








-
Dragon fruit is a perennial climbing vine from the Selenicereus genus of the Cactaceae family, which originated in Mexico and Central America[1]. The relatively mature large-scale planting techniques and the early high returns on investment have prompted the rapid development of China's dragon fruit industry. The planting area expanded from 2,000 in 2010 to 70,000 hm2 in 2021, and the output increased from 36,000 tons in 2010 to 1,600,000 tons in 2021. China now ranks first in the world in both planting area and output of dragon fruit, with an annual output value exceeding 10 billion RMB. The proportion of dragon fruit in tropical fruit production also rose from 0.15% in 2010 to 5.3% in 2020, making it the fifth major South Asian subtropical horticultural fruit after lychee, longan, banana, and mango in China.
The rapid expansion of planting areas has exposed many problems, such as a limited number of varieties, few cold-resistant and pest-resistant varieties, a lack of deep-processed products, and insufficient development of high-value-added products, which severely restrict the healthy and sustainable development of the dragon fruit industry. Therefore, based on the existing domestic dragon fruit germplasm resources, conducting in-depth research on dragon fruit genomics, population genetics and functional genomics are of great significance for developing modern breeding techniques and accelerating the cultivation of new dragon fruit varieties. This review elaborates on the recent research progress of dragon fruit in terms of germplasm resources, genomics, and biological characteristics, hoping to provide some references for subsequent research and breeding of dragon fruit.
-
Dragon fruit is world-renowned for its large, night-blooming flowers, known as the 'Moonlight Cactus', and also commonly referred to as the 'Overlord Flower' by the Chinese. The dragon fruit flower is a typical nocturnal flower that blooms for only one night and wilts the next day. The flower is not only large and beautiful but also edible, with high nutritional value[2]. The stamens and pistils of the dragon fruit flower are of equal or shorter length, and under natural conditions, the dragon fruit exhibits a certain degree of self-incompatibility, leading to a low self-pollination rate[3,4].
Dragon fruit originates from tropical rainforests and prefers to grow in warm, humid, and well-lit environments, climbing on tree trunks or rocks with an optimal growth temperature of 25−35 °C. As a species from the cactus family, dragon fruit exhibits high tolerance to heat, drought, and shade, being light-loving and thriving in both fertile and poor soils[5]. Commercially grown dragon fruits often have triangular fleshy stems, which are the main sites for photosynthesis. The leaves are modified into short spines located in the notches along the wavy edges of the stems.
Dragon fruit typically has a vibrant red or yellow peel and white, red, or violet-red flesh, which are mainly derived from a natural pigment called betalains. As a low-calorie and low-fat fruit, the main sugar component in dragon fruit flesh is glucose[6], it is also rich in dietary fiber, vitamins, various minerals, and high levels of plant albumins and amino acids[7−9]. The fruit is densely packed with numerous black seeds containing unsaturated fatty acids such as linoleic acid, linolenic acid, palmitoleic acid, arachidic acid, and stearic acid[10]. Besides being edible, the flower, fruit, and stem of the dragon fruit can be used medicinally, exhibiting biological activities such as antiproliferative, antimicrobial[11], lipid-lowering, and antioxidant properties[12], and significant anticancer and hypertension-preventive effects, demonstrating great medicinal potential[11].
-
The systematic nomenclature of dragon fruit was once very chaotic, with the same species having different species names. Therefore, it is necessary to clarify the common species names of dragon fruit. Originally, dragon fruit were considered belonging to two genera, Hylocereus and Selenicereus. In 2017, molecular phylogenetic studies showed that the species of the former Hylocereus were nested within the species of Selenicereus, thus merging the two genera into one, now known as Selenicereus[1]. After the merger, Selenicereus contains a total of 33 species (
https://wfoplantlist.org ), with the four most common species used for large-scale commercial cultivation being S. undatus (formerly H. undatus) with red peel and white flesh (Fig. 1a), S. costaricensis (formerly H. costaricensis) with dark magenta peel, and violet-red flesh (Fig. 1b), S. monacanthus (formerly H. monacanthus, also known as H. polyrhizus) with crimson peel and flesh (Fig. 1c), and S. megalanthus (H. megalanthus and S. megalanthus are used interchangeably, now classified as the same species) with yellow peel and white flesh
(Fig. 1d)[1,13,14]. Other varieties, such as the thornless yellow dragon fruit, known abroad as Aussie Gold, have yellow peel (scaly, smooth, and thornless) and white flesh, but they do not belong to S. megalanthus and are more like a mutant variety of S. undatus. Dragon fruit varieties that have pink flesh, or bicolor varieties are usually the hybrids of S. undatus and S. costaricensis.
Figure 1.
The phenotype of bud, flower and fruit of four main commercial dragon fruits. (a) S. undatus has green buds; big, funnel-shaped, and white nocturnal flowers; and oblong fruits with pink peel, and white flesh and covered with green, long, and hard scales. (b) S. costaricensis has green buds along with lighter red edges on the outer perianth; big, funnel-shaped and white nocturnal flowers; and ovoid fruit with dark magenta peel and violet-red flesh, and covered with red, soft and short scales. (c) S. monacanthus has green buds along with bright red edges on the outer perianth; big funnel-shaped and white nocturnal flowers; and ovoid fruit with crimson peel and flesh covered with red, soft and short scales. (d) S. megalanthus has green buds along with dark brown outer perianth; big funnel-shaped (rounder than other species) and white (a little yellow) nocturnal flowers; and ovoid fruit with yellow peel and white (transparent) flesh covered with thorns on tuberculate skin instead of scales.
In China, the most widely cultivated varieties are red-flesh series, including 'Ruanzhidahong', 'Jindu 1', 'Guanhuahong' and 'Guihonglong 1', all of which belong to the species of S. costaricensis. On the other hand, white flesh cultivators like 'Vietnam 1' are introduced from Vietnam and belong to S. undatus, while 'Guanhuabai' and 'Baishuijing' are locally developed new varieties from S. undatus. Unlike earlier cultivated varieties, most commercially used varieties in recent years are self-compatible, which helps reduce labor costs. However, the genetic background of the most collected dragon fruit germplasms remains unknown. Therefore, to promote the breeding of new dragon fruit varieties, it is necessary to conduct detailed and in-depth research on the genetic background and phylogenetic relationships of dragon fruit germplasm resources.
-
The plant genome is the foundation for studying plant gene functions, population genetics, species evolution, and molecular breeding. Thus, the high-quality genome is the key to understanding the mechanisms behind various biological characteristics of plants. In 2021, two reference genomes of S. undatus were published, laying the foundation for in-depth research on dragon fruit[15,16]. The chromosomal-level genome of 'Guanhuabai' ('GHB') dragon fruit (2n = 2x = 22 chromosomes) was assembled using Pacbio, Hi-C, and 10× Genomics sequencing technologies, resulting in a 1.41 Gb genome with scaffold N50 of approximately 127.15 Mb[15]. A total of 27,753 protein-coding genes and 896.31 Mb of repetitive sequences were annotated. The study confirmed that dragon fruit suffered a whole-genome triplication event and a whole-genome duplication event, and inferred that dragon fruit diverged from the Caryophyllales order about 65.3 million years ago (MYA). Additionally, Chen et al. constructed a high-density genetic map based on the 'GHB' × Dahong F1 hybrid population and genetic maps for both parents, screening and identifying numerous SNP loci, providing essential data for subsequent dragon fruit breeding[15]. Based on the 'GHB' dragon fruit genome, Mou et al. discovered that the transcription factors HpDof1.7 and HpDof5.4 can bind to the promoters of HpSuSy1, HpINV2, HpTMT2, and HpSWEET14 to increase their activity, thereby enhancing the expression of the sugar metabolism pathway genes at the transcriptional level and promoting sugar accumulation in dragon fruit[17]. Chen et al. also found that the transcription factor HubHLH159 promotes betacyanin biosynthesis by activating the expression of HuADH1, HuYP76AD1-1, and HuDODA1[18]. Furthermore, Wang et al. preliminarily identified that HuS-RNase2, involved in post-pollination ubiquitination regulation is a candidate female development S-gene in the gametophytic self-incompatibility (GSI) system of dragon fruit[19].
Another version of the S. undatus genome (variety 'David Bowie') assembled a 1.33 Gb dragon fruit genome, with a scaffold N50 of approximately 109.7 Mb, but a contig N50 of only 31 Kb, and predicted 28,992 protein-coding genes[16]. This study estimated that dragon fruit diverged from the giant cereus around 9.18 MYA. They also found that genes related to betacyanin biosynthesis were co-located in a 12 Mb region on one chromosome, and speculated that this may enhance the efficiency of betacyanin synthesis.
Overall, the publication of these two genomes represents a breakthrough in the dragon fruit genome, providing crucial information for a better understanding of the biological characteristics and evolutionary history of dragon fruit, but the integrity of the assembled genomes is low. The genome of the main cultivated violet-red flesh dragon fruit (S. costaricensis) was not studied. Additionally, large-scale studies on population genomics, 3D genomics, and epigenomics, extensively conducted in other plants, have not yet been reported in dragon fruit. Thus, more comprehensive and in-depth studies research on dragon fruit genomics is necessary and urgent.
-
Plant pigments are a type of secondary metabolite produced during life activities, not only giving the plants brilliant flowers and vibrant fruits, but also participating in crucial biological processes such as plant growth and development, photosynthesis, and the maintenance of biodiversity. Moreover, plant pigments carry rich nutritional and biological functions, such as inducing insect pollination, assisting in seed dispersal, performing biological defense functions, and resisting adverse environmental factors[20]. Currently, natural plant pigments are mainly divided into four categories based on their chemical structures: lipid-soluble tetrapyrrole derivatives primarily including chlorophyll (Fig. 2a); lipid-soluble polyene pigments composed of isoprene units, primarily including carotenoids (Fig. 2b); water-soluble polyphenol pigments composed of polyphenol derivatives, represented by flavonoids (Fig. 2c) and anthocyanins (Fig. 2d); and water-soluble pyridine derivative pigments containing nitrogen heterocycles, represented by betalains (Fig. 2e)[21].
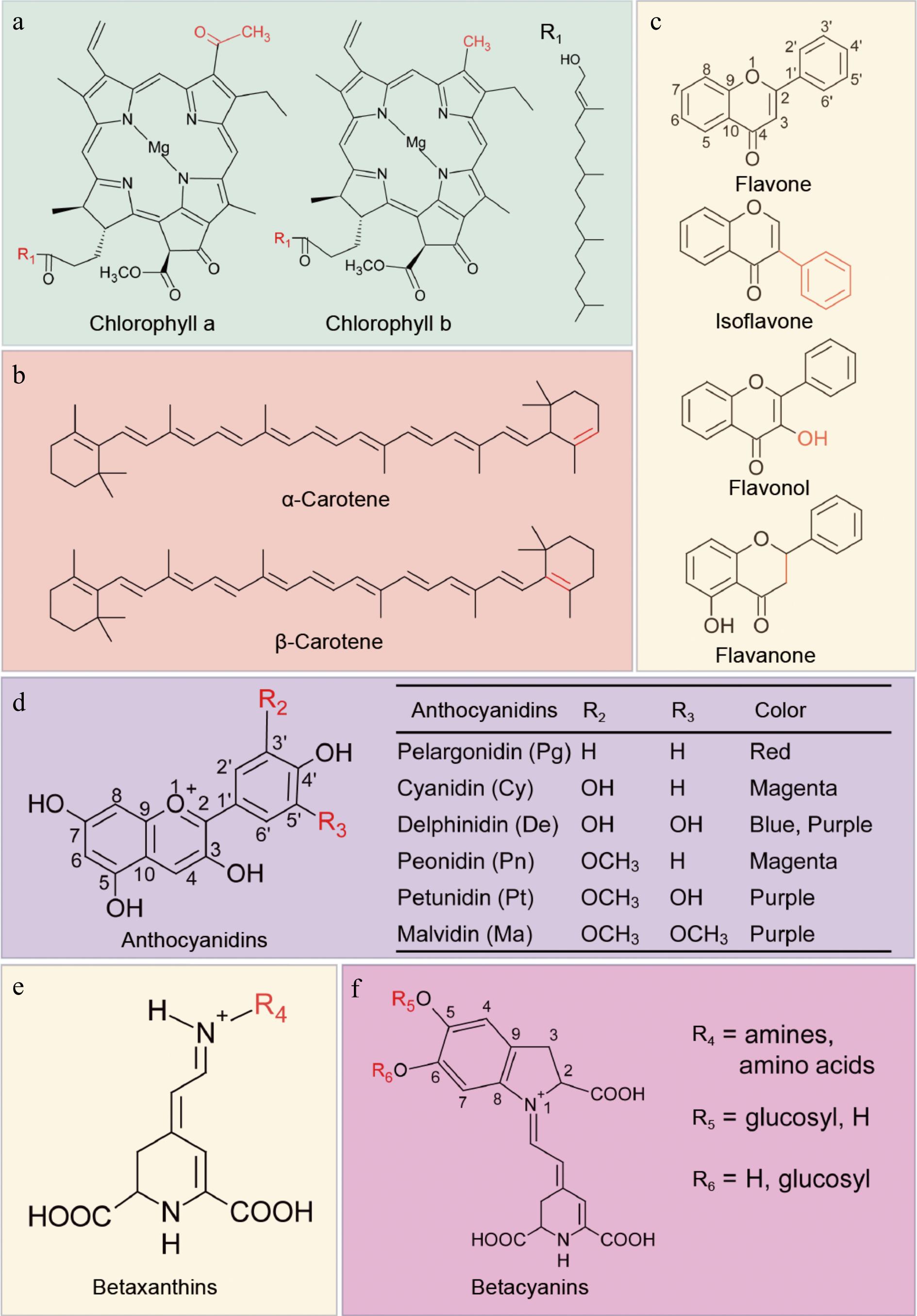
Figure 2.
Chemical structures of four natural plant pigments. (a) Chemical structures of chlorophyll a and b (green). Residue R1 is shown on the right. (b) Chemical structures of carotenes (orange). (c) Chemical structures of flavones and flavonols (yellow), flavone is the core structure for this class of substances. (d) Chemical structures of anthocyanidins. Residues R2, R3 and corresponding color for different type anthocyanidins are listed in the table. (e), (f) Chemical structures of betalains. Composition of residues R4, R5, R6 will produce kinds of betalains. Red groups in (a) − (c) represent the differences of pigments with similar structures.
In plants, betalains are hydrophilic nitrogen-containing pigments that exist only in the Caryophyllales order, and dragon fruit is the only edible fruit that contains amounts of betalains. Betalains are water-soluble tyrosine-derived alkaloid pigments, chemically known as trimethylamine betaine or trimethylglycine, with a chemical structure similar to methionine and choline, all belonging to the quaternary ammonium alkaloid class[22]. The content of betalains in dragon fruit is an important indicator and parameter determining the harvest time (color) and quality of the fruit[23]. As a natural pigment, betalains not only have the advantages of being non-toxic, edible, and highly safe but also have significant industrial value due to their unique physiological functions and medicinal value[24].
Physicochemical properties of betalains
-
Betalains, as a class of natural pigments, not only have advantages such as non-toxicity, edibility, and high safety but also possess significant industrial value due to their unique physiological functions and medicinal properties[25]. However, the stability of natural pigments is relatively poor, which increases the difficulty of their research and reutilization[26]. In dragon fruit, the stability of betalains, including betacyanins and betaxanthins is closely related to factors such as light, temperature, oxygen, pH value, enzymes, water activity, and various metal ions[27,28].
Betalains exhibit polar and ionizable physicochemical properties in aqueous solutions, so changes in pH affect their stability. Research has found that an appropriate amount of organic acids can enhance the stability of dragon fruit betalains. Acidic conditions (pH 5-6) facilitate the extraction and color protection of betacyanins, whereas under alkaline or slightly alkaline conditions, betalains can undergo irreversible fading due to oxidation-reduction reactions or isomerization reactions[29,30]. Under alkaline conditions, they can decompose into the precursor compound betalamic acid. The polarity of betalains also makes them insoluble in the most organic polar or semi-polar solvents, except for water and mixtures of low-molecular weight alcohols[31,32].
Additionally, the light and thermal stability of dragon fruit betalains is relatively poor. High temperatures can lead to the degradation of betacyanin, causing the pigment to fade, and even complete loss of color under extreme cases. In contrast to the slow degradation of pigments in darkness, exposure to light accelerates pigment decomposition[33−35]. Furthermore, metal ions also affect the stability of betalains. Complexation with metal ions can either darken or lighten the color of betalains. For instance, reductive metal ions such as Fe3+, Al3+, Cu2+, and Cr3+ can accelerate the degradation of betacyanins, leading to a lighter color[36−38]. Therefore, various factors affect the stability of betalain pigments, and it is important to manage these factors during processing, application, transportation, and storage to maintain the stability of betalain pigments.
The unique color presentation of betalains is derived from the conjugated diene of 1,7-diazaheptamethin, giving them the absorption peaks in both the UV and visible light regions[39,40]. Specifically, betacyanins have a maximum absorption value around 535−538 nm, while betaxanthins have a maximum absorption range of 460−480 nm[41−43]. Interestingly, at pH < 3, betalains exhibit a color shift effect in the 535−540 nm absorption range, with color deepening in the 575−650 nm range. As the pH increases, betalains may exhibit a bathochromic shift (red shift), where the absorption peak moves to longer wavelengths (lower energy) in alkaline conditions[44,45].
Distribution of betalains and its incompatibility to anthocyanins
-
Unlike the widely present anthocyanins and carotenoids in the plant kingdom, the distribution of betalains in nature is remarkably limited being found only in a few families of specific lineages. To date, betalains have been confirmed to be mainly present in two major lineages: the Basidiomycota phylum of fungi[46] and the Caryophyllales order of flowering plants[47]. Additionally, studies have found them in the diazotrophic bacteria (Gluconacetobacter diazotrophicus)[48]. Betalains serve as the primary pigments in core families of the Caryophyllales order, like Chenopodiaceae[49], Amaranthaceae[50], Cactaceae, and Nyctaginaceae[51,52]. They impart vivid colors to the flower, fruit, peel, and stem of these plants[53,54].
Interestingly, researchers found a mutual exclusivity between anthocyanins and betalains in core Caryophyllales plants, while anthocyanins and betalains cannot be coexisted in the same plants[55−57]. Currently, there is no consensus on the evolutionary reason behind this mutual exclusivity, but recent discoveries have shed some light on this phenomenon. For example: A high frequency of loss of the key late-stage anthocyanin synthesis gene TT19 has been observed in betalains-producing plants, and the recurrent loss of its homologs correlates with the transition to betalains pigment deposition[54,58]. Comparative analysis of DFR and ANS gene expression in betalains-producing and anthocyanin-producing plants revealed that the expression levels of DFR and ANS are nearly nonexistent in most organs and tissues of betalains-producing plants[55]. The R2R3 MYB-type regulator BvMYB1 in betalains-producing plants lacks the important conserved residues for interaction with bHLH, preventing interaction with the bHLH partners in anthocyanin-producing plants, thus inhibiting anthocyanin synthesis[59−61]. Although the key steps in betalains biosynthesis are well understood, the unique evolutionary patterns of betalains origin in the Caryophyllales and the molecular mechanisms underlying their mutual exclusivity with anthocyanins still require further elucidation.
Betalains classification
-
Betalamic acid, a unique chromophore in betalains pigments found in plants[62], accumulates primarily in vacuoles within plant cells, predominantly in the epidermis and subepidermal tissues of plants[63]. Betalamic acid can form different types of betalains pigments by conjugating with cyclo-DOPA derivatives, amino acids, and amines[64]. Based on the structural characteristics and photochemical properties of the pigment products formed, betalains can be divided into two main classes: red-violet betacyanins and yellow betaxanthins. Betacyanins are formed by the condensation of betalamic acid with cyclo-DOPA, while betaxanthins are derivatives formed by the binding of betalamic acid with various amines and amino acids[65].
Compared to betacyanins, research on betaxanthins is relatively limited. On one hand, betalamic acid can form different types of betaxanthins by binding with various amino acids. For example, the coloration of Portulaca grandiflora petals includes types such as portulacaxanthin-II (tyrosine-betaxanthin) and portulacaxanthin-III (glycine-betaxanthin)[66]. On the other hand, betalamic acid can bind with amines to produce different colors, such as the color changes observed in Mirabilis jalapa petals with miraxanthin-III (tyramine-betaxanthin) and miraxanthin-V (dopamine-betaxanthin)[67]. Furthermore, studies have shown that expressing genes related to betaxanthin biosynthesis through gene editing and petal-specific methods can enhance the vividness of ornamental plant colors, making it more aesthetically valuable compared to engineering flavonoid biosynthesis[68].
Betacyanins are primarily derived from various organs of plants such as beetroot[68], purslane fruits[51], globe amaranth[69], and quinoa grains[70]. As edible natural pigments, they find extensive applications in industries such as medical, healthcare, cosmetics, and food processing. Betacyanins can be further classified into seven types based on their chemical structure: betanin-type, amaranthin-type, oleracin-type, apiocactin-type, melocactin-/bougainvillein-r type, gomphrenin-type, and glabranin-/bougainvillein-v type (Fig. 3)[71,72]. Within these frameworks, combining different groups results in more specific color types, for instance, the gomphrenin type produces gomphrenin (gomphrenin I), globosin (gomphrenin II), basellin (gomphrenin III), and gandolin (gomphrenin IV) pigment types[73]. Similarly, amaranthin-types can further differentiate into amaranthin, sinapoyl-amaranthin, argentianin, and celosianin based on functional groups attached to their shared C5 framework[69,74−76]. Both betacyanins and betaxanthins can undergo various reactions such as glycosylation, acylation, decarboxylation, and isomerization on their basic frameworks to achieve different color effects ranging from yellow to purple in various plant tissues.
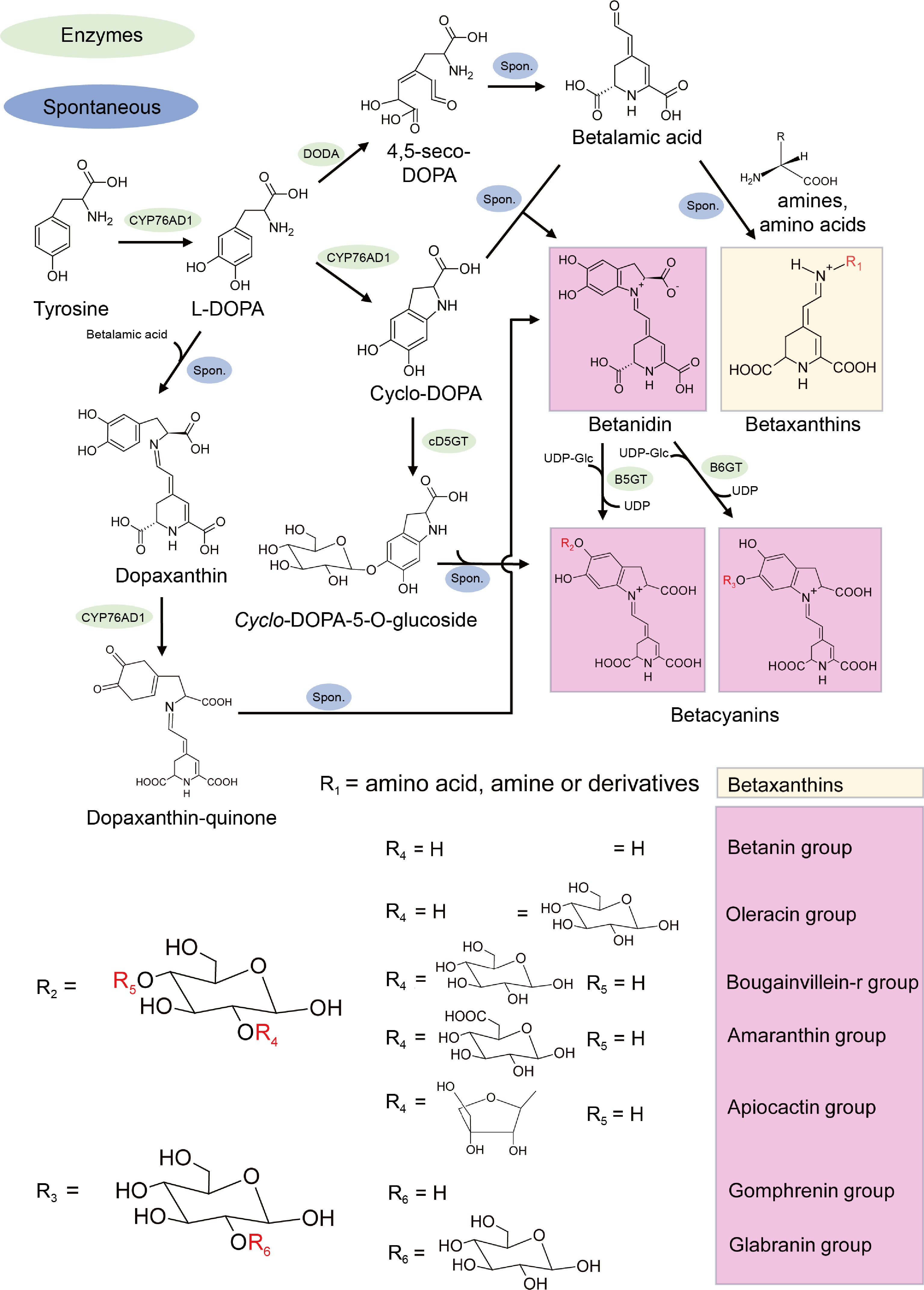
Figure 3.
Schematic pathway of betalains biosynthesis pathway. DODA, DOPA dioxygenase; cD5GT, cyclo-DOPA-5-O-glucosyltransferase; B5GT, betanidin-5-O-glucosyltransferase; B6GT, betanidin-6-Oglucosyltransferase; UDP, uridine-5'-diphosphate. Composition of residues R1-6 forms betaxanthins and seven type betacyanin pigments. The green color highlights the enzymes.
Betalains biosynthesis
-
Unlike most plants, the content of betacyanins in dragon fruit determines the coloration of both peel and pulp. Due to its high nutritional value, antioxidant capacity, and anti-inflammatory properties, the biosynthesis of betacyanins has become a significant research focus with economic value[77,78]. The biosynthesis of betalains in dragon fruit is as follows.
In most betacyanin-producing plants, arogenate serves as a precursor, undergoing decarboxylation under the action of ADH (arogenate dehydrogenase) to produce tyrosine[79]. Tyrosine undergoes hydroxylation catalyzed by cytochrome P450 enzyme CYP76AD1 to form L-DOPA (Fig. 3)[80]. L-DOPA undergoes various reactions. Firstly, under the action of DODA (4,5-dopa-extradiol-dioxygenase), L-DOPA is cleaved into the intermediate 4,5-seco-dopa, which then spontaneously forms betalamic acid (Fig. 3)[62]. Secondly, L-DOPA undergoes cyclization to form cyclo-DOPA under the catalysis of CYP76AD1 enzyme (Fig. 3)[81]. The synthesis of betacyanins can be divided into two main types. Betalamic acid can form betaxanthins by binding with different amino acids or amines (Fig. 3), or it can spontaneously form betanidin, a precursor of betacyanins, with cyclo-DOPA (Fig. 3). Cyclo-DOPA, under the action of cDOPA5GT (cD5GT, cyclo-DOPA-5-O-glucosyltransferase), produces cyclo-DOPA-5-O-glucoside, which can also spontaneously form betanin, another precursor of betacyanins. Betanin, after modification by B5GT (betanidin-5-O-glucosyltransferase) and B6GT (betanidin-6-Oglucosyltransferase), forms different types of betacyanins[82,83].
In summary, betacyanins can be modified and produced into various types of betacyanin pigments, such as glycosylation, acylation, decarboxylation, and isomerization. It is worth noting that decarboxylation primarily involves the degradation of betacyanin pigments and studies have shown the presence of degradation products of mono-, di-, and tri-carboxy betacyanin glycosides in extracts of red beets and dragon fruit[84,85]. Additionally, several transcription factors (TFs) such as HpWRKY44[86], HpERF1/2/3[87], HmoWRKY40[88], BvMYB1[61], and HuMYB1[89] are involved in the regulation of betacyanin biosynthesis. Reported genes and TFs involved in betalains biosynthesis are listed in Table 1.
Table 1. Reported genes involved in betalains biosynthesis.
Gene name Gene Id Gene function Species Ref. HuADH1 HU03G02979.1 Arogenate dehydrogenases S. undatus (H. undatus) [23] HuYP76AD1-1 HU03G00480.1 Cytochrome P450 enzyme S. undatus (H. undatus) [23] HuDODA1 HU03G01342.1 4,5-DOPA extradiol dioxygenase S. undatus (H. undatus) [23] HucDOPA5GT1 HU07G00239.1 cyclo-DOPA-5-glucosyltransferase S. undatus (H. undatus) [23] HucDOPA5GT2 HU03G00240.1 cyclo-DOPA-5-glucosyltransferase S. undatus (H. undatus) [23] HuMYB1 HU01G00040.1 MYB transcription factor, inhibiting the biosynthesis of betalains via suppressing the expression of HuADH1, HuCYP76AD1-1 and HuDODA1 S. undatus (H. undatus) [23] HubHLH159 HU11G01328.1 bHLH transcription factor, promoting betalains biosynthesis by activating the expression of HuADH1, HuCYP76AD1–1, and HuDODA1 S. undatus (H. undatus) [18] HpWRKY44 WRKY transcription factor, activates the expreesion of HpCytP450-like1, promoting the biosynthesis of betalains S. monacanthus
(Hylocereus polyrhizus)[86] HmoWRKY40 WRKY transcription factor, binding and activating the expression of HmoCYP76AD1 S. monacanthus
(Hylocereus monacanthus)[88] -
Plant self-incompatibility (SI) is a widely existing reproductive isolation mechanism among flowering plants, formed during their natural evolution to prevent inbreeding and species degeneration, thereby safeguarding genetic diversity[90]. SI can be traced back to over 90 MYA across numerous angiosperm taxa, with currently more than 100 plant families and approximately 40% of plant species employing SI to prevent self-pollination and promote outcrossing[91]. In recent years, SI has increasingly become a hot topic in disciplines such as plant molecular biology and crop breeding, with its applications extending to a growing number of economic crops.
Studies have shown that SI mechanisms exhibit both independence and diversity. Origin differences exist between SI genes of different genera, and corresponding incompatibility genes also demonstrate heterogeneity[92]. Broadly, based on floral morphology, SI can be categorized into two major types: heteromorphic SI and homomorphic SI. Heteromorphic SI (HSI) plants produce heterostylous flowers that can be further divided into distyly and tristyly types. Distyly type HSI was determined by a single locus, while tristyly is determined by two loci[93]. Homomorphic SI can further be classified into gametophytic self-incompatibility (GSI) and sporophytic self-incompatibility (SSI) based on differences in the genetic control of pollen self-incompatibility trait. Within the SSI mechanism, the male determinants are predominantly expressed in diploid anthers, resulting in haploid pollen grains showing a diploid S phenotype, prevalent in more than ten plant families such as Brassicaceae and Asteraceae, with extensive research in Brassica[94,95]. In contrast, most plants in nature exhibit GSI, found in families like Rutaceae, Rosaceae, Solanaceae, Papaveraceae, Plantaginaceae, Poaceae, Commelinaceae, and Ranunculaceae[96−100]. GSI, the most common form of SI, has been molecularly characterized by two distinct types of S genes and SI mechanisms. One type is the RNase-based SI, where the female determinant S-RNase and the male determinant F-box protein operate. In this mechanism, non-S-locus acts as a necessary factor, where under incompatible conditions, S-RNases enter self-pollen tubes to degrade pollen RNA through their inherent cytotoxic enzyme activity, thereby inhibiting pollen tube growth. The other GSI mechanism is calcium ion-based self-incompatibility, where the female determinant (PrsS) is a small secreted peptide and the male determinant (PrpS) is a transmembrane receptor. Interaction between PrsS and incompatible pollen triggers a signaling pathway, leading to rapid actin depolymerization, pollen tube inhibition, and programmed cell death, currently found exclusively in Papaveraceae[101−103].
Dragon fruit exhibits typical physiological structures that prevent self-pollination, such as protruding style and stamens shorter than the style, demonstrating self-incompatibility[104]. After self-pollination, pollen germination on the stigma is observed, but internal pollen tube growth is inhibited, illustrating typical GSI of dragon fruit[104,105]. Comparative analysis of GSI between white-fleshed (S. undatus) and red-fleshed (S. costaricensis) dragon fruit reveals stronger SI in red-fleshed varieties, while white-fleshed varieties exhibit relatively weaker SI[106]. Further studies indicate that intense SI in S. monacanthus suppresses pollen tube growth in the ovary, leading to fertilization failure, whereas SI between S. undatus and S. megalanthus is less pronounced[107]. Due to the impact of SI, dragon fruit exhibits low natural pollination and fruit set during cultivation, necessitating biological and artificial pollination methods, thereby increasing labor and cost resources in dragon fruit production, and hindering its commercial promotion[108]. Currently, the GSI mechanism of dragon fruit has not been extensively studied, and the related pollen S-RNase and S proteins have yet to be fully elucidated. Researchers analyzed the SI of dragon fruit varieties 'Kyoto 1' (self-compatible) and 'Cu Sha' (self-incompatible) and preliminarily identified HuS-RNase2 as a candidate gene in the GSI system, involved in the recognition process between pollen and stigma[19].
-
Dragon fruit, as the most economically valuable tropical edible fruit in the Cactaceae family possesses characteristics of low energy, high fiber, and high nutritional health value. Its unique appearance, rich nutritional composition, and health benefits have made it increasingly accepted and loved by consumers. In recent years, researchers have explored multiple areas related to dragon fruit. For instance, comparative metabolomic analysis has identified various organic compounds, active substances, and antioxidants in two main dragon fruit crops, S. undatus, and S. monacanthus[5,109]. Proteomics-based studies have elucidated the molecular mechanisms betalains biosynthesis during the white pulp and red pulp stages in S. monacanthus and the antioxidant signaling pathways in S. undatus[110,111]. Single-cell sequencing has revealed the developmental trajectory of dragon fruit peel during aging and identified early marker genes for fruit resistance[112,113]. Furthermore, numerous studies have reported on transcriptomics around growth, development, flowering mechanisms in Selenicereus species, betalains biosynthesis, responses to abiotic stress, antioxidative properties, and plant pathology mechanisms[78,114−118]. With technological advancements, there is potential for deeper genomic, population genetic, functional genomics, and metabolic studies on global dragon fruit germplasm resources[119]. This research aims to elucidate the molecular and evolutionary mechanisms underlying biological characteristics such as nocturnal flowering, self-incompatibility, betalains biosynthesis regulation, nutritional flavors, and others. These efforts will provide richer genetic resources and practical tools for precise dragon fruit breeding[120].
This work was supported by the Natural Science Foundation of Hainan Province (321RC1031) and Guangxi Key R&D Program Projects (2023AB17031).
-
The authors confirm contribution to the paper as follows: study conception and design: Ding Y, Lu G, Hu H; data collection: Li P, Li Z, Yao H; analysis and interpretation of results: Ding Y, Li P; manuscript preparation: Ding Y, Li P; manuscript midification: Ma X, Lu G, Hu H. All authors reviewed the results and approved the final version of the manuscript.
-
All data synthesized and analyzed during this review are available within the article.
-
The authors declare that they have no conflict of interest.
-
Received 20 August 2024; Accepted 17 October 2024; Published online 12 December 2024
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li P, Ma X, Li Z, Yao H, Lu G, et al. 2024. A review on the advances of dragon fruit. Tropical Plants 3: e041 doi: 10.48130/tp-0024-0041 

A review on the advances of dragon fruit
- Received: 20 August 2024
- Revised: 03 October 2024
- Accepted: 17 October 2024
- Published online: 12 December 2024
Abstract: Over ten years of rapid development, dragon fruit has become China's fifth major South Asian subtropical horticultural fruit, following lychee, longan, banana, and mango. As a climbing cactus, dragon fruit possesses lots of unique biological characteristics, such as blooming at night, being rich in betalains substances, and self-incompatibility. In recent years, with the publication of dragon fruit genomes, the molecular basis of forming dragon fruit's biological characteristics has been extensively studied and explored. This review gives a glimpse into the research progress on biological characteristics, germplasm resource classification, and genome sequencing of dragon fruit, especially focussing on the biosynthesis and characteristic of betalains and the mechanisms of self-incompatibility. A theoretical reference is provided for a comprehensive understanding of the genetic basis of important agronomic traits in dragon fruit and the development of new varieties.
-
Key words:
- Dragon fruit /
- Germplasm /
- Genome /
- Betalain /
- Self-incompatibility