










-
Theobroma cacao L. (cocoa), the source of cocoa beans, an important ingredient in chocolate, is grown worldwide where a favorable tropical environment prevails. Cocoa cultivation is heavily promoted due to the growing demand for cocoa beans and other by-products in the cosmetic, pharmaceutical, and chocolate sectors. It is native to the forested basin of the Amazon River and is a shade-loving tree. About 70% of the world’s cocoa beans come from four West African countries: Ivory Coast, Ghana, Nigeria, and Cameroon. It has been estimated that 40% of the worldwide production of cocoa is lost annually due to five major diseases[1]: black pod (due to Phytophthora spp.), witches’ broom (due to Moniliopthora perniciosa), cocoa swollen shoot virus, vascular streak dieback (due to Oncobasidium theobromae) and frosty pod/moniliasis (due to Moniliophthora roreri). In India, cocoa is cultivated in an area of 97,563 ha with an annual production of 27,072.15 MT and productivity of 669 kg/ha[2]. In India, cocoa (Theobroma cacao L.) is primarily grown as an intercrop within existing coconut (Cocos nucifera L.) and arecanut (Areca catechu L.) plantations in the southern states of Kerala, Karnataka, Tamil Nadu, and Andhra Pradesh. The shaded environment within these mixed cropping systems, combined with the favorable climate during the southwest monsoon season (June−October), creates ideal conditions for the development and spread of black pod disease. A random survey conducted on cocoa gardens across the four states of India during 2009 and 2010 found that black pod disease occurred in over 80% of the surveyed gardens[3]. This indicates a significant prevalence of the disease. Furthermore, it has been observed that both the incidence and severity of the disease are on the rise, corresponding to the expansion of cocoa cultivation areas. This upward trend in disease occurrence poses a serious economic threat, resulting in substantial financial losses for cocoa farmers in India[4]. Black pod disease caused by various species of Phytophthora, is the most serious pathogen in cocoa with losses estimated at 700,000 metric tons[5]. Black pod disease is severe, especially during the colder months when the average minimum temperature is less than 20 °C and humidity is greater than 85%[6]. During peak infestations, at least six fungicide applications are needed to control black pod disease. This relatively high frequency of spraying, coupled with increasing input costs (labor and fungicides) and a lack of knowledge about effective spraying techniques, has led to the adoption of chemical control being very low. An ecofriendly approach for black pod control is the use of resistant genotypes enriched with genes for morphological and agronomic traits of interest[7,8]. Germplasms are a rich source of resistant genes. The classification of genotypes based on their reaction to disease resistance is very important before using it for future cocoa breeding. The level of resistance might be assessed by in vitro screening on leaves and detached pods[9,10]. It has been reported that anatomical characteristics, morphophysiological characteristics of pods, and several biochemical characteristics might confer resistance to black pod disease in cocoa in addition to the disease-resistant genes[11]. In previous studies, international clones and hybrids were characterized genetically and screened for black pod resistance[6,12,13]. Genetic resistance to infection by three species of Phytophthora was assessed in 262 genotypes derived from F1 cacao segregating progeny (TSH 1188 × CCN 51) through the leaf inoculation method[14]. The genetic diversity associated with 30 cocoa accessions resistant to Phytophthora was carried out at the Cocoa Research Centre, Vellanikkara (India)[15]. Transcriptomic analysis of the susceptible clone NA 32 and the disease-resistant clone SCA6 was carried out to characterize basal differences in gene expression, early responses to Phytophthora infection, and polymorphisms in defense genes[16]. The importance of various factors contributing to disease resistance is necessary to develop stable resistance against black pod disease genotypes. The current study aims to evaluate the international clones for black pod resistance based on phenotypical and biochemical characteristics. The understanding of phenes associated with disease resistance could make selection and breeding programs more effective and easier.
-
The experimental material consisted of a diverse array of 24 cocoa international clones that showed resistance to Phytophthora in other countries (Table 1 & Supplementary Table S1). They were introduced from the International Cocoa Germplasm Quarantine House, University of Reading, UK and were maintained at the Cocoa Research Centre, Kerala Agricultural University, Vellanikkara, India as budded plants in the field (Agroecological zone – Central midland, lat − 10.546401, long − 76.277764).
Table 1. List of genotypes and their origin based on information in the International Cacao Germplasm Database (ICGD,
www.icgd.reading.ac.uk ).Sl. no. Genotype Origin 1 AMAZ 12 Ecuador 2 VB 514 Brazil 3 AMAZ 5/25 Ecuador 4 NA 804 Peru 5 GU 249/H Guiana 6 NA 33 Peru 7 NA 702 Peru 8 GU 183/G Guiana 9 GU 269/V Guiana 10 ICS 100 Trinidad and Tobago 11 LAF 1 Costa Rica 12 CHUNDALE Wayanad 13 PNG 290 Papua New Guinea 14 PNG 299 Papua New Guinea 15 1MC 105 Peru 16 PA 70 Peru 17 GU 125C Guiana 18 GU 226/V Guiana 19 WA 40 Malaysia 20 SPA 9 Colombia 21 TSA 792 Trinidad and Tobago 22 GU 195/V Guiana 23 SNK 413 Cameroon 24 LCTEEN 162-1010 Ecuador Screening test for Phytophthora resistance
Pathogen isolation
-
Phytophthora palmivora used in this investigation was isolated from the infected cocoa pods on carrot agar medium during the monsoon season which spans from June to August. For further experimental purposes, the isolates were maintained on potato dextrose agar plates after being purified using the hyphal tip culture method.
Artificial inoculation of pathogen by the detached pod test (DPT)
-
The detached pod inoculation method was used to evaluate the resistance of cocoa accessions[9]. Fresh, healthy, immature pods of the same size and age (four months old) were selected from 24 genotypes. Pods from highly susceptible genotype (CCRP 8) to black pod served as the control. The pod length and width were measured. The pods were thoroughly cleaned with soap and water before being disinfected with 70% ethanol. The inoculation of the pod with the pathogen was done through the non-pricking method. Using this technique, a disc of Phytophthora culture was applied onto the surface of the pod that had been moistened with cotton with sterile water. The inoculated pods were incubated in polythene bags with a cotton pad dampened with sterile water to create humidity. Every cocoa genotype was maintained in three replications. The lesion's length and width were measured alternatively for 10 d (Fig. 1). The following formula[17] was used to determine the percentage of infection.
$ \begin{split}&\rm Percent\;pod\;area\;infection=\\&\rm\dfrac{Average\;length\;of\;lesion\;\times Average\;width\;of\;lesion}{Length \times width\;of\;pod}\times 100 \end{split}$ 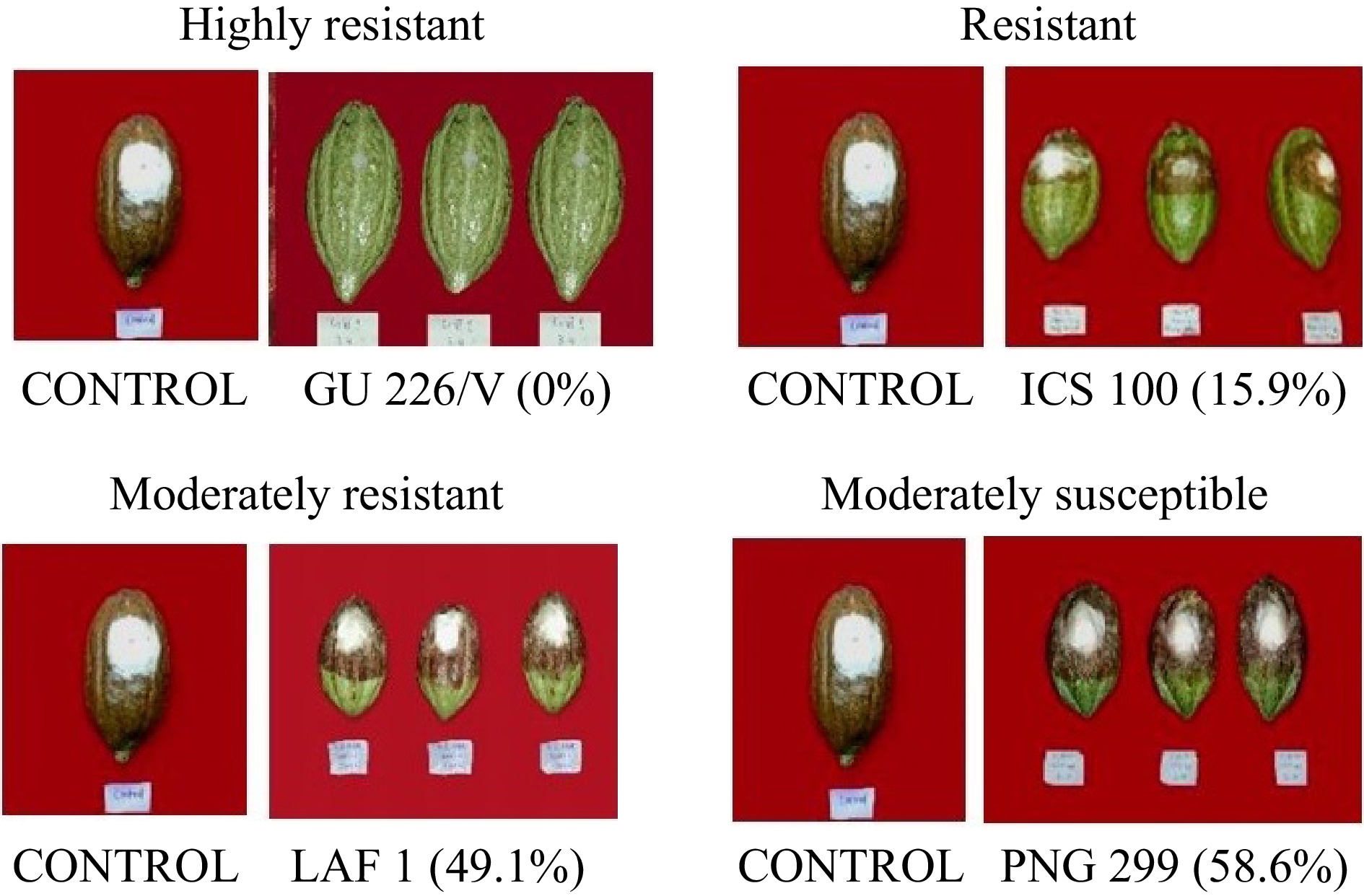
Figure 1.
Different categories of disease incidence under the non-pricking method of pod inoculation based on score chart.
Grouping of the genotypes was done as per the class as mentioned in Table 2[18].
Table 2. Score chart based on pod area infection.
Sl. no. Score Pod area infection 1 Highly resistant 0−15% 2 Resistant 15.1%−25% 3 Moderately resistant 25.1%−50% 4 Moderately susceptible 50.1%−75% 5 Susceptible > 75% Assessment of morphological, yield, and biochemical parameters
-
The following morphological and yield characteristics of ripened pods viz. bean color, pod weight, pod shape, ridge thickness, number of beans per pod, single dry bean weight, and yield on exotic genotypes of cocoa were noted. The ridge thickness was determined using a vernier caliper. The proposed descriptor[19] was used to record the observations. Standard procedures were adhered to in the analysis of biochemical attributes, such as the polyphenol content, fat, wax, calcium, potassium, and sodium content in three replications.
Estimation of polyphenol content
-
Polyphenol content was estimated through the folin-ciocalteu (FC) reagent method[20]. To estimate pod husk polyphenol, dewaxed cocoa pod husk was taken as the sample. Cocoa bean powder/ pod husk powder (500 mg) was defatted with 80% ethanol, centrifuged, and the supernatant collected. After evaporation to remove excess ethanol, the residue was dissolved in distilled water (40 mL). A 0.2 mL aliquot was mixed with water and FC reagent, followed by the addition of Na2CO3 solution. After incubation, absorbance was measured at 650 nm.
Estimation of fat
-
Fat was extracted with the aid of soxhlet apparatus[21]. Cocoa bean powder (10 g) was defatted with petroleum ether. The powder was wrapped in blotting paper, placed in the extraction tube, and solvent was added. After approximately 6 h, the fat settled at the bottom of the flask. The collected fat was transferred to a pre-weighed beaker, evaporated to remove solvent, and weighed to determine the percentage.
Estimation of wax
-
Wax content was estimated by the colorimetric method[22]. Three medium matured fresh pods were collected from each genotype and immersed in 15 ml chloroform for 15 s. The extract evaporated in a boiling water bath until the smell of chloroform could not be detected. After adding 5 ml of wax reagent, samples were placed in boiling water for 30 min. After cooling, 12 ml of deionized water was added. Several minutes were allowed for color development and cooling, extract was filtered using filter paper and then the optical density of the sample was read at 590 nm.
Estimation of minerals
-
Minerals like calcium, potassium, and sodium were assessed through a flame photometer[23]. Defatted cocoa powder (500 mg) was mixed with concentrated nitric acid for overnight pre-digestion. After digestion, 2 mL of 10% perchloric acid was added until the solution turned colorless. Distilled water (100 mL) was then added after cooling the mixture. From the flame photometer concentration in ppm, the mineral content was calculated.
Statistical data analysis
-
Correlation analysis between black pod infection and morphological/biochemical characteristics such as pod shape, ridge thickness, pod rugosity, polyphenol content, husk polyphenol content, fat content, wax, calcium, potassium, and sodium content were carried out and highly correlated characteristics were used for regression analysis. A binomial logistic regression model was employed to determine phenes contributing to black pod infection. It is a univariate/multivariate technique applied to estimate the probability that a characteristics is present by predicting a binary-dependent outcome from a set of explanatory variables and it is used for modeling binary response data. When a response is binary, it can have two values: 0 or 1, which, depending on the kind of study, can indicate resistant or susceptible. The dependent variable in this model is categorical.
A logistic model is used to predict the effect of change in the independent variable on the probability of belonging to a group when the dependent variable is dichotomous[24]. The correlation and regression analysis were done using the software IBM SPSS statistics 20.
Identification of genetic stock for black pod resistance breeding
-
The resistance was ranked along with morphological/biochemical traits that contributed to resistance based on the level of resistance of the 24 cocoa genotypes to black pod. The genotypes with more than two Kg of dry bean weight per tree per year was considered as high yielders, one to two Kg as moderate yielders, and less than one Kg were considered as low yielders. Pod base constriction was scored as follows: 0 – absent, 1 – slight, 2 – intermediate, 3 – strong and rugosity was done as follows, 0 – absent, 3 – slight, 5 – intermediate and 7 – intense[19]. The biochemical parameters such as fat, calcium, polyphenol content, and wax, as well as pod basal constriction and rugosity were ranked from highest to lowest (the highest value in the data with rank 1).
-
The relative susceptibility of 24 cocoa international clones used in the present study to Phytophthora by non-pricking methods are presented in Table 3. In the non-pricking method, out of the 24 international clones tested, 11 were grouped as highly resistant, with maximum resistance observed in genotype GU226/V (0%), GU249/H (0.2%), and NA 702(0.9%). The genotypes GU195/V (15.2%), ICS 100 (15.6%), SPA 9 (17.9%), LCTEEN 162-1010 (15.6%), and GU125C (24.26%) were classified as resistant. Six genotypes were under the class moderately resistant with an infection range from 25.1 to 50%. Accessions CHUNDALE (62.7%) and PNG299 (58.6%) were under the class moderately susceptible (Fig. 1).
Table 3. Classification of exotic genotypes based on percent pod area infection by the non-pricking method.
Sl. no. Genotypes Pod parameters (cm) Lesion size (cm) Classification based on percent pod area infection L W L W HR R MR MS S 1 AMAZ 12 20.5 26.9 6.3 10.2 13.04% 2 VB 514 23.2 24.2 7.1 7.2 8.83% 3 AMAZ 5/25 23.6 30.1 6.6 8.3 7.70% 4 NA 804 21.5 24.9 6.1 7.3 8.40% 5 GU 249/H 16.6 24.5 1.1 1 0.20% 6 NA 33 17.4 23 5.7 5.3 7.40% 7 NA 702 22.5 28.9 2.6 2.3 0.90% 8 GU 183/G 19.7 25.5 4.6 6.6 6.10% 9 GU 269/V 22.4 20.1 4.4 4.1 4.00% 10 ICS 100 18.7 26.1 8.2 9.6 15.90% 11 LAF 1 17.3 24.5 12.3 17 49.10% 12 CHUNDALE 19.3 26 15.5 20.3 62.70% 13 PNG 290 21.5 26.8 13.8 12.2 29.09% 14 PNG 299 17.7 27.2 13.9 20.2 58.60% 15 1MC 105 18.3 23.6 13.3 15.5 47.84% 16 PA 70 21.7 27.3 7.9 11.1 14.80% 17 GU 125C 18.3 24.6 11.5 9.6 24.26% 18 GU 226/V 20.2 23.9 0 0 0.00 19 WA 40 15.4 23.8 11.3 15.7 49.10% 20 SPA 9 19.3 23.2 8.2 9.7 17.90% 21 TSA 792 21.3 25.4 13.1 13.3 36.30% 22 GU 195/V 17.3 26.5 10 7.4 15.20% 23 SNK 413 16.6 23.2 11.4 9.1 27.20% 24 LCTEEN 162-1010 20.1 28.2 8.6 10.4 15.60% Highly resistant (HR), Resistant(R), Moderately resistant (MR), Moderately susceptible (MS), Susceptible (S), Length (L), Width (W). Analysis of morphological, yield, and biochemical characterization of cocoa genotypes
-
The consolidated data on the classification of 24 genotypes based on morphological, yield, and biochemical characteristics are presented in Table 4. Out of 24 genotypes, 13 displayed Angoleta-shaped fruits, four had Criollo-shaped fruits, four had Cundeamore-shaped fruits, two had Amelonado-shaped fruits, and one belonged to Calabacillo. The results of the present study revealed that out of the 24 genotypes, 14 genotypes (58%) expressed dark purple color for beans, and six genotypes (25%) with light purple color, three (13%) genotypes with mixed color for cotyledons, and one (4%) with medium purple. The pod weight and ridge thickness varied from 254 to 630 g and 0.76 to 1.65 cm respectively, across genotypes. The dry weight of beans ranged from 0.7 to 1.5 g respectively. Based on yield statistics, genotype LCTEEN 162–1010 recorded the highest yield with 2.5 kg, while VB 514 recorded the lowest yield at 0.2 kg. Biochemical parameters like total polyphenol (0.66%–13.92%), husk polyphenol (1.05%–4.58%), fat (39.66%–55%), wax (0.95–2.8mg), and calcium (203.25–324.24 mg/100g) were recorded.
Table 4. Characterization of exotic genotypes.
Genotypes Resistance category Characteristics AMAZ 12 Highly resistant PS − Criollo, BC − Dark purple, PW − 313 g, RT − 1.13 cm, BpP − 17, SDBW − 1.0 g, Yd − 0.3 Kg,
PP − 10.32%, HPP − 4.58, Fat − 52.24%, Ca − 320.15 mg/100 g, W − 2.48 mgVB 514 Highly resistant PS − Cundeamore, BC − Mixed, PW − 260 g, RT − 1.28cm, BpP − 19, SDBW − 1.0 g, Yd − 0.2 Kg,
PP − 11.75%, HPP − 4.5, Fat 51.16%, Ca − 310.84 mg/100 g, W − 2.36 mgAMAZ 5/25 Highly resistant PS − Amelonado, BC − Dark purple, PW − 312 g, RT − 1.12, BpP − 32, SDBW − 0.9 g, Yd − 0.5 Kg,
PP − 10.79%, HPP − 4.27%, Fat − 53%, Ca − 311.65 mg/100 g, W − 2.49 mgNA 804 Highly resistant PS − Cundeamore, BC − Dark purple, PW − 290 g, RT − 0.94 cm, BpP − 45, SDBW − 0.9 g, Yd − 0.6 Kg, PP − 11.52%, HPP − 3.64%, Fat − 55%, Ca − 308.15 mg/100 g, W − 2.52 mg GU 249/H Highly resistant PS − Angoleta, BC − Light purple, PW − 466g, RT − 1.48 cm, BpP − 28.9, SDBW − 1.1 g, Yd − 1.1 Kg,
PP − 9.49 %, HPP − 3.57%, Fat − 49.66%, Ca − 312.58 mg/100 g, W − 2.32 mgNA 33 Highly resistant PS − Angoleta, BC − Dark purple, PW − 515g, RT − 1.4cm, BpP − 50, SDBW − 1.2g, Yd − 2.4 Kg,
PP − 10.64%, HPP − 3.48, Fat − 48%, Ca − 298.75 mg/100 g, W − 2.8 mgNA 702 Highly resistant PS − Criollo, BC − Dark purple, PW − 440 g, RT − 1.04 cm, BpP − 54.2, SDBW − 1 g, Yd − 2.4 Kg,
PP − 10.96%, HPP – 3.25, Fat − 45%, Ca − 284.69 mg/100 g, W − 2.41 mgGU 183/G Highly resistant PS − Angoleta, BC − Dark purple, PW − 475 g, RT − 1.6 cm, BpP − 35, SDBW − 0.9 g, Yd − 1.2 Kg,
PP − 11.82%, HPP – 2.54%, Fat − 47%, Ca − 283.42 mg/100 g, W − 2.54 mgGU 269/V Highly resistant PS − Angoleta, BC − Dark purple, PW − 460 g, RT − 1.54 cm, BpP − 31.8, SDBW − 1 g, Yd − 1.3 Kg,
PP − 13.92%, HPP − 3.58%, Fat − 49%, Ca − 292.79 mg/100 g, W − 1.32 mgICS 100 Resistant PS − Angoleta, BC − Medium purple, PW − 506g, RT − 1.46 cm, BpP − 32.33, SDBW − 1.2 g, Yd − 1.5 Kg, PP − 6.89 %, HPP − 2.64%, Fat − 44.66%,Ca − 241.46 mg/100 g, W − 2.24 mg LAF 1 Moderately resistant PS − Criollo, BC − Dark purple, PW − 470 g, RT − 1.42 cm, BpP − 35.2, SDBW − 1.1 g, Yd − 2.4 Kg,
PP − 6.65%, HPP − 2.08%, Fat − 45.33%, Ca − 206.58 mg/100 g, W − 1.82 mgCHUNDALE Moderately susceptible PS − Criollo, BC − Light purple, PW − 544g, RT − 1.12 cm, BpP − 45.4, SDBW − 1.2 g, Yd − 2.1Kg,
PP − 7.70%, HPP − 1.17%, Fat − 53%, Ca − 207.56 mg/100 g, W − 1.35 mgPNG 290 Moderately resistant PS − Angoleta, BC − Dark purple, PW − 604 g, RT − 1.4 cm, BpP − 38, SDBW − 0.8 g, Yd − 1.3 Kg,
PP − 3.98%, HPP − 1.38%, Fat − 44.3%, Ca − 203.25 mg/100 g, W − 1.4mgPNG 299 Moderately susceptible PS − Angoleta, BC − Dark purple, PW − 630g, RT − 1.65 cm, BpP − 44, SDBW − 1.2 g, Yd − 1.5 Kg,
PP − 4.27%, HPP − 4.42%, Fat − 43 %, Ca − 226.87 mg/100g, W − 2.51mg1MC 105 Moderately resistant PS − Cundeamore, BC − Light purple, PW − 350 g, RT − 1.1 cm, BpP − 49, SDBW − 0.9 g, Yd − 1.4 Kg, PP − 3.19%, HPP − 2.34%, Fat − 39.66%, Ca − 215.56 mg/100 g, W − 1.65 mg PA 70 Highly resistant PS − Angoleta, BC − Light purple, PW − 454 g, RT − 1.48 cm, BpP − 48, SDBW − 1.1 g, Yd − 1.8 Kg,
PP − 11.27%, HPP − 4.55%, Fat − 40.6%, Ca − 292.31 mg/100 g, W − 2.48 mgGU 125C Resistant PS − Angoleta, BC − Dark purple, PW − 360 g, RT − 1.6 cm, BpP − 29, SDBW − 0.7 g, Yd − 0.9kg,
PP − 7.17%, HPP − 1.54, Fat − 52%, Ca − 279.46 mg/100 g, W − 1.5 mgGU 226/V Highly resistant PS − Calabacillo, BC − Light purple, PW − 550 g, RT − 1.6 cm, BpP − 42, SDBW − 0.9 g, Yd − 0.7 Kg,
PP − 12.02%, HPP − 1.05%, Fat − 53%, Ca − 324.24 mg/100 g, W − 2.6 mgWA 40 Moderately resistant PS − Cundeamore, BC − Light purple, PW − 540 g, RT − 1.64 cm, BpP − 38, SDBW − 0.9 g, Yd − 1.2 Kg, PP − 4.21 %, HPP − 2.11%, Fat − 41.66 %, Ca − 236.25 mg/100g, W − 0.95 mg SPA 9 Resistant PS − Angoleta, BC − Dark purple, PW − 272 g, RT − 0.76, BpP − 31.6, SDBW − 0.7 g, Yd − 0.4 Kg,
PP − 9.08%, HPP − 2.73%, Fat − 46.0%, Ca − 265.32 mg/100 g, W − 1.45 mgTSA 792 Moderately resistant PS − Angolata, BC − Mixed, PW − 360 g, RT − 1.1 cm, BpP − 53, SDBW − 0.9 g, Yd − 2.4 Kg,
PP − 5.18%, HPP − 1.24, Fat − 42.33%, Ca − 232.48 mg/100 g, W − 1.45 mgGU 195/V Resistant PS − Amelonado, BC − Dark purple, PW − 312 g, RT − 1.12 cm, BpP − 27.8, SDBW − 0.8 g, Yd − 0.5 Kg, PP − 9.35%, HPP − 3.61%, Fat − 45%, Ca − 259.16 mg/100, W − 2.4 mg SNK 413 Moderately resistant PS − Angolata, BC − Mixed, PW − 254 g, RT − 1.34 cm, BpP − 44, SDBW − 0.7 g, Yd − 0.2 Kg,
PP − 0.66%, HPP − 1.86%, Fat − 41.0%, Ca − 235.64 mg/100 g, W − 1.4 mgLCTEEN 162-1010 Resistant PS − Angolata, BC − Dark purple, PW − 508 g, RT − 1.28 cm, BpP − 35.4, SDBW − 1.5 g, Yd − 2.5 kg,
PP − 5.62%, HPP − 3.63%, Fat − 51%, Ca − 220.25 mg/100 g, W − 2.1 mgPod shape (PS), Bean color (BC), Pod weight (PW), Ridge thickness (RT), No. of beans/pod (BpP), Single dry bean weight (SDBW), Yield (Yd) (Total dry bean weight/pod/year), Polyphenol content (PP), Husk Polyphenol content (HPP), Fat content (Fat), Calcium content (Ca), Wax (W). Phenotypic traits associated with black pod resistance
-
Correlation studies done on the quantitative attributes indicated that calcium, bean and husk polyphenol, wax, and fat content responded negatively to the black pod infection. The calcium content (0.806) and polyphenol content (0.697) displayed strong negative correlations, whereas the wax content (0.530) and fat content (0.455) showed moderate negative correlations. A weak but negative correlation was exhibited in husk polyphenol (0.388) (Table 5). The result of the binomial regression model, specifically the high value of odds ratio Exp (B) and negative coefficient, indicated a negative relation between black pod infection and the fat and polyphenol content of cocoa beans, as well as husk polyphenol and pod wax.
Table 5. Logistic estimate of quantitative phenes influencing black pod infection.
Variables Coefficient Standard error Wald Significance Exp (B) Expected percentage of improvement overpopulation Fat** −0.455 0.441 1.067 0.302 1.577 Polyphenol** −0.697 0.812 1.464 0.226 0.374 Wax** −0.530 2.685 0.065 0.799 1.983 Husk polyphenol** −0.388 1.265 5.209 0.022 0.056 5.3 Calcium** −0.806 0.089 1.934 0.164 0.884 Constant 29.43 13.719 4.562 0.03 ** Significance value less than 0.03. Among these characteristics husk polyphenol exhibited a significant value less than 0.03 which is the constant. The results are depicted in Table 5. Based on the Exp (B) value from the regression model, when the expected percentage of improvement for disease resistance over the base population was calculated it was found that if the selection is based on the percentage of polyphenol present in the husk, a new population formed from the base population will express 5.3% improved disease resistance.
Correlation studies for qualitative characteristics demonstrated a strong positive correlation with rugosity (0.839) while moderate correlation was observed for pod base (0.518) with the black pod infection (Table 6). When a logistic estimate of qualitative phenes influencing disease resistance was carried out, the model was good with a significance value of 0.01 for only one characteristic i.e. pod rugosity (Table 6). The results of the current study showed a positive relation between infection percentage and pod rugosity. If selection is done within the existing population, based on the absence of basal constriction and a smooth pod surface devoid of rugosity, a marked reduction in infection rate of 84.42% is anticipated.
Table 6. Logistic estimate of qualitative phenes influencing black pod infection.
Variables Coefficient Standard error Wald Significance Exp (B) Expected percentage of improvement overpopulation Pod base** 0.518 0.838 0.244 0.62 0.661 Rugosity** 0.839 0.593 8.119 0.004 5.422 84.42 Constant −0.877 0.346 6.435 0.01 0.416 ** Significance value less than 0.01. Development of Phytophthora palmivora resistant genotype
-
The 24 genotypes were ranked through a comparative analysis of two categories. In the first category the classification was done based on the percentage of infection, distinguishing between highly resistant, resistant, moderately resistant, and moderately susceptible. The second category utilized qualitative and quantitative phenes identified by a binomial logistic regression model that were relevant in contributing to resistance. Factors contributing negatively to the percentage of infection like wax, husk polyphenol content, and factors positively contributing to disease resistance like pod rugosity and pod basal constriction were scored in descending and ascending order respectively (Table 7), which revealed, AMAZ 12, VB 514, AMAZ 5/25, NA 804, GU 249/H, NA 33, NA 702, GU 183/G, GU 269/V, PA 70, and GU 226/V emerging as highly resistant genotypes occupying the first 11 ranks attributing to the significant role of phenes in disease resistance.
Table 7. Genetic stock for black pod resistance breeding in cocoa.
Classes Genotypes Score
for fat contentScore for
polyphenol
contentScore for
wax content
in the huskScore for husk
polyphenol
contentScore for
calcium
contentScore for
pod basal
constrictionScore
for pod
rugosityTotal score Class of yield Rank Highly resistant AMAZ 12 3 10 9 1 2 0 0 25 Low yielder 2 VB 514 5 4 12 3 5 0 0 29 Low yielder 4 AMAZ 5/25 2 8 8 5 4 0 0 27 Low yielder 3 NA 804 1 5 6 6 6 0 0 24 Low yielder 1 GU249/H 7 11 13 9 3 0 0 43 Moderate yielder 8 NA 33 9 9 2 10 7 1 0 38 High yielder 6 NA 702 13 7 10 11 10 0 0 51 High yielder 11 GU183/G 10 3 5 14 11 0 1 44 Moderate yielder 9 GU 269/V 8 1 21 8 8 0 0 46 Moderate yielder 10 PA 70 20 6 3 2 9 0 0 40 Moderate yielder 7 GU226/V 2 2 4 24 1 0 0 33 Low yielder 5 Resistant ICS 100 14 16 14 13 15 0 0 72 Moderate yielder 16 GU125C 4 15 17 19 12 0 3 70 Low yielder 15 SPA 9 11 13 18 12 13 0 0 67 Low yielder 13 GU195/V 13 12 11 7 14 0 1 58 Low yielder 12 LCTEEN 16-1010 6 18 1 20 24 2 3 74 High yielder 17 Moderately resistant LAF 1 12 17 15 17 22 1 7 91 High yielder 19 PNG 290 15 22 19 21 23 0 0 100 Moderate yielder 21 IMC 105 21 23 16 15 20 3 3 101 Moderate yielder 22 WA40 18 21 22 16 16 1 7 101 Moderate yielder 22 TSA 792 17 19 18 22 18 0 5 99 Moderate yielder 20 SNK 413 19 24 19 18 17 0 3 100 Low yielder 21 Moderately susceptible CHUNDALE 2 14 20 23 21 2 7 89 High yielder 18 PNG 299 16 20 7 4 19 0 3 69 Moderate yielder 14 Further classification based on yield and resistance portrayed, higher yield potential coupled with higher disease resistance in NA 33, and NA 702. Conversely GU 269/V, GU 249/H, GU 183/G, and PA 70 were categorized under moderate yielders despite exhibiting higher disease resistance.
-
In the current investigation, black pod screening was done by artificially inoculating cocoa pods using the non-pricking approach. The non-pricking method is the most appropriate method to check the phenes influencing black pod resistance in cocoa, as the biochemical reaction occuring due to wounding can be avoided[25].
The importance of various factors contributing to disease resistance is necessary to develop stable resistance against black pod disease genotypes. Thus, the 24 genotypes of cocoa used in the present study were classified based on morphology, yield, and biochemical characteristics to understand the influence of these characteristics on disease resistance. Morphological characteristics are reported to influence disease resistance in crops[26]. The thickness of pod husks serve as a resistance factor to black pod in cocoa[11]. Thirty cocoa accessions were evaluated for Phytophthora resistance and genotypes ICS 41 and ICS 75 with the absence of pod base and acute pod base were reported to be highly resistant[13]. The present study revealed that pod base and pod rugosity had a positive relation with disease infection. It was reported that more resistance was observed in Cundeamore and Angoleta-shaped pods with smooth surface (absence of rugosity) with the absence of basal constriction and it was reported that resistance was mainly due to the smooth surface of Calabacillo and Cundeamore pods conferring less water retention over the surface of the pod[27,28]. In the present study, out of 11 highly resistant genotypes, six were expressing either Cundeamore, Angoleta or Calabacillo-shaped fruits with a smooth surface. Hence, the selection of genotypes having a smooth pod surface or the absence of rugosity can help to minimize black pod infection. Similarly, selecting genotypes having low or absence of pod base constriction can help in lowering the growth of Phytophthora on the pod and hence, minimizing the spread of infection. A regression model was used to estimate the conditional expectation of the dependent characteristics. Logistic regression indicated that morphological phenes mainly influence disease susceptibility or resistance[29]. Quantitative characteristic correlation studies revealed an overall negative correlation with black pod infection. Therefore, limiting the spread of infection might be achieved by selecting genotypes with higher values for characteristics like calcium content, polyphenol content, and wax content. The high value of odds ratio Exp (B) and positive coefficient indicated that ridge thickness, polyphenol content, and calcium content had a positive relation with black pod resistance[15]. The regression studies indicated that wax, polyphenol, and calcium content had a negative relation with black pod infection. Epicuticular wax on pod and leaf surfaces imparts a crucial role in the host plant's resistance to black pod in cocoa (Theobroma cacao L.) and reported that cocoa genotypes that contain more wax were highly impervious to black pod infection compared with those of low wax[30,31]. The high amount of wax on the surface of pod was observed in LCTEEN 60-1010. This study established that the epicuticular wax layer affords cocoa pods greater protection against black pod. Phenolic compounds present in plants possess a role in the plant defense against pathogens[32]. After artificial inoculation of pods by mycelium of Phytophthora megakarya, the content of phenolic compounds significantly increased in tolerant genotypes[33]. It was reported that ICS 41, which is highly resistant with high yield exhibited more polyphenol content[34]. The highest total phenol content was observed in genotype GU 269/V (13.92%) which showed high disease resistance too. This suggests that phenolics play a major role in conferring disease resistance in cocoa. An increase in the level of calcium will trigger the cellular response and result in the production of secondary metabolites, which are directly related to the defense mechanism in plants[35,36]. Calcium content was recorded to be high in GU226/V which showed high resistance to black pod. The study showed that genotype NA 702 had a high score for phenes contributing to resistance, indicating that some other signal transduction pathway may be the reason for the resistance[37]. NA[12], PA[12], and GU[38] groups of clones were found to be resistant and moderately resistant in the previous studies.
Genotypes exhibiting strong disease resistance and high yield potential might be crossed by considering their genetic divergence. This approach aims to generate a favorable hybrid without experiencing adverse heterosis, and it also facilitates gene pyramiding, given that black pod resistance is known to involve multiple genetic factors[39]. Genotypes displaying high resistance but with moderate or low yield might be crossbred with a documented high-yielding variety or another high-yielder from the same gene pool. This breeding strategy is employed to enhance overall yield potential.
-
Control of black pod disease caused by Phytophthora palmivora is a difficult task because of the availability of inoculum in abundant quantities. The heightened prevalence of Phytophthora disease during the rainy season makes the application of fungicides an ineffective control measure. Thus, the breeding of resistant cultivars is the most effective and environmentally sustainable approach to combating this disease. The aforementioned results highlight the non-pricking method's efficacy and imply that morphological characteristics other than internal resistance have an impact on resistance. Binomial logistic regression analysis revealed that black pod infection was inversely correlated with specific phenes, such as calcium content, polyphenol content, and wax. On the other hand, phenes that have a positive association with disease infection include pod rugosity and pod basal constriction. The population that results from selection using these phenes may have higher resistance levels. Accessions demonstrating high resistance can then be strategically employed in subsequent cocoa breeding programs. The observed association between phenotypic traits and black pod resistance needs to be further validated in a larger number of germplasm accessions with diverse genetic backgrounds. Upon completing the validation, these phenotypic traits could be used to complement direct screening of black pod resistance in cocoa breeding programs.
-
The authors confirm contribution to the paper as follows: study conception and design, interpretation of data: Minimol JS; critical review of manuscript: Suma B, Plappallil SK; experiment execution in the field and observation recording: Mary A; draft manuscript preparation, data analysis: Shija TK, Jose S; final approval of the manuscript to be published: Minimol JS. All authors reviewed the results and agreed the final version of the manuscript.
-
The data that support the findings of the study are available on request from the corresponding author.
The authors thank Mondelez India Pvt. Ltd and State Plan, Government of India for their support.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Anova for quantitative characters of cocoa.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Minimol JS, Suma B, Mary A, Shija TK, Jose S, et al. 2024. Evaluating black pod resistance in cocoa (Theobroma cacao L.) and its relationship with phenotypical and biochemical characteristics. Beverage Plant Research 4: e038 doi: 10.48130/bpr-0024-0028 

Evaluating black pod resistance in cocoa (Theobroma cacao L.) and its relationship with phenotypical and biochemical characteristics
- Received: 22 April 2024
- Revised: 10 June 2024
- Accepted: 08 July 2024
- Published online: 18 October 2024
Abstract: Disease associated with Phytophthora is a great menace to cocoa. Chemical control measures are however proven to be ineffective. Hence, the most feasible and effective method to combat this situation is using resistant planting material as they are the source of genetic variance. The level of resistance against black pod disease was assessed among 24 international clones of cocoa by the non pricking method of detached pod inoculation. In the non-pricking method, 11 genotypes were grouped under the highly resistant category. The binomial logistic regression model revealed that phenes like polyphenol content, wax, husk polyphenol, and calcium content had a positive influence on Phytophthora resistance. Upon completing further validation, these phenes might be considered for the selection of resistant genotypes from a population. From the present investigation, it was found accessions NA 33, NA 702, and PA 70 showed maximum resistance to disease with high yield potential which could be utilized for further genetic improvement programs in cocoa.
-
Key words:
- Black pod /
- Cocoa /
- Disease /
- Phytophthora /
- Resistance