acephala) was identified. The number of chloroplasts in the 6-219W mutant were considerably reduced, those chloroplasts appeared crumpled, and the thylakoid membranes could not be observed under transmission electron microscopy. Genetic analysis revealed that boap1 (Brassica oleracea albino plant1), a single recessive gene, is responsible for this lethal albino trait. Fine mapping demonstrated that boap1 is located at a 300 kb interval between the InDel markers PW404 and PW406 on chromosome 8. Based on the B. oleracea genome annotation, a candidate gene, BolC08g019310.2J, was identified within the target interval. Sequence analysis revealed a 3-nucleotide (GAT) deletion in the coding sequence of BolC08g019310.2J in the 6-219W mutant, leading to the absence of the amino acid methionine at position 298 that is conserved in Arabidopsis thaliana, B. rapa, and B. oleracea. CRISPR/Cas9 technology was used to knock out the BolC08g019310.2J gene in NB11, and the generation of boap1 mutants with an albino phenotype confirmed BolC08g019310.2J as the causal gene. Subcellular localization indicated that the BoAP1 protein operates in chloroplasts. These results revealed that the BoAP1 mutation disrupted the normal development of chloroplasts in 6-219W, leading to a lethal albino phenotype. The present research lays a foundation for the in-depth study of the molecular mechanism regulating chloroplast development.
-
Key words:
- Chloroplast development /
- Albino lethal mutant /
- Map-based cloning /
- CRISPR/Cas9