








-
Kiwifruit (Actinidia chinensis), known as the 'King of fruits', has important economic, ecological, and social benefits. However, its susceptibility to fungal infection after harvest causes huge economic losses and poses potential safety concerns. Botrytis cinerea, caused by Botrytis cinerea, is one of the main postharvest diseases of kiwifruit, which can lead to a spoilage rate above 20%[1].
Plant hormones[2] are chemicals produced by plants, which, at low concentrations, affect their vital activities by regulating physiological processes, such as growth[3], development, reproduction, and stress response. Jasmonic acid (JA)[4] is a plant hormone closely related to plant resistance that plays an important role in plant growth, development, and stress response. It is widely regarded as a signaling molecule in plants that regulates physiological processes, such as disease resistance[5], stress resistance[6], flowering, and fruit ripening. JA also participates in plant defense response[7]. Synthesis of JA in plants increases under unfavorable environments[8], thereby promoting the initiation of defense mechanisms. Together with other plant hormones, JA regulates plant growth, development, and stress response[9].
SHAM is an inhibitor of lipoxygenase (LOX), a key enzyme in the JA biosynthetic pathway[10]. Therefore, SHAM may be involved in the regulation of plant responses to stress[11−13]. In tomato, 5-aminolevulinic acid (ALA) was reported to enhance oxidative stress tolerance at low temperatures through the nitric oxide (NO), JA, and hydrogen peroxide (H2O2) signaling pathways. Through plant culture, biochemical analysis, and gene expression studies, it was found that ALA pretreatment could enhance the activity of antioxidant enzymes in tomato and enhanced the tolerance to cold injury through a synergistic effect of JA and H2O2 and a downstream signaling effect of NO. These results indicated that ALA plays an important role in regulating the response of tomato[14] to low temperatures, providing a potential molecular mechanism for improving the cold tolerance of crops. In Brassica napus[15], the inhibitory effects of polyunsaturated fatty acids (PUFA) and methyl jasmonate (MeJA) on osmosis-induced proline accumulation were studied. PUFA and MeJA inhibited this accumulation, and MeJA exhibited an antagonistic effect on abscisic acid (ABA), a plant hormone that affects drought stress response by regulating proline metabolism. In a potato tuber formation study[16], the effects of SHAM on theobromine-induced tuber formation, as well as onwhile endogenous JA and tubular acid (TA) levels and LOX activity, were investigated. SHAM inhibited theobromine-induced potato tuber formation, which is related to JA and TA biosynthesis and regulation of LOX activity. The effects of alternative oxidase (AOX) activity on phenol metabolism were investigated in MeJA-treated hairy root cultures of carrot[17], employing the AOX inhibitors, SHAM and PG. AOX activity was positively correlated with phenylpropane biosynthesis, and MeJA enhanced the accumulation of phenolic acids, flavonoids, and lignin by upregulating AOX and phenylalanine ammonia-lyase (PAL) activities. Moreover, in the endophytic fungus Trichoderma atroviride D16 from Salvia miltiorrhiza[18], the effects of smoke-isolated butenolactone (KAR1) on tanshinone I (T-I) production were investigated, and JA production and signal transduction were analyzed. KAR1 increased the production of T-I by inducing the production of JA, which indicated the role of JA signaling in this process and provided valuable information for improving the content of medicinal ingredients.
The roles of SHAM in plant disease and defense have been extensively discussed. However, research on its effects as an exogenous activator on the postharvest disease of fruits is insufficient, and the mechanisms underlying its effects remain unclear. In this study, the effect of SHAM on Botrytis cinerea resistance in kiwifruit was investigated. We performed high-throughput transcriptome sequencing of SHAM-treated kiwifruit samples, enrichment analysis of differentially expressed genes (DEGs), and analysis of the main metabolic pathways affected by SHAM to unravel the molecular mechanisms involved. The present results provide potential strategies for the biocontrol of postharvest diseases and the promotion of sustainable agriculture.
-
Selected 'Red Yang' kiwifruit (Actinidia chinensis cv. Hongyang) were provided by Sanlei Tiantian Agricultural Development Co., Ltd., Chongqing, China (108.83° E, 29.59° N). 'Hongyang' kiwifruit exhibiting good shape, uniform size, and absence of pests and diseases were selected and transported to the laboratory within 6 h of harvesting. The fruits were disinfected by soaking in 2% sodium hypochlorite solution for 2 min, rinsed with disinfected distilled water, and dried at 25 °C for further use.
SHAM treatment and Botrytis cinerea infection
-
Ten microliter SHAM solution (0.1 mol/L) was injected into the pores of dried fruit, which were subsequently dried again. Thereafter, sterile water was injected three times and Botrytis cinerea (104 spores/mL) was inoculated 24 h later. Diseased and healthy junction samples were collected at different time points (0, 24, 72, and 120 h, corresponding to S_0h, S_24h, S_72h, and S_120h, respectively) and diameters of the fungal lesions were determined using ImageJ software.
Total RNA extraction and transcriptome sequencing
-
The SHAM-treated kiwifruit were inoculated with Botrytis cinerea on day 0, 1, 3, and 5 in triplicate for each of the time points. Total RNA was extracted using a magnetic bead assay kit. The RNA samples were assessed for quality for use in the construction of cDNA library. Transcriptome sequencing was commissioned to Nanjing Ovisen Gene Technology Co., Ltd (Nanjing, China). The constructed library was initially quantified using a Qubit2.0 fluorometer, library insert size detection, and qRT-PCR. The qualified samples were sequenced on an Illumina platform. Finally, the PCR products were purified (AMPure XP system), and the library quality was assessed using an Agilent Bioanalyzer 2100 system. The raw RNA-seq data were deposited in the NCBI SRA database (Accession number: PRJNA1104815).
Quality control
-
Raw data (raw reads) in fastq format were first processed using in-house Perl scripts. In this step, clean data (clean reads) were obtained by removing the reads containing adapters, poly-N, and low-quality reads from the raw data. Simultaneously, the Q20, Q30, GC content and sequence duplication levels of the clean data were calculated. All downstream analyses were based on clean high-quality data, and adaptor sequences and low-quality sequence reads were removed from the datasets. The clean reads were mapped to the reference genome sequence using STAR. Only reads with a perfect match or one mismatch were further analyzed and annotated based on the reference genome. HTSeq (v 0.5.4) was used to count the number of reads mapped to each gene. Gene expression levels were estimated as fragments per kilobase of transcript per million mapped reads (FPKM).
Differential gene expression analysis
-
Differential gene expression analysis of two conditions or groups was performed using the DESeq R package (version 1.10.1). DESeq provides statistical routines for determining differential expression in digital gene expression data, using a model based on a negative binomial distribution. The resulting p values were adjusted using Benjamini and Hochberg's approach for controlling the false discovery rate (FDR). Genes with an adjusted p-value < 0.05 found using DESeq were assigned as differentially expressed. The sequence available at
http://kiwifruitgenome.org/ was used as the reference genome. Different infection times (0, 24, 72, and 120 h) were selected, and DEGs between the SHAM treatment and control groups (SHAM120h : CK120h) were identified based on the filtering criteria of false-discovery rate (FDR) < 0.05 and |log2FC| greater than 1 and less than −1. A volcano plot was generated using the OmicShare tool to analyze the DEGs at different infection times (0, 24, 72, and 120 h). The Venn mapping tool was used to compare DEGs associated with Botrytis cinerea. These DEGs were subjected to the KEGG pathway enrichment analysis. The DEGs expression have been validated by qRT-PCR, and the primers used are listed in Table 1.Table 1. Primers used in the present study.
Purpose Gene ID Primers (5'-3') qRT-PCR β-Actin F ACCGACATTTTCCTGCAACC R AGCAGCTGAGGTTGATCTGT qRT-PCR CEY00_Acc00618 F CGTTGGGTTTGTGCTCTGAAA R TCCTCAAACATGTCACCGGA qRT-PCR CEY00_Acc11227 F CCCATAATCGAAAAGGCCGG R TCACCAGATCGATCAGCCTC qRT-PCR CEY00_Acc16583 F CTTGAACCACCGCAACTTGA R CCTACGCATCTTCCTCCAGT qRT-PCR CEY00_Acc20943 F CTCCGGTGATCTGATCCCTCT R ATTCACTAGTGCCAGGCCTT qRT-PCR CEY00_Acc22367 F CCGAGACTTTGTTGCTTGCTC R CAGACCCTCCCGATTCTCTC qRT-PCR CEY00_Acc23338 F TTCCAACTCCTCCATCCCAC R GCCGATGATGGGTGAAGAAC qRT-PCR CEY00_Acc24966 F ACTGGTCGTCTGCTCTGAAA R GCCCCATCACTATCCGGTAA qRT-PCR CEY00_Acc27491 F TTGTTCAGTTGATGCAGGGCC R AAACAGTGCACCCGAGTTTC qRT-PCR CEY00_Acc29568 F ACCAGACTGCCCATTCATGA R GCACGGCTTTGTCAGATGAT Statistical analyses
-
Statistical analyses were performed using SPSS 18.0. The Student's t-test was used to compare physiological indices and relative qRT-PCR expression data (p < 0.05).
-
The appearance and morphology of kiwifruit in the SHAM and CK groups differed at different days postinfection (dpi) (Fig. 1). At 0, 1, 3, and 5 dpi, the lesion diameters in the SHAM and CK groups were 0.30 and 0.30 cm, 0.39 and 0.37 cm, 2.41 and 1.49 cm, and 3.03 and 2.02 cm, respectively. At 0 dpi, the lesion diameters in both kiwifruit groups were in the 0–3 cm range. However, differences between the two groups increased over time. At 5 dpi, the lesion diameter in the CK group increased to 2.02 cm, whereas that in the SHAM group was 3.03 cm. Thus, the lesion diameter in the SHAM group was 1.02 cm higher than that in the CK group, which further confirmed that SHAM treatment reduced the resistance of kiwifruit to Botrytis cinerea. These results indicate that appropriate SHAM treatment can reduce the resistance of kiwifruit to Botrytis cinerea.
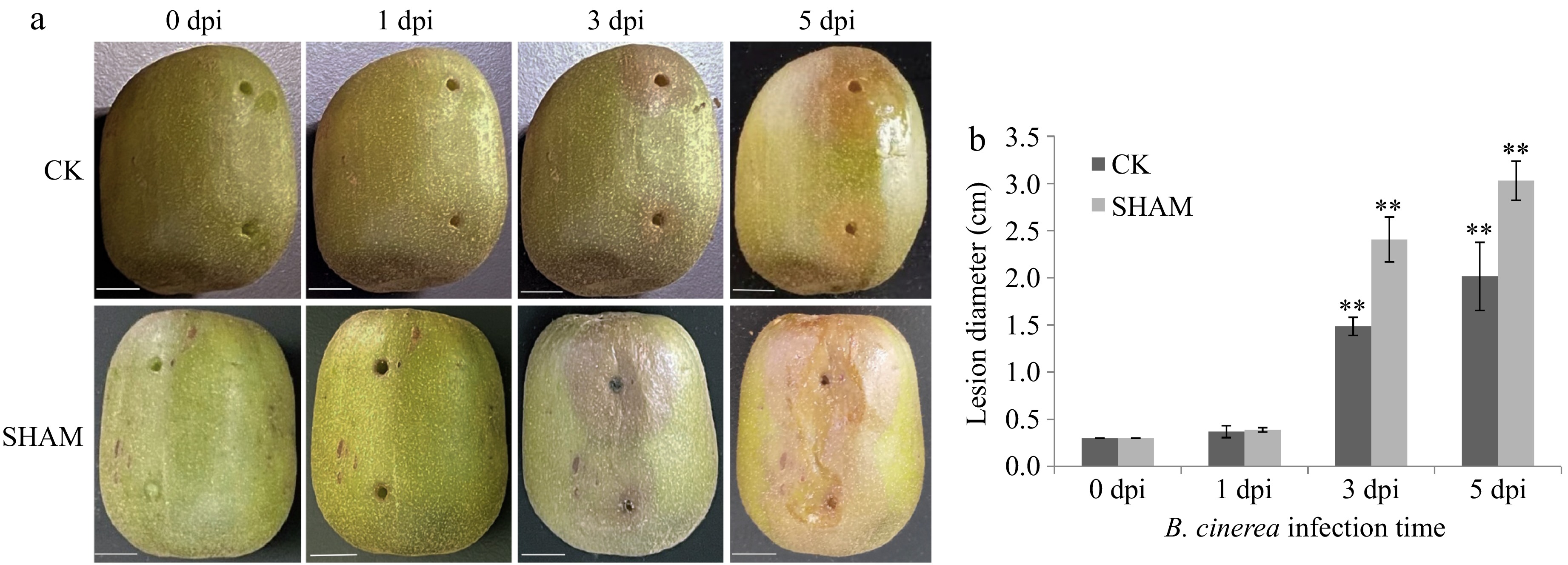
Figure 1.
Changes in the appearance, morphology, and lesion diameter of kiwifruit in the SHAM and CK groups induced by Botrytis cinerea at different days postinfection. (a) Changes in the morphological characteristics of kiwifruit at 0, 1, 3, and 5 d post infection (dpi). (b) Changes in the lesion diameter in kiwifruit in the SHAM and CK groups at different time points. '**' represents extremely significantly different (p < 0.01).
Transcriptome sequence analysis
Sequencing data statistics
-
The SHAM-treated kiwifruit was inoculated with Botrytis cinerea, and the inoculation was repeated three times at three time points on day 0 (C_0h vs S_0h), day 1 (C_24h vs S_24h), day 3 (C_72h vs S_72h), and day 5 (C_120h vs S_120h). Transcriptome sequencing was performed using the kiwifruit tissue. A total of 24 groups of cDNA libraries were sequenced. The average values for Q20 and Q30 were 97.67% and 93.31%, respectively, indicating a good sequencing quality. Statistics of the sequencing data (such as clean reads and clean bases) and a comparison of clean reads with reference genomes are provided in Tables 2 & 3.
Table 2. Statistics of sequencing data.
Sample name Raw reads Raw bases Clean reads Clean bases Error rate Q20 Q30 GC content C_0h_1 47524076 7.16G 47391498 7.16G 0.03% 97.65% 93.24% 46.76% C_0h_2 46500794 7.01G 46365486 7G 0.03% 97.81% 93.62% 46.71% C_0h_3 40160680 6.05G 40052962 6.05G 0.03% 97.69% 93.27% 46.65% C_24h_1 42614752 6.42G 42488828 6.42G 0.03% 97.58% 93.05% 46.57% C_24h_2 43818432 6.6G 43697220 6.6G 0.03% 97.61% 93.09% 46.66% C_24h_3 39659046 5.98G 39568666 5.97G 0.03% 97.84% 93.61% 46.81% C_72h_1 39707836 5.98G 39601106 5.98G 0.03% 97.59% 93.07% 46.75% C_72h_2 39646984 5.98G 39546362 5.97G 0.03% 97.83% 93.63% 46.86% C_72h_3 40853092 6.16G 40733706 6.15G 0.03% 97.63% 93.18% 46.87% C_120h_1 38720330 5.83G 38616554 5.83G 0.03% 97.64% 93.17% 47.17% C_120h_2 45336366 6.83G 45202468 6.83G 0.03% 97.63% 93.15% 46.95% C_120h_3 42699226 6.43G 42577918 6.43G 0.03% 97.70% 93.31% 46.80% S_0h_1 47755908 7.2G 47622990 7.19G 0.03% 97.70% 93.31% 46.48% S_0h_2 45493394 6.85G 45362482 6.85G 0.03% 97.83% 93.63% 46.61% S_0h_3 42211600 6.36G 42099856 6.36G 0.03% 97.69% 93.28% 46.62% S_24h_1 44252586 6.67G 44112658 6.66G 0.03% 97.57% 93.04% 46.90% S_24h_2 38169744 5.75G 38062452 5.75G 0.03% 97.62% 93.15% 46.95% S_24h_3 38973972 5.87G 38878598 5.87G 0.03% 97.87% 93.72% 47.05% S_72h_1 44924206 6.77G 44725720 6.75G 0.03% 97.32% 92.49% 47.12% S_72h_2 41152860 6.2G 41023726 6.19G 0.03% 97.86% 93.72% 47.19% S_72h_3 40050140 6.04G 39942390 6.03G 0.03% 97.62% 93.14% 46.96% S_120h_1 37938536 5.72G 37839310 5.71G 0.03% 97.61% 93.12% 47.26% S_120h_2 42220850 6.36G 42087276 6.36G 0.03% 97.58% 93.15% 47.34% S_120h_3 42243486 6.36G 42114890 6.36G 0.03% 97.69% 93.37% 47.24% Table 3. Comparison of rReads and reference sequences.
Sample name Total reads Total mapped Multiple mapped Uniquely mapped Read-1 Read-2 Readsmapto'+' Readsmapto'–' Non-splicereads Splicereads C_0h_1 47391498 44964944 (94.88%) 1713494 (3.62%) 43251450 (91.26%) 21625725 (45.63%) 21625725 (45.63%) 21625725 (45.63%) 21625725 (45.63%) 26448459 (55.81%) 16802991 (35.46%) C_0h_2 46365486 44059000 (95.03%) 1625532 (3.51%) 42433468 (91.52%) 21216734 (45.76%) 21216734 (45.76%) 21216734 (45.76%) 21216734 (45.76%) 25835515 (55.72%) 16597953 (35.8%) C_0h_3 40052962 38138198 (95.22%) 1437330 (3.59%) 36700868 (91.63%) 18350434 (45.82%) 18350434 (45.82%) 18350434 (45.82%) 18350434 (45.82%) 22123727 (55.24%) 14577141 (36.39%) C_24h_1 42488828 39228228 (92.33%) 1541524 (3.63%) 37686704 (88.7%) 18843352 (44.35%) 18843352 (44.35%) 18843352 (44.35%) 18843352 (44.35%) 23334117 (54.92%) 14352587 (33.78%) C_24h_2 43697220 41131072 (94.13%) 1626420 (3.72%) 39504652 (90.41%) 19752326 (45.2%) 19752326 (45.2%) 19752326 (45.2%) 19752326 (45.2%) 23913823 (54.73%) 15590829 (35.68%) C_24h_3 39568666 36938290 (93.35%) 1450522 (3.67%) 35487768 (89.69%) 17743884 (44.84%) 17743884 (44.84%) 17743884 (44.84%) 17743884 (44.84%) 21343865 (53.94%) 14143903 (35.75%) C_72h_1 39601106 37270004 (94.11%) 1464402 (3.7%) 35805602 (90.42%) 17902801 (45.21%) 17902801 (45.21%) 17902801 (45.21%) 17902801 (45.21%) 21764204 (54.96%) 14041398 (35.46%) C_72h_2 39546362 37400570 (94.57%) 1504068 (3.8%) 35896502 (90.77%) 17948251 (45.39%) 17948251 (45.39%) 17948251 (45.39%) 17948251 (45.39%) 22088098 (55.85%) 13808404 (34.92%) C_72h_3 40733706 38245790 (93.89%) 1521084 (3.73%) 36724706 (90.16%) 18362353 (45.08%) 18362353 (45.08%) 18362353 (45.08%) 18362353 (45.08%) 22622406 (55.54%) 14102300 (34.62%) C_120h_1 38616554 33816150 (87.57%) 1574710 (4.08%) 32241440 (83.49%) 16120720 (41.75%) 16120720 (41.75%) 16120720 (41.75%) 16120720 (41.75%) 19925731 (51.6%) 12315709 (31.89%) C_120h_2 45202468 41753966 (92.37%) 1695002 (3.75%) 40058964 (88.62%) 20029482 (44.31%) 20029482 (44.31%) 20029482 (44.31%) 20029482 (44.31%) 24363679 (53.9%) 15695285 (34.72%) C_120h_3 42577918 39318158 (92.34%) 1846690 (4.34%) 37471468 (88.01%) 18735734 (44%) 18735734 (44%) 18735734 (44%) 18735734 (44%) 22576173 (53.02%) 14895295 (34.98%) S_0h_1 47622990 43144188 (90.6%) 1804166 (3.79%) 41340022 (86.81%) 20670011 (43.4%) 20670011 (43.4%) 20670011 (43.4%) 20670011 (43.4%) 25591719 (53.74%) 15748303 (33.07%) S_0h_2 45362482 41979524 (92.54%) 1554640 (3.43%) 40424884 (89.12%) 20212442 (44.56%) 20212442 (44.56%) 20212442 (44.56%) 20212442 (44.56%) 24638686 (54.32%) 15786198 (34.8%) S_0h_3 42099856 39315626 (93.39%) 1510886 (3.59%) 37804740 (89.8%) 18902370 (44.9%) 18902370 (44.9%) 18902370 (44.9%) 18902370 (44.9%) 22678429 (53.87%) 15126311 (35.93%) S_24h_1 44112658 41846144 (94.86%) 1662310 (3.77%) 40183834 (91.09%) 20091917 (45.55%) 20091917 (45.55%) 20091917 (45.55%) 20091917 (45.55%) 24359883 (55.22%) 15823951 (35.87%) S_24h_2 38062452 36153600 (94.98%) 1412136 (3.71%) 34741464 (91.27%) 17370732 (45.64%) 17370732 (45.64%) 17370732 (45.64%) 17370732 (45.64%) 21234325 (55.79%) 13507139 (35.49%) S_24h_3 38878598 36883350 (94.87%) 1465624 (3.77%) 35417726 (91.1%) 17708863 (45.55%) 17708863 (45.55%) 17708863 (45.55%) 17708863 (45.55%) 21502098 (55.31%) 13915628 (35.79%) S_72h_1 44725720 41128742 (91.96%) 1876526 (4.2%) 39252216 (87.76%) 19626108 (43.88%) 19626108 (43.88%) 19626108 (43.88%) 19626108 (43.88%) 24421759 (54.6%) 14830457 (33.16%) S_72h_2 41023726 38514036 (93.88%) 1702480 (4.15%) 36811556 (89.73%) 18405778 (44.87%) 18405778 (44.87%) 18405778 (44.87%) 18405778 (44.87%) 22568830 (55.01%) 14242726 (34.72%) S_72h_3 39942390 37195056 (93.12%) 1558874 (3.9%) 35636182 (89.22%) 17818091 (44.61%) 17818091 (44.61%) 17818091 (44.61%) 17818091 (44.61%) 21984331 (55.04%) 13651851 (34.18%) S_120h_1 37839310 34998938 (92.49%) 1566774 (4.14%) 33432164 (88.35%) 16716082 (44.18%) 16716082 (44.18%) 16716082 (44.18%) 16716082 (44.18%) 20896975 (55.23%) 12535189 (33.13%) S_120h_2 42087276 38673976 (91.89%) 1863200 (4.43%) 36810776 (87.46%) 18405388 (43.73%) 18405388 (43.73%) 18405388 (43.73%) 18405388 (43.73%) 23284707 (55.32%) 13526069 (32.14%) S_120h_3 42114890 38897188 (92.36%) 1769032 (4.2%) 37128156 (88.16%) 18564078 (44.08%) 18564078 (44.08%) 18564078 (44.08%) 18564078 (44.08%) 23428900 (55.63%) 13699256 (32.53%) Overall analysis of differentially expressed genes
-
Transcriptome sequencing revealed 37,159 expressed genes. According to the criteria of FDR < 0.05 and |log2FC| greater than 1, the DEGs between the SHAM treatment and control groups were identified (Fig. 2). In the S_24h vs S_0h comparison, 714 DEGs were identified, of which 272 genes were downregulated and 442 were upregulated. In the S_72h vs S_0h comparison, 1,511 DEGs were identified, of which 571 were downregulated and 940 were upregulated. A total of 4,181 DEGs were identified in the S_120h vs S_0h comparison, of which 1,448 were downregulated and 2,733 were upregulated. In the S_0h vs C_0h comparison, the number of DEGs (88) was small, the DEGs were not obvious, and the difference between the up- and downregulated genes was small, highlighting the authenticity of these results.
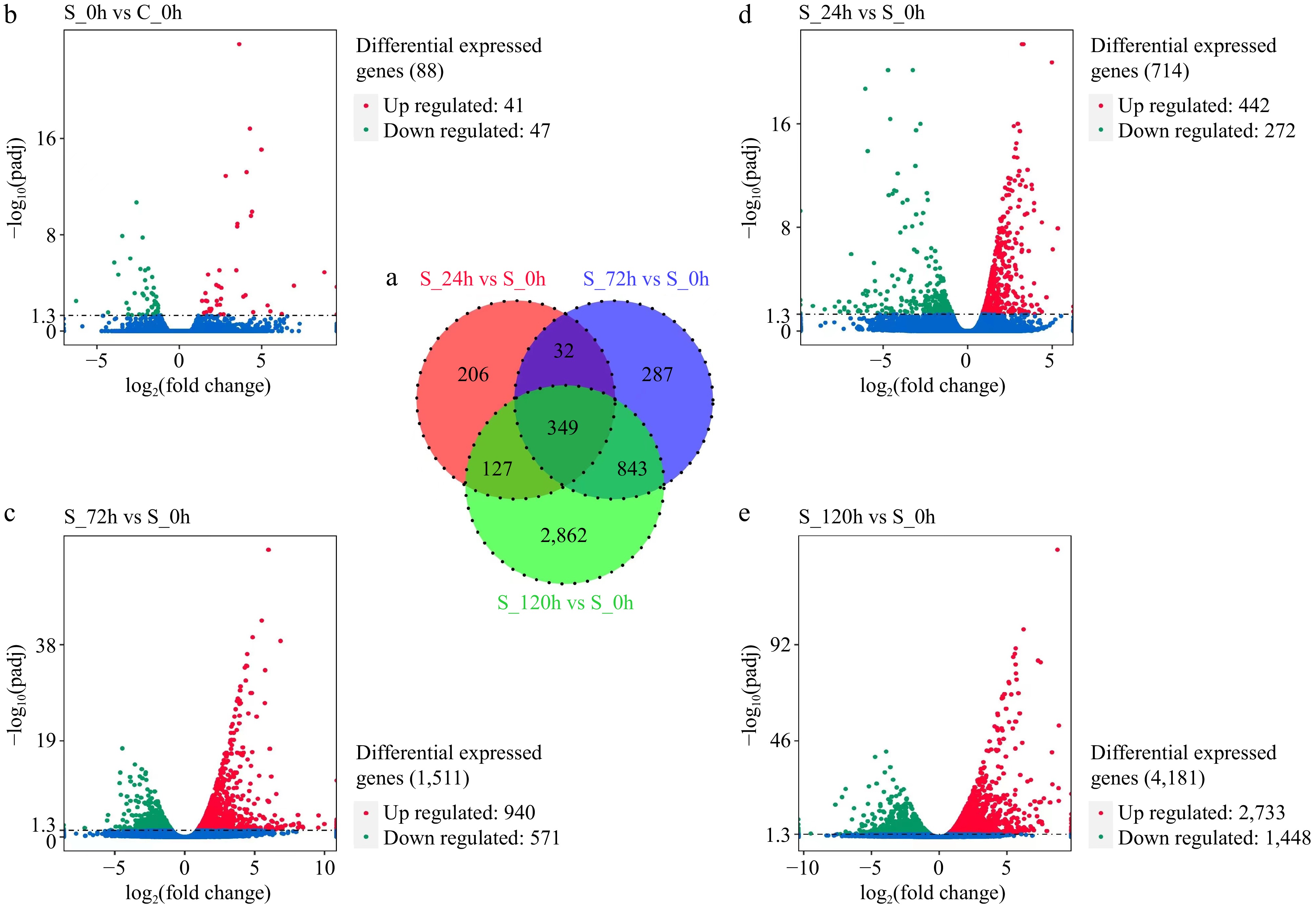
Figure 2.
(a) Venn diagram of differentially expressed genes (DEGs) between different groups (S_24h vs S_0h, S_72h vs S_0h, and S_120h vs S_0h). (b)–(e) Volcano plots of DEGs in different samples. Each dot represents a DEG. Red and green represent significantly up and downregulated DEGs.
The DEGs were associated with flavonoid biosynthesis (CEY00_Acc08970: chalcone synthase; CEY00_Acc24272: spermidine hydroxycinnamoyl transferase; CEY00_ACC24272: spermidine hydroxycinnamoyl transferase; CEY00_Acc16583: trans-cinnamate 4-monooxygenase; CEY00_Acc01005: dihydroflavonol 4-reductase; and CEY00_Acc32390: flavonoid 3',5'-hydroxylase), phenylpropanoid biosynthesis (CEY00_Acc02578: caffeic acid 3-O-methyltransferase; CEY00_Acc11227: caffeoyl-CoA O-methyltransferase; CEY00_Acc20943: phenylalanine ammonia-lyase; CEY00_Acc02557: 4-coumarate-CoA ligase like; and CEY00_Acc01851: cationic peroxidase), stilbene compound (CEY00_Acc11227: caffeoyl-CoA O-methyltransferase; CEY00_Acc24272: spermidine hydroxycinnamoyl transferase; CEY00_Acc16583: trans-cinnamate 4-monooxygenase; CEY00_Acc07635: spermidine hydroxycinnamoyl transferase; and CEY00_Acc29568: shikimate O-hydroxycinnamoyltransferase), MAPK signal pathways - Plant (CEY00_Acc22367: mitogen-activated protein kinase; CEY00_Acc03023: WRKY transcription factor 33; CEY00_Acc04863: endochitinase; CEY00_Acc20092: nucleoside diphosphate kinase; and CEY00_Acc00618: EIN3-binding F-box protein), plant hormone signal transduction (CEY00_Acc00550), indole-3-acetic acid-amido synthetase (CEY00_Acc24308: transcription factor like; CEY00_Acc00888: xyloglucan endotransglucosylase/hydrolase protein; CEY00_Acc11805: histidine kinase; and CEY00_Acc07445: basic form of pathogenesis-related protein endoplasmin like), and plant-pathogen interactions (CEY00_Acc23552: endoplasmin like; CEY00_Acc31880: pathogenesis-related transcriptional activator like; CEY00_Acc19497: disease resistance protein; CEY00_Acc07445: basic form of pathogenesis-related protein like; CEY00_Acc00158: RPM1-interacting protein).
In summary, these results proved that SHAM treatment regulates the resistance response of kiwifruit to Botrytis cinerea.
KEGG pathway enrichment analysis of differentially expressed genes
-
KEGG pathway enrichment analysis of the abovementioned DEGs was performed. In the S_24h vs S_0h_ALL comparison, 16 genes were enriched in the flavonoid biosynthesis pathway, 14 in the MAPK signaling pathway-plant, 10 in cysteine and methionine metabolism pathways, 12 in the starch and sucrose metabolism pathway, 10 in amino sugar and nucleotide sugar metabolism pathways, 19 in plant hormone signal transduction pathways, and four in the pentose phosphate pathway. In the S_72h vs S_0h_ALL comparison, 21 DEGs were enriched in carbon fixation in photosynthetic organisms pathways, 58 in carbon metabolism pathways, 43 in amino acid biosynthesis pathways, 172 in biosynthesis of secondary metabolites pathways, 24 in the propanoate metabolism pathway, 26 in the starch and sucrose metabolism pathway, nine in the pentose phosphate pathway. In the S_120h vs S_0h_ALL comparison, 148 DEGs were enriched in the carbon metabolism pathway, 120 in the biosynthesis of amino acids, 82 in the glycolysis/gluconeogenesis pathway, 48 in carbon fixation in photosynthetic organisms pathways, 52 in the pyruvate metabolism pathway, and 58 in the starch and sucrose metabolism pathway, and significant changes were observed for these pathways. Figure 3 also shows several other pathways with different enrichment factors and Q values, indicating varying degrees of variation in these pathways under the experimental conditions.
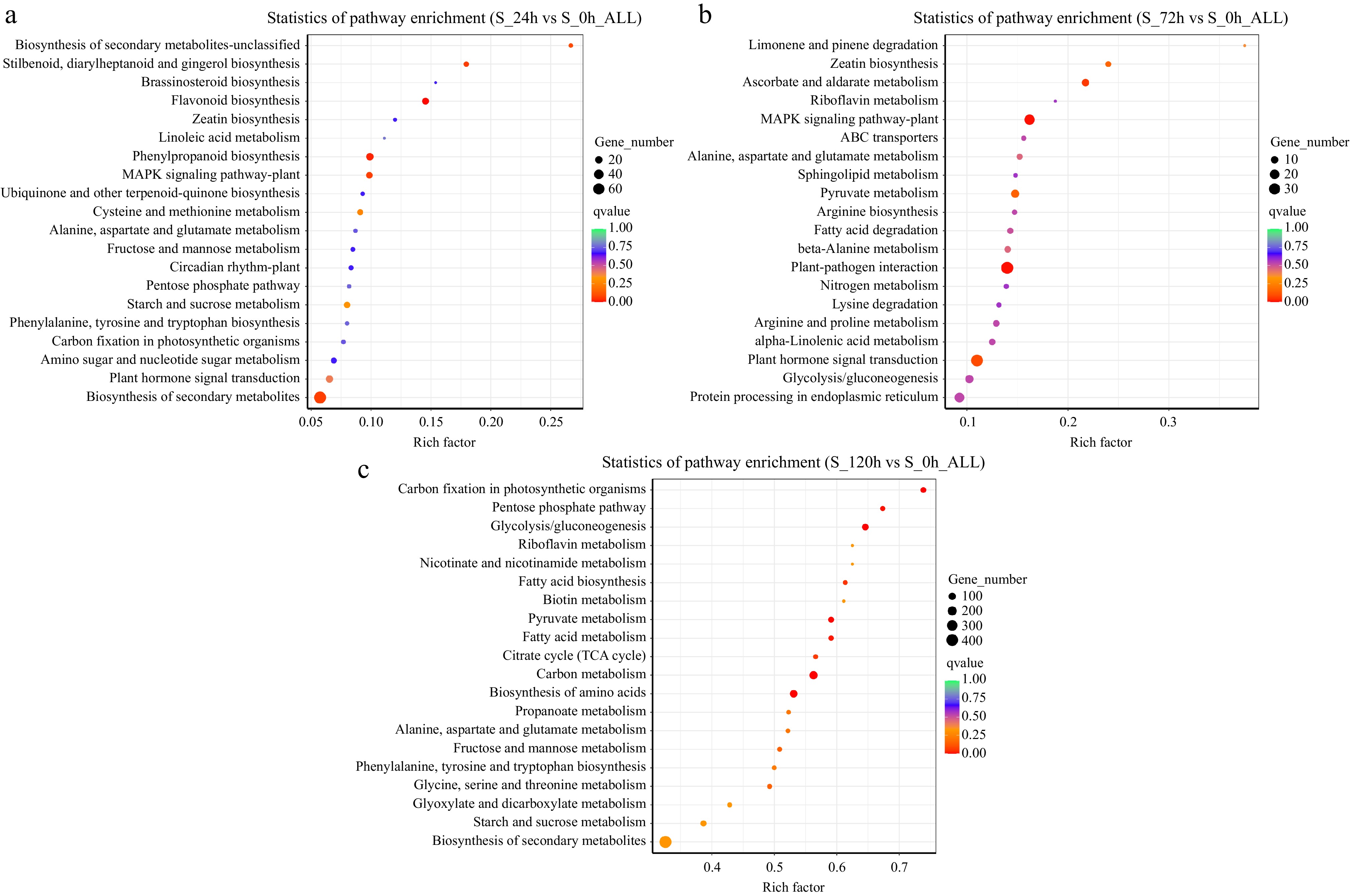
Figure 3.
(a)–(c) Analysis of enriched metabolic pathways in three groups. The top 20 most significant metabolic pathways (based on the p-values) are depicted, with each bubble representing a metabolic pathway. The size of the bubble represents the amount of metabolites in the pathway, as assessed through topological analysis. The color of the bubble represents the p-value of the enrichment analysis (i.e., −log10 p-value); the darker the color, the smaller the p value and the more significant the degree of enrichment.
KEGG analysis showed that the metabolic pathways 'flavonoid biosynthesis', 'phenylpropane biosynthesis', 'stilbene compounds', 'diarylheptane compounds and gingerol biosynthesis', 'MAPK signaling pathway', 'plant hormone signal transduction', 'Zeatin biosynthesis', and 'plant-pathogen interaction' were highly enriched. Compared with that in the CK group, the expression levels of most of the genes in these pathways in the SHAM120h group were upregulated after infection with Botrytis cinerea (Fig. 4).
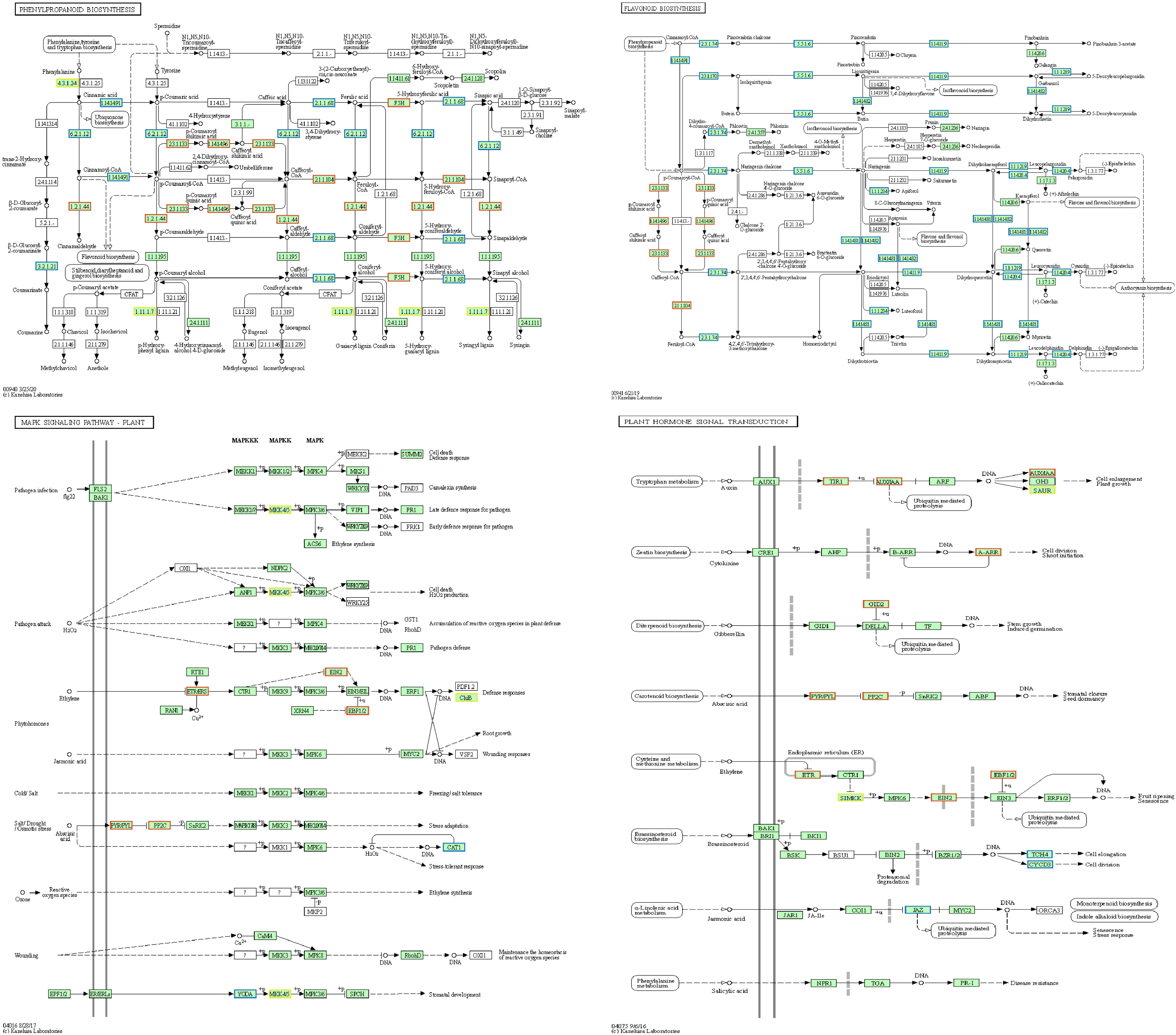
Figure 4.
Diagram showing the important KEGG pathways.
qRT-PCR validation of transcriptome data
-
For qRT-PCR validation of the transcriptome data, nine DEGs enriched for plant disease resistance pathways were randomly selected. The trend of qRT-PCR results was consistent with the transcriptome sequencing results, confirming the validity of the latter (Fig. 5).
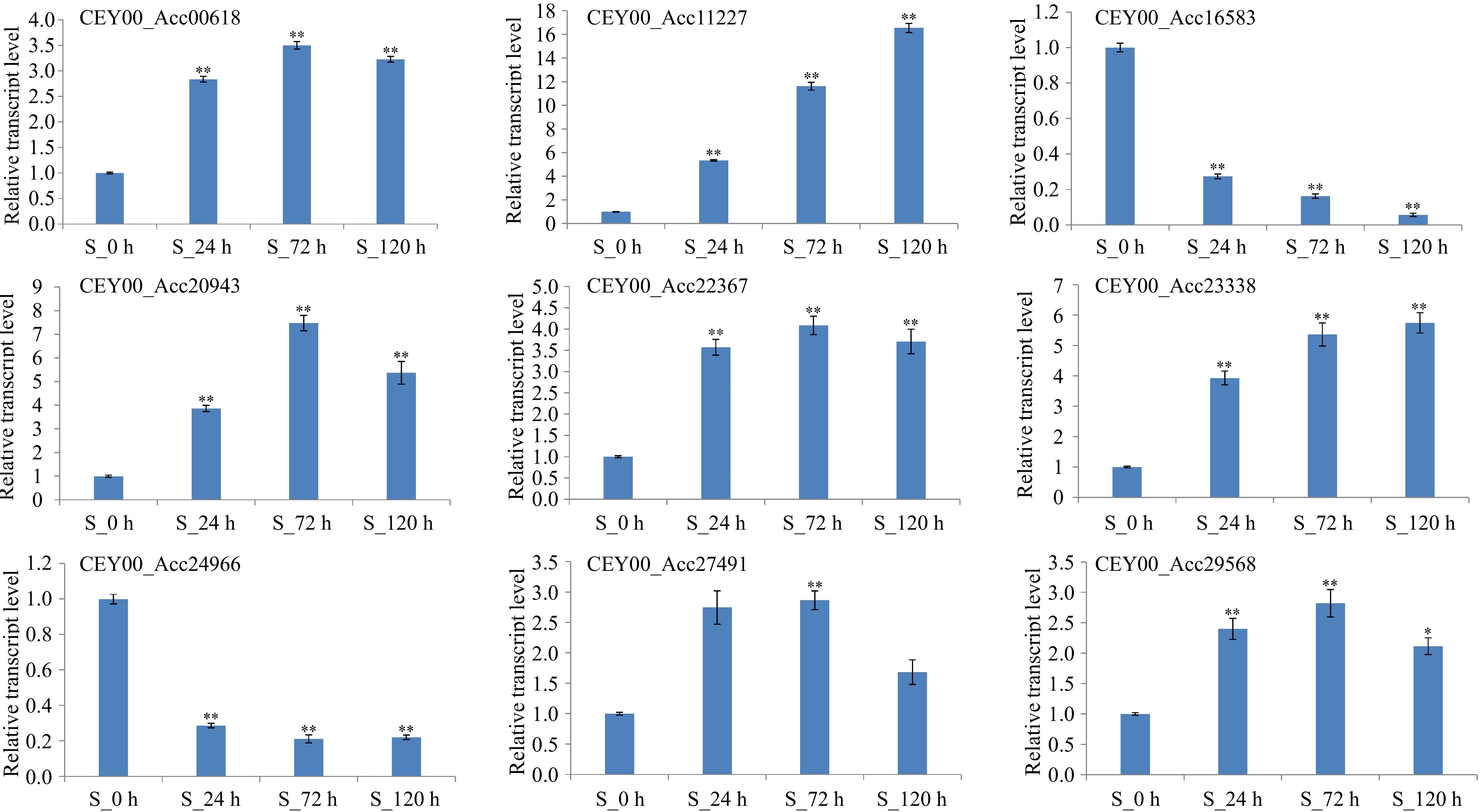
Figure 5.
qRT-PCR analysis of nine genes enriched for plant disease resistance pathways. CEY00_Acc00618: EIN3-binding F-box protein; CEY00_Acc11227: Caffeoyl-CoA O-methyltransferase; CEY00_Acc16583: Trans-cinnamate 4-monooxygenase; CEY00_Acc20943: Phenylalanine ammonia-lyase; CEY00_Acc22367: Mitogen-activated protein kinase; CEY00_Acc23338: Cytokinin dehydrogenase; CEY00_Acc24966: Chalcone synthase; CEY00_Acc27491: Ethylene receptor like; CEY00_Acc29568: Shikimate O-hydroxycinnamoyltransferase. '*' represents significantly different (p < 0.05), '**' represents extremely significantly different (p < 0.01).
-
In this study, SHAM treatment of kiwifruit fruit was used to decipher the mechanism of its action in plant disease resistance. Several genes related to key metabolic pathways were found to be differentially regulated by SHAM. These genes can provide a new strategy and scientific basis for the prevention and control of kiwifruit Botrytis cinerea. Flavonoid biosynthesis[19] is the process of conversion of simple precursor molecules, such as the aromatic amino acid phenylalanine, into flavonoids via a series of enzyme-catalyzed reactions. Flavonoids play important roles in plant physiology and ecology and are key regulators of plant growth and development, adaptation to the environment, resistance to external pressure, and interaction with other organisms. The MAPK signaling pathway-plant[20] is an important signal transduction pathway in plants that is involved in the regulation of plant responses to environmental stress, growth, development, interaction, and other biological processes[21], and is an important mechanism in the adaptation of plants to environmental changes and in their survival. Cys and methionine[22] play important roles through their involvement in various biological processes, including protein synthesis, antioxidant reactions, sulfur metabolism, and methylation and are crucial for maintaining life activities and normal cell functions. The starch and sucrose metabolic pathway[23] not only provides the energy and carbon necessary for plant growth but is also an important food source for humans and animals. The regulation and balance of starch and sucrose metabolic pathways are essential for maintaining normal energy metabolism, growth, and development of organisms. Amino sugar and nucleotide sugar metabolism regulates the formation of cellular structures and cell functions and participates in several important biological processes, such as signal transduction and energy metabolism. Normal metabolism and balance are essential for the proper growth, development, and functional maintenance of cells and organisms. Plant hormone signal transduction is involved in the regulation of physiological processes in plants[2] and transmission of hormonal signals to the interior of cells, affecting gene expression and metabolic pathways for controlling growth, development, and physiological status. This process involves the synthesis, transport, perception, and signaling of hormones. The pentose phosphate pathway plays important biological roles in cells, including NADPH production, nucleic acid synthesis, and regulation of glucose metabolism, and is critical for normal cellular functions and metabolic regulation. Carbon fixation in photosynthetic organisms[24] refers to the process of converting inorganic carbon (such as carbon dioxide) into organic compounds through photosynthesis. This process occurs in plants, algae, and some bacteria, and produces organic matter, including glucose, which provides energy and is a source of carbon for living organisms. Carbon metabolism is the process of synthesis, degradation, and transformation of carbon compounds in organisms[25] that plays an important role in energy generation, organic synthesis, establishment of cell structures, maintenance of carbon balance, and environmental adaptation. It is an important basis for biological activities. Biosynthesis of amino acids[26] refers to the biochemical processes of converting simple precursor molecules, such as glycine and pyruvate, into amino acids through a series of enzyme-catalyzed reactions. Amino acid biosynthesis plays important roles in protein synthesis, growth and development, metabolic regulation, stress resistance, and signal transduction and is required for vital activities in organisms. Propanoate metabolism has important physiological and biochemical functions in organisms, including energy production, carbon supply, metabolic regulation, and organic degradation, which are crucial for cell survival and functional maintenance. The biosynthesis of amino acids is essential for cell survival, growth, and function. It not only forms the building blocks of proteins but is also involved in physiological processes, such as regulation of metabolic balance, synthesis of other biomolecules, and provision of energy and carbon sources. Glycolysis and gluconeogenesis are important carbohydrate metabolic pathways responsible for the breakdown and synthesis of glucose, respectively, for maintaining the stability of cellular energy supply and blood sugar levels. The two pathways interact with each other and together maintain the metabolic balance and physiological functions in the body. Carbon fixation is an important metabolic process in photosynthetic organisms. It supports organic synthesis, cell growth, and division and plays important roles in oxygen production and carbon dioxide reduction, which are crucial for maintaining the stability of the ecosystem and regulating the climate. Pyruvate metabolism is an important metabolic pathway in various organisms. It not only participates in energy production and organic synthesis, but also regulates the redox balance and other metabolic pathways, playing important roles in maintaining cell function and survival.
To conclude, in this study, the role of SHAM, an inhibitor of the JA synthesis pathway, was explored in plant disease resistance, and the potential mechanisms of its action. It was found that to a certain extent, an appropriate concentration of SHAM can reduce the resistance of kiwifruit to Botrytis cinerea. To further understand the molecular mechanism behind this phenomenon, transcriptome sequencing analysis was conducted of fruit materials collected at 0, 24, 48, and 96 h of infection with Botrytis cinerea at the junction of disease and health. Using strict statistical screening criteria (FDR < 0.05 and |log2FC| > 1), 714, 1,511, and 4,181 DEGs were identified at the different infection stages. Most of these DEGs were upregulated in SHAM-treated kiwifruit. KEGG pathway enrichment analysis showed that these genes were mainly enriched for metabolic pathways closely related to plant disease resistance, including 'flavonoid biosynthesis', 'Phenylpropane biosynthesis', 'MAPK signaling pathway', and 'plant hormone signal transduction'. These findings not only provide a new perspective for understanding the role of SHAM in enhancing the resistance of kiwifruit to Botrytis cinerea, but also provide a scientific basis for devising strategies to control Botrytis cinerea in kiwifruit and have practical applicability.
-
The authors confirm contribution to the paper as follows: study conceptualization: Li Z; methodology: Yang J, Ma Y, Li Z; investigation: Yang J, Ma Y; formal analysis: Yang J, Ma Y, Li Z, Zhang H, Wang X, Zeng T; writing – original draft: Yang J; writing – review & editing, supervision, project administration, funding acquisition: Li Z. All authors reviewed the results and approved the final version of the manuscript.
-
The raw RNA-seq data were deposited in the NCBI SRA database (Accession number: PRJNA1104815).
This work was supported by the Natural Science Foundation of Chongqing (CSTB2023NSCQ-MSX0066), and the National Natural Science Foundation of China, China (32001351).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang J, Ma Y, Li Z, Zhong H, Wang X, et al. 2024. Mechanistic elucidation of the role of Salicylhydroxamic acid methyl ester in resistance of kiwifruit to Botrytis cinerea. Technology in Horticulture 4: e026 doi: 10.48130/tihort-0024-0023 

Mechanistic elucidation of the role of Salicylhydroxamic acid methyl ester in resistance of kiwifruit to Botrytis cinerea
- Received: 20 July 2024
- Revised: 05 September 2024
- Accepted: 19 September 2024
- Published online: 04 November 2024
Abstract: Kiwifruit (Actinidia chinensis) is a high-economic-value fruit crop cultivated worldwide. Botrytis cinerea infection, which causes Botrytis cinerea of kiwifruit after harvest, poses serious challenges to its production and storage. Salicylhydroxamic acid methyl ester (SHAM), an inhibitor of jasmonic acid synthesis, is involved in disease resistance in plants; however, its mechanism of action remains unclear. In this study, kiwifruit were infected with Botrytis cinerea after treatment with different concentrations of SHAM for 24 h. Treatment with an appropriate concentration of SHAM reduced the resistance of kiwifruit to Botrytis cinerea. After 0, 24, 48, and 120 h of Botrytis cinerea infection, fruit materials at the junction of disease and health were collected and analyzed using transcriptome sequencing. Differentially expressed genes with an FDR < 0.05 and |log2FC| > 1 were identified. Compared with the 0 h infection control group, 714, 1,511, and 4,181 differentially expressed genes were identified at 24, 48, and 96 h of infection, and most of the differentially expressed genes were regulated in the SHAM treatment group fruits. KEGG pathway analysis revealed that the related genes were mainly enriched for 'flavonoid biosynthesis', 'Phenylpropane biosynthesis', 'Mitogen-Activated Protein Kinase signal pathway', 'plant hormone signal transduction', and other metabolic pathways. These results unravel the molecular mechanism underlying the effect of SHAM in the resistance to Botrytis cinerea and provide an important basis for safe and efficient control of Botrytis cinerea in kiwifruit production.
-
Key words:
- Kiwi fruit /
- Botrytis cinerea /
- SHAM /
- Molecular mechanism /
- Resistances