










-
In recent decades, the demand for coffee has been continuously increasing, and the global consumption increased by 1%−2% every year[1]. According to the statistics of the International Coffee Organization, the annual market value of coffee is about 200 billion US dollars, and it is expected that future consumption will continue to drive the demand for coffee[2]. Coffee is a tropical plant, belonging to Rubiaceae. There are 124 species of coffee in the genus Coffea[3], but only two species − Arabica (C. arabica) and Robusta (C. canephora) are commercially traded in the international market. Arabica coffee grows well in a cool climate and at high altitude (1,000−2,100 m) while Robusta grows better in a hot and humid climate at lower altitude (100−1,000 m)[4]. At present, the main coffee production areas are located between 25° north latitude and 30° south latitude, among which Brazil, Indonesia, India, Uganda, and Vietnam are the main production areas worldwide[5].
Recently, many studies have been carried out on how the quality of coffee beans are affected by species, geographical origin, postharvest processing and production processing[4,6], Luca et al.[7] characterized the effects of different roasting conditions on coffee of different geographical origin by high performance liquid chromatograph (HPLC) with photo-diode array detector, near infrared spectrum instrument (NIRS) and stoichiometry, and concluded that different roasting conditions and geographical origin influenced the properties of roasted coffee beans. And the aroma and flavor of coffee beans has been widely studied, and many conclusions obtained. Kučera et al.[8] analyzed espresso coffee with different roasting degrees (light, medium, medium dark and dark) by ultra-performance liquid chromatography-tandem mass spectrometry. The obtained raw data were analyzed using multivariate statistics to assess the difference between each degree of baking. Cui et al.[9] found that coffee made from the coffee beans of two species exhibited a different, special flavor, as Arabica beans exhibited a 'baked' flavor and Robusta beans exhibited a 'sweet' flavor. The sensory properties, especially the flavor compounds of coffee beans are obviously influenced by production area[5]. In addition, roasting speed and roasting degree also have a significant effect on the aroma components of coffee beans[1,10].
As well as sensory properties, physicochemical properties of roasted coffee beans are also affected during the production procedure[11]. However, little research has focussed on the physicochemical properties, and at the same time excluded the influence of species, production area and roasting degree. Furthermore, most recent research has focused on the coffee beverage rather than coffee beans. This research chose two species of coffee beans (Arabica and Robusta beans) from five production areas (Brazil, India, Indonesia, Uganda and Vietnam), and roasted them to four different degrees (medium light, medium, medium dark and dark), to investigate the difference on physicochemical properties, such as the concentrations of protein, fat, organic acids, chlorogenic acid compounds (CACs) and the composition of volatile compounds (VCs). The results of this research can provide a theoretical basis for coffee bean selection for the relevant industry.
-
Arabica and Robusta beans produced in five areas were used, and the details are shown in Table 1. Sulfuric acid, hydrochloric acid and sodium hydroxide were analytical pure grade and from Wuxi Zhanwang Chemical Reagent Co., Ltd (Wuxi, China). Acetonitrile and methanol were chromatographically pure grade and from TEDIA Co., Ltd, USA. The water used in all experiments was ultrapure water.
Table 1. Dat for 40 coffee beans used in the study.
Production area/region Species Roasting degree Medium light Medium Medium dark Dark Brazil/Espirito Santo Arabica Robusta India/Karnataka Arabica Robusta Indonesia/Sumatra Arabica Robusta Uganda/Elgon Arabica Robusta Vietnam/Lam Dong Arabica Robusta Methods
Sample preparation
-
The coffee bean were reasted with 7 grade fire to four roasting degrees (medium light, medium, medium dark and dark) using a roaster (Tino Probat Inc., Germany), then ground to 0.9 mm powder after 24 h storage. The pictures of the forty samples are shown in Table 1.
Protein and fat determination
-
The experiments were carried out in accordance with the method in GB 5009.5—2016 and GB 5009.6—2016, and using Automatic Kieldahl apparatus and a digestion furnace (Hanon Advanced Technology Group Co., Ltd, China).
Organic acid determination
-
Sample preparation: For exact details please refer to the operation procedure in GB 5009.157—2016. The samples were weighed to 1−2 g in a 50 mL volumetric flask. Then, 30 mL 0.1% phosphoric acid solution was added, followed by ultrasonic treatment for 20 min. Next, 0.1% phosphoric acid solution was added to the volume scale. The flask is then shaken and filtered using a 0.22 μm organic membrane. Samples were then stored at 4 °C in the dark.
HPLC with an ultraviolet detector (1260, Agilent Technologies Co., Ltd) and a chromatographic column (CAPCELL PAK C18 MG II, 4.6 mm × 250 mm, 5 μm, OSAKA SODA CO., Ltd) was used to determine the acid concentrations, and the determined parameters are shown in Table 1.
CACs determination
-
Sample preparation: Please refer to the operation procedure in Hu et al.'s research[12]. Samples of 1−2 g were weighed in a 50 mL brown volumetric flask. Then, 30 mL methanol-0.1% phosphoric acid (50:50, V/V) solution, was added followed by ultrasonic treatment for 20 min. Methanol-0.1% phosphoric acid (50:50, V/V) solution was then added to the volume scale. The mixture was shaken and then filtered using a 0.22 μm organic membrane. Samples were stored at 4 °C in the dark.
HPLC was used to determine the CACs concentration, and the determined parameters are shown in Table 2.
Table 2. Chromatographic parameters for organic acid and CACs determination.
Parameters Organic acids CACs Speed 0.3 mL/min 1.0 mL/min Column temperature 40 °C 30 °C Wavelength 210 nm 327 nm Moving Phase Methanol (A)-0.1% phosphoric acid solution (B) Elution program 0.00~20.00 min:
10%A-90%B0.00~20.00 min:
20%A-80%B20.01~25.00 min: 100%A 20.01~45.00 min:
35%A-65%B25.01~35.00 min: 10%A-90%B 45.01~55.00 min:
40%A-60%B55.01~60.00 min:
20%A-80%BVCs determination
-
Firstly, 0.3 g of sample was weighed in a 20 mL headspace sampler, the samples were then analysed by Ultra-fast E-nose (Heracles NEO, Alpha MOS, France)[13], and the parameters are shown in Table 3.
Table 3. E-nose parameters for VCs determination.
Parameters Value Incubation temperature 60 °C Incubation duration 30 min Injection volume 5000 μL Injection speed 125 μL/s Injector temperature 200 °C Injection time 45 s Trapping temperature 40 °C Trapping duration 50 s Split 10 mL/min Final temperature 250 °C Initial temperature 50 °C (20 s) Temperature program 1 °C/s−80 °C (20 s) 3 °C/s−250 °C (40 s) Acquisition duration 167 s Fid temperature 260 °C FID gain 12 Statistical analysis
-
Experimental samples were measured three times for each experiment. Data was processed by ExcelTM and Minitab19. One-way analysis of variance (ANOVA) and statistical difference (Fisher Test) was performed at p < 0.05 using Minitab19. Principal Components Analysis (PCA) was performed by Origin 2017. Graphs were made by Origin 2017 and Photoshop CS5.
-
Protein and fat are important components of coffee beans, fats especially play an important role in the sensory attribute of coffee[14]. In addition, protein and fat are also important substances in the Maillard reaction during coffee roasting, which can contribute to the formation of pleasant flavor and color of roasted coffee beans. The results of protein and fat determination are shown in Table 4 and Fig. 1. The protein concentration of Arabica beans with different roasting degrees from all production areas was significantly lower than that of Robusta beans, as the average protein concentration of Arabica beans was about 14.3 g/100 g while that of Robusta beans was about 18.5 g/100 g. Thus, Robusta beans from Brazil, India, Indonesia, Uganda and Vietnam may have more protein than Arabica beans. With the increase of roasting degrees, it is obvious that the protein concentration of Arabica and Robusta beans from five production areas increased. It may also be closely related to the decrease in moisture content. Separating the samples into two groups according to species, a one-way ANOVA was performed at p < 0.05 using Minitab19 for each group. Arabica beans from Brazil and India, and Robusta beans from India and Vietnam had higher protein concentration, while both Arabica and Robusta beans from Indonesia and Uganda had lower protein concentration. Thus, production area may also have a significant influence on the protein concentration of coffee beans, which needs further investigation.
Table 4. Protein and fat concentration of each coffee bean (g/100 g).
Sample Protein Fat Brazil-A-1 14.85 ± 0.07jkl 6.3 ± 0.28cdefghijk Brazil-A-2 15 ± 0.14ijk 5.85 ± 0.49fghijklm Brazil-A-3 15.25 ± 0.07ij 6.7 ± 0.14bcdefghi Brazil-A-4 15.75 ± 0.07i 7.95 ± 0.49b India-A-1 14.6 ± 0.14jklm 5.15 ± 0.49jklmno India-A-2 14.7 ± 0.42jklm 5.6 ± 0.42hijklmn India-A-3 14.6 ± 0.28jklm 5.6 ± 0.28hijklmn India-A-4 15.05 ± 0.07ijk 7.15 ± 0.07bcdefg Indonesia-A-1 13.35 ± 0.07op 4.5 ± 0.57mno Indonesia-A-2 13.6 ± 0nop 6.2 ± 0.14defghijkl Indonesia-A-3 14.15 ± 0.21lmn 6.8 ± 0.42bcdefgh Indonesia-A-4 14.5 ± 0.14jklm 7.6 ± 0.14bcd Uganda-A-1 12.55 ± 0.07q 5.3 ± 0.57ijklmno Uganda-A-2 12.85 ± 0.07pq 6.75 ± 0.92bcdefghi Uganda-A-3 13.1 ± 0.14pq 7.95 ± 0.64b Uganda-A-4 13.15 ± 0.07pq 10.05 ± 0.35a Vietnam-A-1 14.07 ± 0.21mno 6.35 ± 0.52cdefghijk Vietnam-A-2 14.45 ± 0.25klm 7.36 ± 0.36bcde Vietnam-A-3 14.69 ± 0.13jklm 7.74 ± 0.06bc Vietnam-A-4 14.85 ± 0.07jkl 9.46 ± 0.35a Brazil-R-1 17.78 ± 0.3efg 4.07 ± 0.3o Brazil-R-2 18.36 ± 0.19def 5 ± 0.21jklmno Brazil-R-3 18.8 ± 0.15cd 4.87 ± 0.33klmno Brazil-R-4 18.93 ± 0.05bcd 5.84 ± 0.21fghijklm India-R-1 19.04 ± 0.08bcd 4.85 ± 0.02lmno India-R-2 19.52 ± 0.02abc 5.3 ± 0.24ijklmno India-R-3 19.66 ± 0.01ab 5.92 ± 0.07efghijklm India-R-4 19.86 ± 0.1a 6.19 ± 0.25defghijkl Indonesia-R-1 17.02 ± 0.12h 4.33 ± 0.16no Indonesia-R-2 17.34 ± 0.37gh 5.72 ± 0.33ghijklmn Indonesia-R-3 17.57 ± 0.19gh 5.99 ± 0.21efghijklm Indonesia-R-4 18.37 ± 14def 5.99 ± 7.1efghijklm Uganda-R-1 17.44 ± 0.09gh 6.15 ± 0.17defghijkl Uganda-R-2 17.68 ± 0.03fgh 5.49 ± 0.3hijklmno Uganda-R-3 17.82 ± 0.35efg 6.45 ± 0.47cdefghij Uganda-R-4 18.48 ± 0.17de 6.39 ± 0.01cdefghij Vietnam-R-1 18.47 ± 0.17de 5.52 ± 0.28hijklmno Vietnam-R-2 19.39 ± 0.07abc 5.57 ± 0.35hijklmn Vietnam-R-3 19.45 ± 0.24abc 6.02 ± 0.14efghijkl Vietnam-R-4 19.94 ± 0.16a 7.32 ± 0.09bcdef *A: Arabica bean; R: Robusta bean
1: medium light; 2: medium; 3: medium dark; 4: dark
Letter code: samples with the same letter code are not significantly different (p < 0.05)
Figure 1. Protein and fat concentration of coffee beans (g/100 g). (a) & (b), protein concentration; (c) & (d), fat concentration. Blue represents Arabica beans, orange represents Robusta beans. 1, medium light; 2, medium; 3, medium dark; 4, dark.
The fat concentration of coffee beans was generally positively correlated to the roasting degree, which was also closely associated with the decrease of moisture content. And, fat concentration of Arabica beans was obviously higher than that of Robusta beans. Thus, roasting degree and species may be significant factors that affect the fat concentration of coffee beans. According to Fig. 1d, there was no significant difference among fat concentration of coffee beans from each production area. Thus, production area might not influence the fat concentration of coffee beans.
Organic acid and chlorogenic acid compounds
-
During the roasting of coffee beans, organic acids are produced while CACs are broken down. Similar to fat and protein, organic acids are also important compounds in the sensory properties of coffee. As they determine the pH value, which is related to the acidity of coffee[13]. The results of organic acids and CACs are shown in Table 5. One-way ANOVA was performed at p < 0.05 using Minitab19 in each species of coffee beans, and the results are shown in Fig. 2.
Table 5. Organic acids and CACs concentration of each coffee bean (g/100 g).
Sample Organic acids Chlorogenic acids compounds Tartaric
acidMalic
acidCitric
acidSuccinic
acidFumaric
acidTotal Neochlorogenic
acidChlorogenic
acidCryptochlorogenic
acidIsochlorogenic
acid AIsochlorogenic
acid BIsochlorogenic
acid CTotal Brazil-A-1 3.48 ± 0.24 0.46 ± 0.15 0.44 ± 0.06 1.37 ± 0.27 0 ± 0 5.75 ± 0.71ghijklmn 0.45 ± 0.02 0.97 ± 0.08 0.58 ± 0.07 0.03 ± 0.01 0.06 ± 0.02 0.06 ± 0.01 2.16 ± 0.2de Brazil-A-2 3.44 ± 0.23 0.48 ± 0.11 0.64 ± 0.06 2.13 ± 0.28 0 ± 0 6.67 ± 0.21cdefg 0.33 ± 0 0.67 ± 0.02 0.38 ± 0.02 0.02 ± 0 0.03 ± 0 0.03 ± 0 1.45 ± 0.01fghi Brazil-A-3 3.97 ± 0.13 0.53 ± 0.06 0.69 ± 0.06 2.49 ± 0.29 0 ± 0 7.67 ± 0.04bcd 0.22 ± 0 0.41 ± 0.01 0.28 ± 0.02 0 ± 0 0.02 ± 0.01 0.02 ± 0 0.94 ± 0.04jkl Brazil-A-4 4.18 ± 0.23 0.64 ± 0.05 1.04 ± 0.07 1.94 ± 0.11 0 ± 0 7.8 ± 0.36bc 0.13 ± 0 0.22 ± 0.01 0.15 ± 0.01 0 ± 0 0 ± 0 0 ± 0 0.5 ± 0.02nop India-A-1 2.61 ± 0.25 0.52 ± 0.04 0.49 ± 0.04 1.59 ± 0.05 0 ± 0 5.2 ± 0.12hijklmnop 0.71 ± 0.1 1.42 ± 0.04 0.85 ± 0.07 0.05 ± 0.01 0.08 ± 0.01 0.08 ± 0.01 3.19 ± 0.24ab India-A-2 3.58 ± 0.11 0.55 ± 0.06 0.8 ± 0.04 0.49 ± 0.09 0 ± 0 5.41 ± 0.3ghijklmnop 0.38 ± 0.05 0.74 ± 0.07 0.48 ± 0.1 0.02 ± 0 0.03 ± 0.01 0.03 ± 0 1.68 ± 0.23fgh India-A-3 3.62 ± 0.05 0.73 ± 0.07 1.01 ± 0.11 2.77 ± 0.01 0 ± 0 8.13 ± 0.25b 0.27 ± 0.02 0.49 ± 0.02 0.32 ± 0.02 0.01 ± 0 0.02 ± 0 0.02 ± 0 1.13 ± 0.07ijk India-A-4 3.75 ± 0.06 0.81 ± 0.1 2.6 ± 0.07 2.77 ± 0.25 0 ± 0 9.93 ± 0.49a 0.15 ± 0 0.27 ± 0.01 0.19 ± 0.02 0 ± 0 0.01 ± 0.01 0 ± 0.01 0.62 ± 0.03lmnop Indonesia-A-1 2.9 ± 0.28 0.33 ± 0.01 1.08 ± 0.06 1.03 ± 0.05 0 ± 0.01 5.33 ± 0.39ghijklmnop 0.67 ± 0.02 1.51 ± 0.08 0.81 ± 0.02 0.05 ± 0.01 0.09 ± 0.01 0.09 ± 0.01 3.22 ± 0.14a Indonesia-A-2 3.53 ± 0.17 0.59 ± 0.08 1.05 ± 0.06 1.11 ± 0.07 0.02 ± 0.01 6.29 ± 0.27defghij 0.5 ± 0.01 1.11 ± 0.02 0.65 ± 0.07 0.03 ± 0.01 0.06 ± 0.01 0.05 ± 0 2.41 ± 0.12cd Indonesia-A-3 3.9 ± 0.16 0.51 ± 0.04 0.71 ± 0.15 1.47 ± 0.04 0 ± 0 6.57 ± 0.38cdefgh 0.31 ± 0.01 0.58 ± 0.01 0.36 ± 0.01 0.01 ± 0 0.03 ± 0 0.03 ± 0 1.32 ± 0.04hij Indonesia-A-4 4.87 ± 0.23 0.73 ± 0.07 1.01 ± 0.04 1.19 ± 0.06 0 ± 0 7.8 ± 0.39bc 0.15 ± 0 0.25 ± 0.01 0.17 ± 0 0 ± 0 0 ± 0.01 0 ± 0.01 0.58 ± 0.01lmnop Uganda-A-1 2.92 ± 0.19 0.57 ± 0.11 1.47 ± 0.24 1.52 ± 0.24 0 ± 0.01 6.48 ± 0.78cdefghi 0.6 ± 0 1.28 ± 0 0.74 ± 0 0.04 ± 0 0.07 ± 0 0.07 ± 0 2.79 ± 0.02bc Uganda-A-2 3.07 ± 0.11 0.55 ± 0.15 1.81 ± 0.23 2.2 ± 0.16 0.01 ± 0.01 7.62 ± 0.64bcde 0.4 ± 0.01 0.86 ± 0.04 0.49 ± 0.01 0.02 ± 0 0.04 ± 0 0.04 ± 0 1.85 ± 0.07ef Uganda-A-3 3.47 ± 0.08 0.55 ± 0.15 2.04 ± 0.13 2.5 ± 0.22 0.01 ± 0.01 8.56 ± 0.4ab 0.26 ± 0.01 0.51 ± 0 0.32 ± 0 0.02 ± 0 0.02 ± 0 0.02 ± 0 1.14 ± 0.02ijk Uganda-A-4 3.57 ± 0.11 0.48 ± 0.1 0.61 ± 0.04 1.7 ± 0.14 0.02 ± 0 6.38 ± 0.38cdefghij 0.14 ± 0.01 0.24 ± 0.02 0.17 ± 0.01 0 ± 0 0 ± 0 0 ± 0 0.56 ± 0.04lmnop Vietnam-A-1 1.28 ± 0.11 0.32 ± 0.05 1.55 ± 0.13 2.21 ± 0.18 0.01 ± 0 5.36 ± 0.46ghijklmnop 0.48 ± 0.04 1.18 ± 0.08 0.59 ± 0.02 0.04 ± 0 0.08 ± 0.01 0.08 ± 0 2.45 ± 0.15cd Vietnam-A-2 1.46 ± 0.05 0.3 ± 0.04 1.27 ± 0.08 3.1 ± 0.01 0.02 ± 0 6.14 ± 0.09fghijkl 0.29 ± 0.01 0.65 ± 0.02 0.38 ± 0.04 0.02 ± 0 0.03 ± 0 0.04 ± 0.01 1.41 ± 0.08ghi Vietnam-A-3 1.73 ± 0.06 0.25 ± 0.03 1 ± 0.01 3.2 ± 0.07 0.01 ± 0.01 6.19 ± 0.18efghijk 0.17 ± 0.01 0.35 ± 0.02 0.23 ± 0.04 0.01 ± 0 0.02 ± 0 0.02 ± 0 0.8 ± 0.07klmno Vietnam-A-4 1.97 ± 0.1 1.11 ± 0.04 0.79 ± 0.03 3.41 ± 0.04 0.03 ± 0.01 7.3 ± 0.21bcdef 0.11 ± 0.02 0.19 ± 0.02 0.13 ± 0.02 0 ± 0 0 ± 0 0 ± 0 0.42 ± 0.06op Brazil-R-1 2.19 ± 0.12 0.1 ± 0.01 0.39 ± 0.03 0.14 ± 0.02 0 ± 0.01 2.83 ± 0.18s 0.65 ± 0.04 1.43 ± 0.04 0.8 ± 0.02 0.05 ± 0 0.09 ± 0 0.09 ± 0.01 3.11 ± 0.11ab Brazil-R-2 2.66 ± 0.38 0.39 ± 0.04 0.27 ± 0.04 0 ± 0 0.01 ± 0.01 3.33 ± 0.37rs 0.39 ± 0.03 0.78 ± 0.05 0.48 ± 0.02 0.03 ± 0 0.04 ± 0 0.04 ± 0 1.75 ± 0.1efg Brazil-R-3 3.04 ± 0.1 0.73 ± 0.07 0.16 ± 0.04 0.41 ± 0.02 0 ± 0 4.33 ± 0.24nopqr 0.16 ± 0 0.29 ± 0.01 0.2 ± 0.01 0 ± 0 0 ± 0.01 0 ± 0.01 0.66 ± 0.01lmnop Brazil-R-4 3.8 ± 0.25 0.3 ± 0.04 0.09 ± 0.02 0.38 ± 0 0 ± 0 4.58 ± 0.27mnopqr 0.08 ± 0.01 0.13 ± 0.01 0.09 ± 0.01 0 ± 0 0 ± 0 0 ± 0 0.29 ± 0.03p India-R-1 3.06 ± 0.21 0.28 ± 0.04 0.28 ± 0.05 0 ± 0 0 ± 0 3.62 ± 0.3qrs 0.5 ± 0.01 0.99 ± 0.03 0.63 ± 0.04 0.06 ± 0.02 0.1 ± 0 0.09 ± 0 2.36 ± 0.1d India-R-2 3.44 ± 0.19 0.85 ± 0.09 0.23 ± 0.03 0 ± 0 0 ± 0 4.52 ± 0.24mnopqr 0.29 ± 0 0.57 ± 0.04 0.36 ± 0.02 0.02 ± 0.01 0.04 ± 0 0.04 ± 0 1.33 ± 0.08hij India-R-3 3.15 ± 0.21 1.01 ± 0.03 0.22 ± 0.01 0.4 ± 0.03 0.02 ± 0.01 4.8 ± 0.15klmnopq 0.21 ± 0 0.39 ± 0.03 0.26 ± 0.01 0.01 ± 0 0.03 ± 0.01 0.02 ± 0 0.92 ± 0.06jklm India-R-4 3.87 ± 0.19 1.26 ± 0.14 0.26 ± 0.06 0.5 ± 0.04 0.02 ± 0 5.91 ± 0.43fghijklm 0.12 ± 0.01 0.21 ± 0.01 0.14 ± 0 0.01 ± 0 0.01 ± 0 0.01 ± 0 0.51 ± 0.03mnop Indonesia-R-1 2.74 ± 0.22 0.32 ± 0.03 0.68 ± 0.05 0.34 ± 0.06 0.01 ± 0.01 4.09 ± 0.37pqrs 0.5 ± 0.02 0.99 ± 0.02 0.62 ± 0.02 0.05 ± 0 0.09 ± 0 0.09 ± 0.01 2.32 ± 0.08d Indonesia-R-2 3.14 ± 0.15 0.43 ± 0.02 0.47 ± 0.07 0.23 ± 0 0.01 ± 0.01 4.27 ± 0.21opqr 0.33 ± 0.01 0.68 ± 0.05 0.45 ± 0.06 0.03 ± 0.01 0.08 ± 0.01 0.05 ± 0.01 1.63 ± 0.14fgh Indonesia-R-3 3.63 ± 0.11 0.47 ± 0.01 0.29 ± 0.02 0.33 ± 0.02 0.01 ± 0.01 4.73 ± 0.15lmnopqr 0.14 ± 0.02 0.25 ± 0.02 0.16 ± 0.01 0.01 ± 0 0.01 ± 0 0.01 ± 0 0.58 ± 0.05lmnop Indonesia-R-4 3.66 ± 3.78 0.16 ± 0.48 0.22 ± 0.6 0.38 ± 1.44 0.01 ± 0.004 4.43 ± 0.37nopqr 0.1 ± 0.3 0.17 ± 0.577 0.12 ± 0.353 0 ± 0.011 0 ± 0.024 0 ± 0.024 0.4 ± 1.289op Uganda-R-1 3.27 ± 0.15 0.31 ± 0.04 0.54 ± 0.06 0.25 ± 0.04 0.01 ± 0 4.38 ± 0.29nopqr 0.5 ± 0.01 1.08 ± 0.05 0.67 ± 0.07 0.05 ± 0.01 0.08 ± 0.01 0.08 ± 0 2.46 ± 0.15cd Uganda-R-2 3.52 ± 0.2 0.25 ± 0.13 0.45 ± 0.04 0.3 ± 0.02 0.02 ± 0 4.53 ± 0.06mnopqr 0.39 ± 0.03 0.77 ± 0.06 0.5 ± 0.06 0.03 ± 0.01 0.05 ± 0.01 0.05 ± 0 1.78 ± 0.18efg Uganda-R-3 3.9 ± 0.33 0.16 ± 0.01 0.11 ± 0.01 0.37 ± 0.08 0.02 ± 0 4.56 ± 0.41mnopqr 0.1 ± 0.12 0.35 ± 0.02 0.27 ± 0.06 0 ± 0 0.02 ± 0 0.01 ± 0.01 0.76 ± 0.05klmno Uganda-R-4 4.01 ± 0.17 0.25 ± 0.13 0.11 ± 0.01 0.41 ± 0.03 0.02 ± 0.01 4.8 ± 0.07klmnopq 0.07 ± 0.02 0.15 ± 0.01 0.1 ± 0.01 0 ± 0 0 ± 0 0 ± 0 0.32 ± 0.03p Vietnam-R-1 3.96 ± 0.08 0.16 ± 0.08 0.46 ± 0.07 0.38 ± 0.04 0 ± 0 4.96 ± 0.1jklmnopq 0.7 ± 0.02 1.42 ± 0.05 0.88 ± 0.05 0.08 ± 0 0.15 ± 0.01 0.14 ± 0 3.36 ± 0.14a Vietnam-R-2 3.94 ± 0.21 0.5 ± 0.04 0.2 ± 0 0.48 ± 0.04 0.01 ± 0.01 5.13 ± 0.13ijklmnop 0.3 ± 0.01 0.54 ± 0.01 0.37 ± 0.02 0.02 ± 0 0.08 ± 0 0 ± 0 1.31 ± 0.04hij Vietnam-R-3 4.69 ± 0.14 0.81 ± 0.13 0.19 ± 0.02 0.5 ± 0.06 0 ± 0 6.19 ± 0.35efghijk 0.2 ± 0 0.36 ± 0 0.24 ± 0 0.01 ± 0 0.02 ± 0 0.01 ± 0.01 0.85 ± 0.02klmn Vietnam-R-4 4.92 ± 0.12 0.5 ± 0.01 0.14 ± 0.01 0 ± 0 0 ± 0 5.56 ± 0.13ghijklmno 0.07 ± 0 0.12 ± 0 0.08 ± 0 0 ± 0 0 ± 0 0 ± 0 0.28 ± 0.01p *A: Arabica bean; R: Robusta bean;
1: medium light, 2: medium, 3: medium dark; 4: dark;
Letter code: Samples with the same letter code are not significantly different (P < 0.05).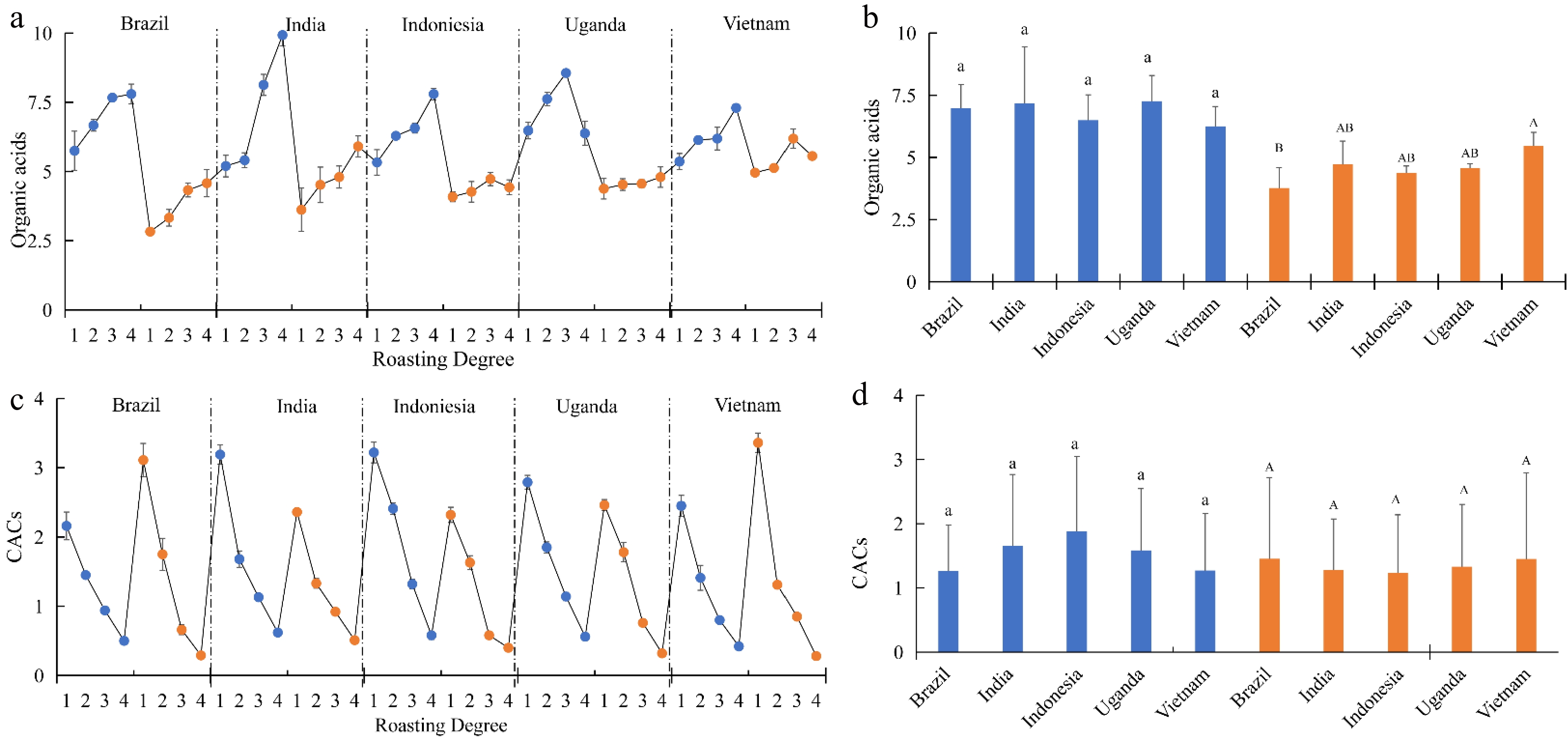
Figure 2. Organic acid and CACs concentration of coffee beans (g/100 g). (a) & (b), organic acid concentration; (c) & (d), CACs concentration. Blue represents Arabica beans, Orange represents Robusta beans. 1, medium light; 2, medium; 3, medium dark; 4, dark.
According to Fig. 2a, for most coffee beans, the total organic acid concentration increased with the increase of roasting degree. However, when the roasting degree of Arabica beans from Uganda and Robusta beans from Vietnam was dark, the concentration of total organic acids fell significantly, which corresponds to the results of Wang[15]. And the significant decrease of total concentration was mainly as a result of the decease of citric acid and succinic acid, which are easily decomposed at high temperature[8]. Between the groups of different species, the value of Robusta beans was lower, and the value was significantly different (Fig. 2b). While the value of Arabica was not different from each other. Thus, production area may influence the organic acid concentration of Robusta beans.
For the concentration of CACs of each coffee beans, it had an obvious negative correlation with roasting degree. With regard to production area, there was no significant influence on the CACs concentration of two species of coffee beans (Fig. 2d). Also, the value of Arabica beans was similar to that of Robusta. Therefore, the concentration of CACs may only be influenced by roasting degree.
Volatile compounds
-
VCs are the main contributors to the odor of coffee, usually related to roasted coffee oil[16]. VCs can be composed of various chemical compounds, such as aldehydes, esters, ketones, alcohols, hydrocarbons, phenols, carboxylic acids, pyridine, pyrazines, furans and so on. The result of each volatile compound of each sample are shown in Supplemental Table S1. The total amount of VCs of coffee beans are shown in Fig. 3. For different species of coffee beans, there were some unique compounds. Arabica beans contained furfural and butyraldehyde isopropyl ester, and Robusta beans contained methylheptenone. Also, the total amount of VCs were also different. As the value of Arabica beans was higher than that of Robusta beans, the total amount of VCs of most coffee beans increased with the increase of roasting degree. As for the production area, there was no significant difference between each species of coffee beans (Fig. 3b).
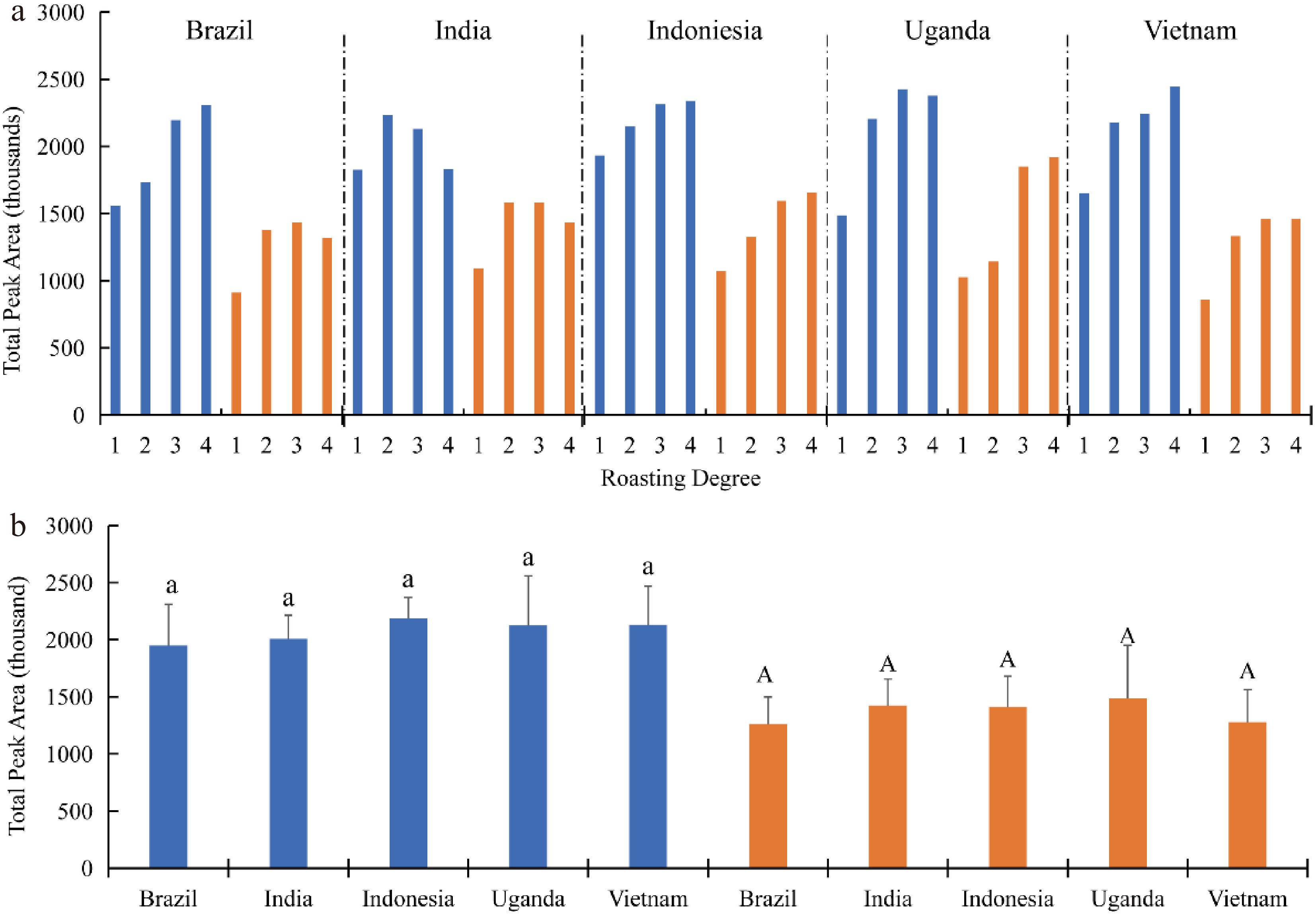
Figure 3. Total amount of VCs of different coffee beans. Blue represents Arabica beans, Orange represents Robusta beans. 1, medium light; 2, medium; 3, medium dark; 4, dark.
Principal component analysis
-
PCA was performed on five indexes (protein concentration, fat concentration, total organic acid concentration, total CACs concentration and total amount of VCs) of 40 samples, and the results are shown in Table 6. Three principal components were extracted, and the cumulative variance contribution rate was 93.1%. Based on PC1 and PC2, a discriminant analysis was performed among the 40 samples, shown in Fig. 4. The results clearly showed that Arabica bean and Robusta bean, and the beans with different roasting degrees were discriminated by PC1 and PC2. While coffee beans from the same production area did not cluster together clearly. Thus, species and roasting degrees can distinguish coffee beans better than production area, which corresponds to the above results. Coffee beans produced in different areas may have the same or similar origin. In addition, Brazil, India, Indonesia, Uganda and Vietnam are located on a similar latitude. So, the same species of coffee bean from these production areas is mostly likely to show similar characteristics.
Table 6. Loading table of each index of coffee bean
Index PC1 PC2 PC3 Protein −0.39 0.613 −0.162 Fat 0.489 0.249 0.501 Total organic acids 0.501 −0.097 −0.806 Total CACs 0.272 −0.74 0.118 Total VCs 0.532 −0.066 0.241 RVC (%) 60.1 26.5 6.6 Cumulative RVC (%) 93.1 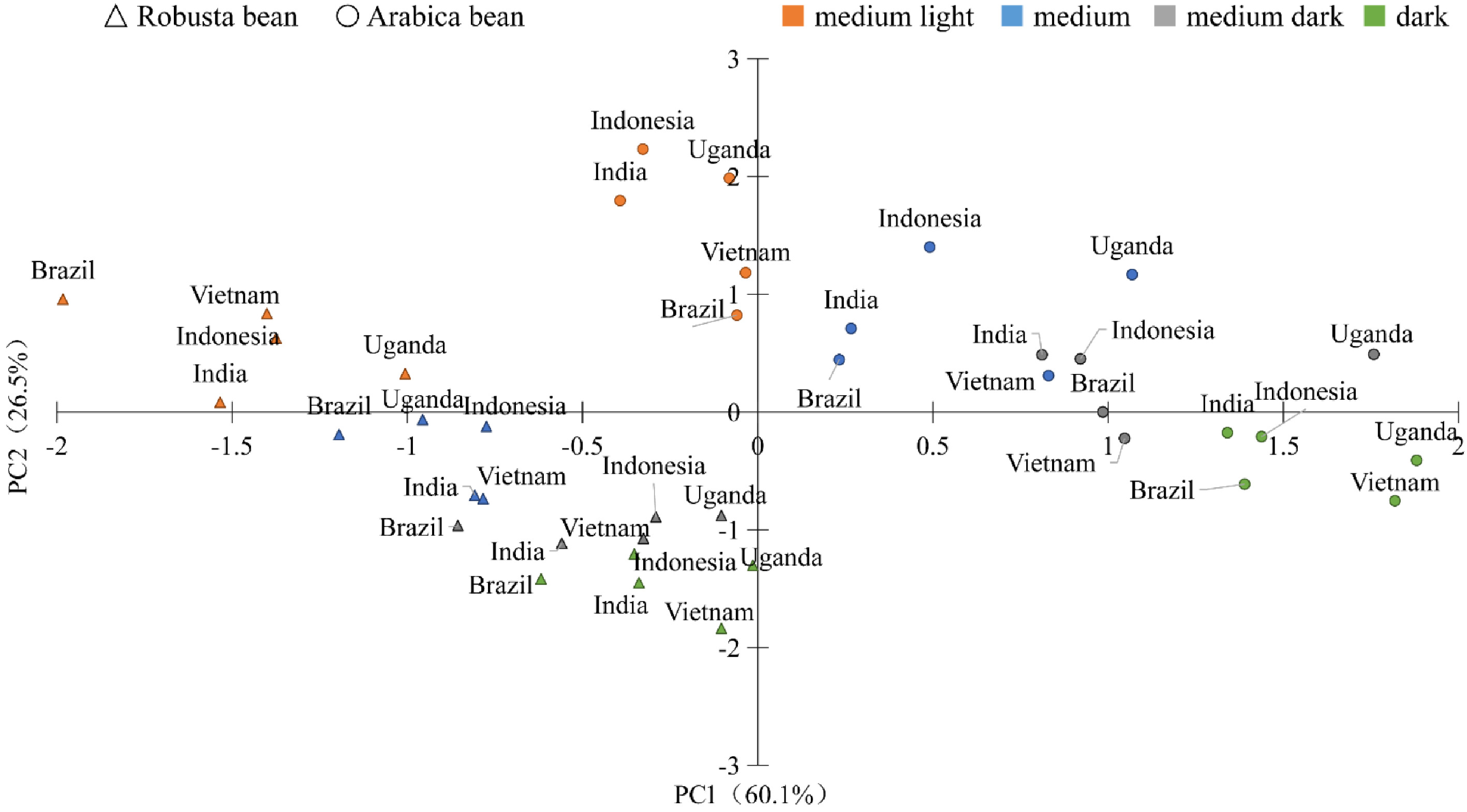
Figure 4. Discriminant results of coffee beans.
-
In this research, coffee beans of two species (Arabica beans and Robusta beans) from five different production areas (Brazil, India, Indonesia, Uganda and Vietnam) with four different roasting degrees (medium light, medium, medium dark and dark), and a total of 40 samples, were used. The protein, fat, organic acids, CACs and VCs were measured to investigate the difference among the samples. The results suggested that coffee beans of two species had obviously different substance concentrations, as Arabica coffee beans had higher concentrations of fat and organic acids, and total amount of VCs, while Robusta beans had higher concentrations of protein. Due to the increasing loss of moisture, the concentration of protein, fat, organic acids, and the total amount of VCs of coffee beans increased with the increase of roasting degree. The concentration of CACs was lower, while the roasting degree of coffee beans was higher, as CACs decomposed at high temperature. Compared to species and roasting degree, production area influenced only part of the index. The concentration of protein of two species of coffee beans and the concentration of organic acids of Robusta beans were affected significantly by production area. For further conclusions on the influence of production area on coffee beans, further research is required. Based on the results of the measurement of protein, fat, organic acids, CACs and VCs, PCA was performed to illustrate the correlation among species, production area and roasting degree with coffee beans. The discriminant analysis indicated that species and roasting degrees could differentiate the coffee beans clearly. While production area could not differentiate the coffee beans well.
The results of this research conclude the difference between Arabica and Robusta beans, and coffee beans with different roasting degrees from different production areas. It can provide a theoretical basis for coffee bean selection for the relevant industry.
- The authors declare that they have no conflict of interest.
- Supplemental Table1 Amount of Volatile Compounds of Each Coffee Bean.
- Copyright: © 2022 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu X, Fei Y, Wang W, Lei S, Cheng C, et al. 2022. Physicochemical difference of coffee beans with different species, production areas and roasting degrees. Beverage Plant Research 2: 7 doi: 10.48130/BPR-2022-0007

Physicochemical difference of coffee beans with different species, production areas and roasting degrees
- Received: 31 December 2021
- Accepted: 17 March 2022
- Published online: 12 April 2022
Abstract: In recent decades, the demand for coffee has seen a continuous increase, and the aroma and flavor of coffee has been widely studied. The current research chose coffee beans of two species (Coffea arabica and C. canephora) from five production areas (Brazil, India, Indonesia, Uganda and Vietnam) with four different roasting degrees (medium light, medium, medium dark and dark), to investigate the difference on physicochemical properties. The results showed that Arabica coffee beans had higher concentrations of fat and organic acids, and total amount of volatile compounds, whereas Robusta beans had higher concentrations of protein. With the increase of roasting degree, the concentrations of protein, fat, organic acids, and the total amount of volatile compounds of coffee beans increased, while the concentrations of chlorogenic acid compounds decreased. The discriminant analysis indicated that the tested coffee beans could be clearly discriminated by species and roasting degrees, but not by production area. The results of this research conclude the physicochemical difference of Arabica and Robusta beans with different roasting degrees. The results can provide a theoretical basis for coffee bean selection for the relevant industries.
-
Key words:
- Coffee beans /
- Species /
- Production Area /
- Roasting degree