









-
Strawberry (Fragaria × ananassa Duch.) is characterized by its vibrant colors and rich aromas. This unique and delectable fruit, with its sweet-tart flavor profile, is not only rich in essential nutrients such as vitamins, minerals, and dietary fiber, but also contains natural antioxidants, including anthocyanins, flavonoids, and phenolic compounds. Strawberries are highly valued by both producers and consumers due to their short cultivation period, extended harvest season, and significant economic benefits[1]. The quality of strawberries can be assessed through both external and internal attributes. External quality primarily encompasses fruit size, shape, and color[2,3]. Internal quality evaluation indicators include edibility, sugar content, organic acid content, vitamin C, and other nutritional components. Among these indicators, the anthocyanin content significantly influences fruit coloration, while soluble sugar and organic acid contents play crucial roles in determining fruit flavor, collectively contributing to the overall quality of strawberries[4]. As fruit quality is not determined by a single indicator, the enhancement of comprehensive quality is the primary factor influencing its market value. Previous research has predominantly focused on the impact of individual quality indicators, leaving a gap in the investigation of key factors regulating the formation of comprehensive fruit quality. In particular, the role of related transcription factors in this process requires further exploration.
The MYB transcription factor family is one of the largest transcription factor families in plants[5], extensively involved in various physiological and biochemical processes, including plant growth and development, environmental response, and physiological metabolism[6−8]. The MYB domain typically comprises one to four incomplete repeat sequences R, with each repeat sequence containing 50 to 53 amino acid residues and featuring three α-helices. The second and third helices, along with hydrophobic fragments, form a three-dimensional helix-turn-helix (HTH) structure, facilitating the binding of transcription factors to specific target genes. The third helix directly interacts with the major groove of DNA[9]. Based on the number of MYB domains, transcription factors in the MYB family are classified into 1R (R1/2, R3-MYB), 2R (R2R3-MYB), 3R (R1R2R3-MYB), and 4R[10].
MYB44 is a characteristic R2R3-MYB transcription factor with approximately 100 members in the plant genome[11]. The MYB44 gene has been closely associated with stress response in plants. In potatoes, StMYB44 negatively regulates phosphorus transport by inhibiting the expression of the phosphorus transfer-related gene StPHO1[12]. Overexpression of the MYB44 gene in Arabidopsis significantly enhances the plant's salt and drought tolerance[13]. Research has also identified a correlation between MYB44 and fruit quality. In grapes, VvMYB44-1 functions as a transcriptional repressor with an EAR inhibitory motif, and its expression is strongly induced by high-temperature treatment, negatively regulating anthocyanin biosynthesis[14]. In strawberry, the R2R3 MYB transcription inhibitor FaMYB44.2 negatively regulates the content of soluble sugars in the fruit[15]. As a highly homologous gene, FaMYB44.1 appears to have a more potent function, but its comprehensive regulation of strawberry fruit quality requires further investigation.
The DAP-seq technique is widely used in studies of quality regulation in horticultural crops, such as in Oolong tea. A study employing DAP-seq technology found that the increased number and accessibility of accessible chromatin regions in artificial hybrid Oolong tea contribute to the regulation of alternatively spliced expressed genes, influencing metabolite formation[16]. Additionally, DAP-seq was utilized to analyze key genes involved in strawberry fruit softening, ripening, and anthocyanin biosynthesis pathways regulated by FvMAPK6, revealing the role of FvRIF in the strawberry fruit ripening regulatory network[17]. Consequently, the elucidation of FaMYB44.1-mediated downstream molecular regulatory network using DAP-seq analysis is of great significance for improving the overall quality of strawberry, identifying quality-related molecular targets, and establishing an integrated regulatory network for strawberry fruit quality.
-
The 'Benihoppe' strawberry plants were cultivated in pots within a greenhouse environment under controlled conditions. These conditions included a photoperiod of 12 h light and 12 h dark, a light intensity of 300 μmol·m−2·s−1, a relative humidity of 65%, and day/night temperatures of 25 and 15 °C, respectively. For the Virus-Induced Gene Silencing (VIGS) experiment, strawberry fruits of uniform shape and size at the white fruit stage were carefully selected.
RNA extraction, fragment amplification, and expression vector construction
-
The whole-genome sequence data and genome annotation files of cultivated strawberry were obtained from the Genome Database for Rosaceae (
https://www.rosaceae.org/ ) [Reference Genome Version, Fragaria × ananassa Camarosa Genome Assembly v1.0 & Annotation v1.0.a1]. Initially, the total RNA was extracted from strawberry fruits and subsequently converted into cDNA. This cDNA fragment served as a template for cloning the FaMYB44.1 gene (augustus_masked-Fvb2-3-processed-gene-197.3). Using fragment amplification, recombinant plasmids were constructed through homologous recombination of the cloned product with the pTRV2 (TRV-FaMYB44.1) and IL60 (OE-FaMYB44.1) vectors. The primers used for RT-PCR and qRT-PCR in this study are listed in Supplementary Table S1.Transient transformation of strawberry fruits
-
The initial step involved preparing the infiltration solution. The recombinant plasmid, which tested positive, was introduced into Agrobacterium tumefaciens strain GV3101. The bacteria were then cultured in Luria-Bertani (LB) liquid medium supplemented with appropriate antibiotics at 28 °C for approximately 12 h, until the bacterial solution reached a 600 nm optical density (OD600) of approximately 0.8.
Strawberries at the white fruit stage were selected for the study. The infestation solution was carefully introduced from the fruit's tip to the hollow pith using a 1 mL syringe. This process continued until the fruit's surface exhibited a macerated appearance and ceased to absorb more solutions. To elucidate the role of FaMYB44.1 during fruit ripening, ten white fruits were chosen for infestation in each experimental group, with the inclusion of a negative control for comparison.
Extraction and determination of anthocyanin
-
The sample for testing was pulverized using liquid nitrogen, and subsequently, 0.5 g was precisely weighed and added to 10 mL of 1% hydrochloric acid methanol solution. The mixture was thoroughly agitated and subjected to 24 h of maceration in the dark for further analysis. The method for determining anthocyanin content was based on the procedure described by Yu et al.[18].
Determination of total acid content
-
A precisely weighed 0.5 g sample of fresh strawberry was thoroughly ground. An appropriate volume of carbon dioxide-free distilled water was added to the sample. The mixture was heated at 75 °C in a water bath for 30 min. After cooling to room temperature, the volume was adjusted to 20 mL. The entire 20 mL solution was filtered through dry filter paper, and the filtrate was collected. For the determination and calculation of total acid content, specific reference was made according to the method described by Yu et al.[19].
Determination of soluble sugar content
-
The content of soluble sugars was measured using High-Performance Liquid Chromatography (HPLC) equipment manufactured by Shimadzu, Japan. For each analysis, 3 g of strawberry samples were weighed, homogenized, and centrifuged. The entire supernatant was then aspirated into a new centrifuge tube and adjusted to a volume of 15 mL. This solution was filtered through a 0.22 μm aqueous filter membrane, and the resulting filtrate was collected for measurement. The experiment was repeated three times to ensure reliability. A standard curve was plotted, and the analytical method was adapted from Fang et al.[20].
DAP-seq analysis
-
The research process began with the extraction of total DNA from strawberries (fruits at all stages of development, roots, stems, leaves, and flowers). The full-length sequence of the FaMYB44.1 gene was ligated into the Halo Tag expression vector. Subsequently, the recombinant vector was subjected to in vitro expression using cloning and vector ligation reagents. Following this, DAP-seq was performed by Gidi Bio-Tech Co., Ltd. in Guangzhou. Deep Tools software was further utilized to analyze the length and depth information of genomic loci, quantify the enrichment fold of peaks, and evaluate their statistical significance[21].
Following PCR amplification, purification, and sequencing of the DNA fragments, the MACS2 analysis software was employed for peak data processing. Subsequently, the distribution of peaks within the red-faced strawberry genome was analyzed using Homer, while the ChIPseeker (R package) was utilized for functional annotation of genes associated with these peaks. Lastly, protein-binding motifs were predicted using the MEME-Chip suite[22].
In the Gene Ontology (GO) analysis, peak-related genes were mapped to various terms in the GO database, quantifying the number of peak-related genes associated with specific GO functions. For pathway significance enrichment analysis, the Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway was utilized as the unit of analysis. Subsequently, the hypergeometric test was employed to identify significant GO terms and pathways enriched in peak-related genes relative to the background.
Initially, the MEME-chip from the MEME Suite was employed to identify statistically significant motif sequences within the peak sequences. Subsequently, the FIMO tool, also part of the MEME Suite, was utilized to precisely locate all motifs predicted by MEME and DREME within the peak sequences, thereby identifying potential transcription factor binding sites.
Statistical analysis
-
Three biological replicates were conducted for each sample in the experiment to ensure data reliability. All data were presented as mean ± standard deviation (SD). Statistical analysis was performed using SPSS software. Statistical significance was determined using a t-test (* p < 0.05 and ** p < 0.01).
-
To elucidate the function of FaMYB44.1 in strawberry fruit ripening and quality, a transient transformation technique was employed to suppress FaMYB44.1 gene expression at the white fruit stage. Five days after injection of TRV-FaMYB44.1 resuspension, phenotypic alterations were examined in strawberries, and relevant quality parameters were assessed. The findings revealed an accelerated ripening process in FaMYB44.1 transgenic strawberries, accompanied by a decrease in the relative expression level of FaMYB44.1 compared to the control group (Fig. 1a & b). Furthermore, FaMYB44.1 transgenic strawberries exhibited significantly higher anthocyanin content than the control group, while soluble sugar content showed a slight, non-significant increase. Notably, the organic acid content was significantly reduced in the transgenic fruits (Fig. 1c–e). Further experimental validation was performed through transient overexpression of FaMYB44.1 in strawberry fruits. (Supplementary Fig. S1). The results demonstrated that FaMYB44.1 overexpression significantly delayed fruit ripening, as evidenced by reduced anthocyanin accumulation and elevated organic acid levels compared to control fruits. These findings provide reciprocal genetic evidence that strongly corroborates initial observations from the silencing experiments, confirming the dual regulatory role of FaMYB44.1 in modulating both anthocyanin biosynthesis and organic acid metabolism during strawberry fruit ripening.
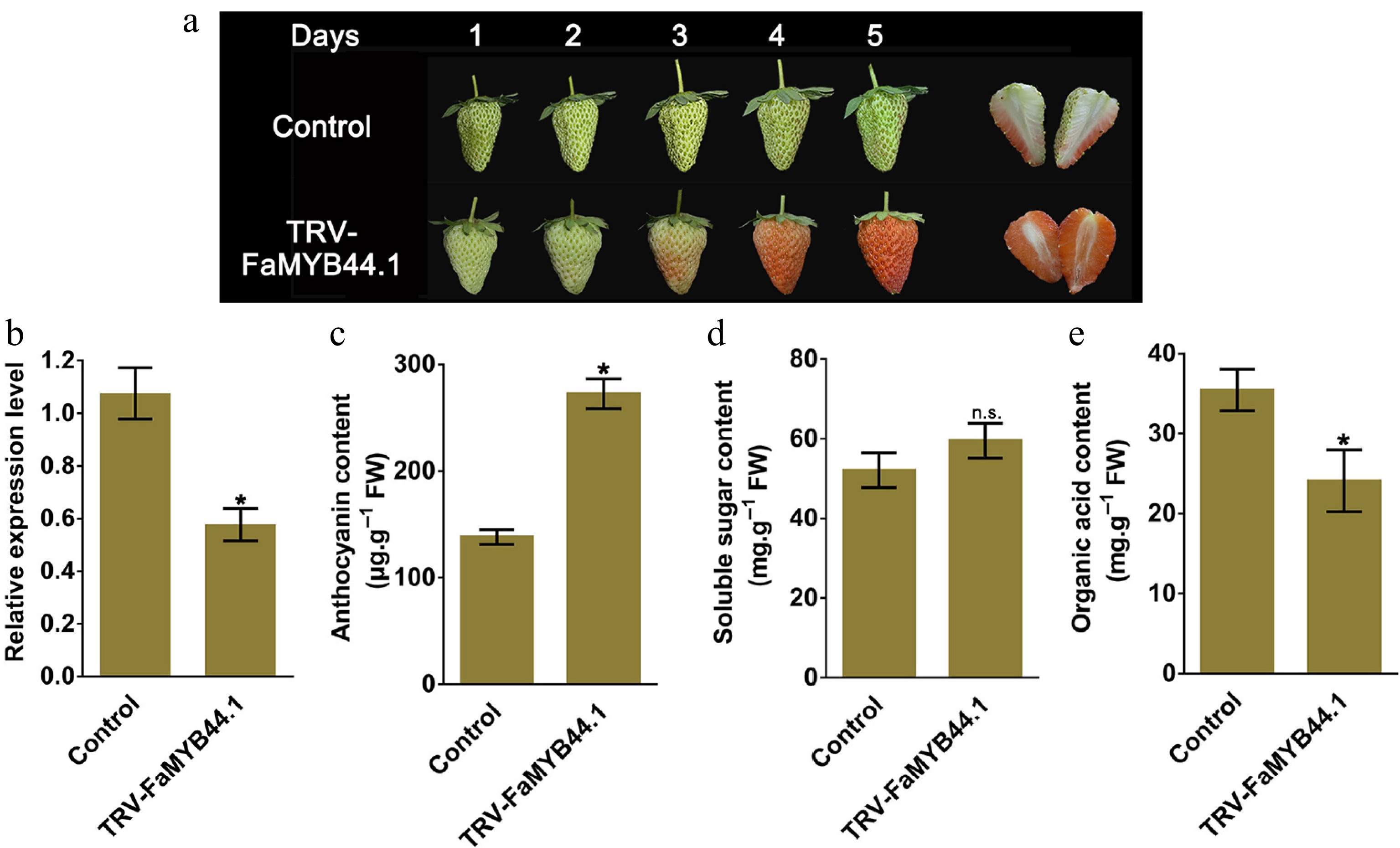
Figure 1.
Five days after injecting the FaMYB44.1 bacterial suspension, changes in strawberry fruit characteristics and related parameters were measured. (a) Phenotypic changes. (b) FaMYB44.1 expression levels. (c) Anthocyanin content. (d) Soluble sugar concentration. (e) Organic acid content in TRV-FaMYB44.1 fruits. Statistical significance compared to the control group is indicated as follows: n.s. = No significant difference; * = p < 0.05. Values represent the meaning of three biological replicates ± SD.
Peak scanning on a whole-genome scale using DAP-Seq sequencing
-
FaMYB44.1, a representative MYB transcription factor, directly regulates the transcription of downstream target genes, thereby influencing various quality indicators. To further investigate the FaMYB44.1 transcription factor and refine the regulatory network of downstream quality-related genes, genomic DNA was extracted from strawberry fruits for DAP-seq analysis. To ensure the accuracy of the sequencing analysis, single-sample peak detection was initially performed, followed by intra-group peak merging to generate unified peak data. The results revealed that FaMYB44.1-R1 and FaMYB44.1-R2 identified 28,229 and 29,149 peaks, respectively (DAP sequencing enrichment regions, q-value < 0.05), with average peak lengths of 439 and 448 bp, average depths of 93 and 98 bp, and peak binding sites accounting for 7.17% and 7.55% of the genome. The reproducibility between the two samples was high, with peak lengths predominantly distributed around 300 bp and minimal distribution beyond 1,500 bp (Fig. 2a & b). The sequencing depth within the peak regions was concentrated in the range of five to seven (plotted on a log2 scale); as the depth increased, the proportion of peaks above a given sequencing depth rapidly decreased, approaching zero (Fig. 2c & d).
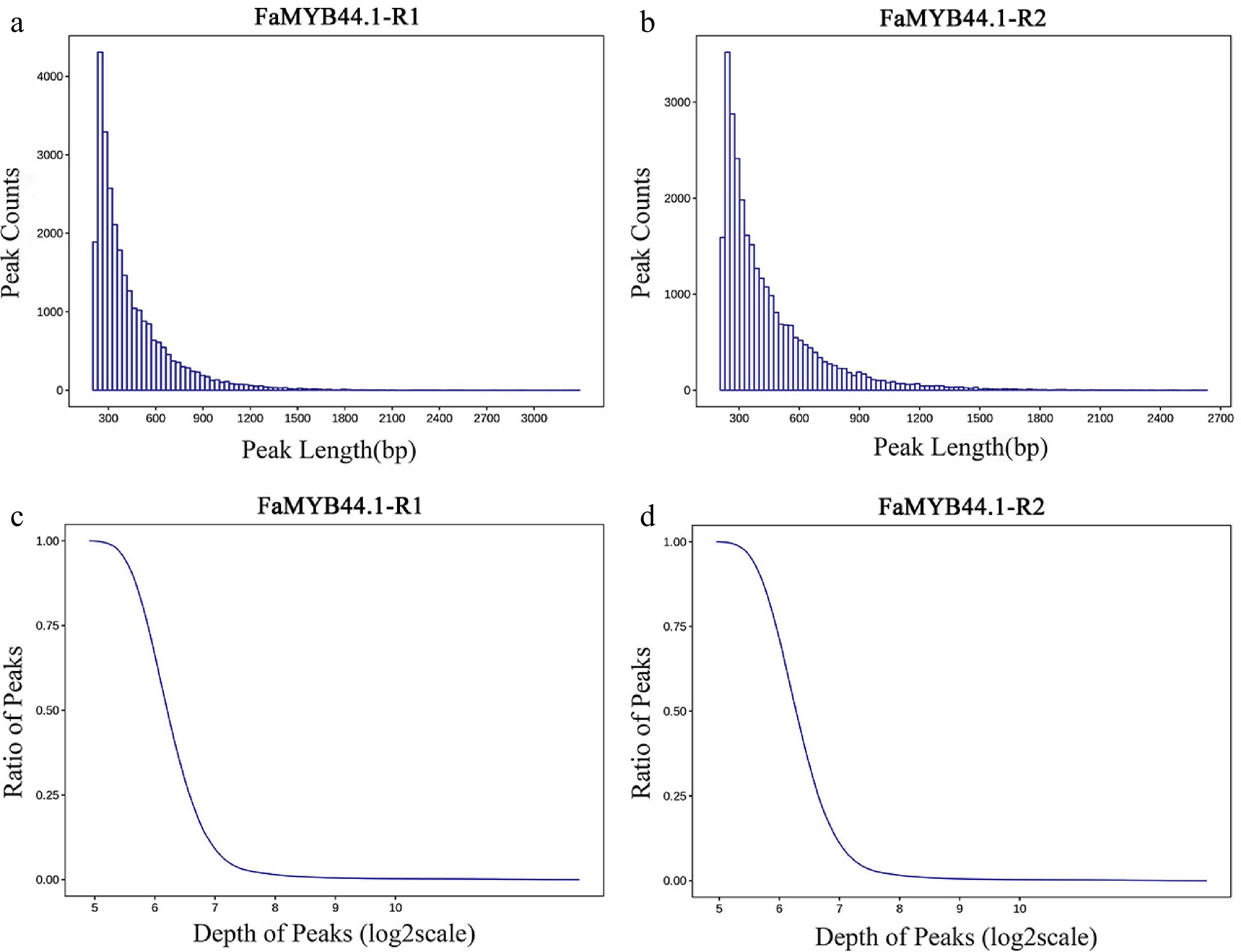
Figure 2.
Distribution of peak length and depth. (a) Peak length distribution for FaMYB44.1-R1, demonstrating a concentration around 300 bp. (b) Peak length distribution for FaMYB44.1-R2, exhibiting similar characteristics to FaMYB44.1-R1 with a peak around 300 bp. (c) Peak ratio vs sequencing depth (log2 scale) for FaMYB44.1-R1, displaying a concentration in the five to seven range. (d) Peak ratio vs sequencing depth (log2 scale) for FaMYB44.1-R2, reflecting the pattern observed in FaMYB44.1-R1.
Figure 3 illustrates the distribution of enrichment folds and confidence levels [–log10(p-value)] for peaks in FaMYB44.1-R1 and FaMYB44.1-R2. The analysis reveals that the enrichment folds of peaks in both FaMYB44.1-R1 and FaMYB44.1-R2 are primarily concentrated between two and six (Fig. 3a & b), while the –log10(p-value) values predominantly fall between zero and 100 (Fig. 3c & d). These findings indicate a high degree of sample reproducibility, thereby facilitating the merging of grouped peaks.
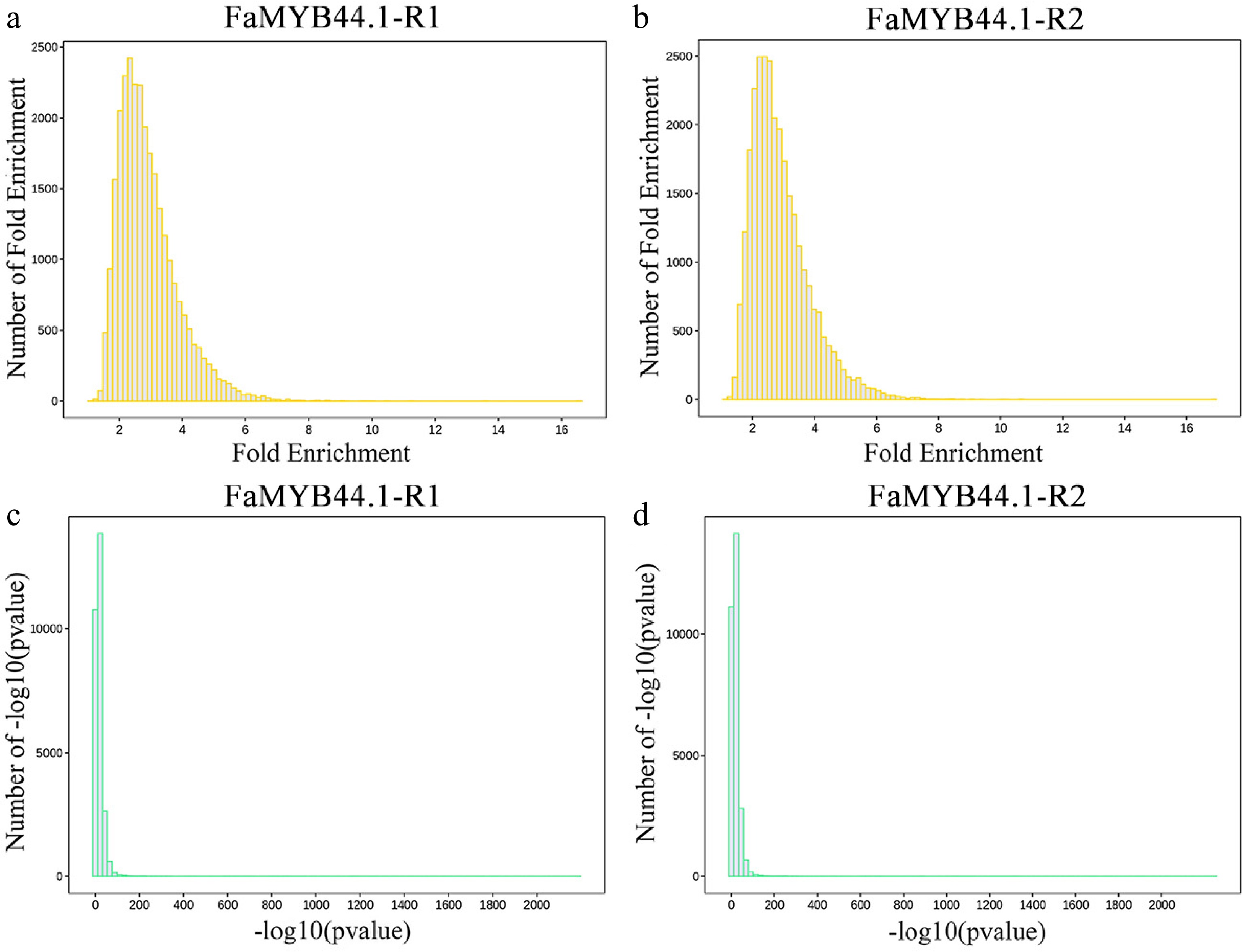
Figure 3.
Distribution of peak enrichment multiples and confidence levels. (a) FaMYB44.1-R1 fold enrichment distribution, with peaks predominantly ranging from two to six folds. (b) FaMYB44.1-R2 fold enrichment distribution, exhibiting a comparable peak concentration. (c) FaMYB44.1-R1 –log10(p-value) distribution, demonstrating significant peaks concentrated between zero and 100. (d) FaMYB44.1-R2 –log10(p-value) distribution, reflecting a similar significant peak concentration as observed in FaMYB44.1-R1.
Distribution sites of peaks in the genomic region
-
The analysis involved filtering peaks between replicate samples within the group, preserving shared peaks with over 50% overlap for subsequent examination. The findings revealed that FaMYB44.1-R1 exhibited 4,278 unique peaks, while FaMYB44.1-R2 demonstrated 5,318 unique peaks; collectively, the two samples shared 23,563 peaks (Fig. 4a). The distribution of peaks indicated that intronic regions (comprising other intron and first intron) accounted for 46.37% (35.01% + 11.36%), followed by promoters (≤ 1 kb + 1–2 kb) and untranslated regions (UTRs) (5' UTR + 3' UTR), representing 16.20% (11.84% + 4.36%) and 14.99% (2.93% + 12.06%), respectively. Conversely, the proportion of downstream peaks (≤ 500) was minimal, constituting only 4.36% (Fig. 4b).
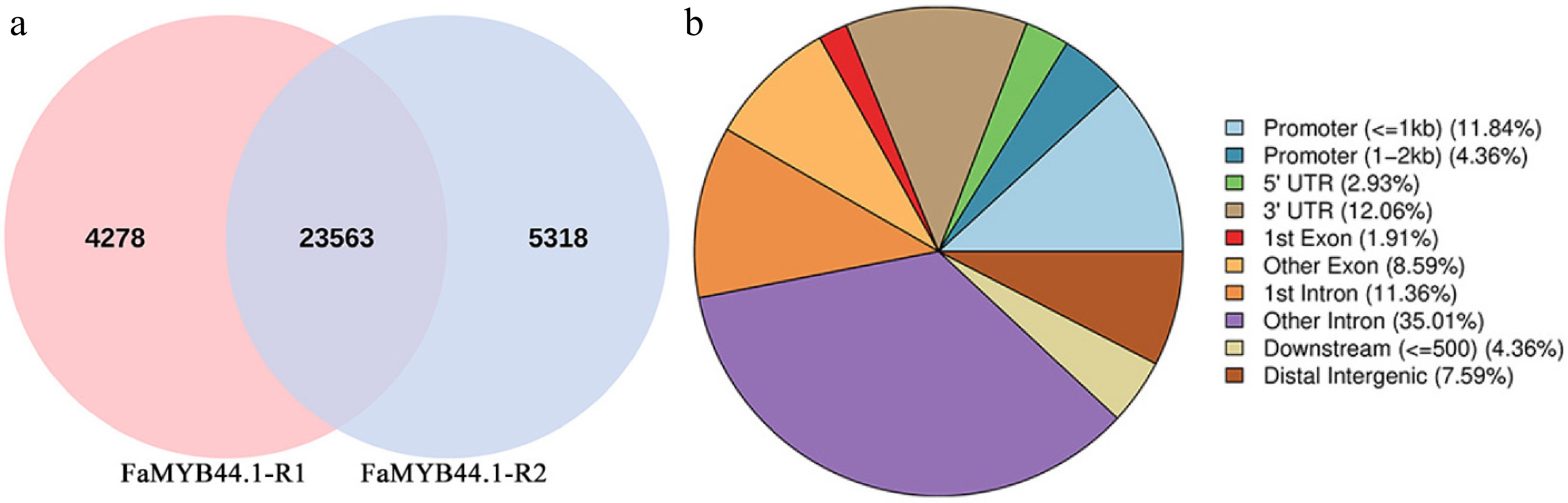
Figure 4.
Venn diagram of peaks and pie chart of peak distribution on gene functional elements. (a) Venn diagram depicting 4,278 unique peaks for FaMYB44.1-R1, 5,318 for FaMYB44.1-R2, and 23,563 shared peaks. (b) Pie chart illustrating peak distribution across gene elements, with intronic regions comprising 46.37%, promoters 16.20%, and UTRs 14.99%, while downstream peaks constitute the smallest proportion at 4.36%.
GO functional analysis of peak-related genes
-
Integrating peak region information from the genome with gene annotation data, GO enrichment analysis was conducted on the identified target genes associated with these peaks. The analysis revealed significant enrichment patterns across different categories. Within the cellular components (CC) category, target genes exhibited notable enrichment in organelle parts, intracellular organelle parts, and membrane-bound organelles (Fig. 5a). In the molecular function (MF) category, significant enrichment was observed in nucleoside binding and purine nucleoside binding (Fig. 5b). The biological process (BP) category demonstrated enrichment in cellular processes of single organisms, transport, and regulation of biological processes (Fig. 5c).
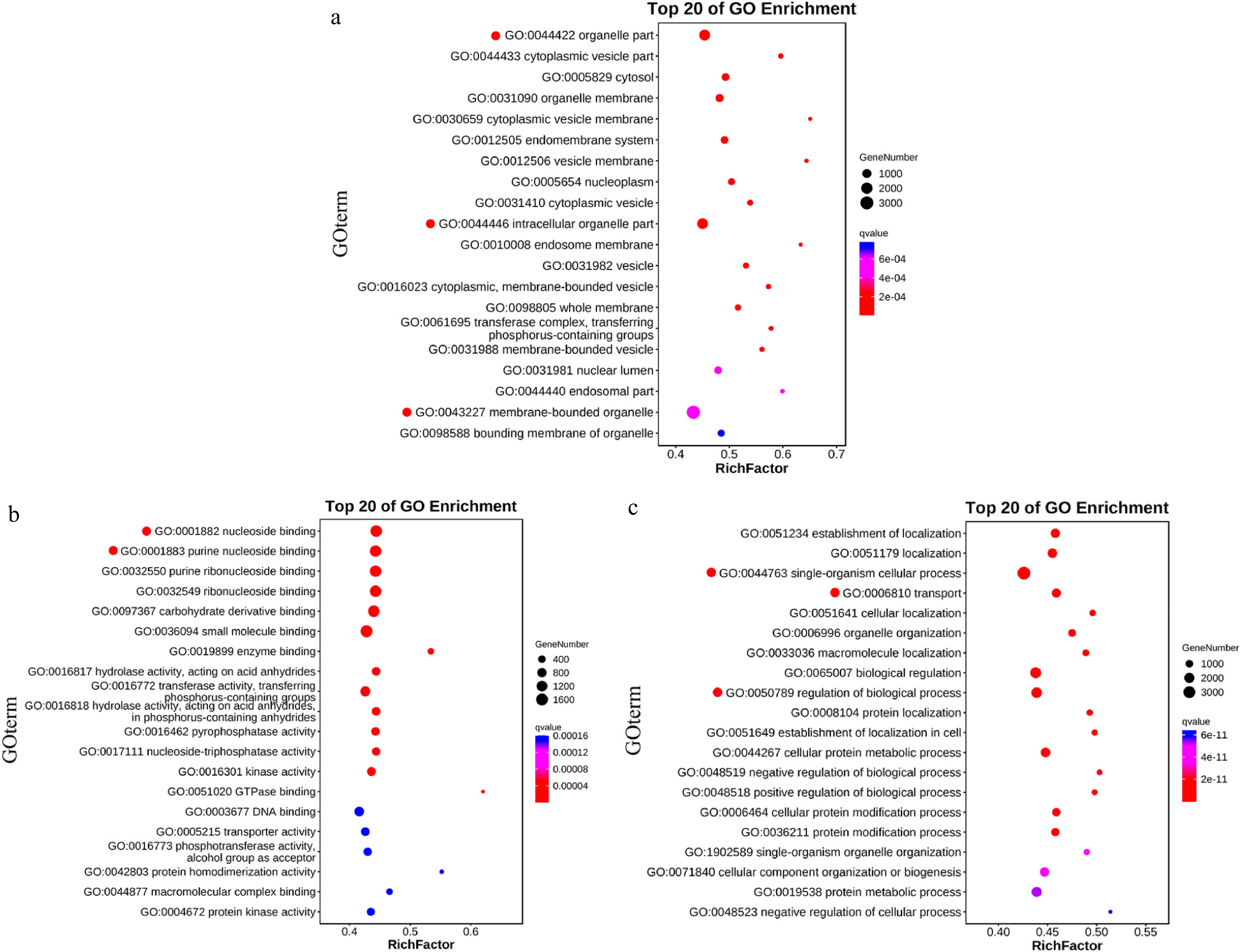
Figure 5.
Enrichment bubble chart. (a) CC enrichment demonstrates significant representation in organelle parts, intracellular organelle parts, and membrane-bound organelles. (b) MF enrichment indicates substantial enrichment in nucleoside binding and purine nucleoside binding. (c) BP enrichment reveals notable representation in cellular processes of single organisms, transport, and regulation of biological processes.
KEGG functional analysis of peak-related genes
-
Based on the identified peak-related target genes, KEGG enrichment analysis was conducted. The results revealed that the top three pathways in the KEGG enrichment analysis were RNA degradation, the mRNA surveillance pathway, and nucleocytoplasmic transport (Fig. 6a). Further investigation of quality-related pathways, including those associated with sugars, acids, and anthocyanins, demonstrated that the downstream pathways regulated by FaMYB44.1 encompass carbohydrate metabolism pathways linked to soluble sugars, such as the pentose phosphate pathway, starch and sucrose metabolism, glycolysis/gluconeogenesis, and fructose and mannose metabolism. Additionally, the citrate cycle (TCA cycle) pathway relevant to organic acids and biosynthetic pathways for other secondary metabolites related to fruit colour, including flavonoid biosynthesis and flavone and flavanol biosynthesis, were identified. Moreover, the analysis indicated that genes such as SDH3-1, FLS, and SPS1 are crucial target genes within the regulatory network controlled by FaMYB44.1 (Fig. 6b).
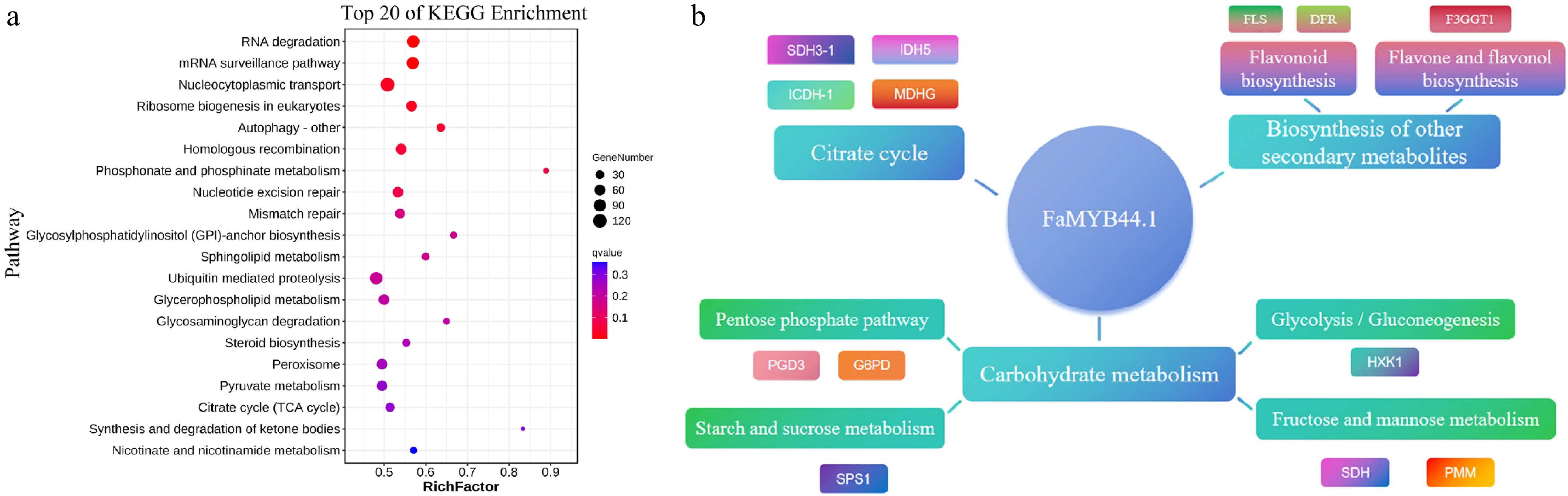
Figure 6.
KEGG pathway enrichment and regulatory network of FaMYB44.1. (a) Top 20 KEGG pathways enriched among peak-associated genes, emphasizing RNA degradation, mRNA surveillance, and nucleocytoplasmic transport. (b) Regulatory network centered on FaMYB44.1, linking to pathways involved in carbohydrate metabolism, citrate cycle, and secondary metabolite biosynthesis, with key target genes including SDH3-1, FLS, and SPS1.
Analysis of peak-binding characteristic motifs
-
FaMYB44.1, a typical MYB transcription factor in strawberries, regulates the transcription of target genes by recognizing specific DNA sequences. This function contributes to a complex regulatory system that influences fruit quality and ripening. Utilizing the MEME Suite (
http://meme-suite.org/ ) for motif analysis of peak binding characteristics, nine MEME motifs were identified, one DREME motif, and two CentriMo motifs that met the significance threshold (e < 0.05). Figure 7a displays the sequences of the top six motifs. Additionally, a comparative analysis using the motif Tomtom revealed that the secondary motif of MEME-1 closely resembles the known DOF5.4 (MA1280.1), which belongs to the C4-type zinc finger factor; both motifs contain a DREME module (Fig. 7b). These findings suggest that DREME may serve as the binding site for FaMYB44.1 in strawberries.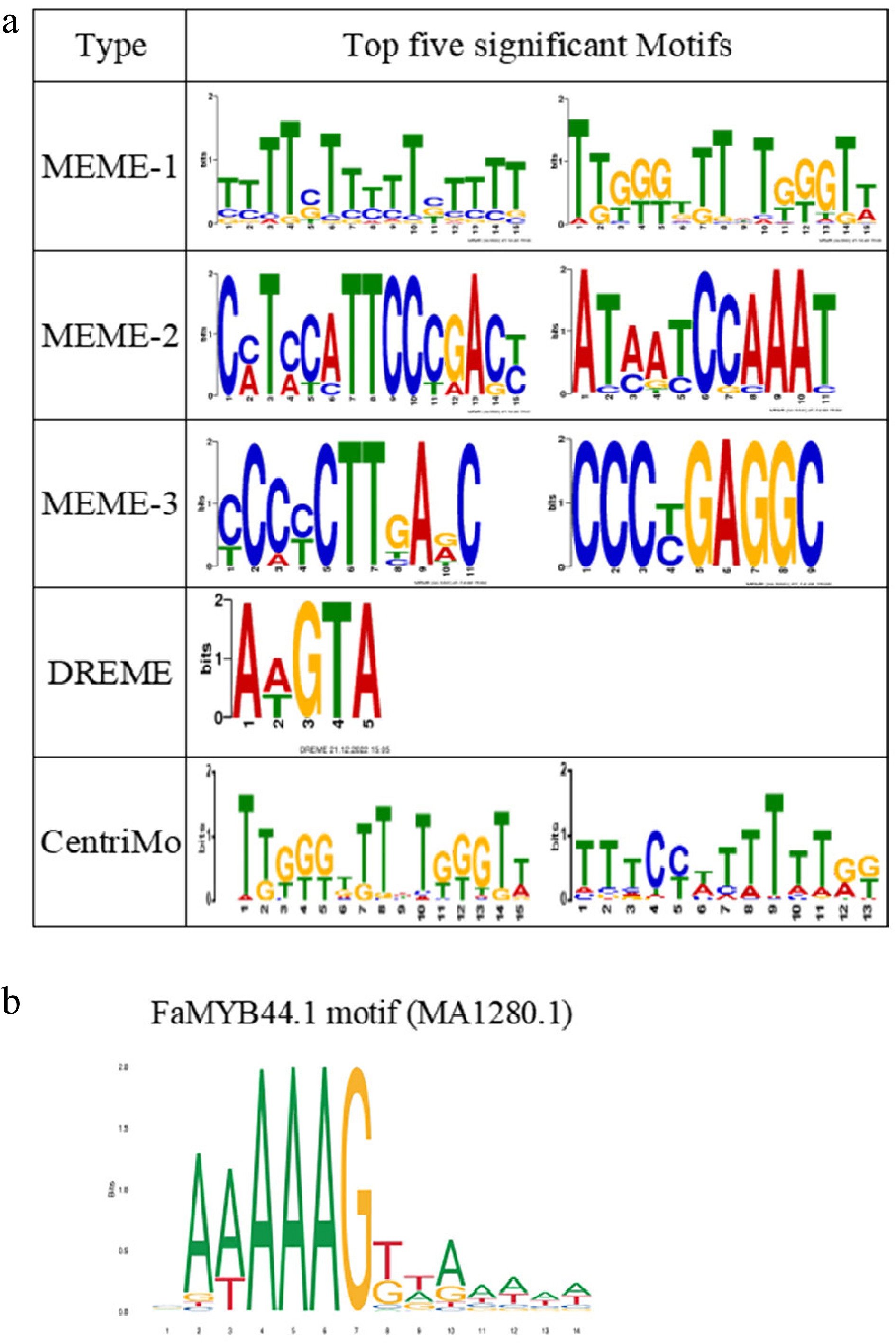
Figure 7.
Identification of FaMYB44.1 DNA binding motifs. (a) The five most significant motifs identified using MEME Suite, DREME, and CentriMo, displaying the consensus sequences and their frequency logos. (b) The FaMYB44.1 motif (MA1280.1) closely resembles the secondary motif of MEME-1, suggesting its potential binding site in strawberry.
-
Strawberries, widely renowned for their vibrant color (appearance quality) and rich flavor (intrinsic taste quality), are a popular fruit globally. The regulation of strawberry quality involves multiple intricate biological processes, including anthocyanin synthesis, organic acid metabolism, and sugar accumulation. Members of the MYB transcription factor family play a pivotal role in plant secondary metabolism, particularly in anthocyanin synthesis and accumulation. Liu et al. identified 146 R2R3-MYB transcription factor genes in 'Nanguo pear' through genome-wide identification and expression analysis, revealing that the expression levels of 12 genes positively correlated with anthocyanin accumulation[23]. An et al. discovered that MdMYB308L interacts with MdbHLH33, enhancing the expression of genes related to anthocyanin biosynthesis and consequently increasing anthocyanin accumulation in apple fruits[24]. Furthermore, Zhang et al. demonstrated that FaRAV1 promotes anthocyanin accumulation in strawberries by directly activating the promoters of anthocyanin pathway genes[25].
Members of the MYB family have been implicated in various primary metabolic processes in plants, including organic acid metabolism, sugar synthesis, and catabolism. For example, Liu et al. demonstrated through a dual-luciferase assay that FaMYB5 activates FaCS2 expression while suppressing FaACO and FaGAD transcription, thereby influencing citric acid accumulation[26]. Wang et al. reported that the R3-MYB transcription factor CsCPC inhibits citric acid accumulation in citrus by downregulating CsPH1 and CsPH5 expression[27]. Chen et al. showed that MYBS1 and MYBS2 play essential roles in sugar signaling in Arabidopsis[28]. Yuan et al. identified MYB169, MYB181, and MYB192 as potential regulators of C4 photosynthesis[29]. Current research indicates that MYB transcription factors are involved in both primary and secondary metabolic processes in fruits. However, studies on their comprehensive regulation of fruit quality through primary and secondary metabolism remain limited. In this study, based on physiological data (Fig. 1) showing that suppression of FaMYB44.1 expression significantly enhanced anthocyanin accumulation while reducing organic acid content, and DAP-seq analysis (Fig. 6b) revealing that FaMYB44.1 binds to promoter regions of key anthocyanin biosynthesis genes (DFR, FLS, F3GGT1, etc.) and organic acid metabolism-related genes (SDH3-1, IDH5, ICDH-1, MDHG, etc.), it is proposed that FaMYB44.1 plays dual regulatory roles in pigmentation and acid metabolism. FaMYB44.1 is demonstrated to significantly enhance the overall quality of strawberry fruits by affecting multiple primary and secondary metabolic pathways, particularly influencing quality indicators such as anthocyanin and organic acid levels.
FaMYB44.1 possesses two highly homologous counterparts in strawberries, FaMYB44.2 and FaMYB44.3, all of which are evolutionarily closely related to genes such as AtMYB44 in Arabidopsis. Research has demonstrated that FaMYB44.1, FaMYB44.2, and FaMYB44.3 exhibit distinct expression patterns during fruit ripening[15]. Specifically, the expression of FaMYB44.2 increases progressively with fruit development and maturation, correlating with the pattern of sugar accumulation. In contrast, FaMYB44.3 displays an irregular expression pattern of increase-decrease-increase-decrease. Notably, the expression of FaMYB44.1 rises modestly during the fruit expansion phase and then declines gradually during the color change phase, mirroring the patterns of organic acid and anthocyanin variation. These findings collectively suggest that FaMYB44.1 is involved in multiple metabolic pathways associated with strawberry fruit quality formation and regulates the quality attributes of ripe strawberries.
DAP-seq sequencing analysis is a widely utilized method for elucidating the downstream regulatory networks of transcription factors. Wang et al. employed DAP-seq technology to conduct a comprehensive analysis of the ZBED1 gene's downstream regulatory network[21]. They discovered that the gene's downstream targets are primarily involved in several key biological processes, including axon guidance, axon regeneration, the Hippo signaling pathway, cell cycle, translation, apoptosis, and the PI3K-Akt signaling pathway. These processes play a crucial role in the sexual size dimorphism of Cynoglossus semilaevis. Similarly, Wang et al. applied DAP-seq technology to analyze the downstream regulatory network of the AhHY5 gene[30]. Their findings revealed that AhHY5 directly regulates 519 light-induced genes, predominantly associated with biological processes such as flavonoid metabolism, UV response, signal transduction regulation, and oxidative stress response. To corroborate these conclusions, a comprehensive analysis of the downstream regulatory network of FaMYB44.1 was conducted using DAP-seq technology. The results demonstrated that the downstream target genes of FaMYB44.1 are involved in various metabolic pathways. These include the Citrate cycle and carbohydrate metabolism within primary metabolic processes, as well as Flavonoid biosynthesis and Flavone and flavanol biosynthesis within the Biosynthesis of other secondary metabolites pathway. These findings further support the initial observations.
-
This study investigates the role of FaMYB44.1 in strawberry fruit maturation and quality formation. Transgenic strawberries with silenced FaMYB44.1 expression displayed significant changes in anthocyanin and organic acid levels. Utilizing DAP-seq at the whole-genome level, it was identified that FaMYB44.1 binds to various genomic regions, including promoters, UTRs, exons, and introns, with the highest frequency in intronic sequences. The predicted target genes are overrepresented in multiple primary and secondary metabolic pathways, such as the carbohydrate metabolism pathway associated with soluble sugars, the citrate cycle (TCA cycle) involved in organic acid metabolism, and the flavonoid biosynthesis and flavone and flavanol biosynthesis pathways linked to fruit pigmentation. Motif analysis revealed that DREME is a binding motif for FaMYB44.1 in strawberries. These findings demonstrate that FaMYB44.1 regulates multiple metabolic processes, thereby enhancing overall strawberry fruit quality and offering a valuable gene target for molecular breeding initiatives.
This research was supported by grants from the Natural Science Foundation of Jiangsu Province (Grant No. BK20230572), the Zhongshan Biological Breeding Laboratory (Grant No. ZSBBL-KY2023-08), the National Natural Science Foundation of China (Grant No. 32402527), and the Basic Science (Natural Science) Research Project in Universities of Jiangsu Province (Grant No. 23KJB210015). We appreciate the linguistic assistance provided by TopEdit (
www.topeditsci.com ) during the preparation of this manuscript.-
The authors confirm contributions to the paper as follows: study conception and design, funding acquisition, resources: Yu JQ, Sheng LX; methodology: Liu Y, Bao MY, Zhang R, Lin Y, Li HYH, Yu JQ; software: Liu Y; validation, investigation, data curation: Yu JQ, Liu Y; writing—original draft preparation: Liu Y, Bao MY; writing—review and editing, formal analysis, visualization, supervision, project administration: Yu JQ. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yang Liu, Ming-Yue Bao
- Supplementary Table S1 The primers used for RT-PCR and qRT-PCR in this study.
- Supplementary Fig. S1 FaMYB44.1 was transiently overexpressed in strawberry fruits, and the changes in phenotypes and related indexes were determined after 5 days of treatment. (a) Phenotypic changes, (b) FaMYB44.1 expression levels, (c) anthocyanin content, (d) soluble sugar concentration, and (e) organic acid content in OE-FaMYB44.1 fruits. Statistical significance compared to the control group is indicated as follows: n.s. = No significant difference; * = p < 0.05. Values represent the meaning of three biological replicates ± SD.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Y, Bao MY, Zhang R, Li HYH, Lin Y, et al. 2025. The downstream molecular network of FaMYB44.1, a key factor in strawberry quality regulation, was investigated by DNA affinity purification sequencing. Fruit Research 5: e034 doi: 10.48130/frures-0025-0024 

The downstream molecular network of FaMYB44.1, a key factor in strawberry quality regulation, was investigated by DNA affinity purification sequencing
- Received: 08 May 2025
- Revised: 30 June 2025
- Accepted: 14 July 2025
- Published online: 02 September 2025
Abstract: Strawberry (Fragaria × ananassa Duch.) is a small berry characterized by its bright color and sweet-sour flavor. With the global rise in living standards, the strawberry industry has experienced rapid growth, and enhancing the comprehensive quality of the fruit has become a focal point in current strawberry research. Previous studies often concentrated on improving single quality indices, with a notable lack of regulatory factors for enhancing overall strawberry fruit quality. In this study, transgenic strawberries with inhibited FaMYB44.1 expression were developed using instantaneous transformation technology. Further analysis of related quality indices revealed that FaMYB44.1 is a MYB transcription factor that inhibits fruit ripening. TRV-FaMYB44.1 transgenic strawberry fruits exhibited significantly increased anthocyanin content and markedly decreased organic acid levels. DAP-seq analysis demonstrated that FaMYB44.1 binds to multiple promoters, untranslated regions (UTRs), exons, and introns within the genome. The associated target genes are enriched in various primary and secondary metabolic pathways, including carbohydrate metabolism, citric acid cycle (TCA cycle), flavonoid biosynthesis, and flavonoid and flavanol biosynthesis. Motif analysis identified DREME (AAGTA) as the binding motif of the strawberry FaMYB44.1 transcription factor. This research confirms FaMYB44.1's involvement in regulating diverse metabolic processes, contributing to the improvement of overall strawberry fruit quality, and provides a valuable gene target for molecular breeding endeavors.
-
Key words:
- Strawberry /
- Fruit quality /
- FaMYB44.1 /
- DAP-seq /
- Transcriptional regulation