









-
Global warming can directly change the environmental factors required for the growth and development of crops, resulting in changes in their maturity, growth cycle extension, and declines in yield[1−3]. The response of plants to climate change, especially high temperatures, requires more attention and research as plants grow in soil that cannot rid itself of heat. Plants have formed a relatively complex regulatory network to cope with heat stress based on recent research in model plants such as Arabidopsis and tomatoes[4,5]. The heat stress transcription factor–heat shock protein (Hsf–Hsp) pathway is important among the networks, and the Hsf transcription factor is critical in plant responses to heat stress[3,6].
During the heat stress response, Hsf directly regulates the expression of downstream response genes such as transcription factors, chaperone proteins, enzymes, etc.[3,5]. The Hsfs in plants have conserved domains and can be divided into three categories, including HsfA, HsfB, and HsfC, among which HsfA has the largest number and plays major roles in the heat stress response[6]. HsfA2 is only expressed in stress conditions and is induced most strongly by heat stress compared with other HsfAs[7−9]. Overexpression of HsfA2 in Arabidopsis can improve the heat and anoxia tolerance of transgenic lines[8,10]. At the protein level, HsfA2 interacts with HsfA1 in the nucleus to activate heat stress response genes[11,12]. Recently, HsfA2 could activate the H3K27me3 demethylase relative of early flowering 6 (REF6), which represses HsfA2 and drives early flowering transgenerational thermomemory in Arabidopsis[13]. In addition, HsfA3 could interact with HsfA2 to form heteromeric complexes and drive transcriptional memory after heat stress in Arabidopsis[14].
Actin is an important protein that widely exists in eukaryotic cells and forms microfilaments to maintain the stability of the cell's cytoskeleton structure to ensure that a cell can complete the normal physiological metabolism processes[15]. ACTIN is often used as a control gene to compare the different expressions of target genes in various conditions or tissues, as its stable expression makes it a housekeeping gene[16]. ACTIN genes have been cloned, and their function has been studied in many plants, such as Arabidopsis, rice, maize, soybean, etc.[17,18]. At the cellular level, actin can coordinate the establishment of cell polarity, participate in cell wall synthesis and plasma membrane stability, regulate plasmodesmata transport, and direct dynamic cytoplasmic streaming[19,20]. Actin also plays an important role in plant cell morphogenesis, including cell division, cell elongation and expansion, and stomatal movement[21−23]. There are many actin-interacting proteins, such as actin-binding protein (ABP), LIM domain-containing protein (LIM1), and villins (VLNs), that have been found in the research of pollen tube tip growth[24−26]. Although few reports involve actin in stress responses, several research studies showed that actin-interacting proteins could respond to different stresses. As an ABP, actin depolymerizing factors (ADFs) ADF7 and ADF4 were reported to be involved in osmotic and pathogen stress responses in Arabidopsis[27,28]. In addition, members of the ADF family participate in different stress responses such as aluminum stress, osmotic, heat, cold, salinity, and drought in pigeon pea or Zea maize using genome-wide identification and qRT-PCR analysis[29,30]. These ABPs can regulate actin dynamics to respond to different stresses, which are also associated with the reactive oxygen species (ROS) and calcium pathways[31,32]. However, the molecular mechanism of actin in the plant heat stress response is rarely reported.
Lily is an important cut and potted flower worldwide and occupies an important part of the flower market[33,34]. Most lily varieties prefer cool environments based on their native habitat. However, most areas in China are becoming hotter in the summer overtime, which seriously affects the quality and annual production of cut lily flowers[35]. Therefore, improving the heat resistance of lily is a flower industry and scientific problem that needs to be solved urgently. Previously, we focused on the molecular mechanism of heat stress transcription factors regulating the lily heat stress response and cloned several HsfAs from Lilium longiflorum 'White Heaven', such as LlHsfA1, LlHsfA2, LlHsfA3A, LlHsfA3B, and LlHsfA4[12,36−38]. During this research process, we found that Lilium longiflorum 'White Heaven' showed more tolerance than Lilium oriental hybrids 'Siberia' with the same heat stress treatment; however, the regulatory mechanism of heat resistance differences between these two varieties has not been explored[37]. In this study, HsfA2 was cloned from these two cultivars, and it was found that LlHsfA2 has an AIP3 domain from Lilium longiflorum 'White Heaven', but LoHsfA2 did not contain an AIP3 domain from the Lilium oriental hybrids 'Siberia'. Further, LlHsfA2 could interact with LACTIN, whereas LoHsfA2 could not interact. In addition, LlHsfA2 or LACTIN silenced plants were sensitive to heat stress treatment using barley stripe mosaic virus (BSMV) induced gene silencing in 'White Heaven' and silenced LoHsfA2 were sensitive to heat stress in 'Siberia'. We found a new pathway by which HsfA2 regulates the plant heat stress response at the protein level and provided new theoretical support for solving the molecular mechanism of lily heat resistance.
-
Longiflorum hybrid 'White heaven' (Lilium longiflorum) and Oriental hybrid 'Siberia' (Lilium oriental) commercial bulbous plants were cultured in pots containing peat and vermiculite (1:1) in a greenhouse with conditions of 25/20 °C with a 16 h light/8 h dark photoperiod. Nicotiana benthamiana plants were cultured in a greenhouse with the conditions of a 16 h light/8 h dark photoperiod at 22 °C.
Gene cloning and sequence analysis
-
Total RNA was extracted from 'White heaven' and 'Siberia' leaves using the RNAprep Pure Plant Kit (Vazyme, Nanjing, China) according to the manufacturer's instructions. Then, reverse transcription of 1 µg of RNA using HiScript Q RT SuperMix Kit (Vazyme, Nanjing, China). The full−length sequences of LoHsfA2, LlACTIN, and LoACTIN were cloned using reverse transcription production by designing primers (Supplemental Table S1) according to transcriptome sequence. A homologous comparison of amino acid sequences was performed using the DNAMAN (Version 7) software, and the conserved domain prediction was carried out using the DNAMAN (Version 7) software and the SMART website (
http://smart.embl.de/ ).Subcellular localization assay
-
The coding sequence of LACTIN were amplified by PCR using primers and then cloned into a pNC−Cam1304−subC vector to generate LACTIN−GFP, which was used for subcellular localization. LACTIN−GFP and 35S::GFP (as negative control) were transformed into Agrobacterium tumefaciens strain GV3101 and cultured in Luria−Bertani medium containing Kanamycin B selective antibiotics. The cultivated Agrobacterium was harvested using centrifugation at 3,000 g for 10 min and resuspended with the infection buffer (10 mM 2−(N−morpholino)−ethanesulfonic acid, 10 mM MgCl2, 0.2 μM acetosyringone, pH 5.6) to a final concentration at OD600 = 1.0. Then mixed along with the silencing suppressor P19, placed in the dark for about 3 h, and infected into Nicotiana benthamiana plants with 4–5 young leaves. After 72 h, a Zeiss LSM710 META confocal microscope observed the fluorescence signal and green fluorescent protein (GFP) images were acquired at an excitation of 488 nm and emission of 525 nm[39]. The primer sequences used for subcellular localization are listed in Supplemental Table S2.
Bimolecular fluorescence complementation (BiFC) assay
-
The coding sequence of LACTIN, LlHsfA2, and LoHsfA2 were amplified by PCR using primers containing SpeI and KpnI sites and then cloned into pSPYCE (YCE) or pSPYNE (YNE) to generate YCE−LACTIN, YNE−LlHsfA2, and YNE−LoHsfA2. All vectors and their corresponding empty vectors (as negative controls) were transformed into Agrobacterium tumefaciens strain GV3101 and cultured in Luria−Bertani medium containing Kanamycin B selective antibiotics. The cultivated Agrobacterium was harvested using centrifugation at 3,000 g for 10 min and resuspended with the infection buffer (10 mM 2−(N−morpholino)−ethanesulfonic acid, 10 mM MgCl2, 0.2 μM acetosyringone, pH 5.6) to a final concentration at OD600 = 1.0. Different suspension combinations were mixed along with the nuclear marker NF−YA4−mcherry and silencing suppressor P19, placed in the dark for about 3 h, and infected into Nicotiana benthamiana plants with 4–5 young leaves. After 72 h, a Zeiss LSM710 META confocal microscope observed the fluorescence signal and green fluorescent protein (GFP) images were acquired at an excitation of 488 nm and emission of 525 nm, and the mcherry images were acquired at an excitation of 543 nm and emission of 615 nm[39]. The primer sequences used for BiFC are listed in Supplemental Table S2.
Functional verification of HsfA2 and LACTIN in different lily varieties using the BSMV silencing system
-
The BSMV system was used to examine the function of HsfA2 and ACTIN in Longiflorum hybrid 'White heaven' and Oriental hybrid 'Siberia', which contains three vectors: pCaBS−α, pCaBS−β, and pCaBS−γ, and the targeted fragments were ligated to the pCaBS−γ[35,40]. The target fragments with a 262 bp length of the LoHsfA2 gene and 236 bp length of the LACTIN gene were cloned into the pCaBS−γ ligation independent cloning (LIC) vector and transformed into Agrobacterium spp. strain EHA105. Agrobacterium cells containing pCaBS−α, pCaBS−β, pCaBS−γLIC, LoHsfA2−pCaBS−γLIC, or LACTIN−pCaBS−γLIC vectors were cultured in liquid Luria–Bertani medium containing Kanamycin B. The cultivated agrobacterium cells were collected using centrifugation at 3,000 g for 10 min and resuspended in the infecting buffer (10 mM 2−(N−morpholino) ethanesulfonic acid, 10 mM MgCl2, 0.2 μM acetosyringone, pH 5.6) to a final concentration at OD600 = 1.0. Suspensions of pCaBS−α, pCaBS−β, with LoHsfA2−pCaBS−γLIC, LACTIN−pCaBS−γLIC, or pCaBS−γLIC were mixed according to a ratio of 1:1:1 and placed in dark conditions at room temperature for 3 h. The mixed suspension was injected into different leaves of cultivated varieties using syringes without needles; then, the upper leaves were collected to verify whether the gene was silenced after injecting 10 d. All silenced plants were used to examine the heat stress phenotype by treating them at 42 °C for 24 h in a light incubator and then allowing them to recover at 22–23 °C for 1 d. Photographs were taken. The primers used are listed in Supplemental Table S2.
The degree of wilting of the silent strain and the control group was counted according to the criterion of drooping or not drooping of the upper stem leaf bases, and a plant with more than 80% of the upper stem leaf bases drooping was considered to be wilted, and the formula for calculating the percentage of wilting was: Percentage of wilting = (no. of wilted plants/total no. of plants) × 100%.
Gene expression
-
qRT−PCR was used to examine the expressions of HsfA2 and ACTIN in the silenced plants and analyze the expression levels of HsfA2 and ACTIN in ACTIN−silenced or HsfA2−silenced plants. Leaves of different commercial bulbous plants were collected 10 d after injecting, and RNA was extracted and reverse transcripted for real−time quantitative RT-PCR. The 18S rRNA was used as an internal control, which has been validated in our previous studies. The analysis of qRT−PCR results was performed using the 2−ΔΔCᴛ method. Each experiment included three technical replicates, and two biological tests were performed; at least one result is shown in the study. All relevant primers are listed in Supplemental Table S3.
-
HsfA2 plays an important role in the heat−stress response of plants as an important heat stress transcription factor whose expression is induced rapidly and abundantly by heat[7,9,36]. In order to perform functional analysis and comparison of HsfA2 in different lily varieties, Lilium longiflorum 'White Heaven' and Lilium oriental hybrids 'Siberia' cultivars were used according to their different heat response phenotypes. LlHsfA2 has already been cloned from 'White Heaven' in previous studies[36]. The coding sequence of LoHsfA2 was cloned from the cDNA of 'Siberia' by designing primers according to the transcriptome sequence. The full length of LoHsfA2 was 1,077 bp, encoding a 358 amino acid protein.
Amino acid sequence alignment between LlHsfA2 and LoHsfA2 showed that there were only some differing amino acids between the LlHsfA2 and LoHsfA2 protein sequences (Fig. 1a). The overall homology of LlHsfA2 and LoHsfA2 was high, and the conserved domains in Hsfs, such as the DNA binding domain (DBD), oligomeric domain (HR−A/B), nuclear localization signal (NLS), and activation domain (AHA) and nuclear export signal (NES) are close to each other (Fig. 1a). The SMART website was used to analyze the specific domains in LlHsfA2 and LoHsfA2. The results showed that the AIP3 domain existed in LlHsfA2 from 'White Heaven' (Fig. 1b, d), whereas the same domain was not found in LoHsfA2 from 'Siberia' (Fig. 1c). These results indicate that differences in several amino acids might result in domain diversity between homologous genes from different cultivars.
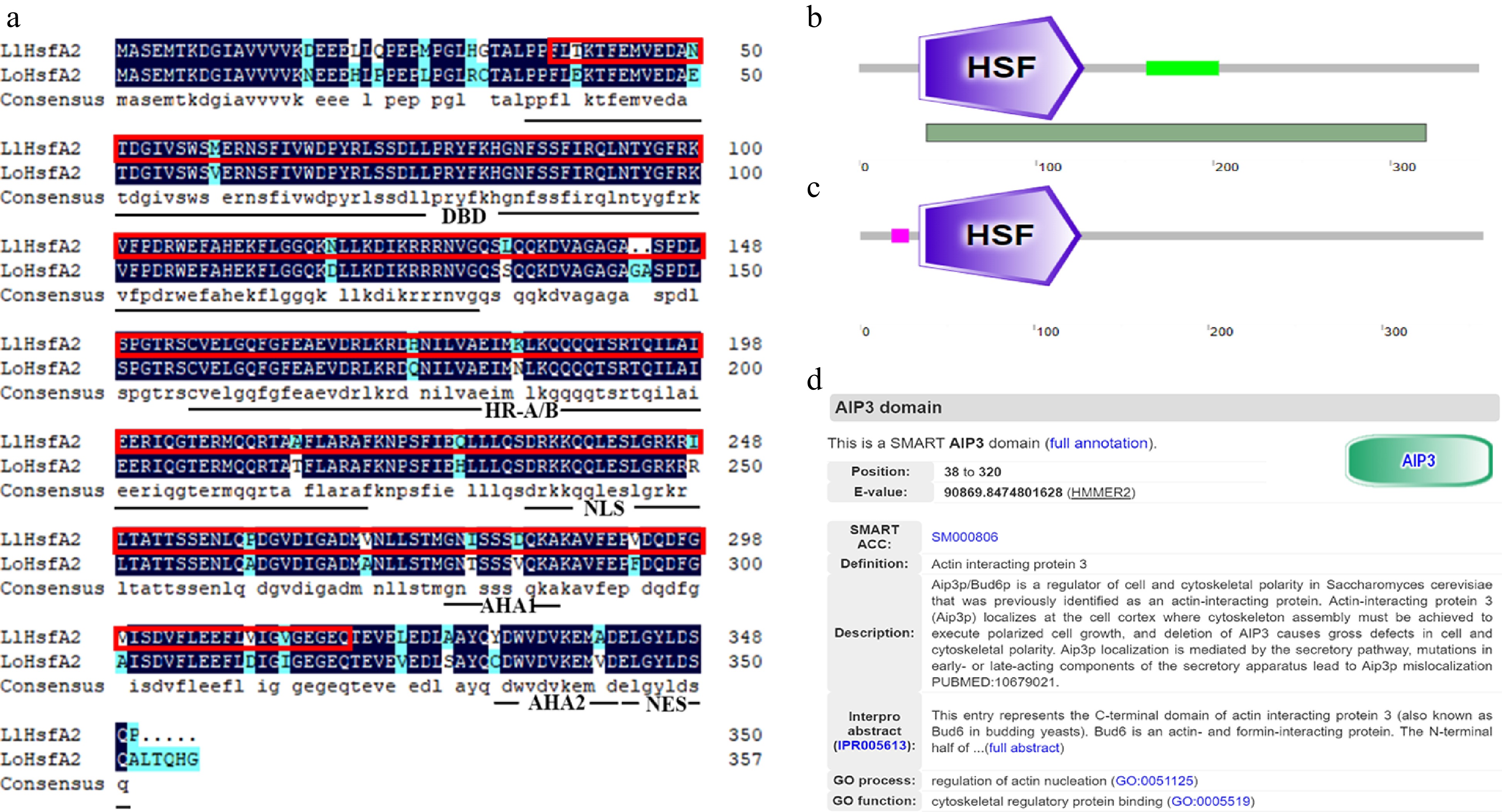
Figure 1.
Amino acid sequence analysis of HsfA2 in 'White Heaven' and 'Siberia'. (a) Amino acid sequence alignment between LlHsfA2 and LoHsfA2. The red boxes indicate the AIP3 domain. DBD, DNA binding domain; HR-A/B: Oligomerization domain; NLS: Nuclear localization sequence; AHA1/AHA2: Activation domains; NES: Nuclear export sequence. (b) The SMART website predicted that LlHsfA2 of 'White Heaven' would contain the HSF and AIP3 domain, and the position of the AIP3 domain was from 38 to 320 amino acids. The dark green bar below represents the location of the AIP3 domain. (c) The SMART website predicted that LoHsfA2 of 'Siberia' only contains the HSF domain. (d) The SMART website comments on the AIP3 domain.
Molecular cloning and sequence analysis of ACTIN in 'White Heaven' and 'Siberia'
-
The AIP3 domain in the protein can bind to ACTIN to affect the interaction between proteins in the relevant studies of Saccharomyces cerevisiae[41]. Therefore, we speculate that LlHsfA2 may interact with LlACTIN to regulate the heat stress response in lily. LlACTIN and LoACTIN were cloned by designing primers according to the transcriptome sequence[35,42]. LlACTIN was cloned from the cDNA of 'White Heaven', which contains a full length of 1,134 bp and encodes a 377 amino acid protein. LoACTIN was cloned from the cDNA of 'Siberia', which contains a full length of 1,134 bp and encodes a 377 amino acid protein. The amino acid sequences of LlACTIN and LoACTIN were homogenously compared. The results showed that the amino acid sequences of LlACTIN and LoACTIN are highly consistent and similar, indicating that the ACTINs are highly conserved in different lily cultivars (Fig. 2a). The phylogenetic tree analysis showed that LACTIN was closest to ACTIN in Tulipa gesneriana, Diospyros lotus, Triadica sebifera, and Vaccinium dunalianum (Fig. 2b).
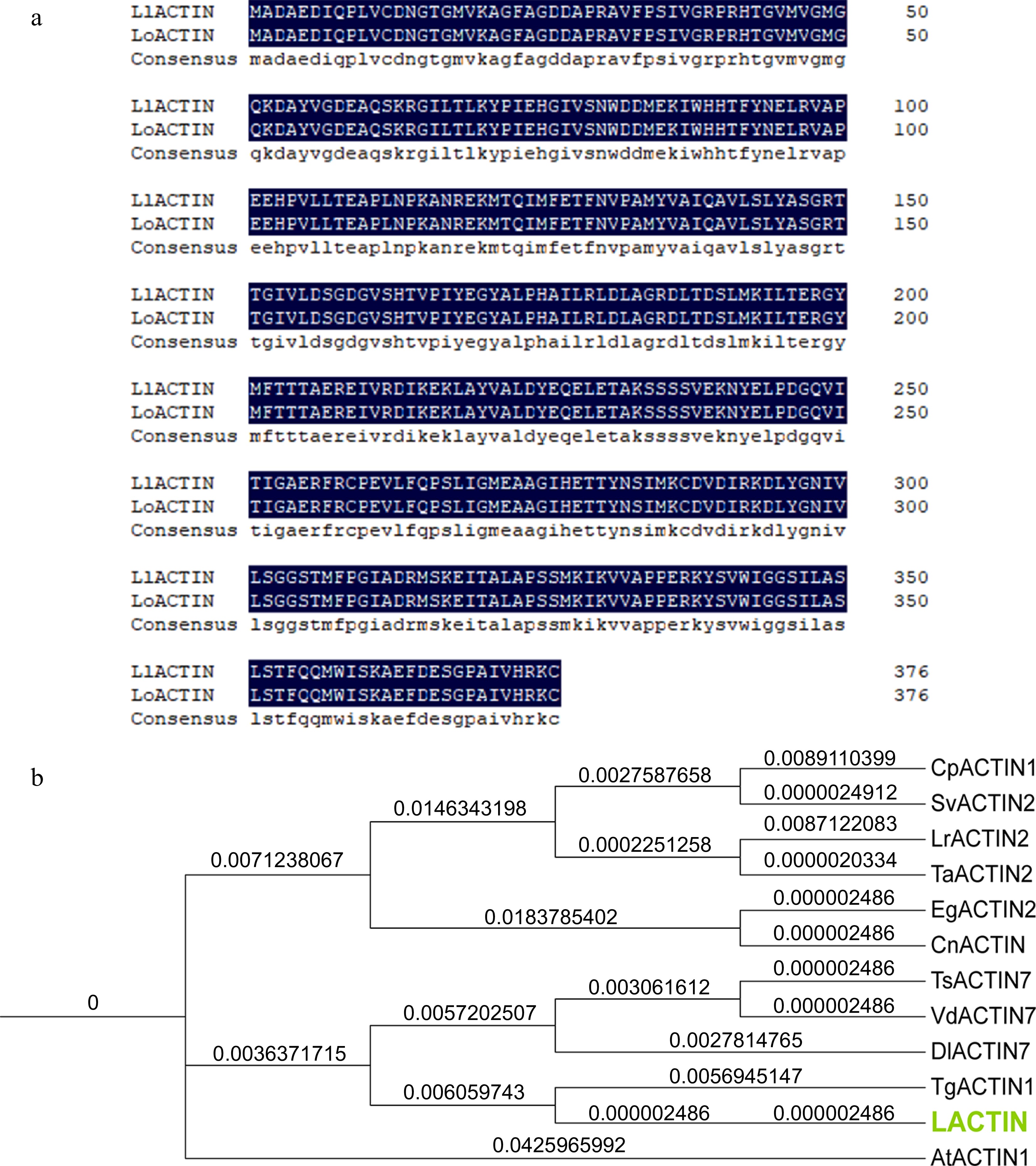
Figure 2.
Amino acid sequence alignment and phylogenetic tree analysis of ACTIN in 'White Heaven' and 'Siberia'. (a) Amino acid sequence alignment between LlACTIN and LoACTIN. (b) Phylogenetic tree of LlACTIN, LoACTIN, and ACTIN in other species. The ACTIN proteins were from: Elaeis guineensis EgACTIN2 (LOC105059495), Cocos nucifera CnACTIN (MH017421.1), Tulipa gesneriana TgActin1 (AB456684.1), Lolium rigidum LrACTIN2(LOC124705928), Triadica sebifera TsACTIN7 (KY656700.1), Diospyros lotus DlACTIN7 (LOC127809754), Cenchrus purpureus CpACTIN1 (MT784734.1), Triticum aestivum TaACTIN2 (LOC123048645), Setaria viridis SvACTIN2 (LOC117839433), and Vaccinium dunalianum VdACTIN7 (OM033722.1). The phylogenetic tree was analyzed by TBtools v1.09876 and drawn by iTOL.
HsfA2 interacts with ACTIN in 'White Heaven' but not in 'Siberia'
-
In a previous study we found that LlHsfA2 was localized in the nucleus[12]. Here, we explored the subcellular localization of LACTIN. LACTIN was inserted into a binary vector containing an eGFP, transformed into Agrobacterium, and injected into N. benthamiana to observe the fluorescent signal. The green fluorescence of LACTIN−GFP was observed in the nucleus and cytoplasm (Fig. 3a). These results indicated that LACTIN was a nucleoplasmic co-localized protein. BiFC was used to verify whether different HsfA2s and ACTINs from 'White Heaven' and 'Siberia' interacted with each other, respectively. HsfA2 and ACTIN of two varieties were inserted into YCE and YNE vectors, transformed into Agrobacterium, and injected into N. benthamiana to observe the fluorescent signal. The results showed that in the combination of LlHsfA2 and LACTIN, the green fluorescence was found in the nucleus and overlapped with the red fluorescence of the nuclear localization marker NF−YA4−mCherry. However, GFP was not observed in the combined LoHsfA2 and LACTIN infection or the control cells (Fig. 3b). These results suggested that LlHsfA2 interacted with LACTIN in the cell nucleus, but LoHsfA2 and LACTIN could not interact with each other.
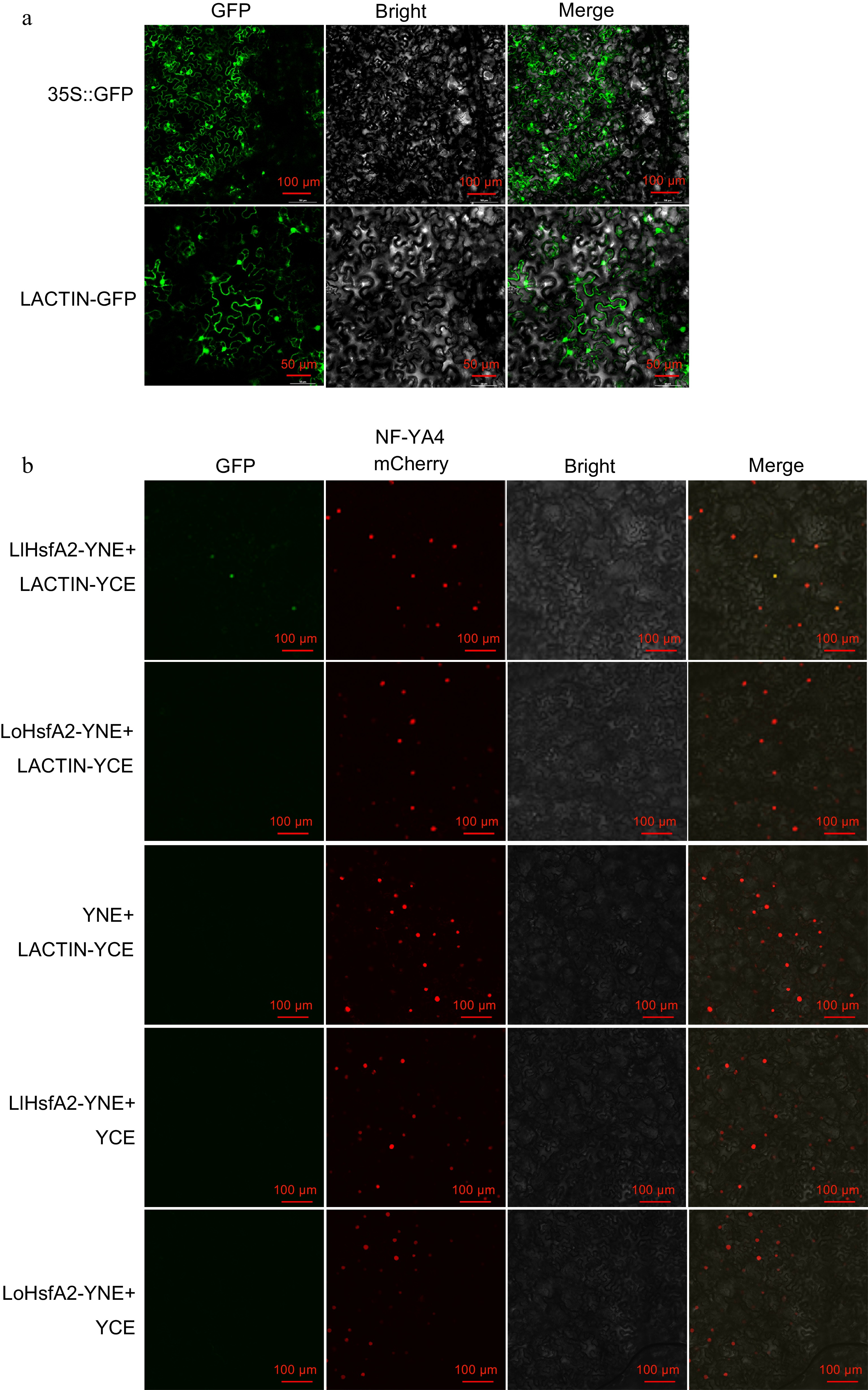
Figure 3.
The interaction relationships of different HsfA2s and ACTINs from 'White Heaven' and 'Siberia'. (a) The subcellular localization of the LACTIN−GFP fusion protein. 35S::GFP was used as the negative control. Green fluorescence were visualized using confocal microscopy 72 h after infiltration. Bars = 100 or 50 μm. (b) BiFC assay. LACTIN−YCE and the empty vector YNE, LlHsfA2-YNE, LoHsfA2-YNE and the empty vector YCE were used as negative controls. NF−YA4−mCherry was used as the nuclear marker. Green and red fluorescence were visualized using confocal microscopy 72 h after infiltration. Bars = 100 μm.
Virus-induced gene silencing of HsfA2 and ACTIN in two varieties
-
In order to verify whether the interaction between LlHsfA2 and LACTIN is involved in the heat stress response in lily, virus-induced gene silencing was used to silence HsfA2 and ACTIN in 'White Heaven' and 'Siberia', respectively, and the heat stress phenotypes of the silenced plants were observed. In a previous study, we silenced LlHsfA2 in 'White Heaven' and found that the silenced strains were sensitive to heat stress treatment[35]. Therefore, LoHsfA2 and LACTIN were silenced in 'White Heaven' or 'Siberia' using BSMV in this study. LoHsfA2 and LACTIN fragments were inserted into BSMV vectors to silence these genes in lily. Using the silencing system, each gene obtained five silenced strains from 20 infected plants and showed the result of two strains. The expressions of LoHsfA2 in 'Siberia' (Fig. 4b), LACTIN in 'White Heaven' (Fig. 4d), and LACTIN in 'Siberia' (Fig. 4f) were decreased compared with their expression in the control plants based on qRT–PCR analysis. All silenced strains were treated at 42 °C for 24 h and allowed to recover for 1 d at room temperature. Compared with the control plants, there was obvious wilting in the leaves of LoHsfA2 silenced 'Siberia' and LACTIN silenced 'White Heaven' (Fig. 4a, c & g), whereas the LACTIN silenced 'Siberia' did not have a different phenotype (Fig. 4e, g). These results indicated that silencing of HsfA2 decreased the thermal tolerance of lilies, and LACTIN silencing decreased the thermal tolerance of 'White Heaven', whereas silencing of LACTIN did not affect the thermal tolerance of 'Siberia'.
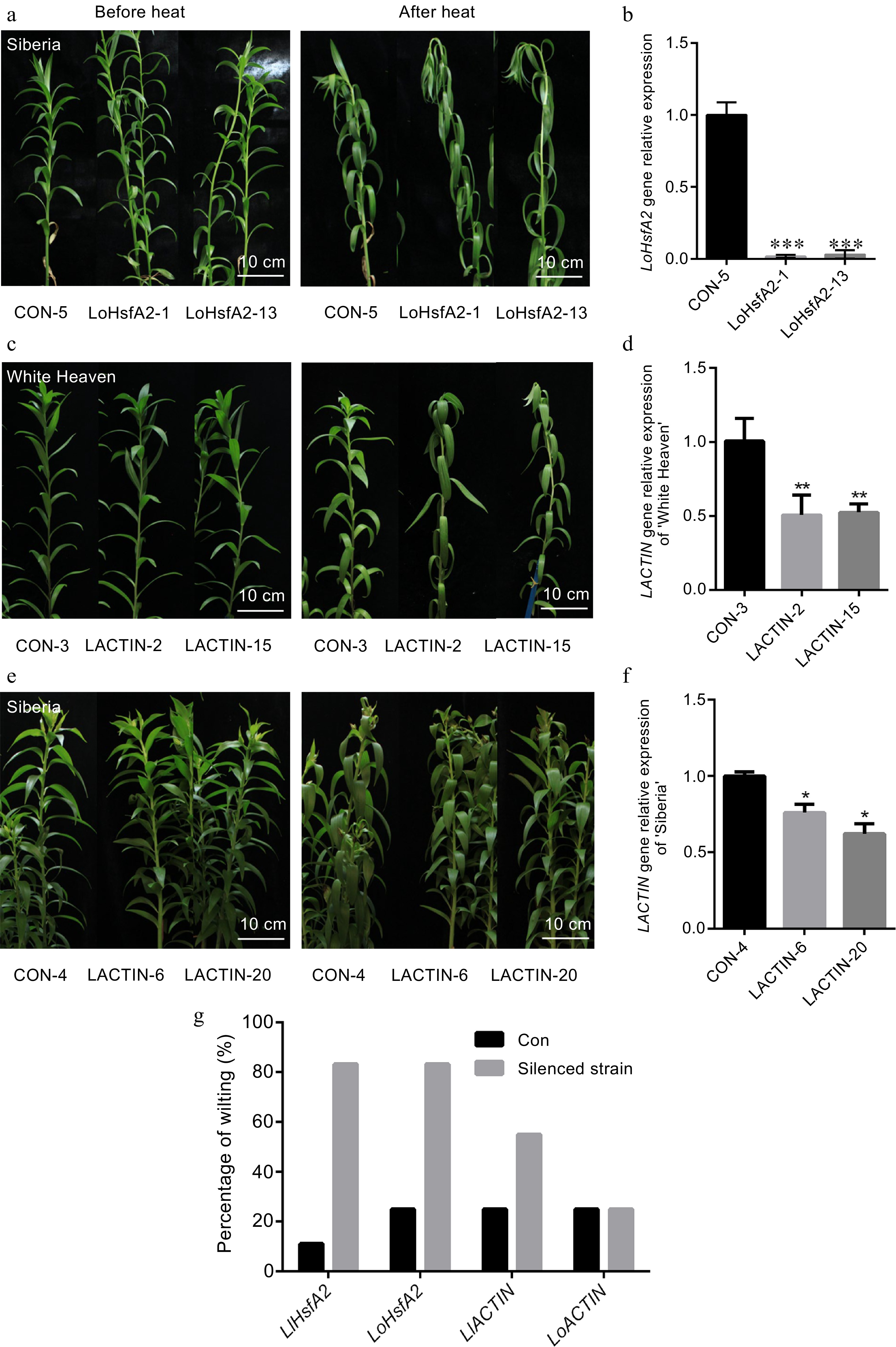
Figure 4.
Heat stress phenotype and gene expression of silenced HsfA2 and ACTIN in 'White Heaven' and 'Siberia'. (a), (c), (e) The phenotypes of the LoHsfA2, LACTIN silenced 'White Heaven' or 'Siberia' lines before heat and recovered at room temperature for 1 d after treatment with heat stress at 42 °C for 24 h. Bar: 10 cm. (b), (d), (f) Expression of LoHsfA2, LlACTIN silenced lines based on qRT-PCR. A T-test analysis of variance was employed to identify treatment means that differed statistically. Samples with different stars are significantly different: * p <0.05, ** p <0.01, and *** p <0.001. (g) Percentage wilting of the LlHsfA2, LoHsfA2, LACTIN silenced 'White Heaven' or 'Siberia' lines and negative control lines recovered at room temperature for 1 d after treatment with heat stress at 42 °C for 24 h.
Silencing of ACTIN or HsfA2 affected the expression of each other in 'White Heaven'
-
In order to examine whether HsfA2 and ACTIN affect each other's expression at the gene level, qRT–PCR was used to detect the expression levels of ACTIN and HsfA2 in the HsfA2 or ACTIN–silenced lines of 'White Heaven' and 'Siberia'. The results showed that the expression of HsfA2 and ACTIN showed a significant change in the ACTIN and HsfA2 silenced 'White Heaven' lines (Fig. 5a, b) but not in the 'Siberia' lines (Fig. 5c, d). Therefore, HsfA2 and ACTIN also affected each other's expression in 'White Heaven' at the gene level. This regulatory relationship may form a positive feedback regulation to mediate heat tolerance and play an important role in the heat stress response of 'White Heaven'.
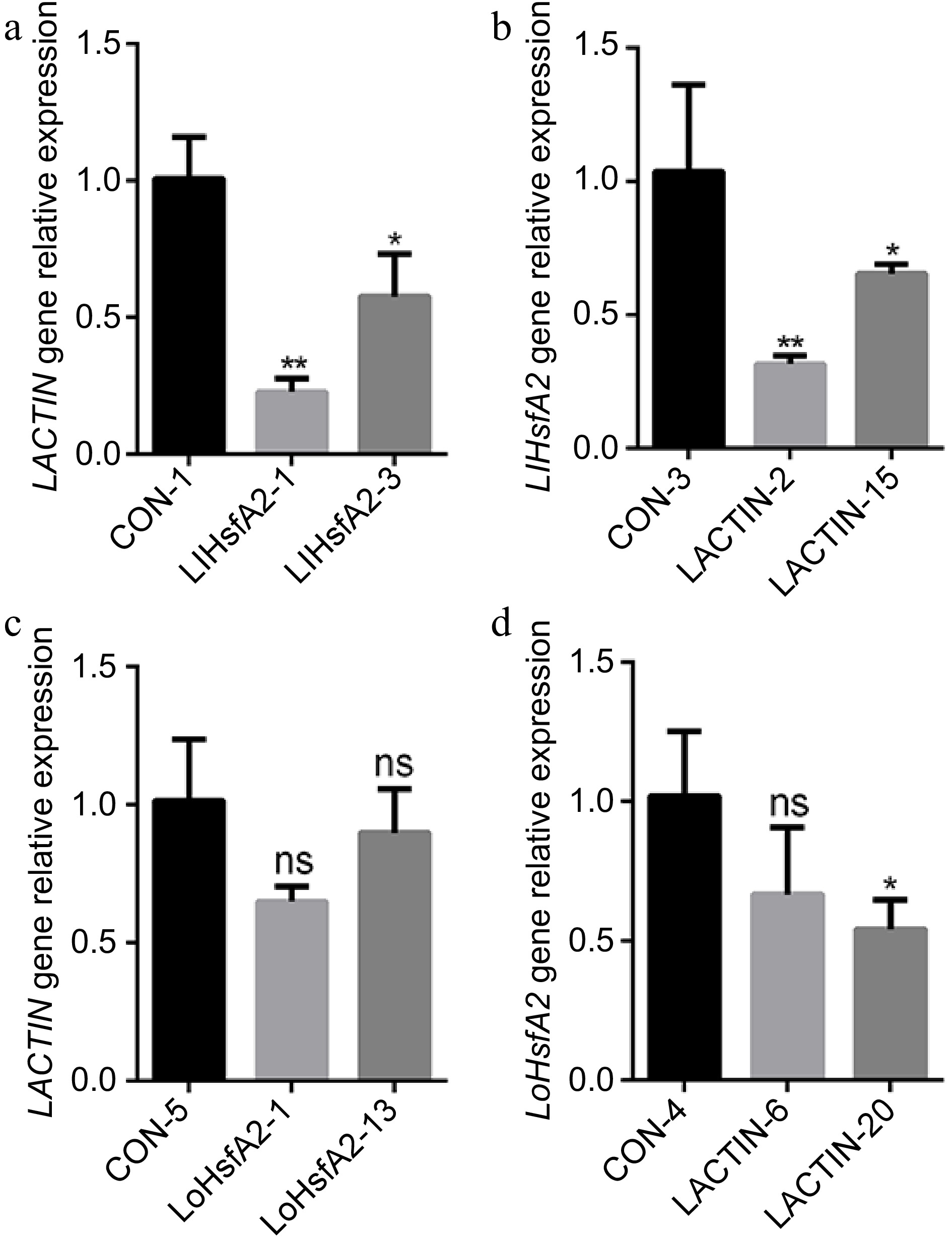
Figure 5.
The expressions of ACTIN and HsfA2 in the HsfA2 or ACTIN-silenced lines of 'White Heaven' and 'Siberia'. (a) The expression of LACTIN in the LlHsfA2 silenced 'White Heaven' line. (b) The expression of LlHsfA2 in the LACTIN silenced 'White Heaven' line. (c) The expression of LACTIN in the LoHsfA2 silenced 'Siberia' line. (d) The expression of LoHsfA2 in the LACTIN silenced 'Siberia' line. A T-test analysis of variance was employed to identify treatment means that differed statistically. Samples with different stars are significantly different: * p <0.05, ** p <0.01; ns: non-significance.
-
When plants are subjected to heat stress in the environment, many transcription factors are induced to regulate the heat stress response and induce the expression of corresponding stress proteins, thus enhancing the heat tolerance of plants in all aspects[6]. Heat stress transcription factors play an important role in all levels of the heat stress response network and complete strict regulatory processes through the Hsf–Hsp pathway[5,43,44]. In this study, we found that LlHsfA2 from 'White Heaven' could interact with ACTIN, which contains the AIP3 domain (Figs 1 & 3b), indicating that HsfA2 regulates the heat stress response through other pathways more than the Hsf–Hsp pathway.
The protein HsfA2 and ACTIN are conserved in different lily varieties, 'White Heaven' and 'Siberia'
-
HsfA2 plays an important role in plant response to heat stress; in the early stage of the heat stress response, HsfA1 regulates the expression of HsfA2, then HsfA2 interacts with HsfA1 at the protein level to regulate other heat stress responses[7,8,36]. In a long-term heat stress environment, HsfA2 was induced highly and accumulated continually, which is an enhancer of acquired thermal tolerance[9]. In this study, we cloned LoHsfA2 from Lilium oriental 'Siberia' and found it was highly homologous with the LlHsfA2 from Lilium longiflorum 'White Heaven'[36] (Fig. 1a), indicating that HsfA2 was conserved in these two varieties. There were only a few amino acid sequences that differed, which caused the presence or absence of the AIP3 domain (Fig. 1). This means that LlHsfA2 contains the AIP3 domain, and LoHsfA2 does not. LlHsfA2 could interact with ACTIN through the AIP3 domain, whereas LoHsfA2 could not function because it lacked this domain (Fig. 3), which may cause differences in heat resistance. This result indicated that although the protein sequence was highly conserved in these two varieties, differences in several amino acids could also lead to differences in the protein domain.
As an important component of the cytoskeleton, actin is required for cell differences, cell growth, and signal transduction, and ACTIN is constantly expressed in various tissues and has a high degree of conservation and homology at the RNA and amino acid levels[27,32,45]. The dynamic conversion of the actin cytoskeleton is essential for a variety of plant physiological processes, which are regulated by ABPs and ADFs[32,46]. Several ADFs, such as ADF4 and ADF7, were reported to regulate the biotic or abiotic stress response through actin dynamics and ROS pathways[28]. In this study, we cloned the ACTIN from 'White Heaven' and 'Siberia', respectively, and found they encode the same amino acid sequence named LACTIN (Fig. 2a), indicating that the evolution of ACTIN in these two varieties was highly conserved, which was consistent with previous research results. In addition, the phylogenetic tree analysis showed that lily's LACTIN was closest to the actin in monocotyledonous plants (Fig. 2b), which also conforms to the conservation in the evolution of actin.
The different interactions between LlHsfA2 and LACTIN lead to diverse heat stress responses in distinct varieties
-
Actin-interacting protein (AIP) was first found to interact with actin in yeast[41]. In this study, we found that LlHsfA2 of Lilium longiflorum 'White Heaven' contains the AIP3 domain and could interact with actin using the BiFC assay. Meanwhile, the LoHsfA2 of Lilium oriental 'Siberia' lacks the AIP3 domain and cannot interact with actin (Figs 1 & 3). Silencing LlHsfA2 or ACTIN induced heat sensitivity in Lilium longiflorum, which means that LlHsfA2 and ACTIN can regulate the heat stress response by interacting with each other (Fig. 4c & g). While in Lilium oriental 'Siberia' that was more sensitive than 'White Heaven' in heat stress, silenced LoHsfA2 plants showed sensitivity to heat stress (Fig. 4a & g), but silenced ACTIN plants have no obvious phenotype compared with the control plants (Fig. 4e). These results indicated that the different interactions between LlHsfA2 and ACTIN affect the diversity of the heat stress response in cultivated varieties. Similar to the ABPs or ADFs, LlHsfA2 could affect actin filament dynamics or Ca2+ or ROS signals to regulate the heat stress response by interacting with actin[27,28]. This regulation mechanism needs further research on whether the HsfA2 could affect actin dynamics.
During the cytoskeleton accumulation and actin aggregation induced by heat stress, small HSPs bind to denatured actin monomers or short oligomers and protect them from aggregation by forming relatively small and highly soluble complexes, which protect the cytoskeleton and the whole cell from damage caused by the accumulation of large insoluble aggregates[47,48]. While studying the effect of heat stress on the actin cytoskeleton of tobacco using two cultured cells, it was found that heat stress induced the depolymerization of actin microfilaments accompanied by the accumulation of Hsp70 binding protein (BiP)[49]. In studies on plants such as tomatoes, it was found that HsfA2 can regulate the transcription of Hsps and interact with Hsps to regulate their activity upon exposure to heat stress, which protects the proteins from being misfolded and losing their functional orientation during heat stress[50−52]. In our study, it was found that in 'White Heaven', LACTIN and LlHsfA2 regulated each other's transcription (Fig. 5a, b). However, no such regulation occurred in 'Siberia' (Fig. 5c, d). Therefore, we speculate that LACTIN may regulate the transcription of LlHsfA2 or interact with LlHsfA2 to affect its protein activity under heat stress, and then LlHsfA2 regulates the transcription of Hsps or interacts with Hsps to regulate the activity of Hsps to prevent LACTIN aggregation and cell damage, forming a positive feedback regulation mode of Hsf–Hsp–ACTIN. In addition, actin also plays an important role in guarding cell turgor regulation and shape changes[20,53,54], influencing the process of programmed cell death[55], altering the cytosolic calcium level[56,57], altering ROS signaling[58], changing organelle morphology[59], and controlling stomatal movement[60]. The interaction of LlHsfA2 and LACTIN may also activate LACTIN and protect cells from heat stress damage in these aspects. However, the specific molecular mechanisms of these assumptions need to be further explored.
-
LlHsfA2 and LACTIN can interact at the protein level and mediate the gene expression of each other in Lilium longiflorum 'White Heaven', which protects cells in 'White Heaven' from heat stress damage. In contrast, this interaction and regulatory relationship was not found in 'Siberia', which may be a reason for the difference in heat resistance between these two varieties. This conclusion provides a new area for further research on the response of lily to heat stress and the cultivation of heat-resistant lily varieties.
-
The authors confirm contribution to the paper as follows: study conception and design: He J, Yi M; data collection: Wang Y, Song C, Tong S, Guo Y, Yang X, Li C, Shao Y; analysis and interpretation of results: Wang Y, Song C, Tong S; helpful discussion provided: Yi M; manuscript review and editing: He J. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This research was funded by the National Natural Science Foundation of China (31972438 and 31300219) and the Chinese Universities Scientific Fund (2018QC096 and 2017YY003).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yue Wang, Cunxu Song, Shi Tong
- Supplemental Table S1 Primers used for LoHsfA2, LlACTIN, and LoACTIN ORF isolation in lily.
- Supplemental Table S2 Primers used for vector construction.
- Supplemental Table S3 Primers used for RT-PCR.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Y, Song C, Tong S, Guo Y, Yang X, et al. 2024. The diversity in interaction between HsfA2 and ACTIN leads to differences in heat stress responses among different lily varieties. Ornamental Plant Research 4: e011 doi: 10.48130/opr-0024-0009 

The diversity in interaction between HsfA2 and ACTIN leads to differences in heat stress responses among different lily varieties
- Received: 25 November 2023
- Revised: 02 February 2024
- Accepted: 26 February 2024
- Published online: 24 April 2024
Abstract: Lily is an important cut flower worldwide and prefers cool environments. The high summer temperature in China affects the quality and quantity of cut lily–flower production. In this study, the heat stress transcription factor HsfA2 was cloned from two cultivated commercial varieties, Lilium longiflorum 'White Heaven' and Lilium oriental 'Siberia', which have different responses to heat stress. An actin-interacting protein 3 (AIP3) domain was found in LlHsfA2 of Lilium longiflorum 'White Heaven' but did not appear in LoHsfA2 of Lilium oriental 'Siberia' using prediction from the simple modular architecture research tool (SMART) website. The genes LlACTIN and LoACTIN were cloned, and their amino acid sequences were found to be the same, so they were named LACTIN. There was an interaction between LlHsfA2 and LACTIN, whereas this interaction did not occur between LoHsfA2 and LACTIN based upon the bimolecular fluorescence complementary (BiFC) experiments. The silenced plants of LlHsfA2 and LACTIN were sensitive to heat stress treatment using barley stripe mosaic virus (BSMV) induced gene silencing in 'White Heaven', and LoHsfA2 silenced plants were also sensitive in 'Siberia'. In contrast, LACTIN silenced plants in 'Siberia' were normal. The expressions of LlHsfA2 or LACTIN were reduced in the LACTIN or LlHsfA2 silenced 'White heaven' plants, whereas the expressions of LoHsfA2 or LACTIN were normal in the LACTIN or LoHsfA2 silenced 'Siberia' plants. In conclusion, the diversity of protein interactions between HsfA2 and ACTIN may affect the distinctness of the heat stress response in different lily varieties, which provides new knowledge for further study on the heat stress response in lily.
-
Key words:
- Lily /
- Heat shock transcription factor /
- Actin /
- Heat tolerance