









-
Red clover (Trifolium pratense) is a perennial species of flowering plant in the family Fabaceae or Leguminosae. Red clover is native to Europe, Western Asia, and northwest Africa, but it is also grown widely in the temperate regions such as Pacific Northwest and the US[1]. It is a quick growing crop adapted to varied environmental conditions, which can be used as an important winter annual in the Deep South[2]. Its finer roots have nodules containing Rhizobia bacteria, with the capacity of fixing nitrogen. Red clover can capture nitrogen (150−250 kg N/ha/year) from the air and feed it into the soil, enriching the soil fertility and providing nourishment to surrounding plants in between seasons[3]. Red clover is also used as a cover crop. The aims include preventing nutrient leaching, reducing soil erosion as well as replacing pesticides to control pests and phytopathogenic diseases in addition to producing pasture mixture, hay, and silage[4]. These advantages of red clover show great value in sustainable agriculture.
According to the EU Novel Food Regulation, red clover is allowed to be used as a food supplement, but not as food or food ingredients[5]. Extracts of red clover flowers have been used extensively for a variety of health protective purposes, such as promoting wound healing and maintenance of bone health[6]. Additionally, red clover flower showed potential value in alternative therapies for menopausal problems, cardiovascular disease, and a range of hormone-dependent diseases such as breast cancer. Several studies indicated that these health-beneficial effects of red clover are derived from secondary plant metabolites belonging to polyphenols with emphasis on the isoflavonoid compounds[6−9]. Bioactive compounds, such as flavonoids, from plant seeds raised a lot of interest[10]. These compounds play active roles reducing the risk of health problems associated with oxidative stress, such as coronary heart disease, and cancer initiation and progression[11]. Although the beneficial compounds have been reported in the seeds of several Trifolium species, the study of Trifolium pratense is little[12]. Currently, wild clover seeds are commonly used as animal feed though they are sources of many nutrients, especially with high protein content (> 16.0% DM)[13].
Oils extracted from Trifolium seeds have shown various health benefits due to high concentrations of bioactive lipids and tocopherols[14]. Clinical studies demonstrated the therapeutic effects of polyunsaturated fatty acids on cardiovascular diseases, Alzheimer’s disease, depression, and various other degenerative neurological disorders[15]. In addition to fatty acids, tocopherols are also important bioactive compounds in plant seed oils. As lipophilic antioxidants, tocopherols show positive effects in protecting polyunsaturated fatty acids from peroxidation[14]. Although red clover leaves were used to produce essential oils, red clover seeds (RCS) oil has been rarely studied[16]. Compared to Soxhlet extraction, supercritical fluid extraction (SFE) has many advantages. SFE provides solvent-free extracts with high selectivity of the targeted components[17]. Extraction using supercritical CO2 is simple and eco-friendly since carbon dioxide is a non-toxic, inert gas with low price. These superiorities make supercritical CO2 extraction a promising method of recovering valuable compounds from plant materials for food and pharmaceutical application[18].
Therefore, it is of great importance to gain an in-depth understanding of the phytochemical profile of RCS and SFE-extracted oil due to their richness of health-benefiting compounds. Recent studies of RCS were limited to measuring the total content of specific group of compounds using colorimetric method[14, 19]. Yet, to explore the potential of RCS for application in food and health care products, it is important to obtain thorough knowledge on the composition of different groups of nutrients and bioactive compounds of RCS at molecular level. This study aims to comprehensively identify the metabolites of RCS and its SFE-extracted oil. The novelties of our study lie in: (1) new findings on comprehensive phytochemical profiles of the study materials using a non-targeted metabolomics approach and; (2) identification and quantification of the compound groups with nutritional, sensorial, and health-promoting properties using targeted mass spectrum (MS) approach. To the best of our knowledge, this is the first report on the thorough investigation of the chemical compositions of RCS and SFE-extracted RCS oil.
-
Organic RCS were purchased from the local store Hankkija in Turku Finland. Trimethylsilyl was purchased from Thermoscientific (Bellefonte, US). Sodium hydroxide, boric acid, potassium chloride, acetyl chloride and potassium carbonate were provided by Sigma-Aldrich (Darmstadt, Germany). The reference standards used in this study included sugars, organic acids, tocopherols, isoflavonoids and flavonoids (Sigma-Aldrich, St. Louis, MO, US). The internal standard of fatty acids was bought from Larodan AB (Solna, Sweden). Other chemicals of liquid chromatography (LC) and MS grade were purchased from Honeywell Riedel-de Haën Co. (Seelze, Germany).
SFE-extracted RCS oil
-
Oil was extracted from RCS by a pilot-scale SFE facility (Chematur Engineering, Karlskoga, Sweden) at Aromtech Ltd (Tornio, Finland). RCS (470 g) were milled and loaded into the extraction vessel. The extraction was carried out for 120 min with CO2 at a flow rate of 0.4 L/min using ethanol (7 g/min) as co-solvent for extraction. The extraction temperature and pressure were 40 °C and 200 bar, respectively. The separation pressure was 50 bars, and the separation temperature was 30 °C. After the extraction, the oil was separated from ethanol by centrifugation and in storage at −80 °C until further analysis.
Dry matter content of RCS
-
The RCS were ground using a mortar and pestle with addition of liquid nitrogen. Approximately 1 g RCS powder was used for drying in the oven at 105 °C until it reached constant weight.
Determination of total content of proteins
-
The total protein content of RCS was determined by a Kjeldahl autoanalyzer (Foss Tecator Ab, Höganäs, Sweden), which was calculated using a conversion factor of 6.25 (Protein content = nitrogen content × 6.25)[20].
Analysis of sugars and organic acids
-
RCS powder (4 g) was extracted with 70% ethanol by 20 min ultrasonication for three times. Each time, the supernatants were separated by centrifuging (15 min, 1,500× g at 4 °C) and collected. The volume of combined supernatants was set to 50 mL with extraction solvent. One milliliter of the supernatant was filtered (0.22 μm PTFE syringe filter, Phenomenex) for gas chromatography (GC) analysis.
The identification and quantification of sugars and organic acids were conducted by an internal standard method in our previous research[21]. The chromatographic system Shimadzu GC-2010 consisted of flame ionization detector (Shimadzu corp., Kyoto, Japan) using a SPB-1 column (30 m × 0.25 mm i.d., 0.25 µm, Supelco, Bellefonte, PA, US). Briefly, RCS extract (0.4 mL) was mixed with 0.2 mL of sorbitol and tartaric acid. The sample was dried by nitrogen flow and mixed with trimethylsilyl (600 µL). The correction factors of the study sugars and organic acids were calculated using the corresponding commercial standards. The identification was based on the comparison of the compound retention times of our samples with those of external standards.
Identification and quantification of phenolic compounds
-
The aqueous ethanolic RCS extract described was used for the analysis of phenolic compounds using our previous method[21]. An ultra-high performance liquid chromatography (UPLC) system equipped with a quadrupole time-of-flight tandem mass spectrometer (Q-TOF) (Bruker Corp., Billerica, MA, US) was applied for the identification using an Aeris Peptide XB-C18 column (150 mm × 4.60 mm, 3.6 μm, Torrance, CA, US). The injection volume was 10 μL. The total flow rate was 1 mL/min. The chromatogram was recorded under 260 nm (for isoflavonoids) and 280 nm, 360 nm (for other flavonoids). MS flow rate was 0.3−0.4 mL/min. Mass full-scan was operated in a range of 20–2,000 m/z under both positive and negative ionization modes. The compounds were identified based on the retention times, UV-absorption spectra and MS spectra by comparing with those of reference compounds and referring to the library and the literatures.
The quantification of the identified phenolic compounds was performed on Shimadzu LC-30AD liquid chromatograph equipped with an SPD-M20A photodiode array detector (Shimadzu Corp., Kyoto, Japan). The same column and chromatographic conditions were used for the UPLC-QTOF analysis. The identified phenolic compounds were quantified using the standard calibration curves (Supplemental Table S1).
Fatty acids analysis
-
Fatty acids in RCS and SFE-extracted oil were determined following the procedure of Christie's research[22]. The total lipid content of RCS was studied using a modified Folch method. Briefly, all the glass tubes were rinsed with chloroform. RCS powder (approximately 100 mg) was weighed to the tube with addition of 900 µL chloroform and 1.5 mL methanol. After 1 h soaking at room temperature, the mixture was homogenized with Ultra Turrax (IKA®-Werke, Germany). Chloroform (2 mL) was added and the homogenization was continued for 2 min followed by 30 min sonication. KCl solution (0.88%, 1.2 mL) was added and vortexed for 5 s, followed by 3 min centrifuging at 3500 rpm. After collecting the lower liquid fraction, 1.5 mL chloroform was added to the tube. The sample was vortexed for 30 s before centrifuging (3 min at 3,500 rpm). The combination of lower liquid fraction with the previous fraction was dried by nitrogen flow at 50 °C. Lipid content was calculated by weighing the glass tube with lipid fraction. Lipid fraction was re-dissolved in 3 mL chloroform. For preparation of fatty acid methyl esters, the lipid solution (containing approximately 10 mg lipid) was transferred to a Pyrex tube, adding the internal standard triheptadecanoin (0.34 mg). After evaporating the solvent by nitrogen flow, acetyl chloride/methanol (1/10, v/v, 2 mL) was added. The tube was incubated at 50 °C overnight. Potassium carbonate (1 M, 2 mL) was carefully added to the tube after cooling down to room temperature. N-hexane (2 ml) was added to the tube and the sample was centrifuged (3 min, 1,000 rcf) after vortexing (10 s). The upper liquid fraction was transferred to a GC vial for analysis.
A Shimadzu GC-2030 equipped with an FID (Shimadzu Corporation, Kyoto, Japan) and DB-23 column (60 m × 0.25 mm × 0.25 μm; Agilent Technologies, J.W. Scientific, Santa Clara, CA, US) was used for the determination and quantification of fatty acids. GC conditions were set as helium flow 1.7 mL/min; 130 °C held 1 min, 6.5 °C/min to 170 °C with no hold, 3.0 °C/min to 205 °C, held for 18 min, 30 °C/min to 230 °C and held for 2 min. The peaks were identified by using external standards GLC-490 (Nu-Check-Prep, Elysian, MN, US) and 37 Component FAME mix (Supelco, St. Louis, MO, US) by comparing the retention times. The quantification was conducted by the internal standard (fatty acid 17:0). The correction factors obtained by analysis of standard mixtures were applied in the quantification of different fatty acids.
Tocopherols
-
Tocopherols were analyzed using the lipid extraction method described previously. After evaporation of chloroform by nitrogen flow at 50 °C, the lipid extract of RCS and SFE oil were dissolved in heptane for HPLC analysis. Tocopherols were analyzed by NP-HPLC-FLD with Shimadzu Nexera XR LC-30 HPLC instrument and RF-20A prominence fluorescence detector (Shimadzu, Kyoto, Japan) using an Phenomenex OOG-4162-EO Luna 3 µm silica column (250 mm × 4.6 mm, pore size 100 Å; Torrance, CA, US). The injection volume was 2 μL. Mobile phase (isocratic, 0.4 mL/min) consisted of 2% 1,4-dioxane and 98% heptane. For quantification, standard curves were prepared using α-, β- and γ-tocopherol solutions at different concentrations. The excitation wavelength was 292 nm and emission wavelength was 325 nm.
Non-targeted metabolomics
-
Sample preparation and compound identification for non-targeted metabolomics were based on previous methods with minor modifications[23]. Briefly, RCS powder (approximately 100 mg) or oil (100 µL) were added to a tube containing 400 µL of cold acetonitrile, respectively. The tubes were kept in an ice box during the sample preparation. The mixture was vertexed for 10 s and centrifuged for 5 min (7,000× g at 4 °C). The supernatants (1 mL) were filtered (0.22 μm PTFE syringe filters, Phenomenex) for further analysis.
The compound identification was performed on the same Q-TOF mass spectrometer as described previously. A Zorbax Eclipse XDB-C18 column (2.1 mm × 100 mm,1.8 µm, Santa Clara, CA, US) was used. The gradient elution were water (solvent A) and methanol (solvent B) both containing 0.1% (v/v) of formic acid. The injection volume was 2 μL. The flow rate was 0.4 mL/min. LC gradient program was followed by 0−10 min to 2%−100% solvent B, 10−14.5 min to 100% B, 14.5−14.51 min to 100%−2% B, 14.51−16.5 min to 2% B. Mass full scan was operated under both positive and negative ionization modes. MS parameters were reported in Supplemental Table S2.
Statistical analysis
-
All data were expressed as mean ± standard deviation based on dry matter content. MS DIAL (version 4.8) was applied to process the raw data from Q-TOF mass spectrometer. The parameters of MS DIAL were set as MS1 tolerance 0.01 Da, MS2 tolerance 0.025 Da, m/z range 50-1500, minimum peak amplitude 3,000 signal counts, and mass slice width 0.1 Da for peak selections. The smoothing level and minimum peak width were 3 scans and 5 scans, respectively. The selected adduct ions include: [M + H]+, [M + NH4]+, [M + Na]+, [M + K]+, [M + CH3OH + H]+, [M + ACN + H]+, [M + H - H2O]+, [2 M + H]+ for the positive mode and [M − H]−, [M + Cl]−, [M − H2O − H]−, [M + FA − H]−, [2 M − H]−, [3 M − H]− for the negative mode. The m/z tolerance was set to 0.015 Da and retention time tolerance was 0.05 min. The database MassBank was utilized in MS-DIAL for additional annotations and mass spectral comparison.
Heatmap was performed using MetaboAnalyst 5.0 (
www.metaboanalyst.ca ). Heatmap was used to analyze the semi-quantitative results of component distribution in RCS and the oil extracted by the equal weight of the seeds. -
As shown in Table 1, the total content of proteins in RCS was 6.9 g/100 g DM. Higher protein contents were reported by a previous research, in which seed protein contents of wild red clover were 14.2−17.3 g/100 g DM using Kjeldahl method[13]. Our study RCS contained 50.0 mg/100 g DM of organic acids, including citric acid and malic acid. Sugars were identified from RCS, including disaccharides (maltose and sucrose), monosaccharides (glucose, fructose, xylose, arabinose and mannose), sugar alcohols (inositol, mannitol and xylitol) and sugar acid (galacturonic acid). In our study, sucrose (2,856.2 mg/100 g DM) and fructose (534.1 mg/100 g DM) were found to be dominant in the sugars, which accounted for 77% and 14% of the total sugars (3,702.8 mg/100 g DM), respectively. To date, no report has been published on the contents and profiles of sugars in RCS despite their postive effect on gut microbe modulation and immunological properties. For instance, arabinosyl substitutional position and ratio in the xylan backbone contributes to the health-beneficial properties of arabinoxylans[24]. The arabinose to xylose (A/X) ratios of the study RCS showed that RCS can be a potential source of arabinoxylans. Arabinoxylan oligosaccharides have prebiotic functions, and they can positively modulate human gut health by promoting the growth of beneficial gut microbes and suppressing pathogenic gut microbes. The mechanism is that oligosaccharides are not digested in the upper gastrointestinal tract but are only fermented in the colon. Short chain fatty acids produced by oligosaccharides during gut fermentation decrease the pH in the colon, which inhibits the growth of pathogenic gut microbes[24,25]. Additionally, the presence of mannose showed the potential of RCS in nutraceutical and pharmaceutical applications due to its health-promoting functions of relieving constipation, upregulating blood lipid metabolism and reducing blood cholesterol and triglyceride levels[26].
Table 1. Content of proteins, lipids, sugars, and organic acids in RCS.
Composition Content Dry matter (%) 88.6 ± 0.1 Proteins (g/100 g DM) 6.9 ± 0.1 Lipids (g/100 g DM) 7.0 ± 1.1 Sugars (mg/100 g DM) 3,702.8 ± 632.7 Arabinose 40.9 ± 2.8 Xylose 13.1 ± 2.1 Xylitol 26.1 ± 4.0 Fructose 534.1 ± 96.3 Glucose 114.9 ± 19.3 Mannose 11.4 ± 1.5 Mannitol 29.4 ± 4.2 Inositol 38.6 ± 5.5 Sucrose 2,856.2 ±496.5 Maltose 31.4 ± 4.3 Galacturonic acid 6.7 ± 0.4 Arabinose to xylose ratio 3.2 ± 0.6 Organic acids (mg/100 g DM) 50.0 ± 4.5 Malic acid 14.0 ± 2.7 Citric acid 36.1 ± 1.9 Phenolic compounds
-
The phenolic compounds were characterized by MS and MS2 (Table 2). The identification was based on the fragmentation pattern, the retention time and UV absorption spectra, and the comparison of those with the commercial standards and relevant literatures. Two flavanonols (taxifolin, taxifolin hexoside), eight flavonols (quercetin hexoside 1, quercetin hexoside 2, quercetin hexoside 3, quercetin 3-O-galactoside, quercetin 3-O-glucoside, quercetin, kaempferol and isorhamnetin), one isoflavone (formononetin coumaroyl hexoside) were identified or preliminarily identified in RCS. In addition, two unknown phenolic compounds were detected.
Table 2. Identification of phenolic compounds in the RCS by UPLC-DAD-ESI-QTOF*.
No. Identification Content
(mg/100 g DM)UV λmax (nm) [M+Na]+/[M+H]+/[M-H]− (m/z) MS2 (m/z) Total phenolic compounds 228.4 ± 51.0 Flavanonols 102.2 ± 22.0 1 Taxifolin hexoside 60.0 ± 13.3 288 489.1321/467.1505/465.1069 467.1505 → 305.0989, 260.0501, 139.0380
465.1069 → 303.0518, 285.04112 Taxifolin 42.2 ± 8.7 289 327.0443/305.0623/303.0533 305.0623 → 287.0522, 259.0577, 231.0629, 195.0270, 153.0171, 149.0221
303.0533 → 285.0416, 275.0568, 259.0618, 241.0515, 217.0515, 178.9993, 153.0199, 125.0257Flavanols 101.6 ± 24.2 4 Quercetin hexoside 1 2.7 ± 0.6 255, 371 487.0793/465.0975/463.0895 465.0975→ 303.0467
463.0895→ 301.0365, 151.00485 Quercetin 3-O-galactoside 1.1 ± 0.2 255, 353 −/465.0978/463.0893 465.0978 → 303.0465
463.0893 → 300.0287, 271.0258, 255.0309, 243.03096 Quercetin 3-O-glucoside 11.7 ± 3.0 255, 353 487.0801/465.0974/463.0903 465.0974 → 303.0469
463.0903 → 300.0286, 271.0257, 255.0307, 243.03097 Quercetin hexoside 2 19.2 ± 4.5 252, 365 487.0799/465.0977/463.0887 465.0977 → 303.0471
463.0887 → 301.03559 Quercetin hexoside 3 1.3 ± 0.2 270, 369 487.0792/465.0978/463.0888 465.0978 → 303.0465
463.0888 → 301.036710 Quercetin 63.1 ± 15.2 255, 370 325.0286/303.0471/301.0377 303.0471 → 229.0473, 153.0169
301.0377 → 273.0402, 178.9992, 151.004412 Kaempferol 1.2 ± 0.2 270, 365 −/287.0523/285.0418 − 13 Isorhamnetin 1.4 ± 0.3 260, 371 −/317.0624/315.0521 − Isoflavones 1.6 ± 0.4 11 Formononetin malonyl- hexoside 1.6 ± 0.4 265, 310 −/517.1283/515.1208 517.1283 → 269.0786
1,031.2513 ([2M-H]−) → 515.1230, 267.0677Unknown compounds 23.0 ± 4.5 3 Unknown compound 1 5.2 ± 0.9 270 427.1527/405.1711/403.1626 427.1527 → 265.1018, 203.0510
403.1626 → 223.0989, 179.10888 Unknown compound 2 17.8 ± 4.2 275, 320 643.2288/−/619.2421 643.2288 → 449.1734
619.2421 → 193.0513, 178.0277, 149.0612* Quantitative results are shown as means ± standard deviation of triplicate analyses. '−' means the MSMS spectrum was not provided. Table 2shows the total content of preliminarily identified phenolic compounds (228.4 mg/100 g DM) in the study seeds. The most abundant groups of phenolic compounds were flavanonols (102.2 mg/100 g DM) and flavonols (101.6 mg/100 g DM), the sum of both accounted for 89% of the total content. Taxifolin hexoside (60.0 mg/100 g DM) and quercetin (63.1 mg/100 g DM) presented the dominant roles in the groups of flavanonols and flavonols, respectively. The isoflavone (formononetin coumaroyl hexoside, 1.6 mg/100 g DM) was found at lower content compared to the other phenolic compounds. Our results suggested lower levels of quercetin, kaempferol, isorhamnetin in comparison than the previous data (671.4, 7.9 and 2.9 mg/100 g DM, respectively) of RCS in the research of Chiriac et al.[19]. This variation was probably caused by different extraction procedures. In their study, phenolic compounds were extracted from RCS with assistance of microwave for 15 min. In addition, Prati et al. reported the presence of taxifolin and taxifolin hexoside in seed extract of red clover, though the content of individual compounds were not determined[10]. The anti-cancer properties of flavanonols and flavanols, and their role in reducing the risk of cardiovascular disease have been well documented[8]. They also showed strong antioxidant and antimicrobial effects[8, 14]. Biochanin A, genistein, daidzein and formononetin, along with their respective glucosides were the common isoflavonoids found in red clover leaves[27]. Interestingly, in our study, only one isoflavonoid glycoside (formononetin coumaroyl hexoside) was observed in RCS, and the content was low. Therefore, the distribution and quantity of isoflavonoids varied in different parts of red clover. Considered as potent phytoestrogens, isoflavonoids have been used as the bioactive compounds for treating hormone-related ailments such as post-menopausal symptoms, pre-menstrual syndrome and dysmenorrhea[28]. Additionally, decoctions of red clover flower and leaves have been orally consumed as ethnomedicines to reduce inflammation as well as to treat chest pain and chronic rheumatism[28].
Fatty acids
-
The total lipid content in our study RCS, as determined by a modified Folch method was 7.0 g/100 g DM (Table 3), which was consist with the result (9.8 g/100 g DM) of the research by Ahmed et al.[14]. Nineteen fatty acids were identified in our seeds with a total content of 46,760.5 µg/g DM, in which polyunsaturated fatty acids were 79%. Polyunsaturated linoleic acid took the dominant role, which accounted for 72% of the total fatty acids. Consistent to our findings, linoleic acid presented as the dominant fatty acid in the seeds of Trifolium pratense (44%), Trifolium repens (65%) and Trifolium resupinatum (52%) in previous researches[14, 29]. Comparatively, linoleic acid was observed with low content (5%−11%) in five other Trifolium species (T. balansae, T. stellatum, T. nigrescens subsp. petrisavi, T. constantinopolitanum, and T. resupinatum var. resupinatum)[12]. As essential fatty acids, high content of linoleic acid in our study samples confers the seeds significant nutritional value. For instance, linoleic acid is the precursor of a number of potent pro-inflammatory mediators, including prostaglandins and leukotrienes, which has led to the development of anti-inflammatory pharmaceuticals[30]. The total content of unsaturated fatty acids in the study RCS was 3.9 folds higher than saturated fatty acids. Saturated fatty acids such as palmitic acid (14%) and steric acid (6%) in RCS have the potential of enhancing the oxidative stability in food products. As reported in previous studies, palmitic acid and stearic acid can be used in solid fat applications, such as margarine, shortening and confectionary industries[31].
Table 3. Composition of and content of lipids, fatty acids and tocopherols in RCS*.
Composition Content Lipids (g/100 g DM) 7.0 ± 1.1 Fatty acids (µg/g DM) 46,760.5 ± 2,686.1 Myristic acid (C14:0) 58.3 ± 6.7 Palmitic acid (C16:0) 6,369.6 ± 338.9 Stearic acid (C18:0) 2,993.1 ± 149.8 Arachidic acid (C20:0) 65.7 ± 3.2 Behenic acid (C22:0) 55.7 ± 3.6 Lignoceric acid (C24:0) 48.1 ± 21.6 Total SFA 9,590.5 ± 507.2 Palmitoleic acid (16:1 n-7) 45.8 ± 2.2 Oleic acid (C18:1 n-9) 178.9 ± 4.6 Vaccenic acid (18:1 n-7) 7.6 ± 2.7 Eicosenoic acid (20:1 n-9) 23.3 ± 4.4 Erucic acid (22:1 n-9) 43.0 ± 8.5 Nervonic acid (24:1 n-9) 8.0 ± 4.6 Total MUFA 310.5 ± 10.4 Linoleic acid (18:2 n-6) 33,769.6 ± 1,971.1 α-Linolenic acid (18:3 n-3) 2,863.8 ± 191.1 γ-Linolenic acid (18:3 n-6) 24.2 ± 5.5 Eicosadienoic acid (20:2 n-6) 2.9 ± 0.4 Dihomo-α-linolenic acid (20:3 n-3) 5.7 ± 1.4 Arachidonic acid (20:4 n-6) 5.7 ± 1.5 Eicosapentaenoic acid (20:5 n-3) 187.6 ± 37.0 Total PUFA 36,859.5 ± 2,188.1 Total n-3 3,057.1 ± 219.0 Total n-6 33,802.4 ± 1,969.7 Tocopherols, mg/100 g DM 94.9 ± 4.4 α-Tocopherol 91.9 ± 4.2 β-Tocopherol 1.3 ± 0.1 γ-Tocopherol 1.7 ± 0.1 δ-Tocopherol − * Results are shown as means ± standard deviation of triplicate analyses. '−' means the compound was not detected in the sample. Tocopherols
-
Three tocopherols (α-, β- and γ-tocopherol) were identified in RCS, with a total content of 94.9 mg/100 g DM. Alfa-tocopherol accounted for 97% (91.9 mg/100 g DM), γ-tocopherol for 2% and β-tocopherol for 1%. Our results in Table 3 suggested the same level of tocopherols as that in previous RCS research (101.7 mg/100 g DM)[14]. Alfa-tocopherol is an important antioxidant for humans with positive effects on the immune system. Alfa-tocopherol-rich RCS showed potential in being an antioxidant and anti-inflammatory agent in disease management[32].
Chemical profiles of RCS oil
Oil yields from supercritical CO2 extraction
-
Using SFE, the oil yield from RCS was 3.7% on a dry weight basis, which was significantly lower than the yield (7%) obtained with the Folch method from the same batch of RCS in the laboratory. This difference could have been ascribed to the lower efficiency of CO2 for extracting polar lipids. Increasing the pressure for SFE could potentially result in a higher yield of oil.
Higher oil yields were reported in Soxhlet-extracted underutilized oilseed crop species of the Fabaceae (Leguminosae) family (19%−36%, dry wt. basis), which included Bauhinia purpurea, Phanera vahlii, Butea monosperma, Caesalpinia crista, Gliricidia sepium, Mimosa pudica and Millettia pinnata[33]. Environmental and genetic factors as well as the extract methods can significantly affect the oil yield, fatty acid composition and physiochemical properties of the oil[34].
Fatty acids
-
Extracted by SFE, the oil quality remained almost unchanged in terms of the profiles of fatty acids. As shown in Table 4, linoleic acid was identified as the major component in RCS oil. Linoleic acid was reported as a common fatty acid in the seed oil of many Fabaceae (Leguminosae) family plants[12, 35]. Omega-6 fatty acids are acknowledged as bioactive lipids with health-beneficial properties, in which linoleic acid accounts for 85%−90% of the dietary intake amount[36]. RCS oil contained rich unsaturated fatty acids such as linoleic acid (61%), oleic acid (16%) and α-linolenic acid (6%), with 91% of which are omega-6 fatty acids. Although essential oil were extracted from red and white clover flowers, our study is the first one showing RCS can be a source for producing oil which is rich in n-6 polyunsaturated fatty acids (PUFAs)[37].
Table 4. Composition and content of fatty acids and tocopherols in RCS oil*.
Composition Content (mg/100 g) % in the
fatty acidsFatty acids 89,482.4 ± 2,515.0 Myristic acid (C14:0) 84.7 ± 1.0 0.09 Palmitic acid (C16:0) 9,445.0 ± 394.8 10.55 Stearic acid (C18:0) 4,955.4 ± 207.3 5.54 Arachidic acid (C20:0) 137.2 ± 13.0 0.15 Behenic acid (C22:0) 27.4 ± 4.7 0.03 Lignoceric acid (C24:0) 63.0 ± 7.6 0.07 Total SFA 14,712.7 ± 599.0 16.44 Palmitoleic acid (16:1 n-7) 81.2 ± 3.8 0.09 Oleic acid (C18:1 n-9) 14,036.8 ± 2,598.0 15.69 Eicosenoic acid (20:1 n-9) 59.6 ± 9.2 0.07 Erucic acid (22:1 n-9) 40.4 ± 4.7 0.05 Nervonic acid (24:1 n-9) 5.0 ± 1.7 0.01 Total MUFA 14,223.0 ± 2,607.4 15.89 Linoleic acid (18:2 n-6) 54,665.6 ± 2,159.7 61.09 α-Linolenic acid (18:3 n-3) 5,432.4 ± 81.8 6.07 γ-Linolenic acid (18:3 n-6) 31.2 ± 3.6 0.03 Eicosadienoic acid (20:2 n-6) 74.2 ± 51.4 0.08 Dihomo-α-linolenic acid (20:3 n-3) 5.4 ± 2.2 0.01 Arachidonic acid (20:4 n-6) 5.4 ± 0.8 0.01 Eicosapentaenoic acid (20:5 n-3) 332.3 ± 5.3 0.37 Total PUFA 60,546.6 ± 2,258.0 67.66 Total n-3 5,770.1 ± 88.3 6.45 Total n-6 54,776.5 ± 2,169.7 61.21 Tocopherols (mg/100 g) 40.0 ± 5.9 − α-Tocopherol 38.9 ± 5.8 − β-Tocopherol 0.7 ± 0.1 − γ-Tocopherol 0.5 ± 0.1 − δ-Tocopherol − − * Results are shown as means ± standard deviation of triplicate analyses. '−' means the compound was not detected in the sample. Tocopherols
-
The tocopherols identified in RCS and SFE-extracted RCS oil followed the same pattern. The total content of tocopherols investigated in the present study was 40.0 mg/100 g oil, of which α-tocopherol was predominant (97%), followed by β-tocopherol (2%) and γ-tocopherol (1%). The tocopherol content of seed oils from five Fabaceae species (Vigna angularis, Phaseolus vulgaris, Phaseolus lunatus, Phaseolus coccineus and Glycine soja,) were reported in the range of 1.5−26.7 mg/100 g oil[36]. Gamma-type was the most abundant tocopherol of all five Fabaceae species. Compared to these legume species, our study showed that the RCS oil extracted by SFE contained higher levels of total tocopherols and different profiles of tocopherols, being a richer source of α-tocopherol.
Non-targeted characterization of RCS and SFE-extracted RCS oil
-
In our study, 44 phytochemicals were putatively identified in RCS and RCS oil, which belonged to more than 13 different chemical classes. In terms of the number of individual compounds, lipids were the most abundant class with 16 compounds annotated, followed by phenolic compounds (n = 9), terpenoid (n = 6) and phytosterol (n = 4). As shown in Fig. 1, the metabolites in RCS and the SFE-extracted oil were arranged with hierarchical clustering based on their abundance.
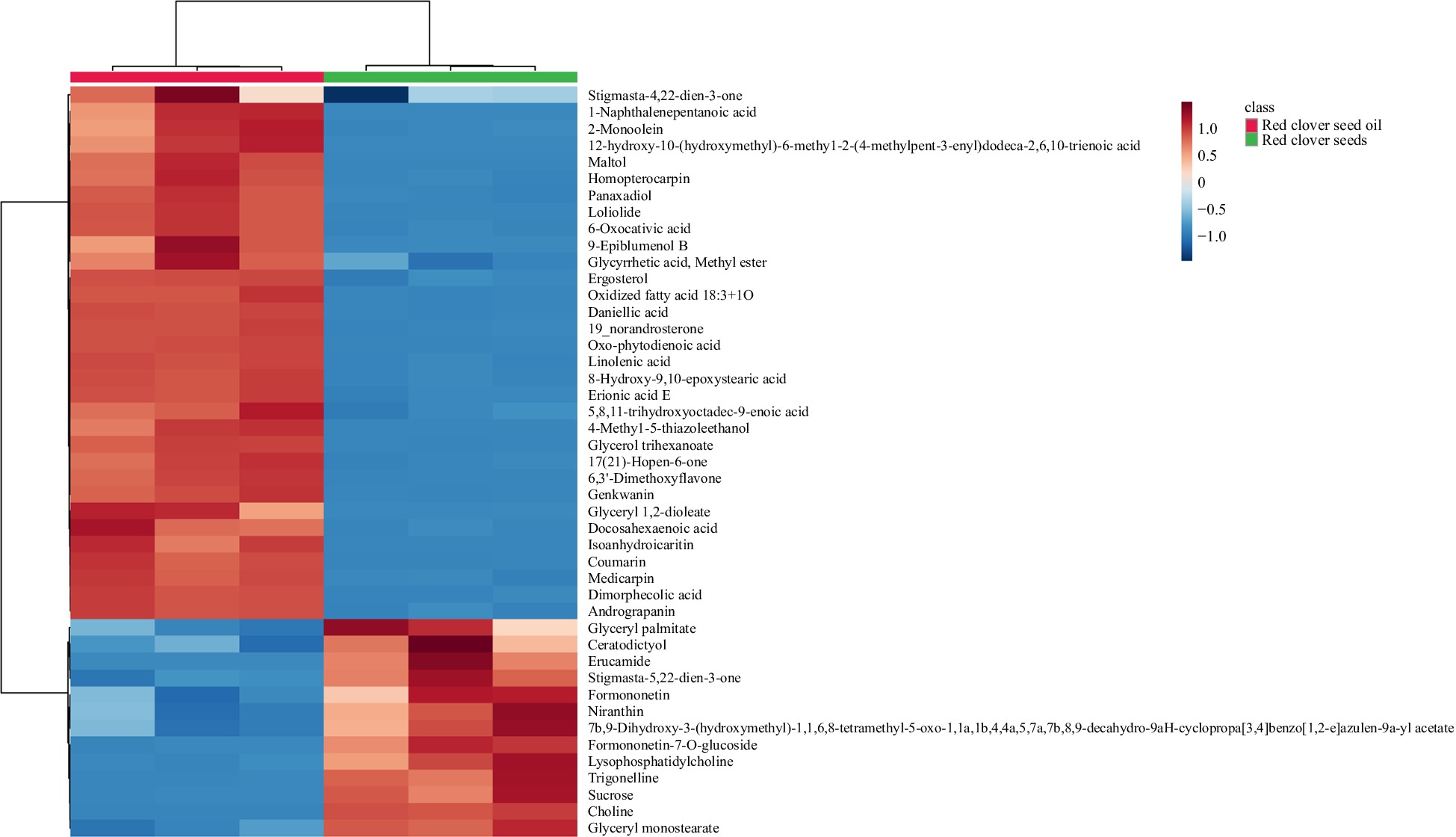
Figure 1.
Heatmap of correlation between phytochemical compounds present in RCS and SFE-extracted oil.
Formononetin, medicarpin and homopterocarpin were identified in the isoflavonoids group in our study. Medicarpin showed significant antiangiogenic and cytotoxic activity, and homopterocarpin has hepatoprotective and antioxidant potential[27, 38, 39]. The presence of these isoflavonoids suggests an interesting potential application of RCS extract and RCS oil in cancer therapy. Unlike the isoflavonoids equally presented in RCS and its oil, other phenolic compounds such as flavonoids (genkwanin, 6,3'-dimethoxyflavone, isoanhydroicaritin), and phenolic acid (erionic acid E) were observed to be more abundant in RCS oil than RCS. Modern pharmacological studies have shown that these compounds were of high importance due to their functions of anti-inflammatory, antioxidant and anti-anxiety[40−42].
Polyunsaturated fatty acids was observed to be more abundant in the SFE-extracted RCS oil than in the RCS in our study. These dietary omega-6 fatty acids have important health benefits since they can reduce blood cholesterol levels[15]. Additionally, monoacylglycerols (glycerol 1-palmitate, 2-monoolein, glyceryl monostearate) were commonly found in vegetable oils. A previous study reported them to be capable of improving loaf volume and texture, although they were not known for being particularly healthy[43]. In our study, phytosterols (19_norandrosterone, stigmasta-4,22-dien-3-one and ergosterol) were dominant in RCS oil, whereas stigmasta-5,22-dien-3-one was found to be more abundant in the RCS. In previous research, phytosterols exerted beneficial hypolipidemic function, as well as anti-cancer, anti-inflammatory, anti-photoaging, anti-osteoarthritic, immunomodulatory, hepatoprotection and antioxidative activities[44].
Terpenoid-metabolites are also compounds with remarkable biological properties such as anti-inflammatory, hepatoprotective, and anti-cancer effects[45]. A wide range of terpenoids were found in RCS and RCS oil including diterpenes (6-Oxocativic acid, daniellic acid, 7b,9-Dihydroxy-3-(hydroxymethyl)-1,1,6,8-tetramethyl-5-oxo-1,1a,1b,4,4a,5,7a,7b,8,9-decahydro-9aH-cyclopropa[3,4]benzo[1,2-e]azulen-9a-yl acetate), triterpene derivatives (panaxadiol, glycyrrhetic acid, methyl ester) and tetraterpene (9-epiblumenol B) indicating their potential for pharmaceutical application.
Furthermore, several compounds with health-promoting properties were determined in our study seeds and oil (Fig. 1). Trigonelline as an alkaloid, was reported to be effective in the hypoglycemic, hypocholesterolemic, antitumor, antimigraine, or antiseptic treatments[46]. Loliolide shows potential in the treatment of patients with obesity as a lipid-lowering agent[47]. Choline is an essential nutrient that is naturally present in some foods and commercially available as a dietary supplement. Choline plays an important role in maintaining the health of the nervous system and in the development of normal brain functions[48]. 4-Methyl-5-thiazoleethanol has pharmacological and biological activities[49]. Previous pharmacological studies showed that andrograpanin exerted anti-inflammatory activity through down-regulating the p38 mitogen-activated protein kinase (MAPKs) signaling pathways[50].
-
To the best of our knowledge, our research is the first study to investigate the phytochemical profiles in RCS and SFE-extracted RCS oil using both targeted and non-targeted analytical methods, which provided a broader image of phytochemical profiles in the study materials. Forty-four phytochemicals were putatively identified in RCS and RCS oil, mainly including lipids, phenolic compounds, terpenoids and phytosterols. Abundant phenolic compounds (including flavanonols, flavonols and isoflavones) were observed in RCS and SFE-extracted oil. Although the oil yield of RCS is not as high as other oilseed crop species in the Fabaceae (Leguminosae) family, the RCS oil has proven to be a rich source of unsaturated fatty acids (mainly linoleic acid and oleic acid) with a high content of α-tocopherol, which has potential for food and pharmaceutical uses.
The study is supported by the Finland-China Food and Health Network (funded by the Finnish Ministry of Education and Culture), FOODNUTRI-Climate Smart Food and Nutrition Research Infrastructure (funded by the Research Council of Finland, No. 337980) and the FIRI 2021 call: Non-roadmap research infrastructures as part of the EU Recovery and Resilience Facility (No. 345916).
-
The authors confirm contribution to the paper as follows: conceptualization: Zhou Y, Tian Y, Selander K, Yang B; methodology: Zhou Y, Tian Y; investigation: Zhou Y, Tian Y, Ollennu-Chuasam P; data curation: Zhou Y, Tian Y; formal analysis and visualization: Zhou Y; writing-original draft: Zhou Y; writing-review & editing: Zhou Y, Tian Y, Kortesniemi M, Selander K, Väänänen K, Yang B; resources: Selander K, Väänänen K; funding acquisition and supervision: Yang B. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files
-
The authors declare that they have no conflict of interest. Baoru Yang is the Editorial Board member of Food Innovation and Advances who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
-
Authors contributed equally: Ying Zhou, Ye Tian
- Supplemental Table S1 External standards applied in quantification of phenolic compounds.
- Supplemental Table S2 ESI-QTOF analytical condition of non-targeted metabolomics.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of China Agricultural University, Zhejiang University and Shenyang Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou Y, Tian Y, Ollennu-Chuasam P, Kortesniemi M, Selander K, et al. 2024. Compositional characteristics of red clover (Trifolium pratense) seeds and supercritical CO2 extracted seed oil as potential sources of bioactive compounds. Food Innovation and Advances 3(1): 11−19 doi: 10.48130/fia-0024-0002 

Compositional characteristics of red clover (Trifolium pratense) seeds and supercritical CO2 extracted seed oil as potential sources of bioactive compounds
- Received: 27 September 2023
- Revised: 20 February 2024
- Accepted: 20 February 2024
- Published online: 28 February 2024
Abstract: Plant seeds from the Fabaceae (Leguminosae) family are commonly edible. However, little has been done to study the phytochemicals of red clover (Trifolium pratense) seeds. Our study aims to obtain comprehensive and novel findings on red clover seeds and supercritical fluid extraction (SFE)-extracted oil, with the purpose of exploring their potential as a new source of functional ingredients for food and health care products. In our study, red clover seed oil was extracted by supercritical CO2. Forty-four phytochemical compounds were preliminarily identified in red clover seeds and the extracted oil by UPLC-ESI-MS/MS metabolomics method. These compounds mainly belong to lipids, phenolic compounds, terpenoids and phytosterols. Red clover seeds contain fatty acids (4,676.1 mg/100 g dried seeds) and bioactive components such as phenolic compounds (228.4 mg/100 g) and tocopherols (94.9 mg/100 g). In red clover seed oil, unsaturated fatty acids are over 83% and are rich in linoleic acid (54.7 g/100 g oil) and oleic acid (14.0 g/100 g oil). These findings provide important guidance for introducing red clover seed oil into pharmaceutical products or as functional foods.