










-
Soil seed banks are a crucial component of forest ecosystems, directly influencing ecosystem structure and function, as well as the assembly and succession of forest communities[1]. It remains unclear whether there are significant linear differences in forest soil seed bank abundance along geographical scales[2]. Additionally, forest community assembly patterns differ between different forest origins (planted forests vs natural forests), and it is uncertain whether these origin differences affect soil seed bank density (SSBD)[3]. Therefore, investigating the distribution patterns and key factors influencing soil seed density between planted and natural forest ecosystems at a macroscale is of great significance for sustainable forest management.
Natural forests are characterized by a series of successional stages of plant communities that develop on primary or secondary bare land[4]. Dominated by native species, these forests can regenerate naturally, boasting complex ecosystems and high biodiversity[5]. In contrast, planted forests are predominantly created through artificial sowing, cultivation, and management, exhibiting uniform age and simplified structure due to human intervention[6]. Compared to natural forests, planted forests generally exhibit lower biodiversity and diminished ecosystem functions[5]. Within both natural and planted forest ecosystems, soil seed banks play a crucial role in maintaining population size and diversity through temporal storage effects[7]. Soil seed banks have the ability to restore degraded ecosystems and accelerate forest succession, so the renewal of natural and planted forests is largely dependent on soil seed banks[8]. Consequently, understanding the dynamics of soil seed banks is of paramount importance in forestry, as it provides valuable insights into the natural regeneration of forests, guiding future forest management practices.
Numerous studies have highlighted the pivotal role of climatic factors, notably temperature and precipitation, in regulating the growth of both planted and natural forests, as well as in resource allocation[3,9]. Likewise, a substantial body of research indicates that temperature and precipitation primarily drive the variances in soil seed banks at the macro scale[10−12]. Hence, climatic factors may exert significant impacts on the soil seed bank of both planted and natural forests. As temperatures rise and rainfall increases, soil seed density significantly decreases[13]. This is mainly because, with higher temperatures and increased precipitation, trees adopt a strategy of rapid investment-reward in resource utilization, leading to faster growth, enhanced inter/intra-species resource competition, increased investment in resource competition, and reduced investment in reproduction. This ultimately results in lower levels of soil seed density[14]. Daylight duration also significantly affects soil seed density, as different daylight durations can alter plant growth cycles, influencing flowering and fruiting times, thus impacting seed production and density[15]. Furthermore, changes in light conditions can affect competitive relationships among plant species, with some plants being more adapted to longer periods of sunlight, while others may have a competitive advantage under shorter light conditions[16]. Such differences can influence species survival and reproduction, subsequently affecting the density of seeds in the soil.
Soil, as the direct living environment for trees, plays a crucial role in their growth, development, and reproduction[17,18]. Research has also found that the resource allocation strategies of both planted and natural forests are significantly constrained by soil nutrients[3]. Therefore, soil nutrients may represent another type of abiotic factor that limits the soil seed bank of forests. Under conditions of ample nutrients, plants may produce more seeds, increasing seed density[19]. Additionally, soil nutrients influence seed viability and germination capacity[20]. The nutritional status of the soil can also affect soil seed density by influencing competitive relationships among plant species[21]. In a nutrient-rich environment, competitively dominant species may prevail, whereas, in a nutrient-poor environment, species with strong adaptability may have a better chance of survival. These differing competitive pressures can impact the density and abundance of species in the seed bank[22]. Soil nutrients also influence the activity of soil microorganisms, which, in turn, affect the physical and chemical properties of the soil, subsequently impacting seed survival and germination[23].
In different developmental stages of forests, trees exhibit various reproductive strategies, which may consequently have an impact on the soil seed bank. Many studies found that forest stand characteristics, such as stand age, mean diameter at breast height, key leaf traits, and forest productivity can influence SSBD[24−26]. With increasing stand age, the intensity of interspecific competition can change, and the microenvironment within the forest, including factors like light, humidity, and soil structure, can undergo alterations, subsequently affecting the soil seed bank[27]. In recent years, numerous studies have highlighted the critical role of key leaf traits in explaining various ecological phenomena. Species with higher specific leaf area (SLA) and lower leaf dry matter content (LDMC) tend to adopt a fast investment-reward resource utilization strategy, allocating more resources to interspecific competition and reducing investment in reproduction[18]. This leads to lower levels of soil seed bank density. Similarly, some research has found that forests with higher productivity typically have greater biomass, resulting in more seed production and increased seed bank density[28]. However, it should be noted that forests with higher productivity may also experience more intense interspecific competition, which can lead to lower soil seed density[27].
In this study, based on SSBD data collected from 537 natural forests and 383 planted forests within China through field surveys and literature sources, the aim is to investigate the differences in SSBD between plantation and natural forests at the macroscale and the key factors driving these differences. To address these questions, the following hypotheses are made: (1) SSBD in planted forests will significantly exceed that in natural forests; (2) Climatic factors will be the primary drivers of the macro-scale differences in SSBD between planted and natural forests; (3) Climatic factors will influence SSBD in planted and natural forests by adjusting soil nutrients and stand characteristics.
-
The density data of the soil seed bank were collected partly from literature searches and partly from field measurements. The specific data are listed in Supplementary Table S1. Relevant peer-reviewed journal articles published between 2005 and 2020 were searched in Web of Science, Google Scholar, and CNKI. The keyword combinations used in the search were 'forest' and 'soil seed bank'. A total of 108 relevant papers containing 623 data points were retrieved. The data in the literature was then screened using the following criteria: (1) The latitude and longitude of the plots should be provided by the study, and the plots should be categorized as either natural or planted forests; (2) The study should provide or allow the calculation of the mean, standard deviation, or standard error of soil seed bank density data in the sample plot; (3) The study should present the results of field studies rather than retrospective or simulation studies; and (4) The sampling period should be outside of peak germination seasons to minimize seasonal effects on soil seed bank density estimates. For the articles meeting the criteria, the index of soil seed density in the 0–10 cm soil surface layer was extracted. If a study has multiple sampling depths from 0 to 10 cm at the same site, these observations were treated as independent samples. In these articles, as much information as possible was collected on tree species, stand age, stand density, tree DBH, and other stand characteristics of each sample plot.
Twenty seven sites in the field were collected and data measured from 297 forest plots. The latitude, longitude, elevation, and slope of each site were recorded for comprehensive analysis, and the site location, tree species, stand age, tree DBH, and stand density documented in real time. At each site, at least four 20 m × 20 m forest plots with typical zonal vegetation were selected. For sampling, the same method was used as described in the literature to measure soil seed bank density: after removing litter from the surface of each sample plot, five soil samples, each with dimensions of 10 cm × 10 cm × 10 cm, were randomly collected. The litter layer was removed to focus on the persistent soil seed bank in the 0–10 cm soil layer, minimizing the effects of short-term seed input and ensuring consistency across sites. The samples were thoroughly mixed and then placed in soil bags, which were sieved to remove debris upon return to the laboratory. The samples were stored in a dry, dark environment until germination experiments began in May of the following year. In May, the labeled soil was evenly spread in germination trays to a depth of about 5 cm. Iron arches were set up over the trays and covered with film to prevent external seeds from entering. The temperature inside the enclosure was maintained at 25–30 °C, with natural light and a humidity level of around 70%. Water was applied every 3–5 d to keep the soil moist. The germination and growth of the seeds were observed and recorded. From the onset of sprouting, daily records were kept of the number of seedlings and their morphological characteristics. Finally, the remaining seeds in the germination trays were checked for germination. The remaining ungerminated seeds were tested for viability using the tetrazolium chloride (TTC) staining method, with seeds soaked in a 1% TTC solution at 30 °C for 24 h. Seeds that displayed a reddish color in their embryos were considered viable[29]. For seeds that did not show a clear TTC staining result, manual examination was performed by cutting to check for intact embryos. The seed bank germination experiment lasted from May to November of the following year. The number of seedlings for all species recorded during the experiment was used to calculate the seed bank density, expressed as the number of seedlings per unit area, for further analysis.
Climate data
-
Climate data, including Mean Annual Temperature (MAT), Mean Annual Precipitation (MAP), Annual Sunlight Duration (ASD), and Mean Annual Evaporation (MAE), were obtained from WorldClim (
https://worldclim.org ) at a 1 km spatial resolution.Soil nutrient data
-
Data for total nitrogen content and total phosphorus content of 0−20 cm soil at a 1 km resolution was extracted from the Harmonized World Soils Database version 2.0 (
https://gaez.fao.org/pages/hwsd ).Forest stand factors
-
Forest stand factors include forest age, forest density, and forest mean diameter at breast height (DBH). Forest age is mainly obtained from the literature reviewed. For literature without forest age information, local forestry bureau and ecological station websites were referred to, as well as consultation with specific personnel in charge. The forest DBH represents the average DBH of all trees (DBH > 5 cm) within the plot. For species identification, local flora references were relied on, and for species that were difficult to classify, the WFO Plant List (
https://wfoplantlist.org ) was used to confirm taxonomic status. For each sample plot, five dominant trees were randomly selected based on their relative dominance (e.g., height and canopy spread) to represent the primary structural characteristics of the stand. The selected trees could either be from the same species or different species, depending on the composition of the plot. To minimize sampling bias, trees with abnormal growth patterns were excluded. Stand-level measurements, such as forest density, were calculated as the number of individual trees per unit area, and species diversity was determined based on the identified species in each plot. Altitude, slope, aspect, and other stand factors of the actual survey plots were measured using handheld GPS devices. Due to the limited number of actual field survey plots, stand factors such as altitude were not considered in the subsequent calculations.Plant functional traits
-
In this study, five plant functional traits were selected to represent diverse strategies of plant resource utilization: leaf area (LA), specific leaf area (SLA), leaf dry matter content (LDMC), leaf nitrogen content (LN), and leaf phosphorus content (LP). The data on plant functional traits of regional tree species collected in the literature were obtained from the TRY database[30]. During field measurements, five dominant trees were randomly selected from each sample plot, ensuring they were situated away from the plot edges. Leaves were collected from various directions at the same height in the middle of the canopy of each selected tree. Twenty leaves of similar maturity, free from diseases and pests, were gathered and stored in ziplock bags for transport to the laboratory. Upon arrival at the laboratory, the leaf area (LA) was measured using a portable laser planimeter (CI-202, Walz, Camma, USA)[31]. Subsequently, the leaves were submerged in water and placed in a dark environment at a constant temperature of 4 °C for 12 h. Once the surface water was absorbed, the saturated fresh weight of the leaves was measured using an electronic balance. The leaves were then placed in an oven at 120 °C for 30 min, followed by drying at 80 °C for 24 h, and the dry weight of the leaves was recorded. Leaf nitrogen content was determined using the Kjeldahl method, while leaf phosphorus content was measured using the Mo-Sb colorimetry method[32]. Specific leaf area (SLA) and leaf dry matter content (LDMC) were calculated using the following formulas: Specific leaf area (SLA) = Leaf area/Leaf dry weight; Leaf dry matter content (LDMC) = Leaf dry weight/Leaf saturated fresh weight. While acknowledging potential differences in plant functional traits among species, this study focused on exploring these traits at the community scale. Therefore, the community-weighted mean value (CWM) was utilized to represent the average trait value of each plot.
$ \text{CWM}=\sum _{\text{i=1}}^{\text{S}}{\text{D}}_{\text{i}}\times\text{Trai}{\text{t}}_{\text{i}} $ (1) where, CWM denotes community-weighted functional trait values, Di is the abundance of dominant species, and Traiti is the specific functional trait[33].
Forest net primary productivity data (NPP)
-
China's MOD17A3H vegetation net primary productivity (NPP) data was obtained from the NASA website (
https://search.earthdata.nasa.gov/search ), with a spatial scale of 500 m and a time scale of years. The NPP estimates were generated using the Carnegie-Ames-Stanford Approach (CASA) model, employing the following calculation method:$ \rm NPP(x,t)=APAR(x,t)\times \varepsilon (x,t)$ (2) where, APAR(x,t) represents the photosynthetically active radiation (PAR) absorbed at the x pixel in the t-th month, with units in MJ/m². ε(x,t) represents the actual light use efficiency at the x pixel in the t-th month, measured in g·C/MJ[34].
Analysis
-
Initially, the soil seed bank density data was transformed logarithmically to normalize it, and all subsequent analyses were performed using these log-transformed data. Before analysis, all variables were standardized for a comparable scale in interpreting parameter estimates.
All data analyses were conducted using R (version 4.2.2,
www.R-project.org ). The 'ggsignif' package was used to test the difference in soil seed bank density between natural forests and planted forests at the 0.05 significance level. To reduce collinearity among multiple plant functional traits, the 'pcaMethods' package was employed for PCA analysis of plant functional traits and the first two principal components, PC1 and PC2 were extracted[35]. The general linear regression model in the 'lme4' package was utilized to analyze the effects of climate, soil, and plant factors on soil seed bank density in plantations and natural forests, with R² used to evaluate model fitting[36]. To visualize the relationship between various factors and soil seed bank density, a correlation heat map was created using the 'linkET' package[37].A multiple linear regression model was constructed, based on modified Akaike information criteria (AICc; ΔAICc < 2) selection procedure to select the best predictors of soil seed bank density. The 'dredge' function in the MuMIn package was used to create all possible subset models, ranking them based on their AICc values (AIC value corrected for sample size), and selecting the model with the lowest AIC value as the optimal model[38]. The contributions of various factors in the optimal model to SSBD were recorded. Variance decomposition was then performed using the rdacca.hp function, assessing the variance contributions of climatic, soil, and plant factors to the optimal model, expressed as percentages[39].
Structural equation models (SEM) can be used to evaluate complex causality between variables by translating hypothetical causality into the expected statistical relationship pattern in the data[40]. To study the direct and indirect effects of each factor on soil seed bank density, a structural equation model was constructed using the 'piecewiseSEM' software package. The SEM model was fitted using the psem function in the 'piecewiseSEM' package, based on generalized least squares, with the optimal model having the smallest AIC score and a Chi-Square p-value greater than 0.05[41].
-
The soil seed bank density (SSBD) of planted forests and natural forests exhibited significant geographical differences. The average SSBD of natural forests was 2.876 m−2, ranging from 1.395 to 4.049 m−2. In contrast, the average SSBD for planted forests was 3.137 m−2, ranging from 1.536 to 3.858 m−2. The difference of SSBD between natural forest and planted forest is very significant, and the SSBD value of planted forest is generally higher than that of natural forest (Fig. 1b).
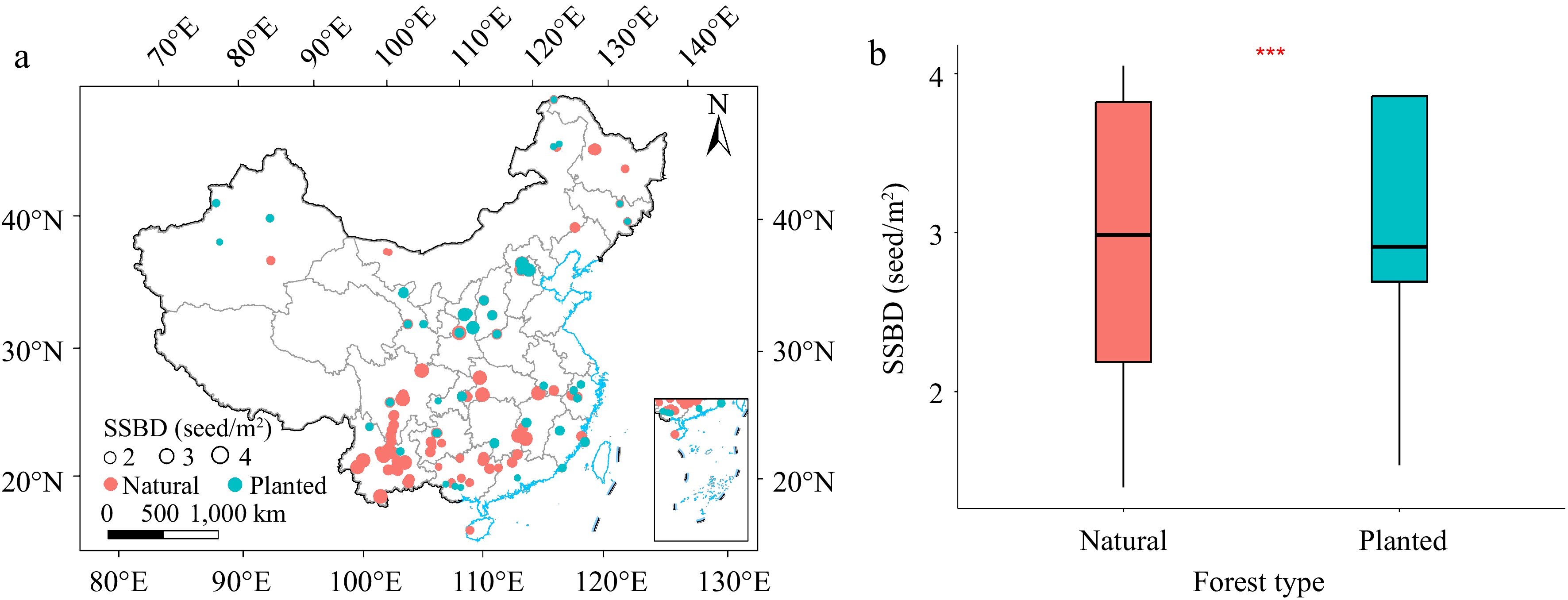
Figure 1.
Spatial distribution of soil seed banks and plot locations in planted and natural forests. (a) Comparison of SSBD between natural forests and planted forests. (b) Significance of the differences was assessed using a t-test, with significance at the 0.001 level. *** p < 0.001.
Both planted and natural forests showed similar trends in SSBD in response to changes in climatic factors. SSBD decreased significantly with increasing temperature and precipitation (p < 0.001), while it increased with longer sunlight exposure and higher evaporation rates. Overall, natural forests exhibited greater climatic plasticity in SSBD (Fig. 2).
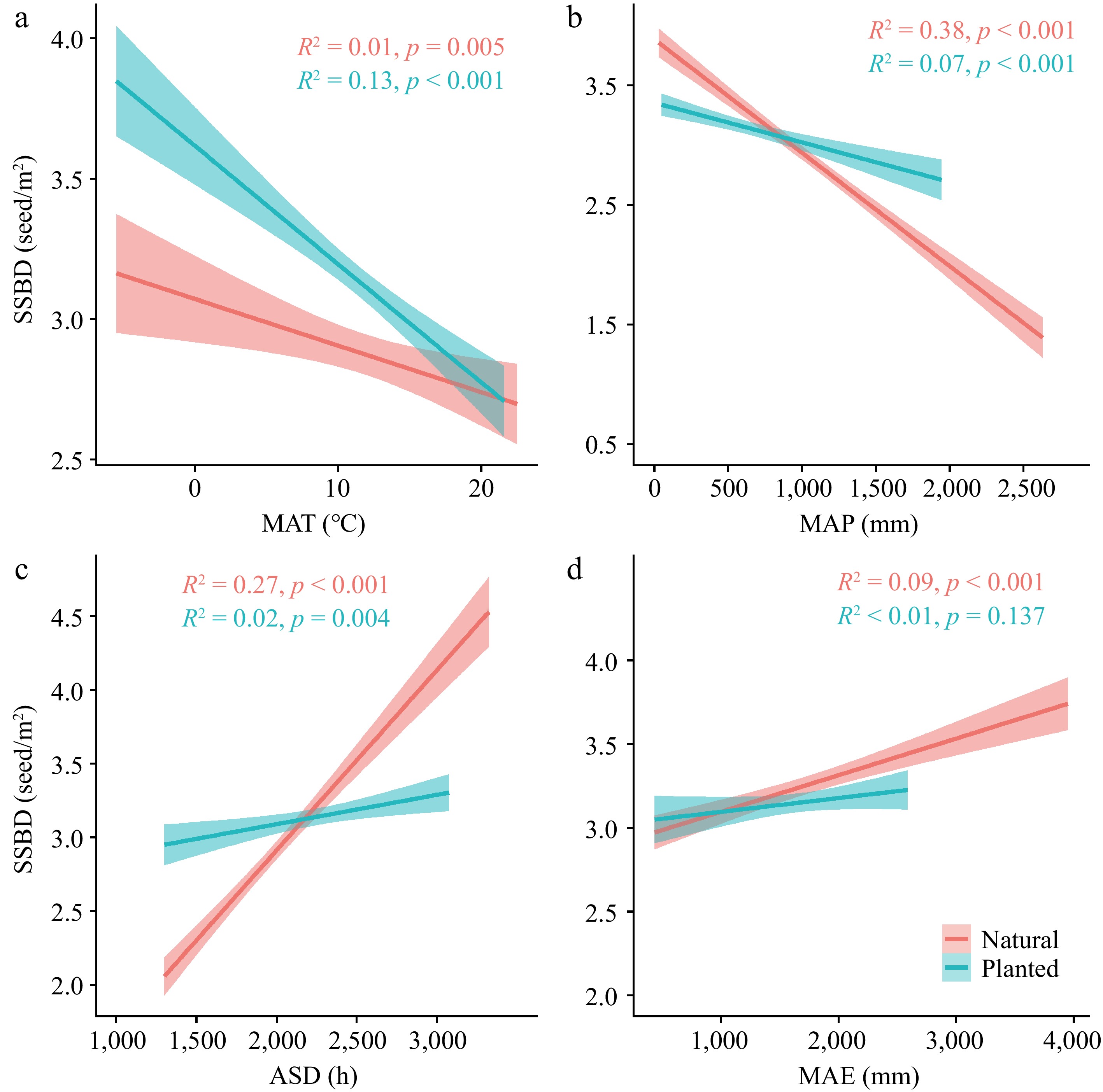
Figure 2.
The relationships between climatic factors and SSBD in natural forests and planted forests. R2 represents the goodness of fit, and P-values indicate significance. Climatic factors include: (a) Mean Annual Temperature (MAT); (b) Mean Annual Precipitation (MAP); (c) Annual Sunlight Duration (ASD); (d) Mean Annual Evaporation (MAE).
Compared to planted forests, SSBD in natural forests was more sensitive to changes in soil nutrients (higher R2). The SSBD of both forest types increased significantly with the increase of soil nitrogen content (Fig. 3a, b). With the increase of soil phosphorus content and soil pH, planted forest SSBD showed a significant decline trend (Fig. 3b, c).
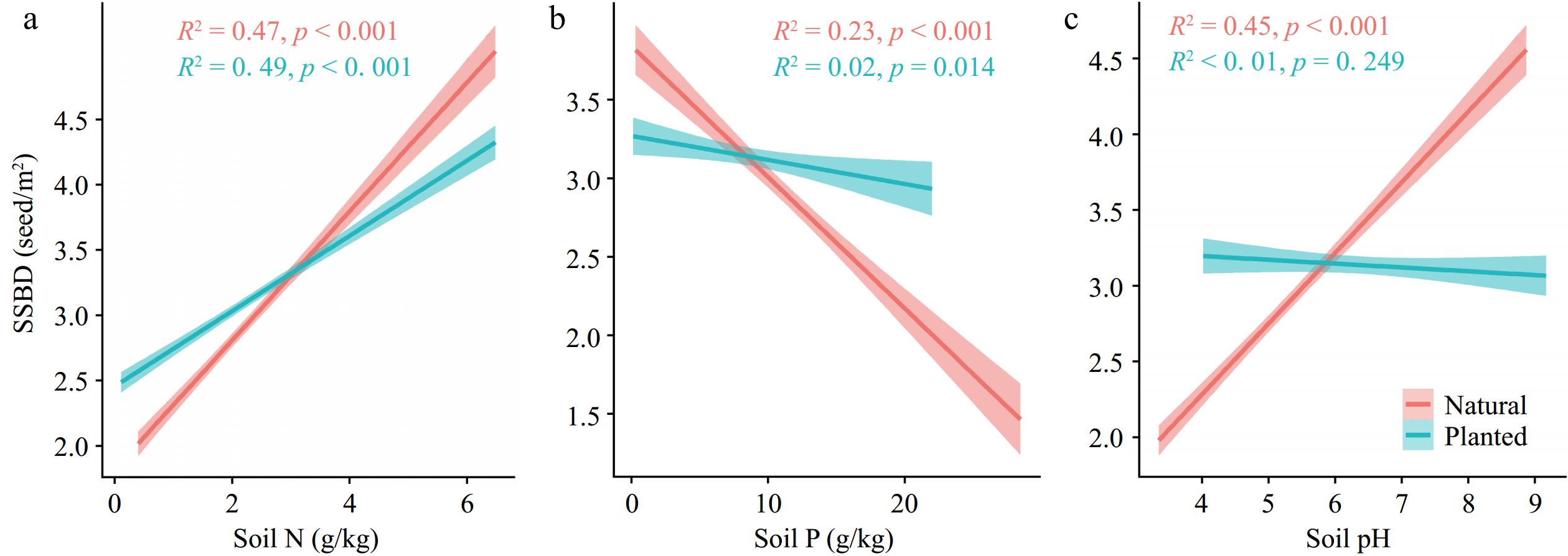
Figure 3.
The relationships between soil factors and SSBD in natural forests and planted forests. R2 represents the goodness of fit, and P-values indicate significance. Soil factors include: (a) Soil total nitrogen content (Soil N); (b) soil total phosphorus content (Soil P); (c) Soil pH.
SSBD in both forest types showed similar trends in response to changes in forest stand factors. SSBD in planted and natural forests positively correlated with forest age and forest DBH, but negatively correlated with stand density and leaf functional traits (Fig. 4). The impact of forest productivity on natural forest SSBD (R2 = 0.17) was greater than on planted forests (R2 < 0.01) (Fig. 4f). There was a general collinear correlation between potential influencing factors of SSBD in planted and natural forests (Fig. 5).
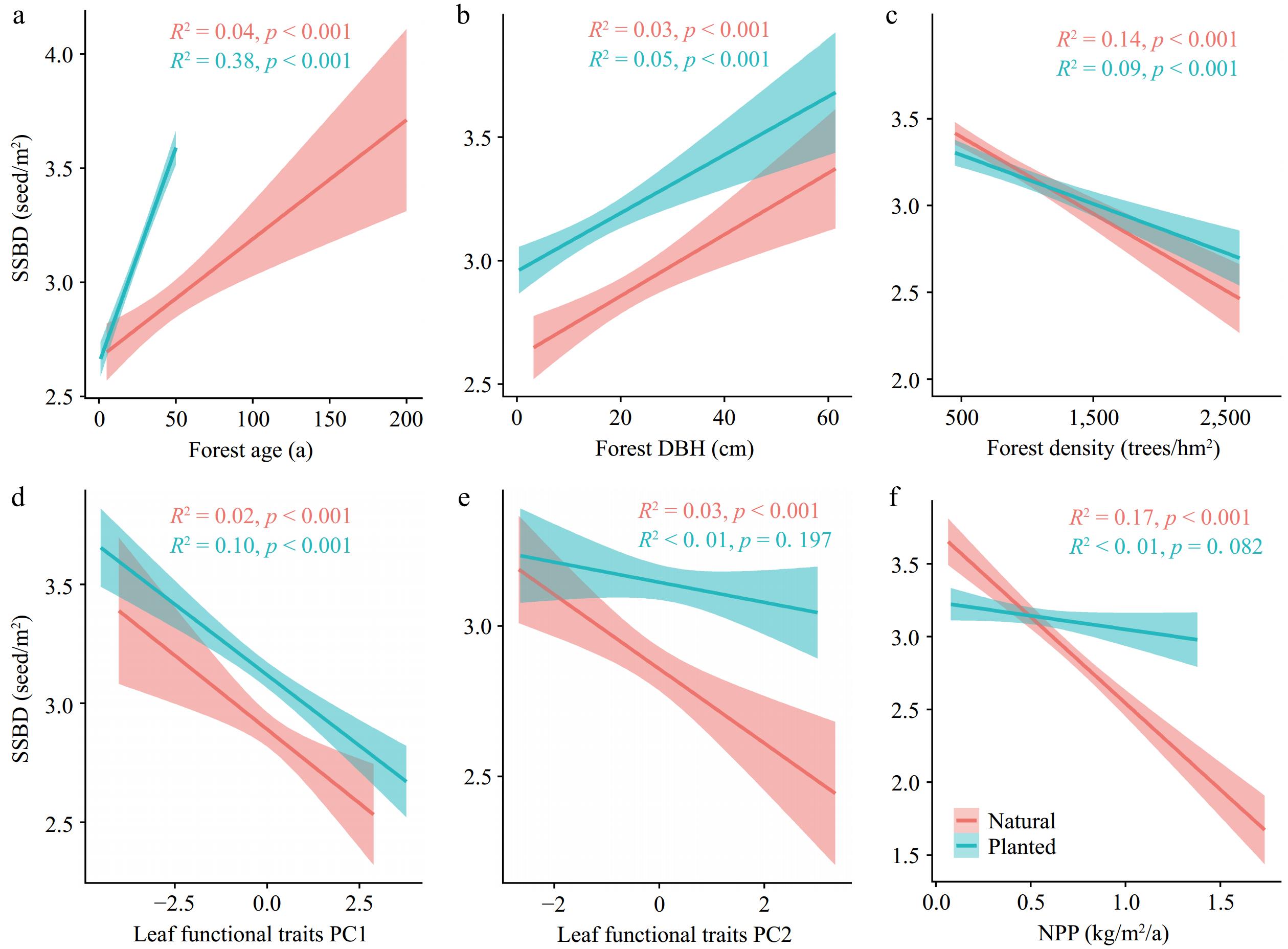
Figure 4.
The relationships between forest stand factors and SSBD in natural forest and planted forests. R2 represents the goodness of fit, and p-values indicate significance. Forest stand factors include: (a) Forest age; (b) Forest diameter at breast height (average DBH); (c) Forest density; (d) Leaf functional traits PC1; (e) Leaf functional traits PC2; (f) Net primary productivity (NPP).
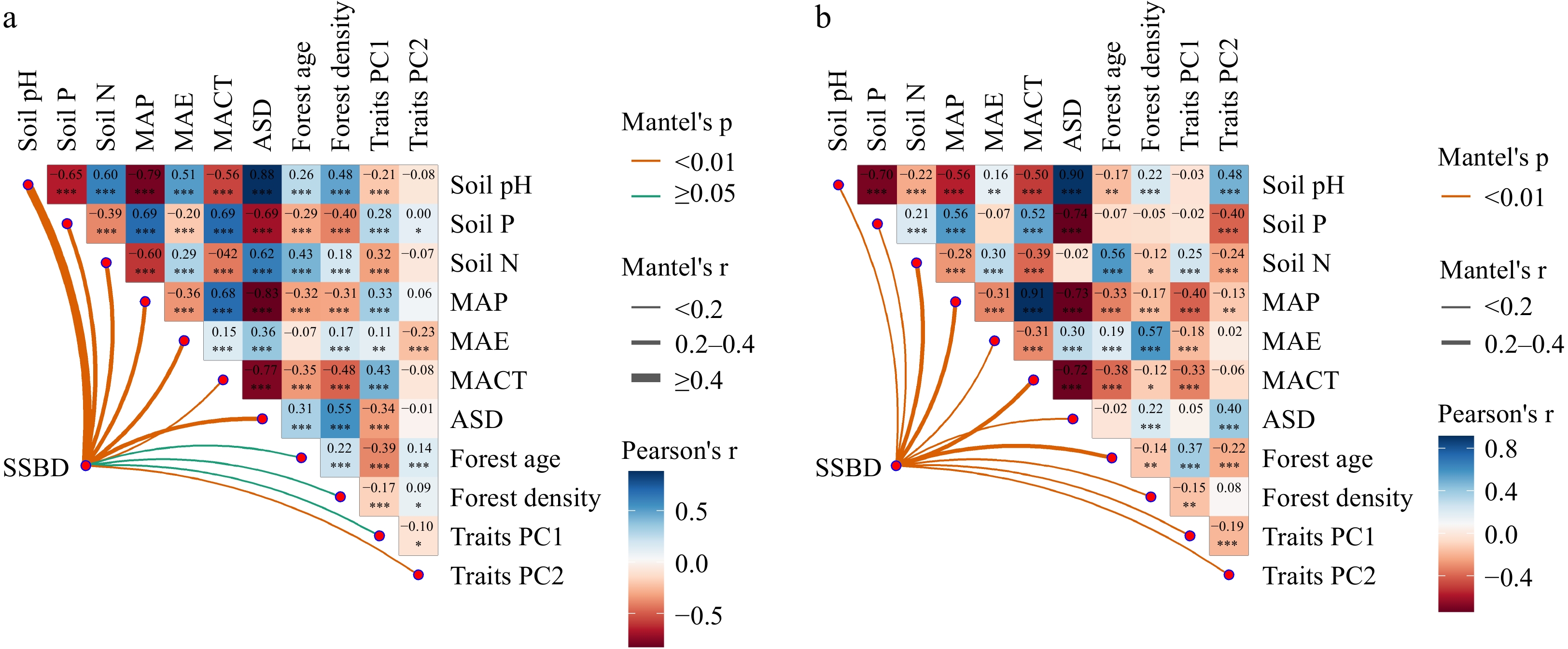
Figure 5.
Multivariate correlation analysis of potential influencing factors on SSBD in (a) natural forests, and (b) planted forests. MAP: Mean annual precipitation; MAE: Mean annual evaporation; MACT: Mean annual coldest month temperature; ASD: Annual sunlight duration; Soil N: Soil total nitrogen content; Soil P: Soil total phosphorus content; Soil pH: Soil pH; Traits PC1: Leaf functional traits PC1; Traits PC2: Leaf functional traits PC2.
All potential influencing factors explained 75.7% of the variance in SSBD for natural forests and 66.1% for planted forests (Fig. 6). Soil nutrient factors (R2 = 0.361; R2 = 0.377) had a stronger explanatory power for the spatial variability of SSBD in both forest types than climatic factors (R2 = 0.301; R2 = 0.073) and forest stand factors (R2 = 0.094; R2 = 0.211) (Fig. 6). Soil pH made the largest independent contribution to the spatial variability of SSBD in natural forests (Fig. 6a), while soil nitrogen content contributed most significantly to the spatial variability of SSBD in planted forests (Fig. 6b).
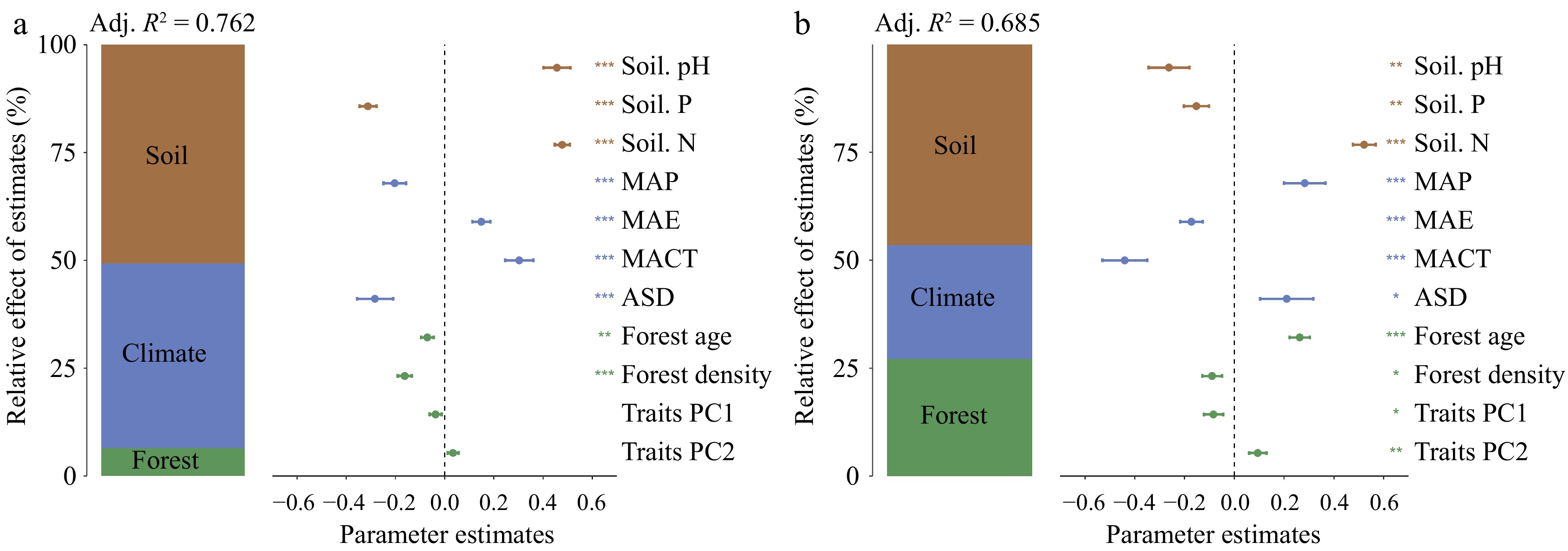
Figure 6.
Impact of potential factors on SSBD in (a) natural forests, and (b) planted forests. The figure presents the average parameter estimates (standardized regression coefficients), related 95% confidence intervals, and the relative importance of each factor, expressed as the percentage of explained variance. The adjusted R2 for the average model and the p-values for each predictive factor are denoted as follows: * p < 0.05; ** p < 0.01; *** p < 0.001.
Soil pH had the greatest direct impact on SSBD in natural forests. MAT and MAP not only directly affected SSBD in natural forests but also indirectly through effects on soil pH, stand density, and forest NPP, with the direct impacts being greater than the indirect ones (Fig. 7a). For planted forests, soil nitrogen content had the greatest direct impact on SSBD. MAE influenced SSBD in planted forests indirectly through its impact on soil nitrogen content, with its indirect effect being greater than the direct effect (Fig. 7b).
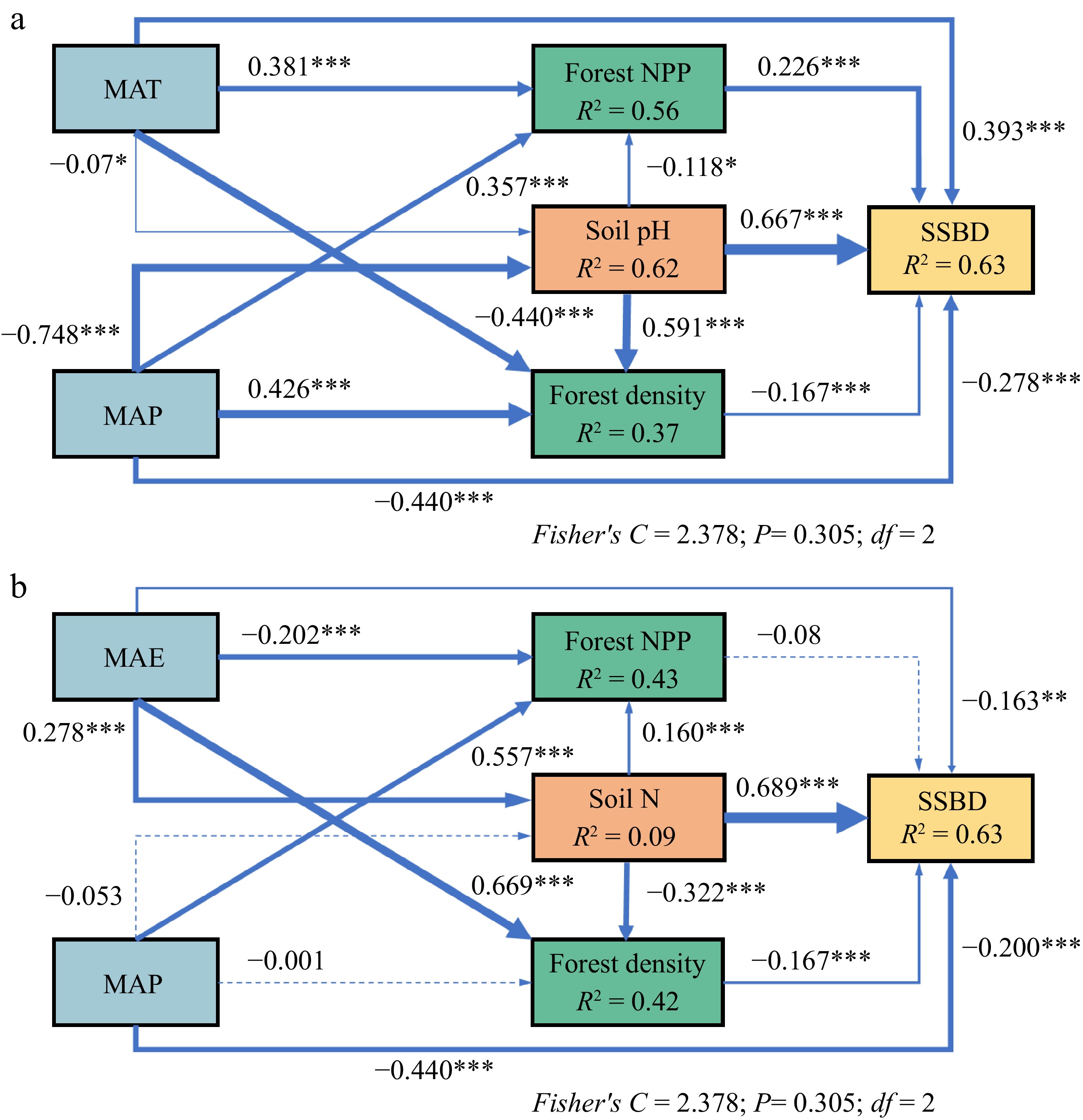
Figure 7.
Relationships between SSBD and climatic factors, soil nutrients, and forest stand factors in (a) natural forests, and (b) planted forests. The path diagrams represent the standardized results of the final Structural Equation Models (SEMs) testing relationships between variables. Numbers alongside the paths indicate the standardized SEM coefficients, and asterisks denote significance (*** p < 0.001; ** p < 0.01; * p < 0.05). R2 indicates the goodness of fit for the generalized additive models. The best SEMs were selected based on the lowest Akaike information criterion.
-
The results of this study show that the SSBD of planted forests is significantly higher than that of natural forests, confirming the first hypothesis. Planted forests, characterized by shorter planting periods and younger ages, tend to have higher SSBD compared to older, mature natural forests growing in their natural state[8,42]. In planted forests, the density and distribution of trees are often carefully planned to maximize land use efficiency and productivity[43]. Intensive planting will increase the coverage of vegetation, and after the soil surface is covered by vegetation, soil erosion and seed loss caused by erosion can be reduced, which is conducive to the accumulation and maintenance of seeds in the soil[44]. Moreover, tree species in planted forests are often selected for high yield or rapid growth, which may produce higher seed outputs, thereby increasing the density of the soil seed bank. Planted forests undergo regular cycles of harvesting and replanting. This periodic human intervention might lead to a regular renewal of seeds in the seed bank, thereby maintaining or increasing its density[45]. Compared to natural forests, planted forests generally harbor (or yield) a large number of light-demanding tree species with broad ecological niches[46]. These tree species often produce abundant seeds, and these seeds can persist in the soil for extended periods.
Numerous studies have shown that climatic factors significantly influence the SSBD in forests[13,47,48]. The experimental results indicate that the SSBD in both planted and natural forests decreases with rising Mean Annual Temperature (MAT). Research suggests that temperature is a key climatic factor affecting seed dormancy and stimulating germination[49]. Cold conditions slow down the metabolic rate of seed embryos and germination rates. Seeds that grow in colder regions tend to have higher longevity and survival rates compared to those in warmer regions[50]. As MAT increases, seed germination rates rise, while seed vitality and persistence decrease. Studies also show a positive correlation between temperature and the frequency of predator activities; higher MAT can increase the predation rate of germinated seeds in the soil[19]. Our results demonstrate that SSBD in planted forests is more sensitive to temperature changes than in natural forests (Fig. 2). This could be due to the forest climate formed in natural forests over time[51]. Natural forests have more developed ecosystems and a stronger resistance to environmental changes, making their SSA less sensitive to increases in MAT compared to planted forests[9]. Therefore, the response of natural forests to MAT rise in SSBD is less sensitive than that of plantation forests. Research shows that SSBD significantly decreases with increased precipitation, consistent with our findings[48]. Increased rainfall can break seed dormancy and stimulate germination. However, early germination is not conducive to seed growth; changes in rainfall affect the longevity of the seed bank, and increased Mean Annual Precipitation (MAP) directly impacts the risk dispersal mechanisms of seeds, potentially causing a decrease in SSBD[13,48]. The results show that SSBD in natural forests is more sensitive to MAP compared to planted forests, possibly because planted forests, due to artificial irrigation, have less water demand. In contrast, natural forests are often in a state of drought and water scarcity, making their soil seed banks more responsive to rainfall compared to those in planted forests[52]. In the present study, other climatic factors also affect the SSBD of planted and natural forests, but according to the results of the comprehensive structural equation model, MAT and MAP are the key climatic factors affecting SSBD in both planted and natural forests.
The experimental results of this study show that there is a close relationship between the SSBD in planted and natural forests and soil nitrogen content, phosphorus content, as well as soil pH. Being in wild and impoverished soils, natural forests are limited by soil nitrogen nutrients, while planted forests, under artificial cultivation, still require timely nitrogen fertilization to ensure normal tree growth[53]. Both planted and natural forests are limited by nitrogen in their soil environments. The development of forests in China is primarily limited by nitrogen elements[54]. Therefore, an increase in soil nitrogen content is conducive to the growth and development of germinating seeds in the soil[55]. Our results show that SSBD in both planted and natural forests is positively correlated with soil nitrogen content. Compared to the limitation of soil nitrogen content on planted and natural forests, the limitation of soil phosphorus content is not very strong. Chen et al. have shown that seed vigor in the soil seed bank is positively correlated with soil available P content, which also explains the experimental results of this study[56]. In planted and natural forests, seed vigor is positively correlated with soil total phosphorus content. Higher seed vigor in soil seeds changes their bet-hedging ability and risk dispersal strategies, increasing their risk of extinction. Therefore, SSBD tends to be lower in environments with higher soil phosphorus content[57]. Seed germination in acidic soils is limited, and as soil pH gradually changes from acidic to neutral, plant efficiency in utilizing soil nutrients increases[58]. In natural forests, an increase in soil pH improved the nutrient uptake efficiency of seeds in the soil and significantly increased SSBD. In planted forests, however, there is no significant linear relationship between SSBD and soil pH, possibly because the soil pH in planted forests, due to artificial afforestation, is mostly neutral[59].
In both planted and natural forests, older forests with larger average diameters at breast height (DBH) typically have longer successional periods[9]. As forests age, the ecosystem gradually evolves towards a more mature state, during which the number of seeds usually increases[27]. In the later stages of succession, interspecific competition among forest trees diminishes, resources shift towards reproduction, and more seeds are produced[60]. Additionally, due to prolonged seed deposition, the soil seed bank gradually accumulates more seeds. Therefore, SSBD shows a positive correlation with forest age and average DBH[27]. Experimental results indicate that forest stand density is significantly negatively correlated with SSBD. Forests with higher stand density have higher canopy closure, resulting in less light reaching the understory vegetation and soil seed germination, making nutrient uptake more difficult and lowering SSBD[61]. Studies have shown that leaf functional traits such as LA, SLA, LDMC, LN, LP, etc. can affect soil structure and nutrient cycling under the influence of leaf litter, thus disturbing soil seed bank density changes. The effect of leaf functional traits on soil seed bank density in natural and planted forest communities was driven by multi-dimensional traits rather than single traits. SSBD in both planted and natural forests decreases with an increase in leaf functional traits PC1 and PC2, indicating a consistent response of SSBD in planted and natural forests to changes in leaf functional traits. Numerous studies have shown that key leaf traits can effectively predict the productivity of forest communities[3,35,62]. In communities with higher productivity, trees allocate more resources to growth and development and engage in greater interspecific competition. Consequently, trees that reduce their reproduction result in fewer seeds produced by trees, resulting in lower SSBD[61,63,64].
Variance decomposition results indicate that, compared to climatic and forest stand factors, soil factors are the primary drivers affecting the SSBD in both planted and natural forests. This finding contradicts Hypothesis 2. Nutrients in the soil directly influence the germination and growth of soil seeds, having a more direct and intense impact than climatic factors, consistent with predictions by Yang et al. regarding global soil seed bank density influencers[2]. This study also found that among the biotic and abiotic factors affecting SSBD, soil pH is the most significant factor for natural forests, while soil nitrogen content is the most significant for planted forests. Similar results were found in Ma et al.'s study of the herb layer seed bank on the Tibetan plateau[13]. Increased soil pH enhances seed persistence, and soil pH might be indirectly influenced by precipitation, affecting SSBD in natural forests. Nitrogen, one of the most limiting factors for plant growth in terrestrial ecosystems plays a key role in influencing seed germination and growth. In planted forest ecosystems, which are generally low in nitrogen, growth is limited by nitrogen availability[53]. Acidic soils may affect seed size, lifespan, and vigor, and increased nitrogen content benefits plant carbon storage and promotes the accumulation of soil organic matter[65]. Therefore, the nitrogen content in planted forests impacts soil nutrients, and increasing nitrogen availability can alter community structure and composition. Increasing the availability of nitrogen can increase the richness of vegetation in the above-ground herbaceous layer, accelerate the growth and propagation of trees, and increase SSBD[66].
Gong et al. found that the interaction between climatic and soil factors significantly affect the ecosystem functions of planted and natural forests[3]. An et al. also discovered in their study of the soil seed bank on the Qinghai-Tibet Plateau that climatic changes affect SSBD by influencing above-ground community structure and soil nutrient availability[48]. This study also found that climatic, soil, and forest stand factors not only have a direct impact on SSBD but also that climatic factors indirectly affect SSBD in planted and natural forests by influencing forest community succession and soil nutrient availability, confirming Hypothesis 3. Rising temperatures accelerate microbial activity in the soil, increasing the decomposition rate of organic substances such as nitrogen and phosphorus. This makes more nutrients available for seeds in the soil[67]. Higher temperatures also increase community productivity, promote tree growth and development, increase forest canopy closure, reduce the light available to understory vegetation, and decrease the richness and density of the soil seed bank[68]. Increased precipitation, on the one hand, raises soil moisture and water content, increasing pathogens around soil seeds, reducing seed vigor and density[13]. On the other hand, increased precipitation limits nutrient transport in plant roots and restricts nitrogen mineralization in soil, reducing nutrients available for seed absorption[67]. Studies have found that precipitation and tree layer productivity are positively correlated; increased precipitation promotes forest tree growth. Trees adopt growth strategies over reproductive strategies with increased rainfall, reducing seed production. Additionally, tree growth increases forest canopy closure, reducing the light required for seed germination[46], thereby affecting SSBD.
This work was supported by the Xinjiang Normal University Young Top Talent Project (Grant No. XJNUQB2023-14), Natural Science Foundation of Xinjiang Uygur Autonomous Region (Grant No. 2022D01A213), Fundamental Research Funds for Universities in Xinjiang (Grant No. XJEDU2023P071), National Natural Science Foundation of China (Grant No. 32201543), Innovation and Entrepreneurship Training Program for College Students in 2023 (Grant No. S202310762004), Xinjiang Normal University Landmark Achievements Cultivation Project (Grant No. XJNUBS2301), and the Xinjiang Graduate Innovation and Entrepreneurship Project and Tianchi Talent Program.
-
The authors confirm contribution to the paper as follows: study conception and design: Gao J, Guo X; data analysis: Wang J, Wang R, Zhang X, Xu J, Zhang X; draft manuscript preparation: Wang J, Guo X, Gao J; manuscript revision: Wang J, Gao J. All authors contributed to the discussion of results, manuscript preparation, and approved the final version.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Jiangfeng Wang, Ru Wang, Xing Zhang
- Supplementary Table S1 Analysis data of this study.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang J, Wang R, Zhang X, Xu J, Zhang X, et al. 2024. Climate, soil, and stand factors collectively shape the macroscopic differences in soil seed bank density between planted and natural forests. Seed Biology 3: e020 doi: 10.48130/seedbio-0024-0020 

Climate, soil, and stand factors collectively shape the macroscopic differences in soil seed bank density between planted and natural forests
- Received: 16 August 2024
- Revised: 16 October 2024
- Accepted: 12 November 2024
- Published online: 09 December 2024
Abstract: Global climate change is intensifying forest degradation, making the soil seed bank density (SSBD) in planted and natural forests a crucial resource for ecosystem restoration. Focusing on soil seed bank density can help us assess the potential of vegetation regeneration and maintain ecosystem stability and function. However, the macro-scale distribution differences and controlling mechanisms of SSBD in these forests remain elusive. This study focuses on the SSBD in 537 natural and 383 planted forest sites across China, examining the specific impacts of climatic, soil, and forest stand factors. This study also predicts the pathways through which these factors modulate SSBD variations in both forest types. The present findings reveal that SSBD is significantly higher in planted forests compared to natural ones (p < 0.001). SSBD shows a marked declining trend with increasing temperature and precipitation (p < 0.001). In contrast, increases in sunlight duration and evapotranspiration positively correlate with SSBD in both forest types. Natural forests exhibit higher sensitivity to soil nutrient changes than planted forests. Both forest types show similar SSBD trends with changes in forest stand factors. Soil pH independently contributes the most to the spatial variation of SSBD in natural forests, while soil nitrogen content is the most significant contributor for planted forests. Mean Annual Temperature (MAT) and Mean Annual Precipitation (MAP) not only directly affect SSBD in natural forests but also indirectly through soil pH, forest stand density, and forest net primary productivity, with direct impacts outweighing the indirect. In planted forests, Mean Annual Evapotranspiration (MAE), Mean Annual Precipitation (MAP), soil nitrogen content, and stand density have a direct and significant impact on SSBD. Additionally, MAE and soil nitrogen content indirectly affect SSBD through forest stand density. The present results reveal that in forest management and administration, attention should not only be given to changes in climatic factors but also to soil nutrient loss.
-
Key words:
- Natural forests /
- Planted forests /
- Climate change /
- Soil nutrient /
- Soil seed bank density