









-
Melatonin (N-acetyl-5 methoxytryptamine, melatonin, MT) is a potent antioxidant commonly present in animals and plants[1]. It protects plants from pathogenic bacteria, drought, salinity, heat, low temperature, and heavy metal stress[2] and regulates plant leaf[3], root[4], and hypocotyl development[5]. The molecular mechanism of melatonin to function has been described previously, for example, melatonin enhances plant resistance to pathogens through the MKK4/5/7/9 (MAP kinase kinase 4/5/7/9)-MPK3/6 (MAP kinase) cascade reaction[6], CAND2/PMTR1 as a possible melatonin receptor is involved in melatonin-dependent regulation of stomatal closure in Arabidopsis[7]. It should however be noted that for many hormones to function, there are usually interacting factors involved. For example, ethylene promotes fruit ripening, but MdERF2 negatively regulates ethylene-mediated apple ripening by interacting with MdERF3[8]. ABA (Abscisic acid) maintains seed dormancy, and ODR1 interacts with bHLH57 in the cell nucleus and negatively regulates ABA levels to reverse seed dormancy[9]. Some interaction factors that are involved in melatonin responses have been identified in multiple species, such as the interaction between the heat shock protein HSP40 and SlSNAT2 (encodes a key enzyme in melatonin synthesis) that enhances plant heat tolerance[10] and the interaction between MeWRKY20/MeWRKY75 and MeTDC/MeASMT that regulates endogenous melatonin levels[11]. However, large numbers of interaction factors in melatonin-mediated plant development remain to be explored.
Lilies are among the most important cut flowers in the world. The quality of commercial bulbs is an important factor that affects the quality of cut flowers, and small bulbs are indispensable for obtaining high-quality com>mercial bulbs; however, the cycle of commercial bulb cultivation is long[12]. Therefore, an important goal is to reduce the cycle of commercial lily bulblet development. Previous research has shown that melatonin promoted lily bulblet development, the authors found that application of 20 μM melatonin increased the number of cells at the base of lily bulblet shoot apex, and up-regulated the expression of cell proliferation genes LoCYCB2-1 (B-type cyclin) and LoPCNA (Proliferating cell nuclear antigen), down-regulated genes that inhibit cell proliferation LoKRP3 (Cyclin-dependent kinase inhibitor), and increased the content of trehalose and starch in bulbs[13]. The authors constructed a possible regulatory network that melatonin regulated bulb development, but the key regulators involved in melatonin-mediated cell proliferation and starch accumulation have not been investigated.
Protein degradation mediated by melatonin is the key factor that slowed leaf senescence in Malus hupehensis[14] and promoted the proliferation of Nicotiana tabacum L. line Bright Yellow 2 (BY-2) cell during sugar starvation[15]. Interestingly, melatonin promotion of lily bulb cell proliferation and accumulation of starch and trehalose are also related to protein degradation[13], which means that protein degradation factors may be key factors involved in melatonin-mediated bulb development. BPM is a ubiquitin-related gene containing the MATH and BTB/POZ structural domains[16]. The BTB/POZ structural domain assembles with CUL3 to form a functional E3 ubiquitin ligase complex, and the MATH structural domain is responsible for the recognition and recruitment of specific substrate proteins, resulting in the posttranslational regulation of the latter[17,18]. RAVs are a class of transcription factors that contain AP2/B3 domains[19], can regulate the plant cell cycle[20], limit seed size[21], and mediate ABA signaling[22]. Almost all ERF/AP2 transcription factors interact with BTB/POZ-MATH proteins in Arabidopsis thaliana[23]. This means that BPM and RAV may mediate multiple metabolic processes of plants in an interactive manner, even including hormonal responses.
However, RAV and BPM belong to a multi-member protein family, and previous research on RAV or BPM has mainly focused on reproductive organs and stress responses; researchers have not determined whether RAV is involved in underground storage organ development and in mediating the melatonin response. In this study, we identified that the LoBPM3-LoRAV module plays an important role in the melatonin-mediated regulation of lily bulblet development by regulating cell proliferation, which has not been previously reported in lily. This report provides a basis for elucidating the molecular mechanism by which melatonin regulates the development of lily bulblets.
-
The morphology of the bulblets was observed to investigate the effect of melatonin on the development of lily bulblets. The lily bulblets inoculated with 20 μM melatonin medium showed white inner scales and upward growth for 7 d, the inner scales were erect and prominent at 20 d, and the bulblets were fuller than the control lily bulblets (Fig. 1a, b). The mean circumference and height of lily bulblets at 20 d were subsequently measured. The mean circumference of the lily bulblets in the control group was 3.43 cm, while the mean circumference of the lily bulblets in the melatonin-treated group was 4.70 cm, which was 37.1% greater than that of the control group (Fig. 1c). The mean bulblet height in the control group was 1.55 cm, while the mean bulblet height in the melatonin-treated group was 1.88 cm, but the difference was not significant (Fig. 1d). These data indicated that melatonin promotes lily bulblet development, including scale growth and radial growth of bulblets.
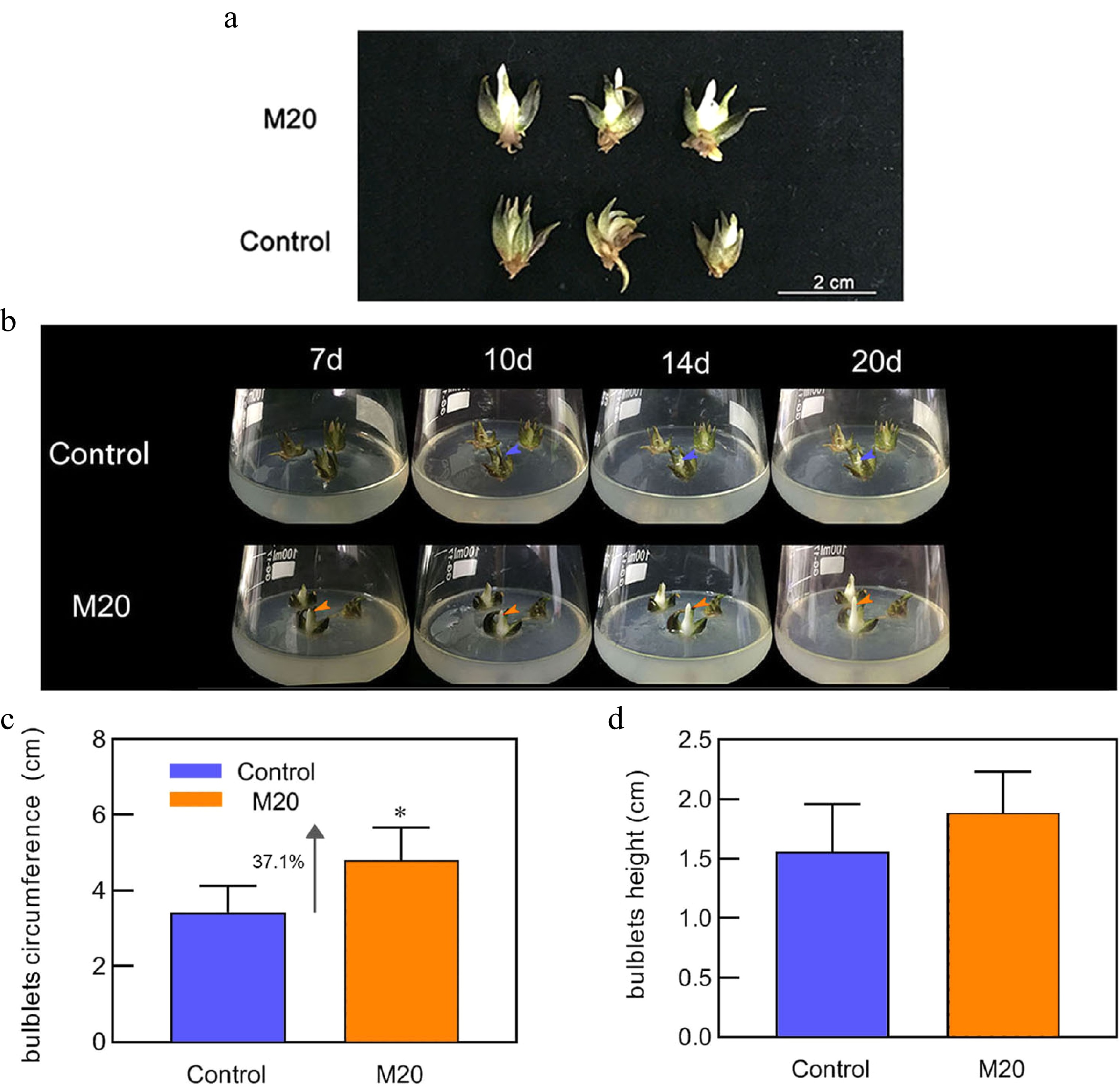
Figure 1.
Morphology of the bulblets in different media. (a) Phenotype of bulblets on day 20 in different treatment groups. (b) Growth status of bulblets in the control and M20 medium (containing 20 μM melatonin) at different incubation time points. Blue and orange arrows indicate internal scales in different treatment groups. (c) Bulblet circumference. (d) Bulblet height. Blue and orange boxes indicate the control and melatonin treatment groups, respectively. Values are presented as the means of three replicates ± SD. Data were analyzed using one-way analysis of variance (ANOVA) followed by Duncan's test; * indicates a significant difference (P < 0.05).
Effects of melatonin on starch and trehalose metabolism in lily bulblets
-
To confirm that the phenotype was valid, we examined indicators related to bulb development. Scale growth is inseparable from cell proliferation, so the expression levels of genes related to cell proliferation were measured. Compared with the control, the expression levels of LoPCNA and LoCYCB2-1 increased by 58% and 52% (Fig. 2a, b), respectively, on day 10 in the group treated with melatonin. The expression level of LoTCP4 ( a class II TCP gene, encoding a TCP family transcription factor) decreased by 3.2-fold and 2.7-fold on days 10 and 20, and the expression level of LoKRP3 (a gene related to the suppression of cell proliferation) decreased by 0.85-fold and 1-fold on days 10 and 20, respectively, in the melatonin-treated group (Fig. 2c, d). These data suggest that melatonin promotes cell proliferation and thus growth of bulblets. Trehalose is a non-reducing disaccharide that can act as a signal to directly or indirectly regulate the plant cell cycle[24]. Starch is an important storage material for bulblets, and ADP glucose pyrophosphorylase (AGPase) is the key enzyme of starch synthesisits synthesis that initiates redox activation by trehalose[25]. We therefore detected the content of starch and trehalose in the lily bulblets cultivated for 10 days. The content of starch and trehalose in the bulblets of the control group were 44.94 mg/g and 23.81 mg/g, respectively. However, the starch and trehalose content in the melatonin-treated group were 57.84 mg/g and 49.64 mg/g, which was increased by 28.7% and 1.08-fold, respectively, compared with the control (Fig. 2e, f). Moreover, the expression of LoAGPase (encodes a rate-limiting enzyme related to starch synthesis) increased by 2.2-fold and 0.6-fold (Fig. 2g) on days 10 and 20, and the expression of LoTPP (trehalose-6-phosphate phosphatase gene, encoding trehalose synthesis enzyme) increased by 2.4-fold and 0.5-fold (Fig. 2h), respectively, in the melatonin-treated group. These data suggest that melatonin promotes the accumulation of bulb starch.
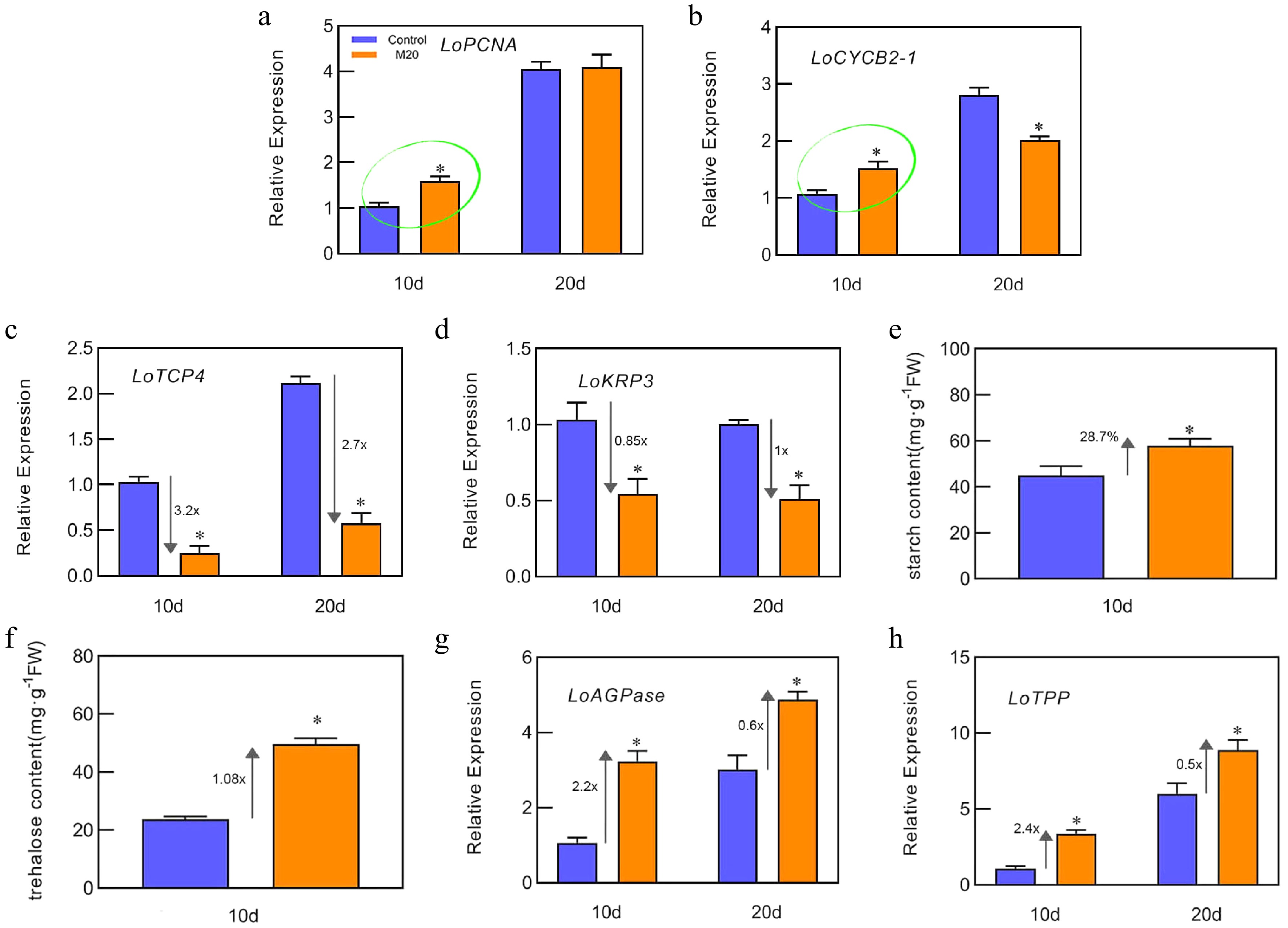
Figure 2.
Effects of melatonin on starch and trehalose metabolism. (a)−(d), (g)−(h) The expression levels of LoPCNA, LoCYCB2-1, LoTCP4, LoKRP3, LoAGPase, and LoTPP on days 10 and 20 of bulblet cultivation, respectively. (e), (f) The content of trehalose and starch in bulblets from the control and melatonin-treated groups on day 10. Expression levels were calculated relative to those of Lilium FP mRNA. Blue and orange boxes indicate the control and melatonin treatment groups, respectively. Values are presented as the means of three replicates ± SD. Data were analyzed using one-way analysis of variance (ANOVA) followed by Duncan's test; * indicates a significant difference (P < 0.05).
Effects of melatonin on LoRAV expression and LoRAV clone analysis
-
Previous studies have shown that AP2 transcription factor can be ubiquitinated by the CULLIN3-base E3 ligase assembled with by BPB/POZ-MATH protein, so we may form a module with LoBPM3 and LoRAV to function[26]. We sought to understand whether the LoBPM3-LoRAV module is involved in melatonin-mediated bulblet development. We first cloned and expressed LoRAV and analyzed the effects of the expression of this gene. The expression of LoRAV was decreased by 36.5% in the melatonin-treated group compared with the control group at day 10 (Fig. 3a). Cloning and sequencing showed that LoRAV contains a complete open reading frame of 1,056 bp, encoding 351 amino acids, with a protein molecular weight of 38.906 kD and a theoretical isoelectric point (pI) of 9.43 (Fig. 3b, c). The protein sequence analysis showed that LoRAV is an AP2/B3 transcription factor containing AP2 and B3 domains at amino acids 46−105 and 171−250, respectively, and a relatively conserved ubiquitination-like motif were observed (Fig. 3d). We also performed a multiple sequence alignment of LoRAV with RAV family member genes of Arabidopsis . As shown in Fig. 3e, the protein sequence of LoRAV are highly conserved and AtRAV1/AtRAV2/AtTEM1.
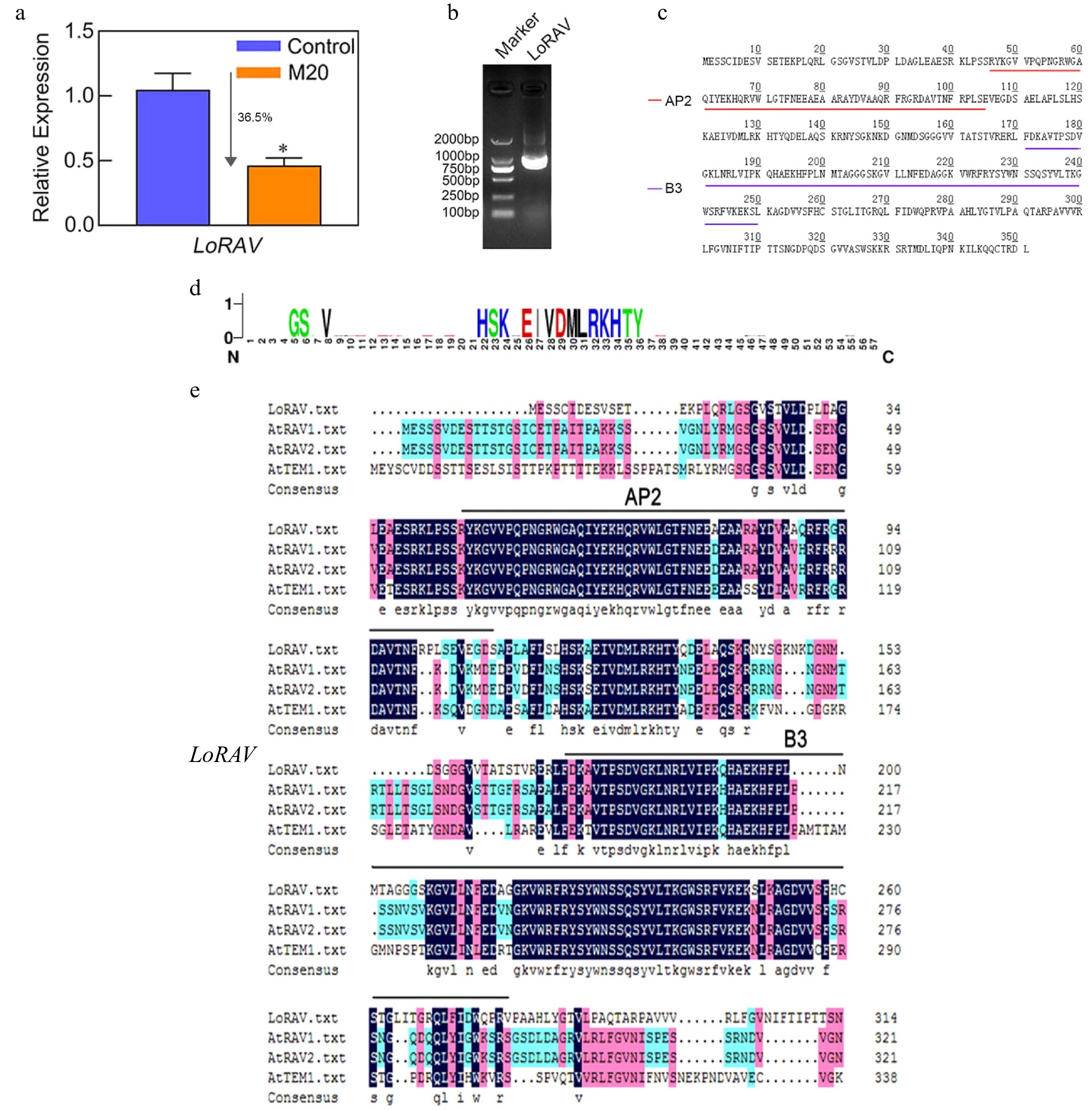
Figure 3.
LoRAV expression, cloning, and amino acid sequence analysis. (a) The expression of the LoRAV gene in bulblets on day 10 in the control and melatonin-treated groups. The expression levels were calculated relative to those of Lilium FP mRNA. Blue and orange boxes indicate the control and melatonin-treated groups, respectively. Values are presented as the means of three replicates ± SD. Data were analyzed using one-way ANOVA followed by Duncan's test; * indicates a significant difference (P < 0.05). (b) The product of LoRAV cloning; the left lane is a 2000 bp DNA marker. (c) The amino acids of LoRAV; red and purple lines indicate the AP2 and B3 structural domains, respectively. (d) LoRAV-like ubiquitinated reciprocal motifs predicted by the Cuckoo work group website. (e) Multiple sequence alignment of LoRAV with RAV family member genes of Arabidopsis.
LoRAV subcellular localization and functional analysis
-
The subcellular localization of the gene was determined to further study LoRAV. As shown in Fig. 4a, the fluorescent signal in the null and control covered all tobacco leaf cells, while the 35S:LoRAV-GFP fusion gene appeared only in the nucleus, indicating that LoRAV was a nuclear gene. We explored the function of LoRAV based on a transient transformation system in lily. To determine the stability of the transient transformation system, we first transiently transformed the lily scales with pRI101-LoRAV-GFP. As shown in Fig. 4b, fluorescence was detected after coculture for 3 d. Next, we transiently transformed lily scales with pRI101-LoRAV. As shown in Fig. 4c, after transient overexpression of LoRAV (LoRAV-OE), the expression of the LoRAV, LoAMY (alpha amylase gene, encoding a starch degradation enzyme) and LoTCP4 genes increased 4.2-fold, 1.3-fold, and 9.8-fold, respectively, compared with the control group (WT), while the expression of the LoAGPase, LoCYCB2-1, and LoPCNA genes decreased 75%, 97% and 85.3%, respectively. When the scales of LoRAV-OE were treated with 20 μM melatonin (MT-LoRAV-OE), the expression of the LoRAV and LoTCP4 genes was down-regulated by 37.2% and 50.7%, respectively, and the expression of LoCYCB2-1 and LoPCNA increased 96.2-fold and 4.7-fold, respectively, compared with the scales in LoRAV-OE, but melatonin had no effect on LoAMY and LoAGPase. These data indicated that melatonin and LoRAV have opposite regulatory effects on the proliferation of bulblets.
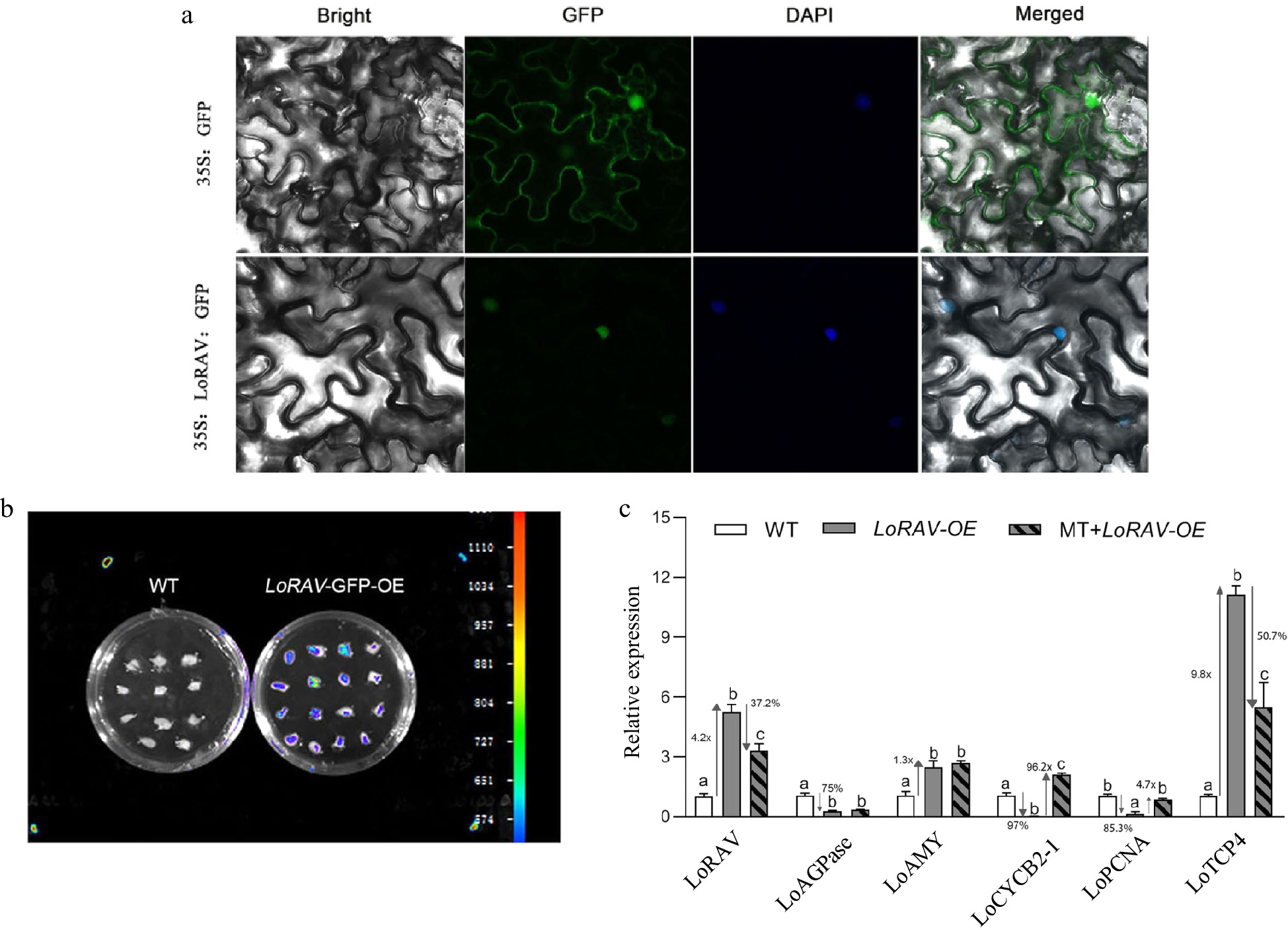
Figure 4.
Subcellular localization and functional analysis of LoRAV. (a) Transient transformation of tobacco leaves with pRI101-LoRAV-GFP; 35S:GFP served as a control. (b) Fluorescence detection after transient transformation of scales with pRI101-LoRAV-GFP. (c) The expression of cell proliferation and starch metabolism genes in bulblets with transient overexpression of LoRAV. White boxes indicate wild type (WT), gray boxes indicate LoRAV transient overexpression (LoRAV-OE), and slash boxes indicate that the LoRAV transient overexpression strain received 20 μM melatonin treatment (MT-LoRAV-OE). The expression levels were calculated relative to those of Lilium FP mRNA. Values are presented as the means of three replicates ± SD. Data were analyzed using one-way ANOVA followed by Duncan's test. Different lowercase letters indicate a significant difference (P < 0.05), and the same lowercase letters indicate no significance.
Effects of melatonin on LoBPM3 expression and LoBPM3 clone analysis
-
A gene encoding the BTB/POZ-MATH protein LoBPM3 was screened in the lily transcriptome database[27]. As shown in Fig. 5a, the expression of the LoBPM3 gene in lily bulbs increased approximately 2.4-fold compared with the control group when the bulbs were treated with melatonin for 24 h, suggesting that melatonin induces the expression of the LoBPM3 gene. Cloning and sequence analysis showed that LoBPM3 contains a complete ORF of 1248 bp, encoding 415 amino acids, with a protein molecular weight of 46.045 kD and a theoretical isoelectric point (pI) of 5.92. The protein sequence analysis showed that LoBPM3 belongs to the BPM protein family and contains two structural domains, MATH and BTB-POZ, at amino acids 39–172 and 194–313, respectively (Fig. 5b, c). We performed a multiple sequence alignment of LoBPM3 with all BPM family member genes of Arabidopsis, They are also highly conservative (Fig. 5d), this further confirms that the gene we cloned is correct. We also performed subcellular localization of LoBPM3, as shown in Fig. 5e, the fluorescent signal in the null and control covered all tobacco leaf cells, while the 35S:LoBPM3-GFP fusion gene appeared only in the nucleus, indicating that LoBPM3 was a nuclear gene.
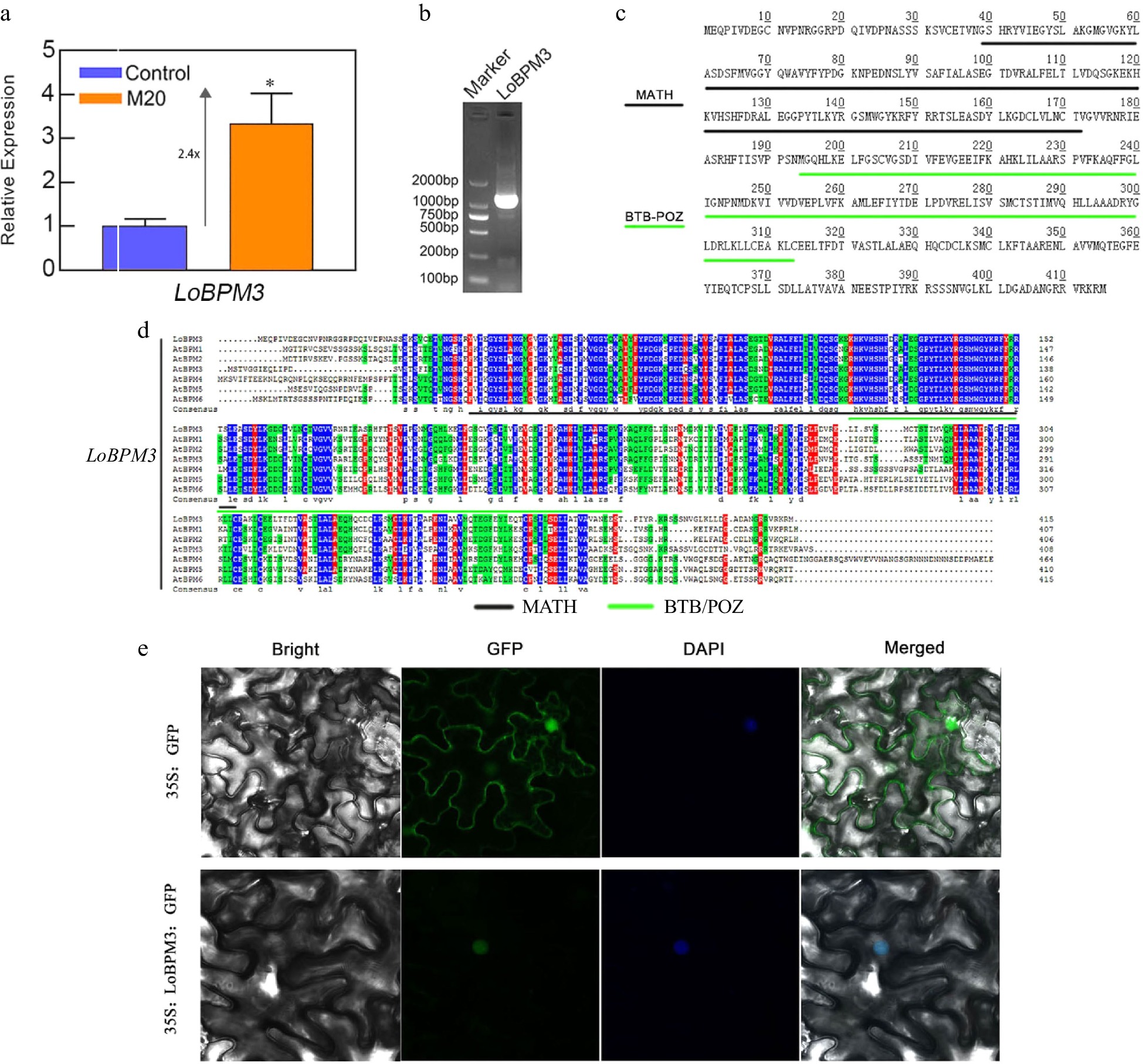
Figure 5.
The expression of LoBPM3, cloning, and amino acid sequence analysis. (a) The expression of the LoBPM3 gene in bulblets after 24 h of treatment of the control and 20 μM melatonin groups. Blue and orange boxes indicate control and 20 μM melatonin treatments, respectively. The expression levels were calculated relative to those of Lilium FP mRNA. Values are presented as the means of three replicates ± SD. Data were analyzed using one-way ANOVA followed by Duncan's test; * indicates a significant difference (P < 0.05). (b) The product of the LoBPM3 clone; the left lane is a 2000 bp DNA marker. (c) The amino acids of LoBPM3, black and green lines indicate the MATH and BTB-POZ structural domains, respectively. (d) Multiple sequence alignment of LoBPM3 with BPM family member genes of Arabidopsis. (e) Transient transformation of tobacco leaves with pRI101-LoBPM3-GFP; 35S:GFP served as a control.
Correlation analysis of LoBPM3 and LoRAV
-
LoBPM3 and LoRAV were inserted into the pGBKT7 vector (empty BD) to analyze their self-activation. Then, yeast receptor Y2H cells were transformed with pGBKT7-53 and pGADT7-T cotransformants as a positive control and pGBKT7 null transformants alone as a negative control. The results are shown in Fig. 6a. The positive control pGBKT7-53+pGADT7-T transformants grew normally in SD/-Trp-His-Ade medium, and pGBKT7-LoRAV, pGBKT7-LoBPM3, and the negative control pGBKT7 transformants failed to grow, indicating that neither LoRAV nor LoBPM3 proteins have self-activation activity. Furthermore, pGBKT7-LoRAV was used as a decoy protein, while LoBPM3 was inserted into the pGADT7 (empty AD) vector as a target protein. The respective transformants were coated on two-deficient (-T-L) and four-deficient (-T-L-A-H) plate media containing X-α-GAL, and only pGBKT7+LoRAV+pGADT7-LoBPM3 transformants on the four-deficient plate (-T-L-A-H) and positive control pGBKT7-53+pGADT7-T grew and stained blue with X-α-GAL (Fig. 6b). This finding indicates that LoRAV interacts with LoBPM3 in yeast cells.
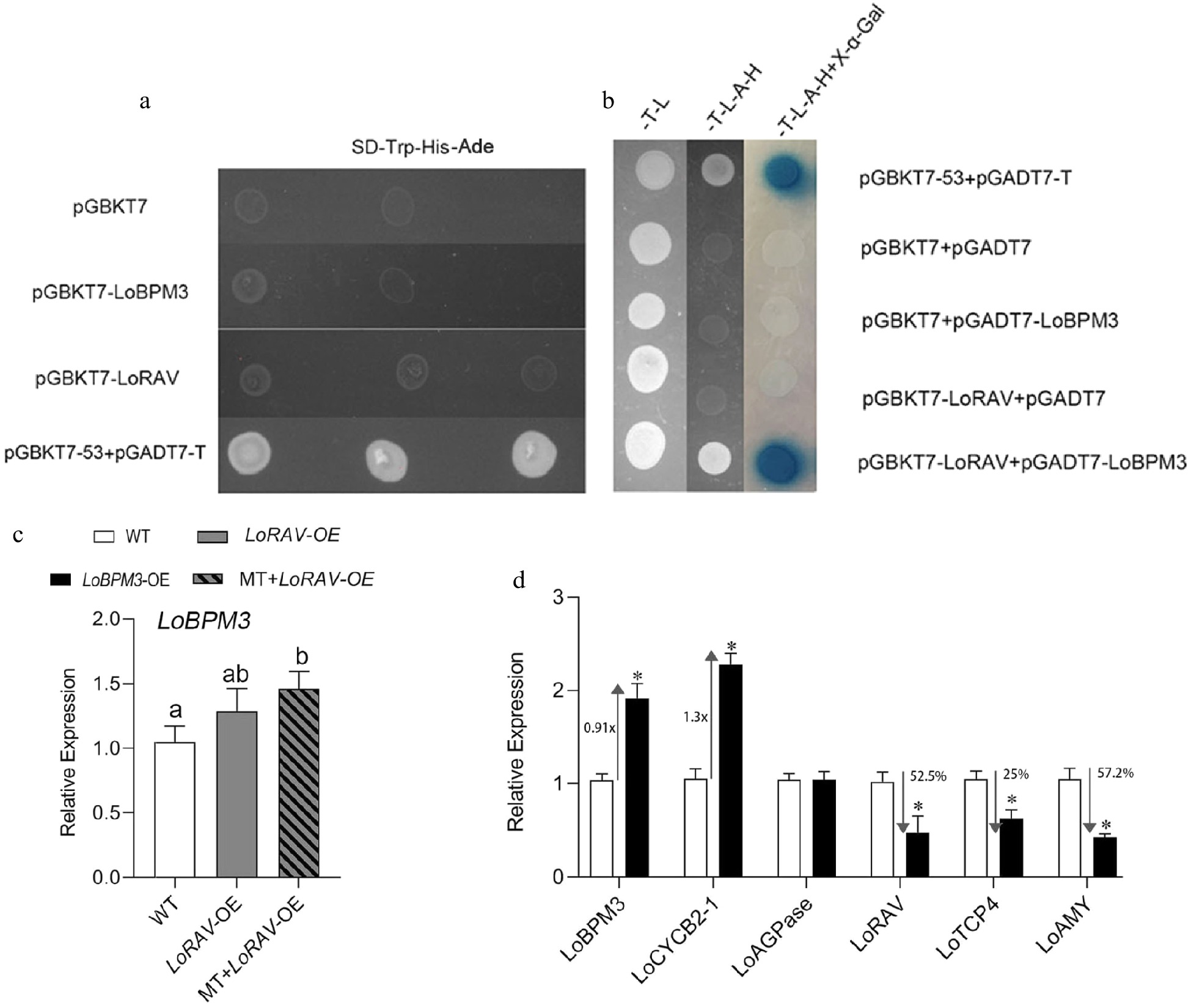
Figure 6.
Analysis of the transcriptional activation of LoRAV and LoBPM3 and their correlation. (a), (b) Fragments of LoRAV and LoBPM3 were ligated into the pGBKT vector (BD, binding domain), and LoBPM3 was ligated into the pGADT7 vector (AD, activation domain). -T-L, SD medium lacking Trp and Leu; -T-L-A-H, SD medium lacking Trp, Leu, His and Ade; -T-L-A-H+X-α-Gal, -T-L-A-H medium containing X-a-gal, and pGBKT7+pGADT7 were used as negative controls (Empty BD+Empty AD). pGBKT53 and pGADT7-T were used as positive controls. The blue plaque indicates the interaction between two proteins. (c) Upstream and downstream relationship between LoRAV and LoBPM3. White boxes indicate the wild-type (WT) strain, gray boxes indicate LoRAV transient overexpression lines (LoRAV-OE), and slashed boxes indicate LoRAV transient overexpression lines that received 20 μM melatonin treatment (MT-LoRAV-OE). (d) The expression of cell proliferation and starch metabolism genes in bulblets with transient overexpression of LoBPM3. Black boxes indicate the LoBPM3 transient overexpression strain (LoBPM3-OE). The expression levels were calculated relative to those of Lilium FP mRNA. Values are presented as the means of three replicates ± SD. Data were analyzed using one-way ANOVA followed by Duncan's test. Different lowercase letters and * indicate a significant difference (P < 0.05), and the same lowercase letters indicate no significance.
Further analysis of the regulatory relationship between LoBPM3 and LoRAV. As shown in Fig. 6c, overexpression of LoRAV alone (LoRAV-OE) had no effect on LoBPM3 expression, but the expression of LoBPM3 was further upregulated when LoRAV-OE was treated with melatonin (LoRAV-OE+MT). Meanwhile, the expression of LoRAV decreased 52.5% after transient overexpression of LoBPM3 (LoBPM3-OE) (Fig. 6d) compared with the control group (WT). These results suggest that melatonin does not regulate LoBPM3 through LoRAV and that LoBPM3 negatively regulates LoRAV. In addition, we also found that the expression of LoBPM3 and LoCYCB2-1 increased 0.91-fold and 1.3-fold, respectively, and the expression of LoAMY and LoTCP4 decreased by 25% and 57.2%, respectively, in the LoBPM3-OE group compared with the WT (Fig. 6d). However, the expression of LoAGPase was not altered after transient overexpression of LoBPM3 (Fig. 6d). These data indicated that LoBPM3 positively regulates cell proliferation.
-
In China, yearly, approximately 90% of commercial lily bulblets are imported from the Netherlands, Chile and other countries, and this high commodity dependence hinders the development of domestic seed resources[28]. As countries become more tolerant of the entry of genetically modified horticultural products into the market, biobreeding techniques are receiving increasing attention. Therefore, a screen for key genes regulating bulb development is particularly important for our attempts to improve lily seed bulb traits in a targeted manner through biobreeding.
Melatonin is beneficial for the development of lily bulblets
-
Previously, we confirmed that 20 μM melatonin promotes the initiation of lily bulblet scale primordia and starch accumulation[13]. It should be noted that previous experiments were performed in a coconut bran substrate cultivation environment, which is nutrient restricted. The authors did not address whether melatonin works only under specific conditions or has broad spectrum effects. In this study, the medium provided rich nutrients for bulb development. We found that the lily bulblets grew to a larger circumference by 20 d after the addition of melatonin to the control medium and grew new scales earlier (Fig. 1). PCNA (proliferating cell nuclear antigen gene)[29] and CYCB2-1 control DNA replication and G2/M transformation in the cell cycle[30], respectively, and are marker genes for cell proliferation. The cyclin-dependent protein kinase inhibitor gene KRP3[31], and the class II TCP genes[32] mainly inhibit cell proliferation. In this study, the expression of LoPCNA and LoCYCB2-1 was upregulated in the early stages by melatonin (Fig. 2a, b), and the expression of LoKRP3 and LoTCP4 was inhibited (Fig. 2c, d). This is consistent with our observed phenotype that the inner scale grows faster in the melatonin-treated group.
Trehalose-6-phosphate phosphatase, encoded by the TPP gene, is the key enzyme in the pathway for trehalose synthesis[33]. AMY (alpha amylase) is mainly responsible for the degradation of starch, and AGPase (ADP-glucose pyrophosphorylase) is a key enzyme in the rate of starch accumulation[34]. The activity of AGP is activated by trehalose, which in turn promotes starch accumulation in plants[25]. In this study, the content of starch and trehalose were increased by day 10 (Fig. 2e, f), and the expression of LoAGPase and LoTPP was upregulated (Fig. 2g, h). These results suggest that melatonin promoted the accumulation of bulb carbohydrate and thus increases the circumference of the bulb.
We suggested that melatonin is beneficial for bulb development via possibly broad-spectrum mechanisms both in the presence or absence of external nutrition, and melatonin is associated with the activation of at least two pathways: cell proliferation and starch metabolism. This conclusion was not drawn in previous studies. Unlike previous experiments, we did not observe root systems in lily bulbs in this study, which may be related to the high levels of cytokinin present in the medium, as higher cytokinin to growth hormone ratios are detrimental to root development[35].
The RAV gene is a negative regulator of bulb cell proliferation
-
Previous experiments suggested that melatonin promotes bulblet development, but major regulators of this process were not identified[13]. In this study, we isolated and identified a gene that participates in the regulation of cell proliferation. The RAV gene is characterized by the inclusion of two structural domains, AP2 and B3, which we confirmed by cloning and sequence analysis (Fig. 3b, c). Surprisingly, LoRAV, although it was localized in the nucleus, has no self-activation activity (Fig. 4a, 6a), suggesting that LoRAV cloned in lily is a transcriptional repressor gene, which may be related to the fact that its protein sequence contains the RLFGV motif, a conserved motif in transcriptional repressors[36]. We suggest that LoRAV is a negative regulator of bulb cell proliferation, and there are several lines of evidence. Firstly, AtRAV1 inhibited the expression of the endosperm cell proliferation genes MINI3 and IKU2 directly in Arabidopsis, limiting seed size[21]; GmRAV controlled the expression of the cell cycle genes GmE2F and GmCYCD3, directly or indirectly[20]; and the domain of LoRAV was highly conserved (Fig. 3e ). Secondly, the expression of cell proliferation-promoting genes (LoCYCB2-1, LoPCNA) were completely inhibited after transient overexpression of LoRAV, and the expression of LoTCP4 was upregulated significantly, but the expression of LoCYCB2-1, LoPCNA, and LoTCP4, but not LoAMY and LoAGPase, was reversed when LoRAV-OE was treated with melatonin (Fig. 4c). These findings show that the function of LoRAV is mainly related to mediating cell proliferation.
LoBPM3 is upstream of LoRAV and negatively regulates LoRAV
-
In this study, we also screened a protein that interacts with LoRAV in lily. Since the whole genome work of the lily has not been completed, we only retrieved a gene encoding MATH-BTB/POZ protein in the transcriptome database[27]. LoBPM3 was cloned successfully in lily through sequence feature analysis (Fig. 5b, c). Interestingly, LoBPM3 is conserved with all member genes of the BPM family of Arabidopsis, and localized in the nucleus (Fig. 5d, e), suggesting that the cloned LoBPM3 in lily may have the biological function of assembing with CULLIN3-base E3 ligase. We found that treatment of lily bulbs with melatonin for 24 h induced LoBPM3 expression (Fig. 5a) and that LoBPM3 interacted with LoRAV in yeast (Fig. 6b), suggesting that melatonin-mediated regulation of LoRAV expression is at least partially related to LoBPM3. Notably, we showed that LoBPM3 may be a positive regulator of cell proliferation because the expression of LoCYCB2-1 was upregulated and the expression of LoTCP4 was downregulated by the overexpression of LoBPM3 (Fig. 6d). We also found that the expression of LoRAV was regulated negatively by overexpression of LoBPM3, but the expression of LoBPM3 was not regulated by overexpression of LoRAV (Fig. 6c). These results indicated that LoBPM3 is upstream of LoRAV and partially weakens the effect of LoRAV. The regulation of LoRAV by LoBPM3 maybe occurs not only at the transcriptional level; previous studies have shown that BPM3 proteins modify target proteins mainly through the ubiquitination pathway, such as MYB56[37], MYC2, MYC3, and MYC4[38]. In the present study, a conserved ubiquitination-like interaction motif, VDML (Fig. 3d), was identified in LoRAV proteins, but whether the LoBPM3 protein negatively regulates LoRAV gene expression by degrading LoRAV proteins through the ubiquitination-like pathway requires further exploration.
-
In summary, we revealed a working module, namely, LoBPM3-LoRAV, that regulates lily bulb development by mediating cell proliferation, we confirmed that LoRAV is a negative regulator of bulb cell proliferation, and we showed that the suppression of LoRAV expression is related to LoBPM3, which was not reported in previous studies of lily. We propose the following model. As shown in Fig. 7, without the addition of melatonin, the inhibitory factor of cell proliferation occupies its main position, the promoting factor of cell proliferation is suppressed, and the bulb grows slowly. When melatonin is administered, some ubiquitination-related genes (such as LoBPM3) come into play, reversing the above situation, weakening the inhibitory factor of cell proliferation, and promoting factor of cell proliferation occupy the main position, and the cells begin to proliferate actively, thereby promoting bulb development.
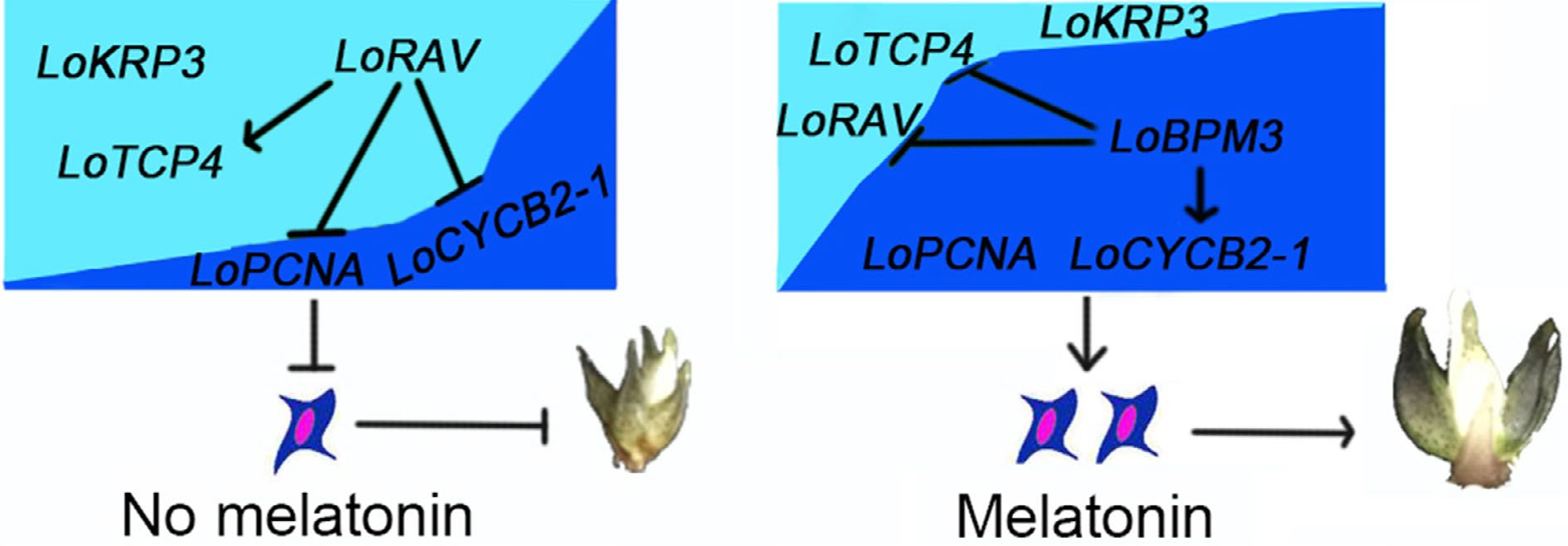
Figure 7.
A working model of melatonin regulating bulb development by mediating cell proliferation. Dark blue and arrows indicate positive interactions, and light blue and flat lines indicate negative interactions.
-
Sterile 'Siberia' bulblets (diameter ≥ 1 cm) were used as the test material, and the material was preserved for use in this laboratory. The reagents included melatonin purchased from Macleans Biotech (Shanghai, China) and 6-benzyladenine (6-BA), naphthalene-1-acetic acid (NAA), agar powder, and sucrose purchased from Sinopharm (Beijing, China). pRI101-ON was purchased from Beijing Bao Ri Doctor Biotechnology Co (Beijing, China). pRI101-GFP is the pRI101-ON vector containing an ORF fragment of GFP between the BamH I and Sac I restriction enzyme sites.
Treatment with melatonin
-
Sterile lily bulblets of the same size were selected and inoculated into 30 ml of control media or 30 ml of M20 media and then placed in a completely dark environment at 25 ± 1 °C for 20 d. The control media was MS (Murashige-Skoog) + 90 g L−1 sugar+7 g L−1 agar + 2 mg L−1 6-BA + 0.5 mg L−1 NAA, and the M20 media was control media supplemented with 20 μM melatonin. The high concentration of sucrose was beneficial for bulb development, and the melatonin concentration (20 μM) was previously determined through preliminary experiments[13]. Three lily bulblets were inoculated in each bottle of medium, and 10 groups were established for each treatment containing three replicates.
Morphological observation and index determination
-
Photographs were captured 7, 10, 14, and 20 d after inoculation to confirm the growth status of lily bulblets. Lily bulblets cultured for up to 20 d were selected, and their circumference, diameter, and height were measured. The lily bulblets were photographed after removal, and the middle width of the bulblets was measured using Image J (Version 1.8.0) software and calculated according to the circumference equation L = πd to determine the circumference. The height of the bulblets was determined from the base of the bulblets to the tip of the middle scale. The starch and trehalose concentrations were determined from randomly selected bulblets cultured for up to 10 d using the anthrone colorimetric technique, strictly according to the instructions of the physiological and biochemical assay kit from Suzhou Ke Min Biotechnology Co. (Jiangsu Province, China).
Total RNA extraction, reverse transcription polymerase chain reaction (PCR) and real-time quantitative PCR assays
-
Bulblets were cultured for up to 10 or 20 d before RNA extraction. Total RNA was extracted using a HiPure HP Plant RNA Mini Kit (Miki, China), and the RNA quality and concentration were determined using 1.0% agarose gel electrophoresis and an Infinite® 200 Pro nucleic acid protein detector (Tecan, Mannedorf, Switzerland). First-strand cDNA synthesis was performed using a Tiangen FastKing cDNA First Strand Synthesis Kit (Tiangen, China) according to the manufacturer’s instructions.
Sequence information of candidate genes was obtained after the retrieval of previously completed lily transcriptome data (NCBI Accession number: PRJNA385210)[27]. An NCBI BlastX analysis of conserved amino acid sequences was performed. Primer 3.0 software was used to design primers (Supplemental Table. S1), and the gene nomenclature was referenced[39]. qRT–PCR analysis was performed using the QuantStudio 3 Real-Time PCR System with a reaction volume of 20 μL: 1 μL of cDNA, 1 μL of forward primer, 1 μL of reverse primer, 7 μL of ddH2O, and 10 μL of 2x SYBR Green qPCR Master Mix (LowROX) (Biomake). The reaction conditions were as follows: predenaturation at 95 °C for 10 min, denaturation at 95 °C for 15 s, 58 °C for 30 s, and 72 °C for 30 s (40 cycles). The FP gene was used as an internal reference[40], and gene expression was calculated using the 2−ΔΔCᴛ method. Each sample included three biological replicates and three technical replicates.
LoRAV and LoBPM3 expression, cloning and bioinformatics analysis
-
Bulblets cultured for up to 10 d and after 24 h of melatonin treatment were selected to test the effects of melatonin on LoRAV and LoBPM3 gene expression, respectively. Cloning primers LoRAV-full-F, LoRAV-full-R; LoBPM3-full-F, and LoBPM3-full-R were designed based on the LoRAV and LoBPM3 transcript sequences (Supplemental Table S1). PCR amplification was performed using the synthesized cDNAs as templates, and the amplified products were detected by electrophoresis on a 1.0% agarose gel. The target fragment was purified using the Novozymes Gel Recovery/DNA Purification Kit (DC301) and cloned into the pMD19-T vector, and the bacteriophage was verified by PCR after the transformation of E. coli and sent to the company for sequencing. Protein signatures were analyzed using ExPASy (
https://web.expasy.org/protparam/ ), multiple amino acid sequence comparisons were performed using DNAman, and the Cuckoo workgroup online website (www.biocuckoo.org ) and WebLogo (http://weblogo.threeplusone.com ) were used to predict and analyze ubiquitination interaction motifs.LoRAV and LoBPM3 subcellular localization
-
Using the correctly sequenced plasmids of LoRAV/LoBPM3 as templates, PCR amplification was performed using the attached Nde I and BamH I subcellular localization primers (Supplemental Table S1), and the amplification products from which the terminator was removed were ligated to the pRI101-GFP vector and then transformed into Agrobacterium tumefaciens EHA105. A. tumefaciens infestation solution was prepared and applied to tobacco leaves. After 2 d of culture under weak light, the infested parts of the leaves were removed, observed under a laser confocal microscope, and photographed.
Analysis of LoRAV and LoBPM3 self-activation activity and yeast two-hybrid assay
-
The analysis of self-activation activity and yeast two-hybrid assays were performed according to the instructions provided with the Coolaber Classic Yeast Transformation Kit. PCR amplification was performed using yeast vector primers (Supplemental Table S1) with attached Nde I and BamH I digestion sites, and the amplification products were ligated to pGBKT7 and pGADT7 vectors to construct LoRAV-BD, LoBPM3-BD and LoBPM3-AD fusion expression vectors. LoRAV-BD and LoBPM3-BD fusion vectors and empty pGBKT7 were transformed into Y2H yeast receptor cells and plated on SD/-Trp-His-Ade medium for the self-activation activity assay. LoBPM3-AD, LoRAV-BD, pGBKT7 and pGADT7 fusion expression vectors were cotransformed into Y2H yeast receptor cells, plated on SD-Trp-Leu and SD-Ade-His-Trp-Leu media, stained with X-a-Gal and used for reciprocal validation. Additional cotransformations were performed with pGBKT7-53 and pGADT7-T as positive controls.
LoRAV and LoBPM3 instant conversion
-
PCR amplification was performed using the overexpression vector primers LoRAV-OE-F, LoRAV-OE-R; LoBPM3-OE-F, LoBPM3-OE-R (Supplemental Table S1) with attached Nde I and BamH I restriction enzyme sites, and the amplification products were ligated to the pRI101-ON vector. Single colonies containing the target gene were screened and inoculated into 1 mL of YEB broth containing 50 mg mL−1 kanamycin and 50 mg mL−1 rifampicin and incubated overnight at 28 °C with shaking. This overnight culture was added to 50 ml of fresh YEB solution containing 50 mg mL−1 kanamycin and 50 mg mL−1 rifampicin and incubated until the OD value was 0.8. Then, the bacteria were centrifuged at 25 °C for 10 min at 5,000 rpm and incubated with 50 mL of a 1/2 MS (without NH4NO3) + 100 μM L−1 acetosyringone + 60 g L−1 sucrose mixture to resuspend the precipitated bacterium, which was the Agrobacterium infestation solution.
The scales of small lily bulblets were peeled and cut into 0.5 cm2 sized squares, inoculated into preculture medium (MS + 0.2 mg L−1 NAA + 30 g L−1 sucrose + 7.0 g L−1 agar), removed after 3 d of preculture, and placed in Agrobacterium infestation solution for 15 min, with 30 scales per treatment. After treatment, the transformed scales were inoculated into coculture medium (MS + 0.2 mg L−1 NAA + 100 μM L−1 AS + 30 g L−1 sucrose + 7.0 g-L−1 agar), and after 3 d of incubation, the scales that transiently overexpressed the LoRAV gene were selected and treated with 30 mL of 20 μM melatonin or distilled water for 24 h. Then, the total RNA was extracted from the scales for subsequent studies.
Data analysis
-
Data were analyzed using IBM SPSS Statistics (SPSS 22.3 Inc., Chicago, IL, USA) using one-way ANOVA followed by Duncan’s test. Differences were considered significant at P < 0.05.
This work was supported by grants from the National Key R&D Program of China (Grant No. 2018YFD1000407), the earmarked fond for CARS (Grant No. CARS-23) and LiaoNing Revitalization Talents Program (Grant No. XLYC2002052). We thank Prof. Aide Wang (Shenyang Agricultural University, China) for providing the pGBKT7 and pGADT7 plasmids for the experiments.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Summary of primers used in this study.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Sun L, Song S, Yang Y, Sun H. 2022. Melatonin regulates lily bulblet development through the LoBPM3-LoRAV module. Ornamental Plant Research 2:12 doi: 10.48130/OPR-2022-0012 

Melatonin regulates lily bulblet development through the LoBPM3-LoRAV module
- Received: 28 April 2022
- Accepted: 10 August 2022
- Published online: 29 August 2022
Abstract: Melatonin regulates lily bulblet development; however, the key interactions have not yet been determined. In this study, we found that adding 20 μM melatonin to the growth medium increased the bulblet circumference and the trehalose and starch content. Moreover, the expression of numerous genes involved in cell proliferation and carbohydrate accumulation were upregulated. Further analysis showed that melatonin downregulated the expression of LoRAV and upregulated the expression of LoBPM3 at an early stage. Cloning and bioinformatics analysis showed that LoRAV is 1056 bp in length and encodes a protein with two structural domains, namely, AP2 and B3; the encoded protein is localized to the nucleus and has no self-activation activity, and this protein interacts with LoBPM3. The expression of LoTCP4 and LoAMY was upregulated, while the expression of LoAGPase, LoCYCB2-1, and LoPCNA was inhibited when LoRAV was transiently overexpressed. However, the trends in the expression of the genes mentioned above, except LoAGPase and LoAMY, were reversed when LoRAV-OE was treated with melatonin. Interestingly, the expression of LoRAV, LoTCP4 and LoAMY was inhibited, and the expression of LoCYCB2-1 was upregulated by transient overexpression of LoBPM3, but the expression of LoBPM3 was not regulated by LoRAV overexpression. Based on these data, we suggest that melatonin promoted bulb development, which was partly associated with regulation of the LoBPM3-LoRAV module; thus, LoRAV negatively regulates cell proliferation of bulblets, and LoBPM3 is located upstream of LoRAV and attenuates its expression.
-
Key words:
- Lilium /
- Melatonin /
- Bulblet development /
- LoBPM3-LoRAV module