









-
Grape (Vitis vinifera L.) is one of the most popular fruits in the world. Table grapes account for approximately 36% of global grape production[1]. In China, table grapes account for 80% of total grape production[2]. In light of this, it is very important to study the aroma of table grapes. The volatile compounds in grapes affect sensory evaluation, which could be the reason that consumers choose certain grapes over others[3,4]. Volatile compounds in fruits are responsible for defining their aroma and flavor. We can obtain grapes with distinct aromas and characteristics for the varying volatile combinations and concentrations[5]. Fruit volatile compounds are mainly comprised of esters, alcohols, aldehydes, ketones, lactones, terpenoids, and apocarotenoids[6]. Many factors affect volatile composition, including the genetic diversity[7,8], viticultural techniques[9,10], degree of maturity[4], climatic conditions[3,11,12], and postharvest storage conditions[4,13]. Among these factors, climate conditions (sunlight, temperature, water status, etc.) were often considered an important factor for grape volatile compounds for the same cultivar[11].
Light is the primary climatic factor affecting volatile composition. As we all know, intensity, quality, and photoperiod are the main factors of light regulation[14]. Sunlight promotes the accumulation of terpenoids and monoterpene, which are the typical aroma components in Muscat grapes[3,12]. Modified canopy management (basal leaf removal) and exposure to appropriate proportions of blue and red light were effective strategies to improve the characteristic aroma[14,15]. Temperature is another important climate factor affecting volatile composition. Generally, excessively high temperature is deemed to have negative effects on fruit metabolism[16]. For example, high temperature in the winter season inhibited most VviCCDs expression than in summer grape berries, which was associated with norisoprenoid accumulation[3]. Temperate zones are more conducive to the formation of aroma substances[17]. Compared with grapes grown under cool conditions, the same grape variety presented a higher concentration of monoterpenes when cultivated under warm conditions[18]. The other factor that influences grape development is water availability. Proper water deficit has been proven to be available for increasing the characteristic aroma contents, especially terpenes and esters[19,20]. Therefore, improving water use efficiency can increase fruit flavor.
Different terrains forms different aroma characteristics. The southern subtropical region of China was not a traditional viticultural area due to the sticky rainy weather and inadequate low-temperature accumulation[3,21]. With the application of grape two-crop-a-year cultivation technology, the above-mentioned problems have been conquered[22], and Guangxi (a province in southern China) has become a unique advantage viticulture area[23]. Due to the plentiful sunlight and temperature accumulation, grape berries could be harvested twice a year[3,22]. Summer grape fruits are the name of grapes harvested in the first growing season, while winter grape fruits are the name of grapes harvested in the second growing season[24]. 'Ruidu Kemei', breeding from a cross between 'Italy' and 'Muscat Louis', is a new table grape variety appropriate to two-crop-a-year cultivation[25,26]. At present, there are few reports about grape volatiles under two-crop-a-year cultivation systems. Recently, Lu et al. compared the volatile profiles of 'Riesling', 'Cabernet Sauvignon', 'Victoria', and 'Muscat Hamburg' grape berries under two-crop-a-year cultivation[3]. However, knowledge about the volatile profiles of two-crop grapes is still very rare. More work needs to be carried out to establish the aroma substance characteristics, and to provide a theoretical basis for improving aroma under two-crop-a-year cultivation systems.
To distinguish grape volatiles under the two-crop-a-year cultivation system, the volatiles in summer and winter berries of 'Ruidu Kemei' were qualitatively and quantitatively analyzed by headspace solid phase microextraction (HS-SPME) combined with gas chromatography-mass spectrometry (GC-MS). Meanwhile, two crops' volatiles were also conducted in relation to climate factors in the present study.
-
This experiment was conducted during two growing seasons in 2022 on 3-year-old 'Ruidu Kemei' grapevines in the vineyards of the Grape and Wine Research Institute, Guangxi Academy of Agricultural Sciences, located in Nanning, Guangxi Province, China (22°36'39" N, 108°13'51" E). In this vineyard, the vines were managed on a canopy frame with a single trunk and were planted in north-south-oriented rows spaced 1.5 m (between vines) × 2.5 m (between rows). Nutrition, pest, water, and fertilizer management was carried out by uniform standards for two-crop-a-year as previously described[3].
The key techniques of two-crop-a-year cultivation systems was described by Cheng et al.[24]. Summer grape fruits (SF) were harvested on July 15th, and winter grape fruits (WF) were harvested on December 31st.
Berry sampling and meteorological data collection
-
Six vines for sampling were chosen based on their relatively consistent growth status. Six biological replicates were conducted in this study, and each biological replicate comprised 90 berries from six clusters of different vines, then sampled berries of each biological replicate were mixed and put into a 50 mL centrifuge tube, and immediately frozen in liquid nitrogen, and stored at −80 °C until needed.
Temperature (°C), relative humidity (%), and solar radiation intensity (W/m2) were acquired according to Cheng et al.[24]. Growing degree days (base 10 °C) were calculated from bloom to harvest according to Bindi et al.[27].
Volatile metabolome methods
Sample preparation and treatment
-
Samples of each biological replicate were ground to powder in liquid nitrogen, and 500 mg powder was transferred immediately to a 20 mL head-space vial (Agilent, Palo Alto, CA, USA), containing NaCl-saturated solution, to inhibit any enzyme reaction. The vials were sealed using crimp-top caps with TFE-silicone headspace septa (Agilent). At the time of SPME analysis, each vial was placed at 60 °C for 5 min, then a 120 μm DVB/CWR/PDMS fiber (Agilent) was exposed to the headspace of the sample for 15 min at 60 °C.
GC-MS conditions
-
After sampling, desorption of the VOCs from the fiber coating was carried out in the injection port of the GC apparatus (Model 8890; Agilent) at 250 °C for 5 min in the splitless mode. The identification and quantification of VOCs was carried out using an Agilent Model 8890 GC and a 7000D mass spectrometer (Agilent), equipped with a 30 m × 0.25 mm × 0.25 μm DB-5MS (5% phenyl-polymethylsiloxane) capillary column. Helium was used as the carrier gas at a linear velocity of 1.2 mL/min. The injector temperature was kept at 250 °C and the detector at 280 °C. The oven temperature was programmed from 40 °C (3.5 min), increasing at 10 °C/min to 100 °C, at 7 °C/min to 180 °C, at 25 °C/min to 280 °C, hold for 5 min. Mass spectra was recorded in electron impact (EI) ionization mode at 70 eV. The quadrupole mass detector, ion source, and transfer line temperatures were set, respectively, at 150, 230, and 280 °C. The MS with selected ion monitoring (SIM) mode was used for the identification and quantification of analytes.
Statistical analysis
Principal component analysis (PCA)
-
Unsupervised PCA was performed by the statistics function prcomp within R (
www.r-project.org ). The data was unit variance scaled before unsupervised PCA.Hierarchical Cluster Analysis (HCA) and Pearson correlation coefficients
-
The HCA results of samples and metabolites were presented as heatmaps with dendrograms, while Pearson correlation coefficients (PCC) between samples were calculated by the cor function in R and presented as only heatmaps. Both HCA and PCC were carried out by the R package ComplexHeatmap. For HCA, normalized signal intensities of metabolites (unit variance scaling) are visualized as a color spectrum.
Differential metabolites selected
-
For two-group analysis, differential metabolites were determined by VIP (VIP > 1) and absolute Log2FC (|Log2FC| ≥ 1.0). VIP values were extracted from OPLS-DA results, which also contain score plots and permutation plots, and was generated using R package MetaboAnalystR. The data was log transform (log) and mean centering before OPLS-DA. To avoid overfitting, a permutation test (200 permutations) was performed.
Kyoto Encyclopedia of Genes and Genomes (KEGG) annotation and enrichment analysis
-
Identified metabolites were annotated using the KEGG Compound database (
www.kegg.jp/kegg/compound , accessed on April 2nd, 2022), annotated metabolites were then mapped to the KEGG Pathway database (www.kegg.jpkegg/pathway.html , accessed on April 2nd, 2022). Pathways with significantly regulated metabolites mapped then fed into MSEA (metabolite sets enrichment analysis), their significance was determined by hypergeometric test's p-values. -
Volatiles in grape berries were affected by meteorological parameters under the double cropping system[3]. Significant differences in meteorological parameters between the two crop growing seasons are shown in Table 1. The summer growing season was from 1 March to 15 July, and the winter growing season was from 1 September to 31 December. In the present study, the active accumulated temperatures for both growing seasons were greater than 3,100 °C (Table 1), meaning that the active accumulated temperatures were sufficient to guarantee normal grape maturity[28]. The active accumulated temperature, the effective accumulated temperature, and the daily average temperature for the summer growing season was higher than those of the winter growing season. However, there were 83.33 h of high temperatures over 35 °C during the summer growing season, which was less than the winter growing season (127.17 h). Moreover, the relative humidity during the summer growing season showed a higher value than the winter growing season. For the solar radiation intensity and cumulative solar radiation, the winter growing season was higher than the summer growing season.
Table 1. Phenology and climatic factors during the two crop-growing seasons in Nanning (China) in 2022.
Meteorological data Summer Winter Phenology 1 Mar−15 Jul 15 Aug−31 Dec Active T (°C) 3,393.53 3,149.98 Effective T (°C) 2,023.53 1,769.98 Average daily temperature (°C) 24.78 21.92 High temperature (> 35 °C) (°C) 83.33 127.17 Relative humidity (%) 86.94 80.78 Solar radiation Intensity (W/m2) 93.86 108.65 Cumulative solar radiation (W/m2) 3,703,264.3 3,805,233.3 Overview of the volatile compounds of summer fruit and winter fruit
-
To figure out the difference between SF and WF, volatile metabolite analysis was applied in this study. A total of 620 metabolites in 15 categories were detected, including 122 terpenoids, 115 esters, 99 heterocyclic compounds, 60 hydrocarbons, 52 ketones, 48 alcohols, 47 aldehydes, 31 aromatics, 11 amines, 11 acids, eight phenols, seven nitrogen compounds, three halogenated hydrocarbons, two sulfur compounds, and four others (Fig. 1a, Supplementary Table S1). There was no difference between SF and WF for 12 categories (Fig. 1b). WF had more terpenoids and heterocyclic compounds than SF. Conversely, SF had more esters (Fig. 1b).
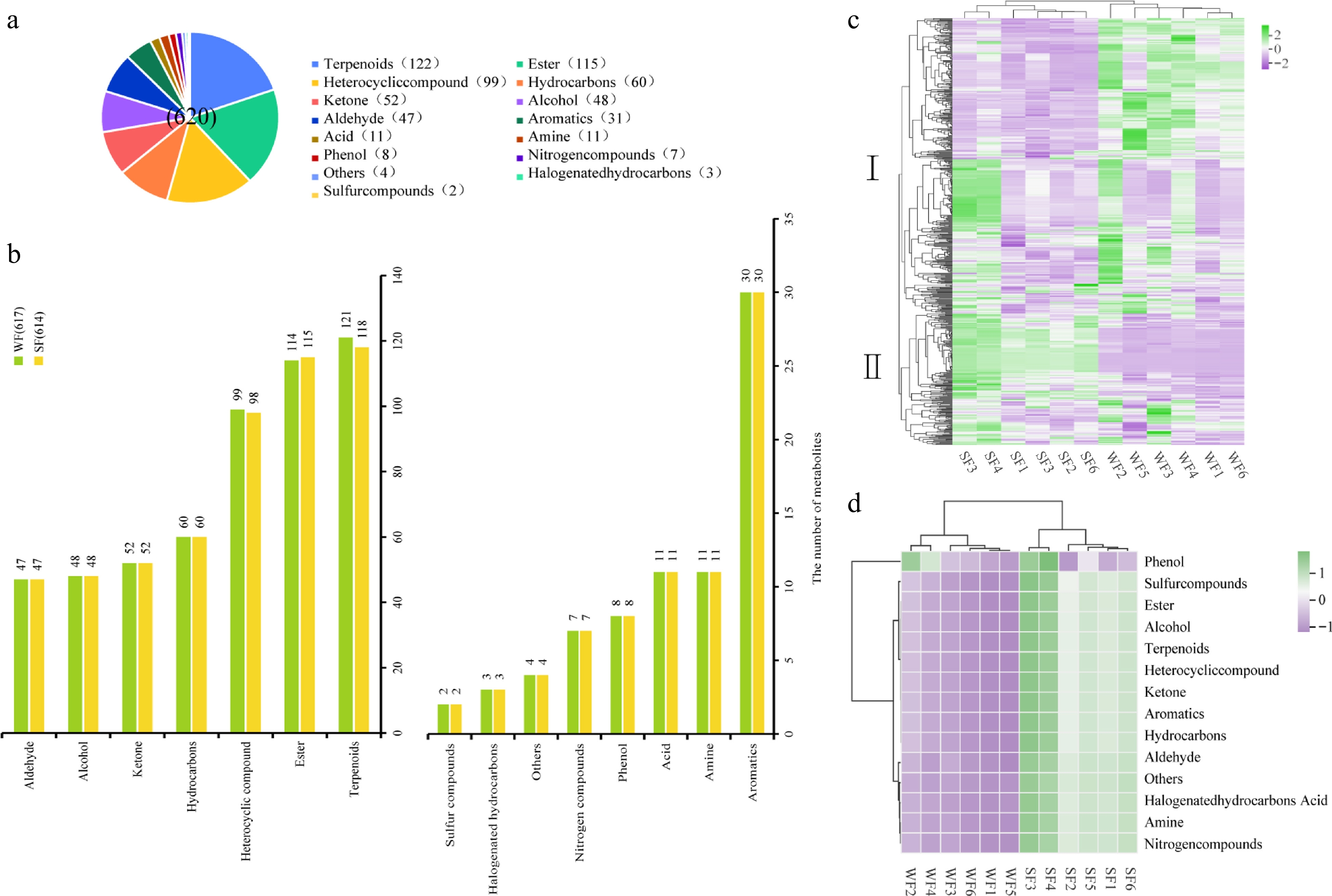
Figure 1.
(a) Categorical all metabolite statistics. (b) Categorical metabolite statistics for SF & WF. (c) All metabolites for hierarchical cluster analysis (HCA). (d) The relative content of classified metabolites for SF & WF.
For the relative metabolite contents, it was found that the metabolites were divided into two clusters, and significant differences could be observed in the substances between SF and WF. The metabolite relative contents in Cluster I were higher in SF, while WF exhibited higher relative contents in Cluster II metabolites (Fig. 1c). Phenols showed little difference between SF and WF, but SF was richer in the 14 other categories than that of WF (Fig. 1d).
SF and WF were evidently distinguished by PCA (Fig. 2a), the explanation rate of the first five principal components reached 87.1% (Fig. 2b). The cluster dendrogram divided SF and WF into two groups, which was consistent with PCA (Fig. 2c). The results indicated that the volatile compounds differed greatly between SF and WF.
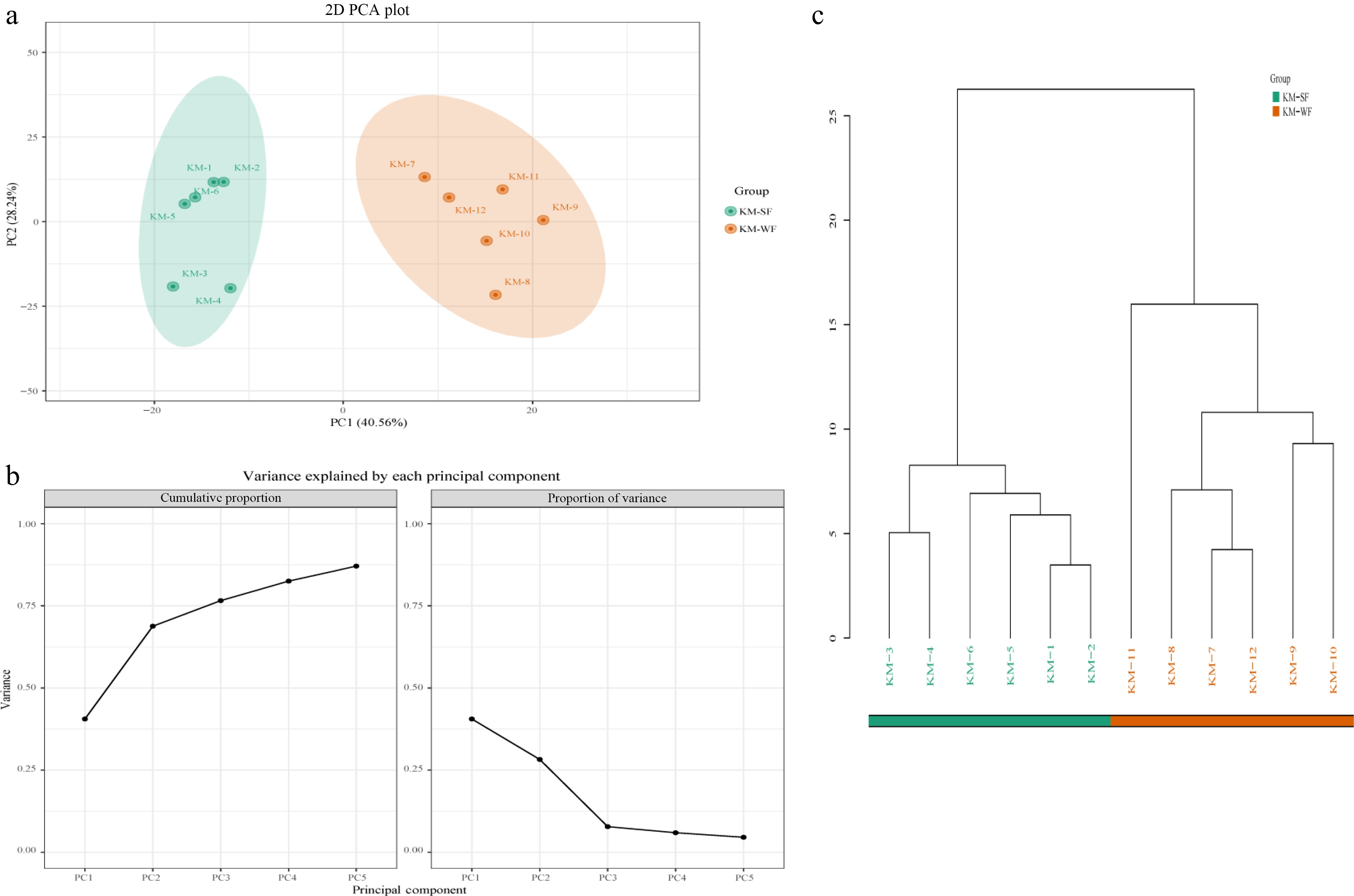
Figure 2.
(a) Principal component analysis (PCA). (b) Grouped principal component analysis explanation rate plot. (c) Sample hierarchical clustering tree.
Analysis of differential volatile compounds
-
To better distinguish volatile compounds between summer fruits and winter fruits, metabolites with fold change ≥ 2 and fold change ≤ 0.5 were selected as significant differences. The comparison SF_vs_WF showed a total of 143 different metabolites accounted for 23.18 % of the total detected substances, including 85 up-regulated metabolites and 58 down-regulated metabolites (Fig. 3a). The metabolites with a higher number for up-regulated were terpenoids, ketone, hydrocarbons, ester, aldehyde, alcohol, halogenated hydrocarbons, acids, and others (Fig. 3b). The metabolites with a higher number for down-regulated were amine, aromatics, nitrogen compounds, phenol, and heterocyclic compounds. It's worth noting that no sulfur compounds showed a statistically significant difference between SF and WF (Fig. 3a & b). For relative content, terpenoids, heterocyclic compounds, esters, and aromatics showed greater difference than other compounds (Fig. 3c).
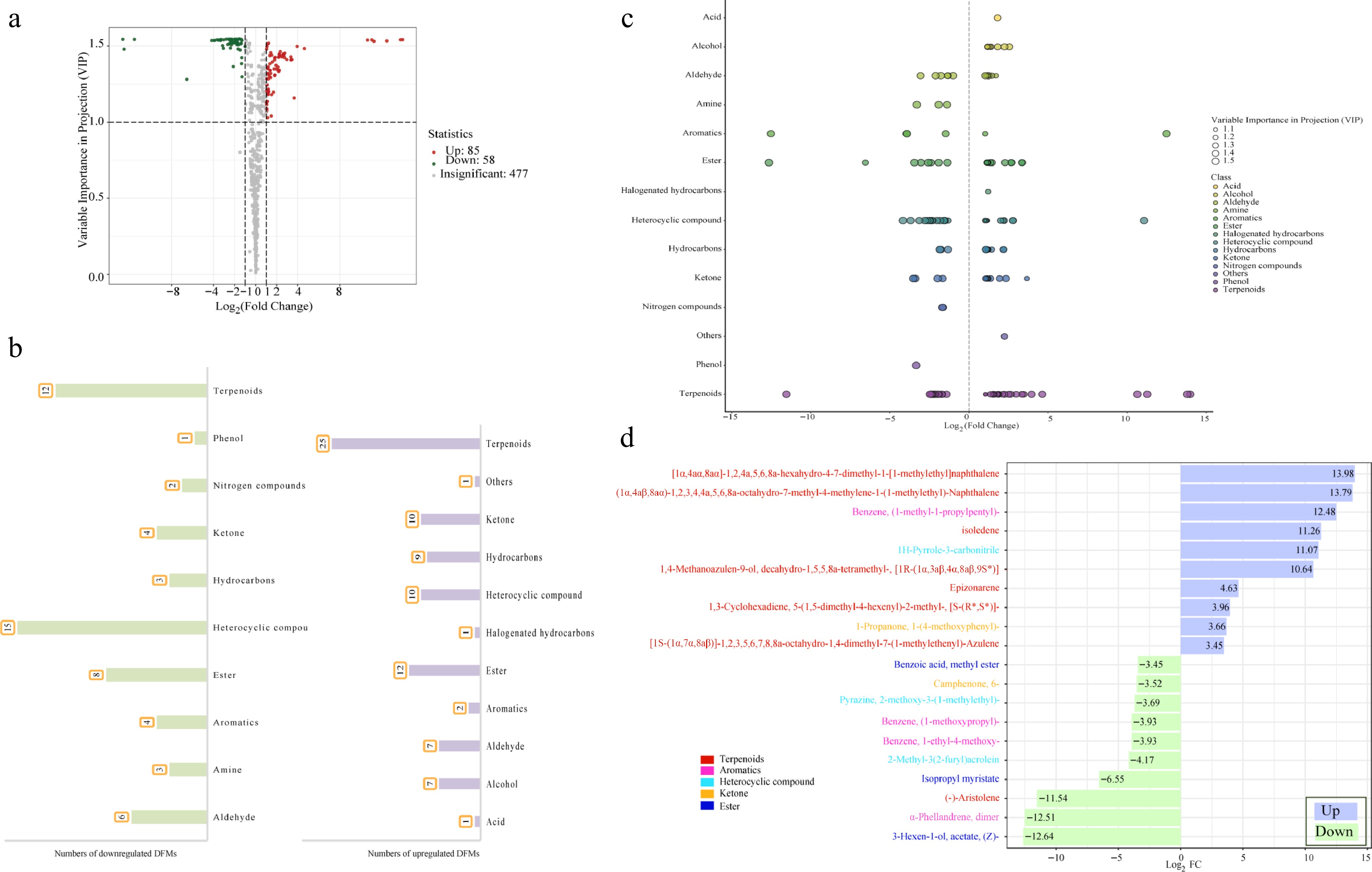
Figure 3.
(a) Volcanic plot of differential volatile compounds. (b) Bar chart of the number of volatile compounds classified for up-regulation & down-regulation. (c) Scatter plot of differential volatile compounds. (d) Bar chart of the top 20 differential volatile compounds.
To determine the metabolites with large differences for the SF_vs_WF comparison, a list of the top 20 substances using Log2FC was made, including 10 up-regulation substances and 10 down-regulation substances (Fig. 3d). There were obvious distinctions between the SF and WF. The top 20 substances using Log2FC contained five categories: terpenoids (8), aromatics (4), heterocyclic compounds (3), esters (3), and ketones (2). The top 10 up-regulation substances contained seven terpenoids, one aromatic, one heterocyclic compound, one ketone, while three esters, three aromatics, two heterocyclic compounds, and one ketone in the top 10 down-regulation substances. These results suggested that, for the top 20 substances using Log2FC, terpenoids were mainly up-regulated in WF, while esters and aromatics were up-regulated in SF. Specifically, the up-regulated and down-regulated substances with the largest Log2FC for SF_vs_WF comparison were [1α,4aα,8aα]-1,2,4a,5,6,8a-hexahydro-4-7-dimethyl-1-[1-methylethyl]naphthalene (terpenoid) and 3-Hexen-1-ol, acetate, (Z)-(ester).
KEGG enrichment analysis of differential volatile compounds
-
Fourty-nine of the 620 metabolites were annotated to 20 KEGG pathways (Supplementary Table S2). Additionally, 13 differential volatile compounds out of 143 differential volatile compounds between SF and WF were primarily annotated and enriched in the following seven pathways: biosynthesis of secondary metabolites, metabolic pathways and sesquiterpenoid and triterpenoid biosynthesis, monoterpenoid biosynthesis, limonene and pinene degradation, terpenoid backbone biosynthesis, and α-Linolenic acid metabolism (Fig. 4a & b; Table 2). Among them, the top three KEGG pathway types were biosynthesis of secondary metabolites, metabolic pathways and sesquiterpenoid, and triterpenoid biosynthesis, accounting for 53.85%, 46.15%, and 38.46% of the total differential volatile compounds annotated in KEGG respectively (Fig. 4a). KEGG annotations and enrichment showed that sesquiterpenoid and triterpenoid biosynthesis, monoterpenoid biosynthesis, limonene and pinene degradation were the main KEGG pathways for the differential volatile compounds between SF and WF (Fig. 4b). Significantly except sesquiterpenoid and triterpenoid biosynthesis, the other six pathways were mainly down-regulated (Table 2). These 13 differential volatile compounds were nine terpenoids, three aldehydes, and one ester (Table 2). Only four terpenoids were more in WF when compared with SF, including (E)-β-Famesene, Naphthalene,1,2,3,5,6,8a-hexahydro-4,7-dimethyl-1-(1-methylethyl)-, (1S-cis)-, α-Farnesene, and (E)-1-Methyl-4-(6-methylhept-5-en-2-ylidene)cyclohex-1-ener (Table 2). All the remaining nine differential volatile compounds were less in WF than in SF (Table 2).
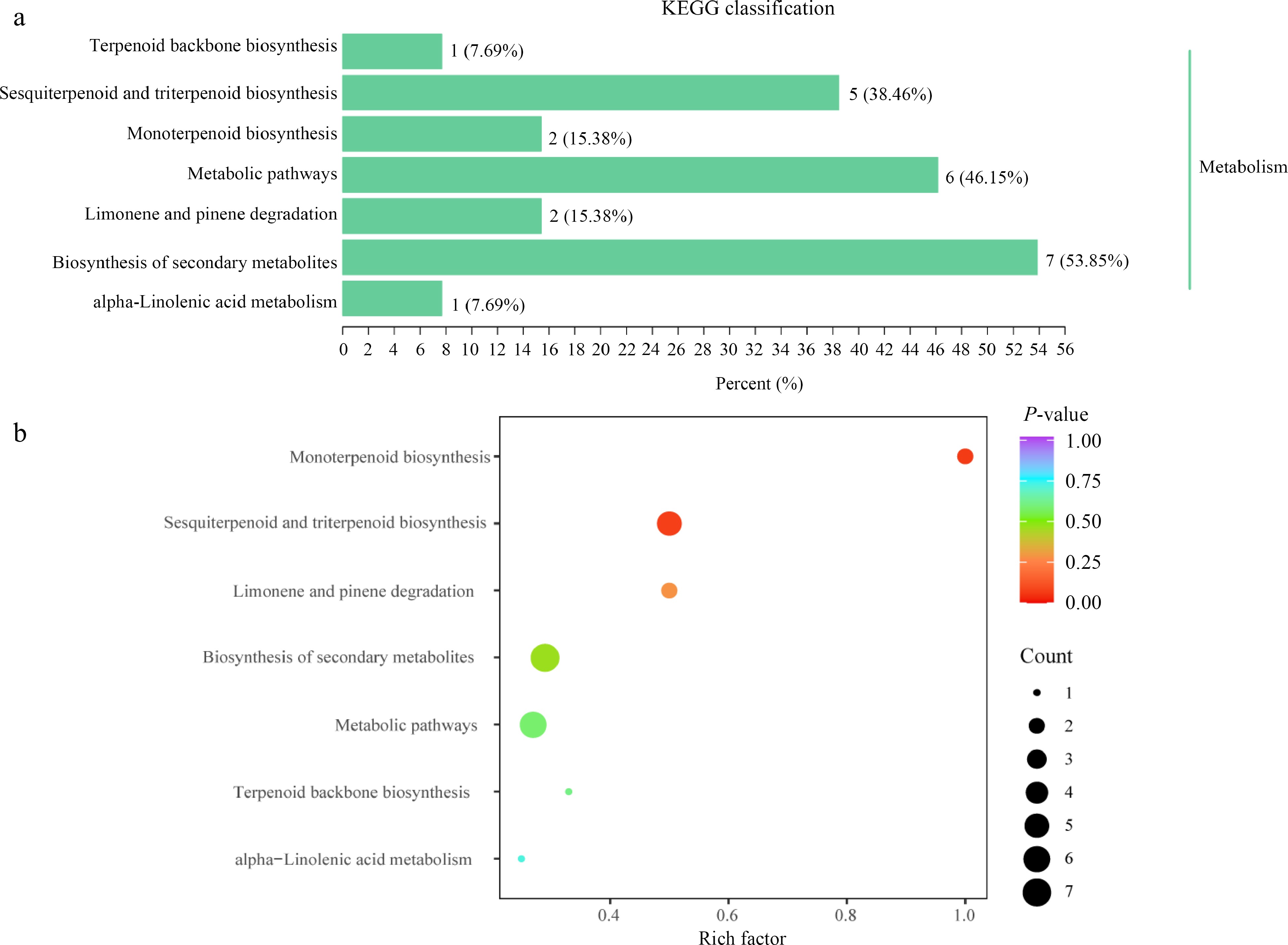
Figure 4.
The classification of the KEGG enrichment pathway. (a) KEGG enrichment analysis of differential volatile compounds. (b) KEGG annotations and enrichment of differential volatile compounds for SF_vs_WF comparison.
Table 2. KEGG functional annotation and enrichment of differential volatile compounds between SF and WF.
Formula Compounds KEGG_pathway Class Odor SF vs WF C10H18O L-α-Terpineol Metabolic pathways, Biosynthesis of secondary metabolites, Monoterpenoid biosynthesis Terpenoids Lilac, floral, terpenic Down C7H6O BenzAldehyde Metabolic pathways Aldehyde Sweet, bitter, almond, cherry Down C8H8O BenzAldehyde, 2-methyl- Metabolic pathways Aldehyde Mild floral, sweet Down C10H18O Bicyclo[3.1.0]hexan-2-ol,
2-methyl-5-(1-methylethyl)-, (1α,2β,5α)-Metabolic pathways, Biosynthesis of secondary metabolites, Monoterpenoid biosynthesis Terpenoids Balsam Down C15H24 Naphthalene, 1,2,3,5,6,8a-hexahydro-4,7-dimethyl-1-(1-methylethyl)-, (1S-cis)- Metabolic pathways, Biosynthesis of secondary metabolites, Sesquiterpenoid and triterpenoid biosynthesis Terpenoids Thyme, herbal, woody, dry Up C7H6O2 2-hydroxy-BenzAldehyde Metabolic pathways Aldehyde Medical, spicy, cinmon, wintergreen, cooling Down C8H14O2 3-Hexen-1-ol, acetate, (Z)- Biosynthesis of secondary metabolites, α-Linolenic acid metabolism Ester Fresh, green, sweet, fruity, ba--, apple, grassy Down C15H24 α-Farnesene Biosynthesis of secondary metabolites, Sesquiterpenoid and triterpenoid biosynthesis Terpenoids Citrus, herbal, lavender, bergamot, myrrh, neroli, green Up C15H24 (E)-1-Methyl-4-(6-methylhept-5-
en-2-ylidene)cyclohex-1-eneBiosynthesis of secondary metabolites, Sesquiterpenoid and triterpenoid biosynthesis Terpenoids − Up C15H24O 2,6,10-Dodecatrienal,
3,7,11-trimethyl-, (E,E)-Biosynthesis of secondary metabolites, Terpenoid backbone biosynthesis, Sesquiterpenoid and triterpenoid biosynthesis Terpenoids − Down C15H24 (E)-β-Famesene Sesquiterpenoid and triterpenoid biosynthesis Terpenoids Woody, citrus, herbal, sweet Up C10H16O Bicyclo[3.1.1]hept-2-ene-2-methanol, 6,6-dimethyl- Limonene and pinene degradation Terpenoids Woody, minty Down C10H16O 3-Oxatricyclo[4.1.1.0(2,4)]octane, 2,7,7-trimethyl- Limonene and pinene degradation Terpenoids Green Down − indicates no annotation of substance. Flavor omics analysis of differential volatile compounds
-
One hundred and fourty-three differential volatile compounds were annotated to 159 sensory flavors (Supplementary Table S3). The top 10 sensory flavors with the highest number of annotations were green (23), fruity (21), herbal (14), woody (14), sweet (13), floral (9), fresh (8), fatty (8), citrus (8), and earthy (7) (Fig. 5a), which were the most important sensory flavors for SF and WF. The top 10 differential volatile compounds with high numbers of sensory flavor features annotation were Hexanoic acid, propyl ester (Ester), 3-Hexen-1-ol,acetate,(Z)-(Ester), Butanoic acid,hexyl ester (Ester), Butanoic acid, octyl ester (Ester), Fenchone (Terpenoids), Isocyclocitral (Aldehyde), Pyrazine, 2-methyl-5-(1-methylethyl)-(Heterocyclic compound), Heptanal (Aldehyde), and Geranyl isobutyrate (Ester), which were the most important differential volatile compounds of sensory flavors for SF and WF (Fig. 5b).
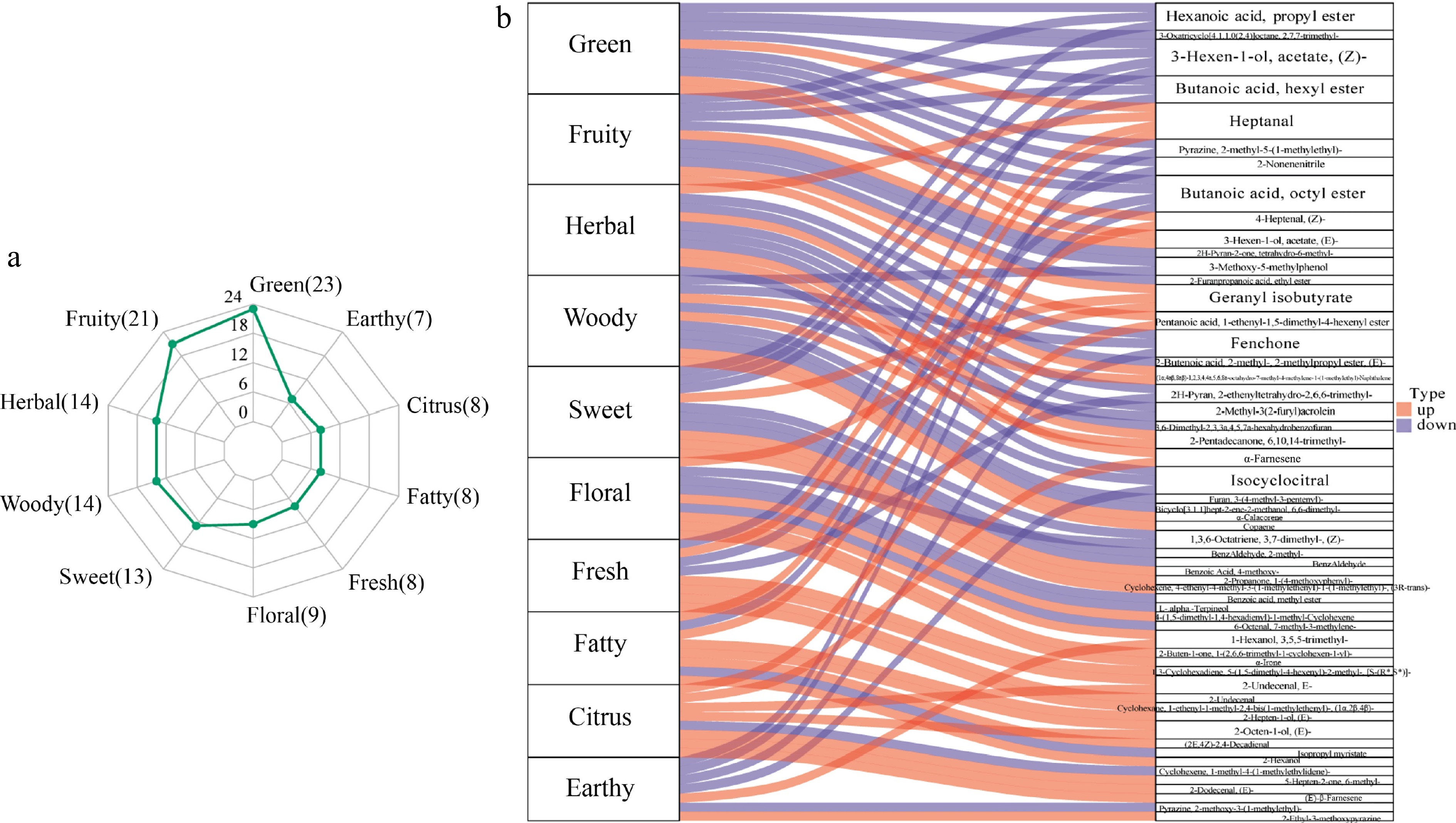
Figure 5.
(a) Radar map for analysis of differential metabolite sensory flavor characteristics. (b) Sankey diagram of flavor omics.
Compared with WF, SF mainly showed green, fruity, herbal, woody, sweet, and earthy, the relevant substances were Hexanoic acid, propyl ester (Ester), 3-Hexen-1-ol,acetate,(Z)- (Ester), Butanoic acid,hexyl ester (Ester), Butanoic acid,octyl ester (Ester), Fenchone (Terpenoids), Isocyclocitral (Terpenoids), Pyrazine, 2-methyl-5-(1-methylethyl)-(Terpenoids), etc (Fig. 5b). WF mainly showed more floral, fresh, fatty, and citrus than SF, according to a higher number of up-regulated metabolites for SF_vs_WF comparison, including 2-Undecenal,E-(Aldehyde), 2-Octen-1-ol,(E)-(Alcohol), 2-Dodecenal,(E)-(Aldehyde), (E)-β-Famesene (Terpenoids), etc (Fig. 5b).
-
Meteorological data differ greatly between the two crop seasons in Guangxi (China). The active accumulated temperature, the effective accumulated temperature, the daily average temperature, and the relative humidity for the summer growing season were higher than those of the winter growing season[3,24,29], which were in line with the present research. However, there were more hours of high temperatures over 35 °C in the winter growing season than that in the summer growing season. This result is the opposite of other study findings[3,24]. Consistently with previous studies[24,29], the solar radiation intensity and cumulative solar radiation was higher in the winter growing season. According to the results of this research and literary references, meteorological data for two crop seasons varies by year.
In the present research, 620 volatile compounds in 15 categories were detected in summer and winter fruits, including 122 terpenoids, 115 esters, 99 heterocyclic compounds, 60 hydrocarbons, 52 ketones, 48 alcohols, 47 aldehydes, 31 aromatics, 11 amines, 11 acids, eight phenols, seven nitrogen compounds, three halogenated hydrocarbons, two sulfur compounds, and four others. These results indicated that terpenoids were the main volatile characteristic substances of 'Ruidu Kemei', followed by esters. It has been confirmed that terpenoids were the characteristic aroma components of muscat flavored varieties, which was consistent with the present study[30].
Grape cultivation in the field was greatly impacted by climate conditions. Berries were influenced greatly by their growing environment in terms of chemical composition. Due to variations in climate between the summer and winter growing seasons, the most important metabolites of grapes perform differently under a two-crop-a-year cultivation system, such as flavonoids[24,29], phenols, carotenoids[28], and volatiles[3]. The present study showed clear differences in the concentration of volatile compounds in response to meteorological data for two crop seasons, which verified the findings of previous research[3], while the compounds of volatiles mainly remained similar for the volatile compounds depending largely on the genotype of the grape cultivar rather than the growing environment[11]. However, when compared with WF, higher volatile compound concentration was observed in berries of summer, which would be caused by more hours of high temperatures over 35 °C during the winter growing season. Generally, lower temperatures were conducive to the accumulation of aromatic substances. Furthermore, this data appears to be related to the higher active accumulated temperatures in the summer growing season, which favored the grape ripening and volatile accumulation in the grape berries[10,31].
To determine the distinction of volatile compounds between summer fruits and winter fruits, 143 significant different metabolites were selected. For number and relative content, terpenoids, heterocyclic compound, ester and aromatics showed greater differences than other compounds. In particular, for the top 20 substances using Log2FC, terpenoids (such as [1α,4aα,8aα]-1,2,4a,5,6,8a-hexahydro-4-7-dimethyl-1-[1-methylethyl]naphthalene) were mainly up-regulated in WF, while esters (such as 3-Hexen-1-ol, acetate, (Z)-) and aromatics (such as Benzene, (1-methoxypropyl)-) were up-regulated in SF, since heat and sunlight stress can reduce the aromatic content of grapes, while less solar radiation intensity favored the higher level of aromatics[9]. The most likely precursors for the esters were lipids and amino acids. Their metabolism during ripening will therefore play an important role in determining both the levels and types of esters formed[32]. It has been reported that cluster sunlight exposure in viticulture in dry-hot climates caused a notable decrease in esters, including ethyl hexanoate and hexyl acetate[15]. Sunlight was advantageous for accumulating terpenoids[3], the activation of terpene synthase genes (VvTPS54 and VvTPS56) and the synthesis of carotenoids in grapes, subsequently leading to the accumulation of terpenoids and norisoprenoids[33,34]. As reported, higher solar radiation intensity and cumulative solar radiation enhanced accumulation of terpenes[12]. Zhang et al. indicated that VvDXS2 and VvDXR were partially linked to differential terpene accumulation for different illumination conditions[35]. Sun et al. found that grape berries grown in rain shelters contain lower levels of terpenoids and norisoprenoids during development, possibly as a result of less light, inhibiting isoprenoids during development[10]. In this study, more hours of high temperatures over 35 °C in the winter growing season than that in the summer growing season, higher solar radiation intensity, and cumulative solar radiation still promoted sesquiterpenoid and triterpenoid biosynthesis, monoterpenoid biosynthesis, limonene, and pinene degradation (Fig. 4b). These results reconfirmed that the increased light exposure was beneficial for terpene accumulation, which could infer that the negative effect of the elevated berry temperature might be surpassed by the beneficial effect of increased synthesis of terpenes induced by light. However, Friedel et al. made an opposite judgment[33]. Thus, grape cultivars might respond differently to climate. In summary, for 'Ruidu Kemei', WF performed better in terpenoids, whereas SF displayed better in esters and aromatics. Based on previous studies of other grape varieties[3], it could be concluded that WF probably always forms higher concentrations of terpenes than SF under a two-crop-a-year cultivation system in the Guangxi region of South China, which has a typical subtropical humid monsoon climate.
Among other qualities, aroma flavor contributes to consumers' acceptance of table grapes[30]. Table grapes' flavor was generally determined by their free volatiles since they were directly detectable and tasteable[36]. Flavors varied from different ingredients and different concentrations of volatile substances. In the present study, the different relative content of volatile substances was the reason for the different flavors of grapes in two growing seasons. Fruit aroma profile visually displayed that the most important sensory flavors for 'Ruidu Kemei' were green, fruity, herbal, woody, sweet, floral, fresh, fatty, citrus, and earthy. Green and fruity were the most critical aroma for 'Ruidu Kemei', due to most volatiles annotated. Green, fruity, herbal, woody, sweet, and earthy were more prominent in the SF, the relevant substances were Hexanoic acid, propyl ester, 3-Hexen-1-ol,acetate,(Z)-, Butanoic acid,hexyl ester, Butanoic acid, octyl ester, Fenchone, Isocyclocitral, Pyrazine, 2-methyl-5-(1-methylethyl)-, etc (Fig. 5b). Compared with SF, for WF, floral was the most prominent fruit smell, followed by fresh and fatty smell, and then citrus smell, the metabolites with the greatest contribution were 2-Undecenal,E-, 2-Octen-1-ol,(E)-, 2-Dodecenal,(E)-, (E)-β-Famesene, etc (Fig. 5b). In general, seasonal differences can be observed in the sensory properties of grape berries from the same variety[37]. Floral dominated in WF for performing better in terpenoids than SF. Due to their association with floral scents, terpenoids may attract appropriate pollinators and facilitate reproduction[37].
-
In the present research, 620 volatile compounds in 15 categories were detected in summer and winter fruits by a GC-MS/MS-based metabolomics approach. Terpenoids were the main volatile characteristic substances of 'Ruidu Kemei', followed by esters. Meteorological data for two crop seasons varied by years. The variational climatic factors in the summer and winter growing seasons were responsible for the difference in volatile metabolites between the two crops of grapes. Compared with the WF, higher active accumulated temperatures in the summer growing season contributed to higher volatile compound concentration in SF. Moreover, terpenoids, heterocyclic compounds, esters, and aromatics showed greater differences than other compounds between SF and WF. In addition, it was demonstrated that the winter cropping cycle promoted the biosynthesis of terpenoids by higher solar radiation intensity and cumulative solar radiation, which lead to more floral fruit smell than SF. On the contrary, more esters and aromatics were observed in SF in response to less solar radiation intensity, cumulative solar radiation and higher active accumulated temperatures in the summer growing season. Flavor omics analysis presented that the most important sensory flavors for 'Ruidu Kemei' were green, fruity, herbal, woody, sweet, floral, fresh, fatty, citrus, and earthy. Green and fruity were the most critical aroma for 'Ruidu Kemei', due to the most volatiles annotated. Green, fruity, herbal, woody, sweet, and earthy were more prominent in the SF. Floral was the most prominent fruit smell in WF. Clarification the characteristics of aroma substances of grape berries in two growing seasons can provide a basis for the scientific control of grape aroma, the improvement of grape quality, and the optimization of grape cultivation technology.
-
The authors confirm contribution to the paper as follows: study conception and design: Guo R, Lin L, Zhang Y; data collection: Lin L, Yu H, Liu J, Shi X; analysis and interpretation of results: Yu H, Huang G; draft manuscript preparation: Yu H; review & editing, project administration and funding acquisition: Guo R, Lin L. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This work was supported by grants from the Guangxi Key Research and Development Program (GuikeAB19245031), the special project for basic scientific research of Guangxi Academy of Agricultural Sciences (Guinongke2021YT127 & Guinongke2023YM111).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Huan Yu, Rongrong Guo
- Supplemental Table S1 The information of all detected metabolites.
- Supplemental Table S2 49 of the 620 metabolites arnotated to 20 KEGG pathways.
- Supplemental Table S3 143 differential volatile compounds amnotated to 159 sensory flavors.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu H, Guo R, Liu J, Shi X, Huang G, et al. 2024. Metabolome provides new insights into the volatile substances in 'Ruidu Kemei' grapes under the two-crop-a-year cultivation system. Fruit Research 4: e035 doi: 10.48130/frures-0024-0029 

Metabolome provides new insights into the volatile substances in 'Ruidu Kemei' grapes under the two-crop-a-year cultivation system
- Received: 29 May 2024
- Revised: 11 July 2024
- Accepted: 31 July 2024
- Published online: 04 November 2024
Abstract: In subtropical regions, the implementation of a two-crop-a-year cultivation system depends on local climatic conditions. Grape volatile compounds vary greatly with the season, due to climate differences, which lead to extreme differences between summer grape fruits (SF) and winter grape fruits (WF). In the present study, a gas chromatography-mass spectrometer (GC-MS) was used to analyze volatile compounds from 'Ruidu Kemei' grapes grown under the two-crop-a-year cultivation system. Results showed that fruits in summer and winter contained 620 volatile compounds in 15 categories. Among them, terpenoids constituted the largest group, with 122 metabolites, followed by 115 esters. This indicated that the main volatile characteristic substances of 'Ruidu Kemei' were terpenoids and esters. Higher volatile compounds in SF might be associated with higher active accumulated temperatures in the summer growing season. In addition, terpenoids, heterocyclic compounds, esters, and aromatics showed greater differences than other compounds between SF and WF. Regarding terpenoids, WF exhibited superior performance, while SF performed better in esters and aromatics. For WF, higher solar radiation intensity promoted the biosynthesis of terpenoids, which lead to more floral characteristics than SF. According to the flavor omics analysis, 'Ruidu Kemei' was primarily characterized by green, fruity, herbal, woody, sweet, floral, fresh, fatty, citrus, and earthy. In the SF, green and fruity flavors were more prominent, while floral was the dominant fruity aroma in WF. This work provides new insights into the metabolism of volatiles in summer and winter grapes and reference for the selection and promotion of varieties with suitable aromas for a two-crop-a-year cultivation system.