









-
The contamination of ecosystems with heavy metals has become a concern due to their toxicity to biota and also due to them being permanently immobilised in soil components. Moreover, metals influence soil microbes by decreasing their population size, diversity and biochemical activity, thereby altering the structure of soil microbial communities[1−3]. Exposure to metals can also result in the establishment of tolerant microbial populations in soil, with important roles in the ecosystem.
Arsenic (As) is a ubiquitous trace metalloid present in almost all environments and widely distributed in soil and water[4]. The amounts of As in non-polluted soils are usually lower than 10 mg·kg−1[5]. The occurrence of high levels of As in soils results from human activities, such as mining and smelting processes, in addition to the use of As-based compounds such as insecticides, herbicides, fungicides, algaecides, sheep dips, wood preservatives, dyestuffs and feed additives[5]. Natural sources of As comprise volcanic activities, wind-born soil particles, sea salt sprays and microbial volatilization[6,7].
Currently available tools for the remediation of soils polluted with metals or metalloids are costly, time consuming, can be dangerous to people and result in the production of secondary waste[8,9]. Phytoremediation has become known as a promising eco-remediation technology by which plants and their associated microbiota are used to eradicate contaminants from polluted soils[10,11], acting through phytoextraction, stabilization, immobilization, volatilization and rhizofiltration[12−15].
Soils contaminated by heavy metals frequently have low percentages of organic matter and available nitrogen, limiting plant development[16]. Legume plants and their associated root nodule bacteria (rhizobia) are essential components of the biogeochemical cycles in both natural and agricultural ecosystems. We consider that this association could aid rehabilitation of disturbed areas by adding fixed atmospheric nitrogen that can be used by other plants (non-legumes). However, the growth and development of many plant species, including legumes, is affected by the occurrence of high amounts of toxic chemical elements such as As.
Compared to other legumes, Lotus species have a higher potential for adaptability to abiotic stresses, frequently surviving in adverse conditions such as those found in polluted soils[17,18]. Lotus is a cosmopolitan genus with two main centers of diversity, the Mediterranean region (including portions of Europe, Africa, and Western Asia) and Western North America[19,20]. The genus Lotus comprises about 100 annual and perennial species[18]. However, only a small number of Lotus species, mainly of agronomic interest, have been studied in relation to their symbionts[21,22]. The species of Lotus that have been domesticated and improved by selection and plant breeding are: L. corniculatus, L.; L. uliginosus Schkuhr., also denoted as L. pedunculatus; L. glaber Mill., also denoted as L. tenuis; L. subbiflorus Lagasca and L. ornithopodioides[18]. On the other hand, L. japonicus is used as a model for genetic and molecular studies[23−25].
L. uliginosus is a perennial legume also used as a tropical forage with a high productive capacity and is recommended for extensive livestock areas in countries of the South American cone and especially in Chile, Brazil and Uruguay. This species was also naturalised in Argentina. Its use is in cattle and sheep rearing and fattening processes and also for soil remediation. It thrives in varied conditions, especially low fertility and moisture as well as soil acidity. It is a species with high spring-summer-autumn forage potential comparable to other traditionally used legumes. This specie has also a outstanding nutritional value, with the presence of condensed tannins, giving it additional nutritional advantages[26]. Due to its importance several authors have studied the diversity and phylogenetic relationships of root nodule bacteria of L. uliginosus collected from fields in different countries, such as Uruguay[27], Portugal[28] and Belgium[29]. These authors indicated that those strains were affiliated with Bradyrhizobium japonicum or Bradyrhizobium sp.
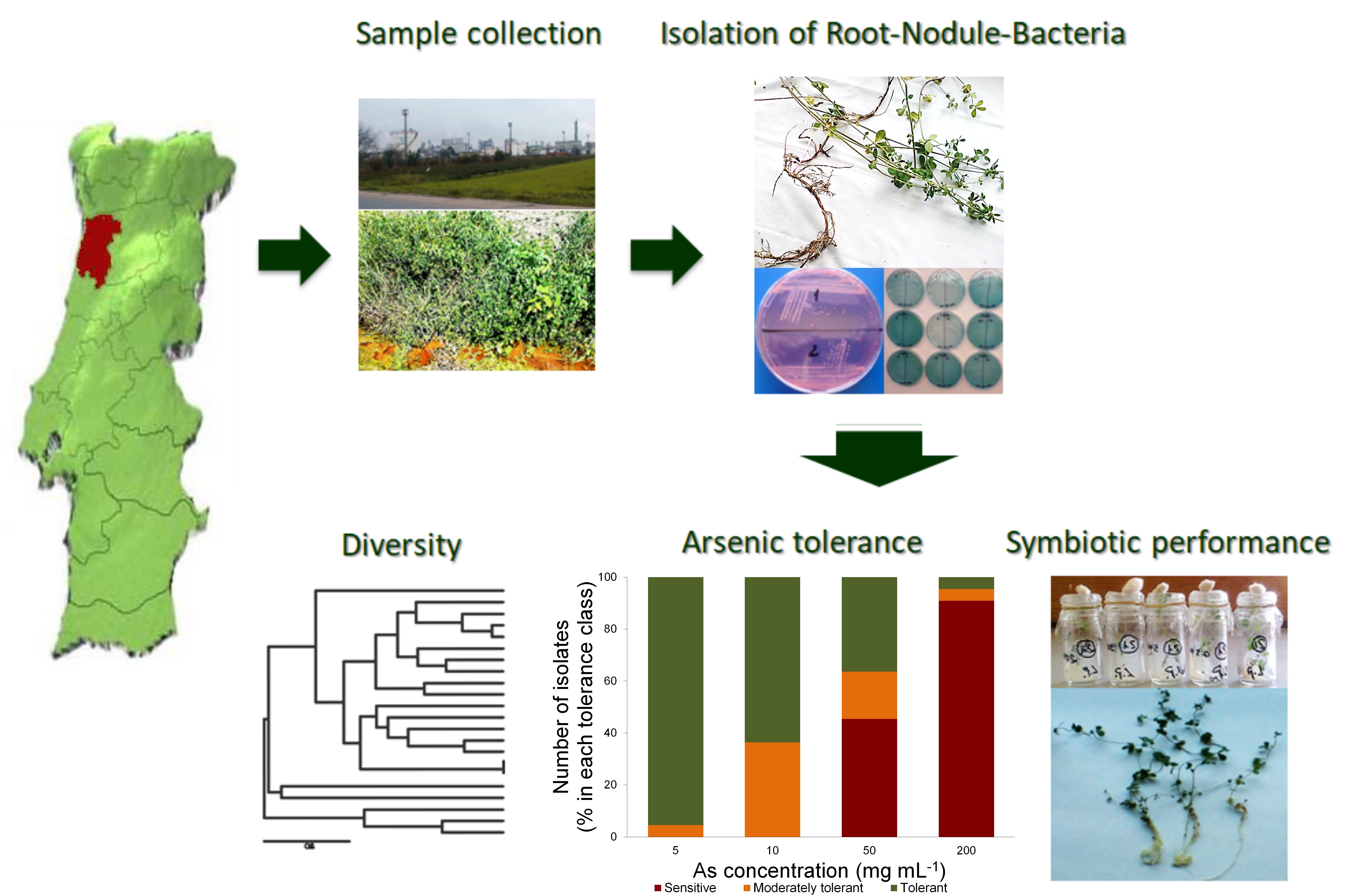
The goal of this research was to investigate a symbiotic legume system effective in nitrogen fixation in a polluted site for possible use during remediation of such soils. In view of the future use of legumes for soil improvement, it is essential to assess the effect of As on the functioning of the symbiosis. Therefore, the objective of this work was also to study the rhizobial population associated with L. uliginosus growing in As contaminated soils with regards to genetic diversity, efficiency of nitrogen-fixation and tolerance to As (Supplemental Fig. S1).
-
Bacteria were isolated from root nodules of Lotus uliginosus growing under field conditions on an industrially contaminated soil, in the centre of Portugal (GPS: 40.778978329572496, −8.59899224806268), containing high levels of arsenic (1.5 × 103 mg·kg−1). Six plants were collected and 5−6 nodules were randomly taken from each plant. The nodules were carefully detached from the roots, surface sterilized (0.25% solution of HgCl2)[28] and washed extensively with sterile water. Afterwards, nodules were individually crushed and spread on a plate with yeast-mannitol agar (YMA) supplemented with congo red dye[30]. Plates were incubated at 28 ºC in the dark for 7-10 d. The isolate purity was checked by examining colony morphology and congo red absorption. Isolates were also cultivated in yeast-mannitol agar (YMA) supplemented with bromothymol blue (BTB)[30]. Isolates were kept at 4 ºC.
Genetic diversity
-
ERIC (Enterobacterial Repetitive Intergenic Consensus) - PCR[31] was used to assess the genetic diversity of 22 isolates obtained from nodules of L. uliginosus as described above. This procedure is used to distinguish strains that are taxonomically very close and has been recognized as appropriate for assessing rhizobial diversity[32,33]. DNA was extracted from bacterial liquid cultures using the Aqua pure genomic DNA extraction kit from Bio-Rad, following the kit protocol specifications. ERIC fingerprints were generated using the primers ERIC1R and ERIC2 previously reported by de Bruijn and Versalovic et al.[31,34]. Matrices of the Dice coefficient were calculated and cluster analysis was performed using the UPGMA (Unweighted Pair Group with Arithmetic Average) algorithm and the program Free Tree[35,36]. The program Tree View (PHILIP) was used for the construction of dendrograms and evaluation of the respective genetic relationships.
16S rRNA region amplification and sequencing
-
The 16S rRNA region was amplified in seven selected isolates using 41F and 1488R primers as described by Weisburg et al.[37]. PCR reactions primers were used at a concentration of 5 μM, together with 15.8 μl of Qiagen kit Taq mix solution (2.5 U of Taq polymerase, 1.5 mM of MgCl2, 200 μM of the different dNTP), and approximately 20 ng of genomic DNA, in a final volume of 20 μl. Amplifications were performed following the conditions of Weisburg et al.[37], using an Eppendorf Mastercycler Gradient thermocycler. PCR products of the amplified 16S rRNA region, with an expected size of about 1500 bp, were confirmed by agarose gel electrophoresis. These products were sequenced, with the same primers used for PCR amplification, with an ABI 3730 XL automated sequencer, by STAB VIDA, Caparica, Portugal. Obtained nucleotide sequences were subjected to quality control and edited as necessary using the DNA chromatogram files in Chromas Lite program (version 2.1.1). Homologous sequences were searched in NCBI (National Center for Biotechnology Information) GenBank database (
www.ncbi.nlm.nih.gov ) using BLASTn tool[38]. Nucleotide sequences were aligned using ClustalW within the MEGA 7 platform. A neighbor-joining phylogenetic tree was constructed using the p-distance model and tested with 1000 bootstrap replication within the MEGA 7 platform[39].NCBI accession numbers
-
Sequences obtained previously were deposited in the NCBI database. Below is a list of all accession codes presented next to the correspondent strain:
Isolate 8: OQ145681
Isolate 10: OQ145686
Isolate 12: OQ145682
Isolate 15: OQ145683
Isolate 21: OQ145684
Isolate 23: OQ145685
Isolate 24: OQ145687
Arsenic tolerance screening of bacterial
isolates -
Arsenic tolerance of isolates was assessed by evaluating growth inhibition in the presence of different levels of As. For each isolate, a cell suspension was prepared in sterile water and 30 µL of a pre-washed re-suspended aliquot (106 cells mL−1) was inoculated in 5 mL of yeast-mannitol (YM) liquid medium containing increasing concentrations of arsenic(III) chloride (AsCl3) (5, 10, 50 and 200 mg of As mL−1). Tubes were kept at 28 ºC on an orbital shaker at 100-rev min−1 for 72 h. Isolate growth was evaluated by a once per day measurement of the optical density at 600 nm. Two replicas for each strain and each concentration were carried out. Finally, the growth of the bacterial isolates was classified into three groups according to the percentage of growth inhibition: sensitive (100%−80%), moderately tolerant (80%−60%) and tolerant (< 60%). Cell suspensions prepared without the addition of As were also included as controls for each isolate.
Effects of arsenic in the symbiosis
-
Lotus uliginosus cv. Sunrise seeds were surface sterilized and rinsed extensively with sterile distilled water according to[30]. Next, seeds were hydrated for 1−2 h in sterilized water and moved to 0.8% w v−1 agar-water plates for 1−2 days until germination. The pre-germinated seeds were moved to slants containing 50 mL of N-free Jensen plant medium[40]. Seedlings were allowed to establish in this nutrient medium before addition of bacterial inocula and As treatments. Each isolate was inoculated by applying 1 mL of bacterial suspension (approximately 108 bacterial cells in 1/4 Jensen medium) on the roots of each seedling (3 replicates per isolate). Arsenic was added to liquid Jensen medium (1/4 diluted) as AsCl3, in order to produce concentrations of 0.5, 5, 10, 20, 100 and 200 mg·mL−1 of As. Plants were supplemented with each As concentration (three replicates per each treatment). Furthermore, controls only with nitrogen (TN) and without nitrogen and not inoculated (T0) were prepared as described by Soares et al. [22]. Plants were incubated for six weeks in a controlled-environment chamber with 16 h light/8 h dark cycle at 23 ºC (day)/18 ºC (night). Plants were observed for nodulation after three and six weeks, and were harvested after six weeks of growth. The colour of the interior of the nodules was also observed and correlated with the presence/absence of leghemoglobin depending on whether they were pink or not, i.e., root nodules coloured pink by leghaemoglobin are caused by a nitrogen-fixing symbiotic relationship between the plant and beneficial bacteria (rhizobia)[41].
Finally, the shoots were dried at 80 ºC for two days. The dry weight (aerial biomass) of inoculated plants (X) was used to determine the index of effectiveness (Es) as described by Ferreira & Marques[42]. TN and T0 represent the dry weight of plants from nitrogen control and from non-inoculated plants, respectively:
$ =\frac{X-T0}{TN-T0}\times 100{\text%} $ Statistical analysis
-
Data were analysed by one-way analysis of variance (ANOVA) with software STATISTICA version 10 (StatSoft), using the Tukey’s honestly significance difference (HSD) test at P ≤ 0.05.
-
In order to address the objective of this research, we isolated bacterial strains from L. uliginosus growing under field conditions in an arsenic contaminated site in the central region of Portugal. In this context, 22 isolates were obtained from root nodules of L. uliginosus. All isolates were slow growers, showed little congo red absorption, and had an alkaline reaction on BTB, indicated by a blue colour, which is usually produced by Bradyrhizobium spp.[30] (Supplemental Fig. S2). Congo red is often incorporated in culture media for isolating rhizobia or for testing the purity of rhizobia cultures. Rhizobia typically do not absorb congo red or absorb it weakly, while other bacteria absorb it strongly.
The assessment of genotypic diversity of the natural population nodulating L. uliginosus was achieved by ERIC-PCR. The analysis of the fingerprinting patterns of each isolate showed the existence of several clusters (Fig. 1). This dendrogram was used to determine the similarities among isolates. Results showed the presence of a high genetic diversity in the population, despite the high contamination by As in the original soil. These isolates showed multiple fingerprinting patterns and no single dominant genotype was apparent from our results.

Figure 1.
Dendrogram showing the diversity of root nodule bacteria (Bradyrhizobium sp.) isolated from L. uliginosus, derived from ERIC-PCR fingerprints using UPGMA method, at 85% similarity.
Arsenic tolerance
-
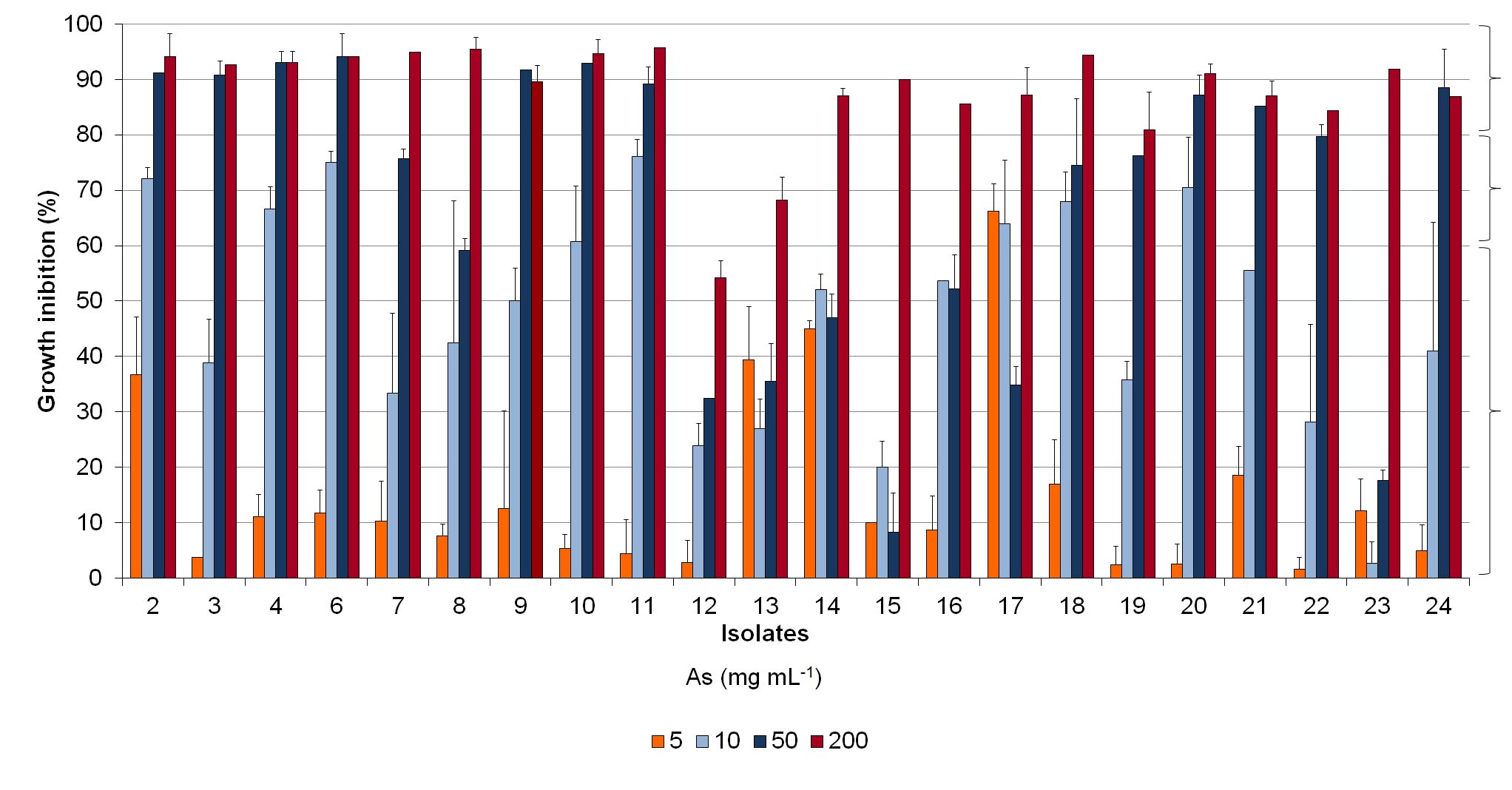
Results obtained for As tolerance after 72 h (Fig. 2, Supplemental Fig. S3 and Table 1) showed that, for the highest concentration tested (200 mg of As mL−1), most isolates were considered As sensitive (percentage of growth inhibition (GI) of 80%−100%). Only two isolates, 13 and 12, were considered as moderately tolerant (GI 60%−80%) and tolerant (GI < 60%), respectively. On the other hand, for 50 mg·mL−1 of As about 36% of isolates (isolates 8, 12, 13, 14, 15, 16, 17 and 23) were tolerant, and 18% were considered moderately tolerant (isolates 7, 18, 19 and 22). However, an inverse situation was verified for 10 and 5 mg·mL−1 of As, where no isolates were found sensitive. While for 10 mg·mL−1 64% of the isolates were tolerant, for the lowest concentration of As used, 5 mg·mL−1, a large majority of isolates (95%) was considered as tolerant.
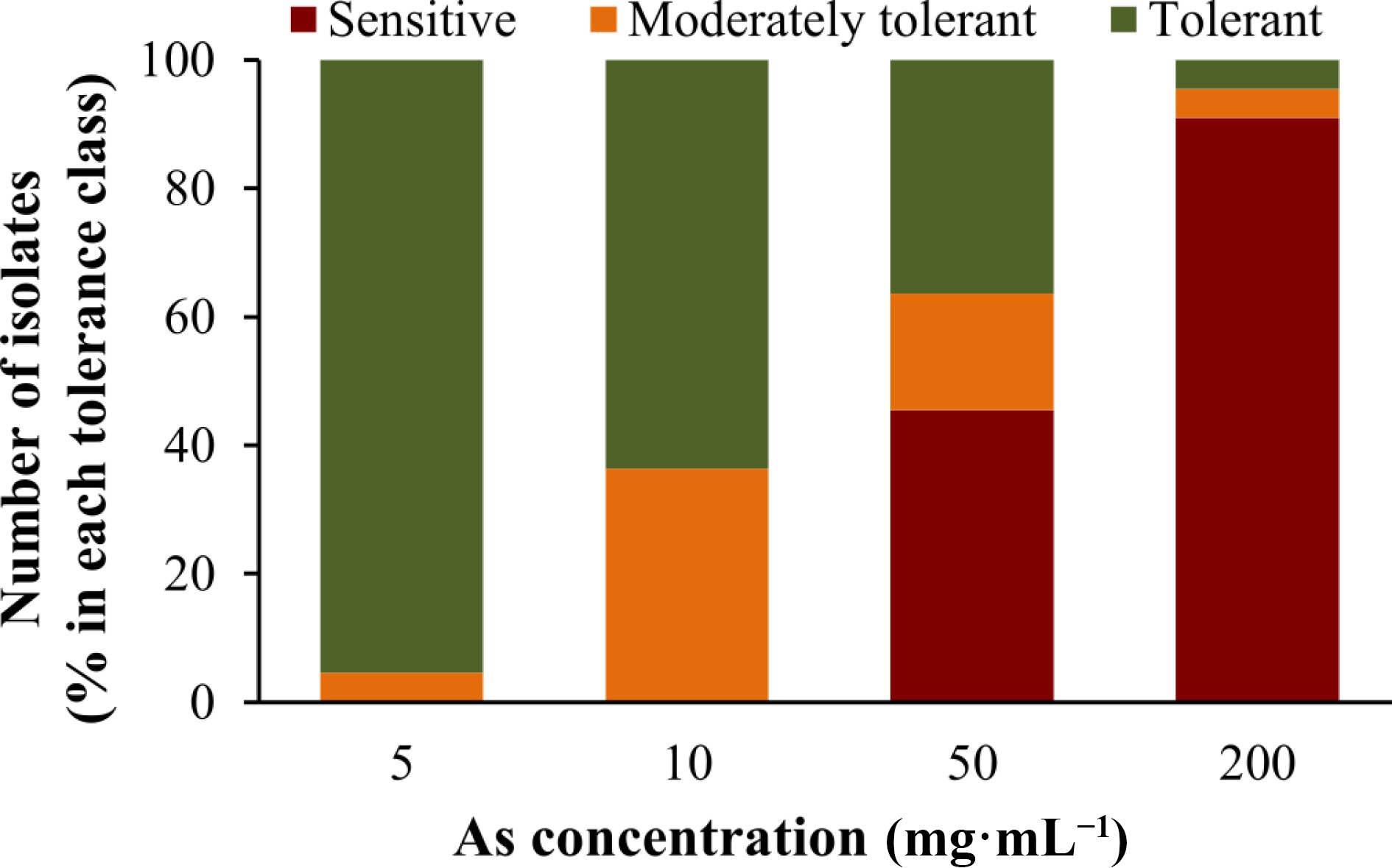
Figure 2.
Tolerance of the L. uliginosus isolates to different As concentrations (5, 10, 50 and 200 mg·mL−1). Isolates were classified according to the percentage of growth inhibition relative to controls grown in the absence of As, being considered sensitive (80%−100%), moderately tolerant (60%−80%) and tolerant (< 60%). Stacked-columns indicate the percentage of isolates in the three tolerance classes for each As concentration.
Table 1. Nodulation phenotype of L. uliginosus plants inoculated with the different isolates upon different As concentrations.
Isolates As concentration (mg·mL−1) 0 0.5 5 10 20 3 w 6 w 3 w 6 w 3 w 6 w 3 w 6 w 3 w 6 w 8 + + + + + + + + − − + + + + + + + + − − + + + + + + + + − − 10 + + − ± − ± − − − − + + − ± − ± − − − − + + − ± − ± − − − − 12 + + + + + + ± ± − − + + + + + + ± ± − − + + + + + + ± ± − − 15 + + − + − + + + − + + + + + + + + + − + + + + + + + + + − + 21 + + − + + + − + − + + + + + + + − + − − + + + + + + − + − − 23 + + − + − + − + − + + + − + − + + + − + + + + + + + + + − − 24 + + + + − + − + − − + + + + − + − + − − + + − − − + − + − − w, incubation weeks; +, presence of nodules; −, absence of nodules; ±, presence of small and white nodules. Effects of arsenic on the symbiosis
-
For the evaluation of the effects of As on the symbiosis with L. uliginosus, seven isolates (namely 8, 10, 12, 15, 21, 23 and 24) belonging to different clusters (Fig. 1) and with different levels of tolerance to As (Supplemental Fig. S3) were chosen to inoculate L. uliginosus cv. Sunrise seedlings. Assays were performed with six As concentrations (0.5, 5, 10, 20, 100 and 200 mg of As mL−1) and results were recorded three and six weeks after the addition of bacterial inocula and As to the seedlings. Briefly, plants did not tolerate the highest As concentrations used, 100 and 200 mg·mL−1, and died one week after As addition. Arsenic toxicity affected the symbiosis and the different isolates also showed different symbiotic performances as the arsenic concentration increased up to 20 mg·mL−1 (Fig. 3).
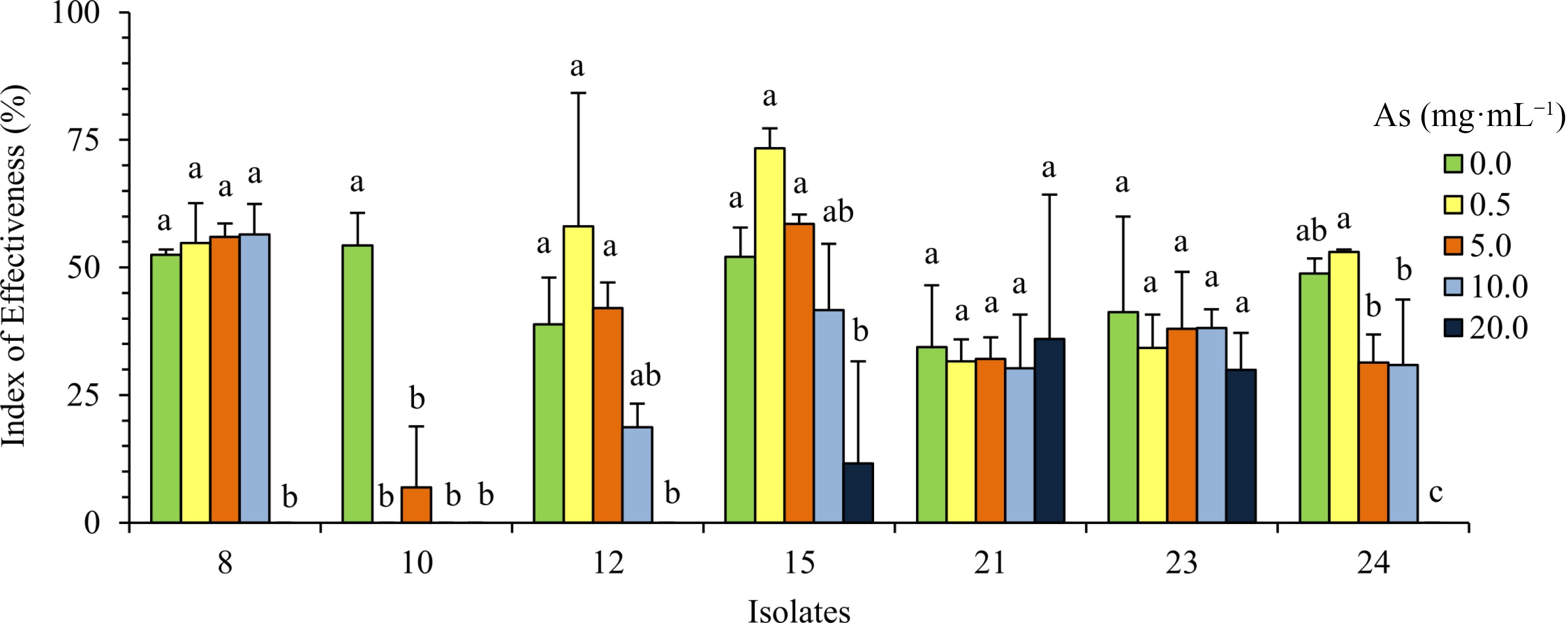
Figure 3.
Effect of As on the symbiosis. Values represent the average of the index of effectiveness (Es) ± SD of L. uliginosus plants inoculated with the bacterial isolates and grown with different As concentrations (0, 0.5, 5, 10 and 20 mg·mL−1). Different letters express significant differences between plants inoculated with each isolate at several As concentrations according to Tukey's HSD test at at P < 0.05.
One of the effects of As in the symbiosis was demonstrated by the delay in nodulation (Table 1), especially for higher concentrations of As tested, when compared to the controls without the addition of As. A more drastic effect was the impairment of the symbiosis by the lack of nodulation. For the highest As concentration allowing plant survival, 20 mg·mL−1, only plants inoculated with isolates 15, 21 and 23 presented pink root nodules (i.e., functional) at least in one of the replicates. In this case, nodulation was delayed, but the symbiosis remained effective in some replicates, depending on the isolate tested. Plants inoculated with isolates 8 and 12 did not show delayed nodulation but nodules were only formed at As concentrations up to 10 mg·mL−1. However, at this concentration plants inoculated with isolate 12 had small and white nodules (i.e., ineffective). Nodulation was also observed (with the presence of pink nodules) in plants inoculated with isolate 24 until 10 mg of As mL−1 but with a delay of 3 weeks. Plants inoculated with isolate 10 and growing with 0.5 and 5 mg of As mL−1 showed also a delay in nodulation and nodules were small and white. This nodulation phenotype indicates an ineffective symbiosis, inversely to what happened with plants grown without the addition of As which had pink nodules.
These plant-inoculation experiments using several As concentrations were also performed to evaluate the symbiotic effectiveness, by determining the shoot dry weight of L. uliginosus plants after 6 weeks of growth. The As concentrations used in these experiments were higher than those usually present in groundwater used for irrigation and in soils in various countries, e.g. 3,10 µg·L−1 and 22 mg·kg−1, respectively, in Argentina, or 3,700 µg·L−1 and 196 mg·kg−1, respectively, in India[43]. All the isolates chosen for these experiments were considered effective in the absence of As and the respective indices of effectiveness (Es), under these conditions, ranged between 35% and 55% (Fig. 3). These isolates also showed different levels of symbiotic performance and indices of effectiveness upon different As concentrations. In general, results were congruent with the observed nodulation phenotypes (Table 1). Isolates 15, 21 and 23 were able to establish an efficient symbiosis with the host plant at 20 mg of As mL−1. Plants inoculated with isolate 15 showed an increase in the aerial part and the consequent increase in the indices of effectiveness at the concentration of 0.5 mg As mL−1, which was near 75% and the highest Es of these experiments. This strain was therefore considered highly effective in the presence of As (0.5 mg·mL−1). However, for the other two isolates, 21 and 23 respectively, no significant differences were found between the various treatments, including the control without As addition, and their respective indices of effectiveness were lower but still considered as effective in nitrogen fixation. On the other hand, isolates 8, and 24, were able to establish an efficient symbiosis until the concentration of 10 mg of As mL−1. For the first isolate, no significant differences were found between the various treatments (0.5, 5 and 10 mg As mL−1) including the control without addition of As. The indices of effectiveness of plants inoculated with isolate 12 and with 0.5 and 5 mg of As mL−1 did not show significant differences between each other and the control (without As addition). However, for 10 mg·mL−1 of As the index of effectiveness was very low (19%) and the symbiosis was considered as ineffective, the plants presenting small and white nodules as previously mentioned. Lastly, plants inoculated with isolate 10 had a very weak performance in the presence of As (with plants having small and white nodules) and were only able to establish an efficient symbiosis when As was completely absent, showing indices of effectiveness significantly different from all the remaining treatments.
Interestingly, among the isolates that were able to establish an efficient symbiosis while sustaining the highest As concentration tolerated by plants, 20 mg·mL−1 (isolates 15, 21 and 23), isolate 15 was also among those with higher indices of effectiveness in the absence of As. Moreover, this isolate had the highest Es shown, near 75% at 0.5 mg·mL−1 of As being considered as highly effective in nitrogen fixation.
16S rRNA region phylogeny
-
Aligned sequences of the partial 16S rRNA region were used to construct the phylogenetic tree shown in Fig. 4. The isolates from this study were all clustered with Bradyrhizobium spp. Sequences obtained from isolates 8, 10, 15, 23 and 24 shared 100% sequence identity and the closest strains were Bradyrhizobium spp. isolated from Lotus uliginosus (Bradyrhizobium sp. 3LBC, 8LBI, SEMIA 839 and NZP2309[21,28]), Cytisus triflorus (Bradyrhizobium sp. CTS12), Cytisus scoparius (Bradyrhizobium genosp. AD Cs6020[44]) Vigna unguiculata L. (Bradyrhizobium sp. VUPMI37) and Ulex europaeus (Bradyrhizobium sp. ICMP 12674[45]). Isolates 12 and 21 shared 100% sequence identity and the closest strains were Bradyrhizobium spp. isolated from Glycine max (Bradyrhizobium japonicum GI-4 and J5 and Bradyrhizobium sp. 323S2[46,47]), Pigeonpea (Bradyrhizobium sp. RP6) and Erythrina brucei (Bradyrhizobium shewense ERR11T [48]).

Figure 4.
Phylogeny of the partial 16S rRNA gene with a total of 1189 aligned positions. Confidence bootstrap values are presented near each node. NCBI GenBank accession codes are presented next to each strain. Isolates obtained in this work are in bold. Rhizobium leguminosarum USDA 2370T (U29386) was selected as an outgroup.
-
Biological nitrogen fixation, including the contribution made by symbioses in the root nodules of legumes, supplies a large proportion of the nitrogen that increases soil fertility in natural and agro-ecosystems[49]. In recent years, the selection of symbiotic or free-living plant growth promoting rhizobacteria with remediation abilities has gained prominence, since such strains could help plants to grow in polluted soils or could even limit the inclusion of contaminants into plant tissues[50]. In particular, the rhizobium-legume symbiotic interaction has been highlighted as a promising tool to be used for bioremediation of As and heavy metals in soils[51−55].
It has been reported by several authors that associations between plants and microorganisms increase the bioremediation potential of plants[56−58]. In this study we found a large diversity of root nodule bacteria isolated from L. uliginosus growing in soils contaminated with As in Portugal. These results are different from previous data indicating a lack of genetic diversity in the population of Rhizobium isolated from Trifolium sp. in an analogous area contaminated by heavy metals[59,60]. However, similar results have been reported by Carrasco et al.[51] and also by Delorme et al.[61], who observed that the presence of rhizobial genotypes in soils with high heavy metal concentration were not different from those existing in soils with low metal levels. These authors showed that the rhizobial population was very diverse and the presence of heavy metals did not lead to a decrease in diversity. According to Rangel et al.[62], a wide diversity of bacteria resistant to As was verified, including several rhizobial genera such as Azorhizobium, Mesorhizobium, Rhizobium, Burkholderia (now either in Paraburkholderia or Trinickia)[63]. Therefore, biological nitrogen fixation has a great potential to be used in the future for phytostabilization purposes, given the high number of host legume species of the mentioned rhizobia that can be tested in the field and also because legumes accumulate heavy metals mainly in roots and show a low level of metal translocation to the shoot[55].
In this study, we have isolated bacteria from root nodules of L. uliginosus growing in soil contaminated by arsenic. These bacteria were morphologically characterized, by growth rate and reaction on BTB, as identical to those strains belonging to the genus Bradyrhizobium. Moreover, L. uliginosus root nodule bacteria seem to be constrained to B. japonicum and Bradyrhizobium sp. symbionts, rarely harbouring endophytic bacteria[27,28]. These results were confirmed for a set of seven isolates (namely, 8, 10, 12, 15, 21, 23 and 24) used to test the effects of As in the symbiosis, which were molecularly identified as Bradyrhizobium spp., using 16S rRNA region sequencing. A detailed phylogenetic analysis comprising appropriate molecular markers[22] could reveal if these isolates represent novel species or described ones, and compare them with the ones already described taxonomically from L. uliginosus that did not originate from As-contaminated soils[18, 24].
In the tests performed to assess As tolerance, it was found that all the isolates were able to grow at concentrations of 5 and 10 mg As mL−1 revealing higher tolerance than observed by Deepika et al.[64] for Rhizobium radiobacter (strain VBCK1062) when using 10 mM As in YM liquid medium.
In the tests carried out with L. uliginosus plants, the effects of As on nodulation and growth were recorded three and six weeks after seeding. The number of nodules and growth of the plants were affected by the presence of As in the medium, producing a severe effect in the early stage of nodulation. This could be due to the effect of As on rhizobia survival and initial tolerance[65]. The negative effects of As in the nodulation process found in our study are similar to those described by several authors. In fact, Reichman[66] observed that treatment with As induced a significant decrease in the total and effective number of nodules in soybean. Also, Pajuelo et al.[67] found important proof of damage by As on roots as well as a reduction of root hair number, which was associated with a decrease of around 90% in rhizobial infection number in plants of Medicago sativa. Moreover, the reduction in nodulation observed in legumes exposed to other metals, such as Cu2+, Cd2+ and Hg2+, was also attributed to the damage of metals on root hairs[68]. On the other hand, Bustingorri et al.[69] demonstrated that As causes membrane damage and decreases the chlorophyll content. This metalloid has no known biological function and fundamentally disrupts the cell growth and metabolism of living cells[64]. The adaptive tolerance response occurring in some rhizobial strains to the perturbation caused by As contamination, was also reported by others[47]. Our results confirmed that some Bradyrhizobium isolates tested for the evaluation of the effects of As in the symbiosis showed capacity to establish an efficient symbiosis, even in the presence of the highest As concentration (20 mg·mL−1), mainly isolate 15. This isolate had high effectiveness when compared to the As-free inoculated controls and to the TN and T0 controls.
Biological nitrogen fixation, including the contribution made by symbioses with legumes, supplies a large proportion of the nitrogen that increases soil fertility in natural and agro-ecosystems[49]. In recent years, the selection of symbiotic or free-living plant growth promoting rhizobacteria with remediation abilities has gained prominence, since such strains could help plants to grow in polluted soils or could even limit the inclusion of contaminants into plant tissues[50]. In particular, the Rhizobium-legume symbiotic interaction has been highlighted as a promising tool to be used for bioremediation of As and heavy metals in soils[51−54]. In this study, we have isolated bacteria from L. uliginosus with the capacity to establish an efficient symbiosis, even in the presence of high concentrations of As.
We consider Bradyrhizobium isolate 15, as the most promising to be tested in situ for their applicability for phytoremediation in sites polluted by this metalloid.
-
The main impact of this work is the possible use of autochthonous legume plants and their micro-symbionts, such as the symbioses with L. uliginosus and Bradyrhizobium, for the phytostabilization of contaminated soils, helping its fertilization. Results reveal that root nodule bacteria isolated from L. uliginosus growing in polluted soils, mainly those tolerant to the highest concentration of arsenic, can be symbiotically effective upon high As concentrations. This can be a very interesting aspect and can be used in the future, in bioremediation experiments on contaminated soils using native legumes, since it could have a positive ecological impact in those sites. We consider that the dual function of As bioremediation plus soil nitrogen enhancement can be achieved by effective symbiotic nodulation in affected As-metal-soils. Such a tolerant and functional symbiosis can support vegetation cover, stabilizing As-contaminated soils, and consequently it could be a smart practice for phytostabilization.
This work was supported by the cooperation project between Portugal and Argentina: 'The genus Lotus and their utilization for the restoration of soils contaminated with heavy metals. The biochemistry and their symbionts', FCT/DREBM 00264, Proc. 4.1.3 and also by the European project: 'Raising the bio-based industrial feedstock capacity of marginal Lands (Margin Up)', nº101082089.
-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Schematic representation of the experimental approach used in the isolation and characterization of root nodule bacteria from Lotus uliginosus growing in arsenic contaminated soils.
- Supplemental Fig. S2 Bromothymol blue pH-reaction by the different isolated strains from Lotus uliginosus.
- Supplemental Fig. S3 Tolerance of bacteria isolated from Lotus uliginosus to As. Columns indicate the percentage of growth inhibition ± SD after 72h incubation in liquid YM medium supplemented with different concentrations of As (5, 10, 50 and 200 mg mL-1), relative to cultures grown in the absence of As.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Soares R, Fareleira P, Colavolpe B, Ruiz OA, Videira e Castro I. 2023. Root nodule bacteria isolated from Lotus uliginosus for future use in phytostabilization of arsenic contaminated soils. Grass Research 3:8 doi: 10.48130/GR-2023-0008 

Root nodule bacteria isolated from Lotus uliginosus for future use in phytostabilization of arsenic contaminated soils
- Received: 12 January 2023
- Accepted: 20 April 2023
- Published online: 10 May 2023
Abstract: In recent decades there has been growing concern around heavy metals and metalloid contamination in soil. Arsenic (As) is a ubiquitous trace metalloid. The high levels of this metalloid in soils are a consequence of human activities and also from natural inputs. In general, the biodiversity of microorganisms and plants decreases drastically in contaminated soils. The knowledge that some leguminous plants, mainly certain species of Lotus, are growing well in such soils has attracted our attention for studying symbioses that are well adapted to harsh conditions. In this work we studied the rhizobial population existing in the root nodules of native Lotus uliginosus Sch. growing in a central region of Portugal. This legume grows in soils particularly affected by As due the discharge of industrial liquid effluents from fertilizer and chemical facilities. Diversity and tolerance to different concentrations of As of root nodule bacteria were studied. Our results showed that the symbioses between L. uliginosus and As tolerant Bradyrhizobium isolates were efficient when a nutrient medium containing high As concentrations was used. The present work highlights the capacity of L. uliginosus to grow and establish nitrogen-fixing symbioses in soils strongly contaminated with As and its potential for future use to promote vegetation cover to stabilize As contaminated soils.
{kind=link}
{kind=link}
{kind=link}